
Homeostasis as the Mechanism of Evolution

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Homeostasis is Anything But Static
3. The Historic Concept of Homeostasis, from Bernard to Cannon
4. Negative Feedback
5. Homeostatic Imbalance
6. The Cart and Horse Diachronic Perspective
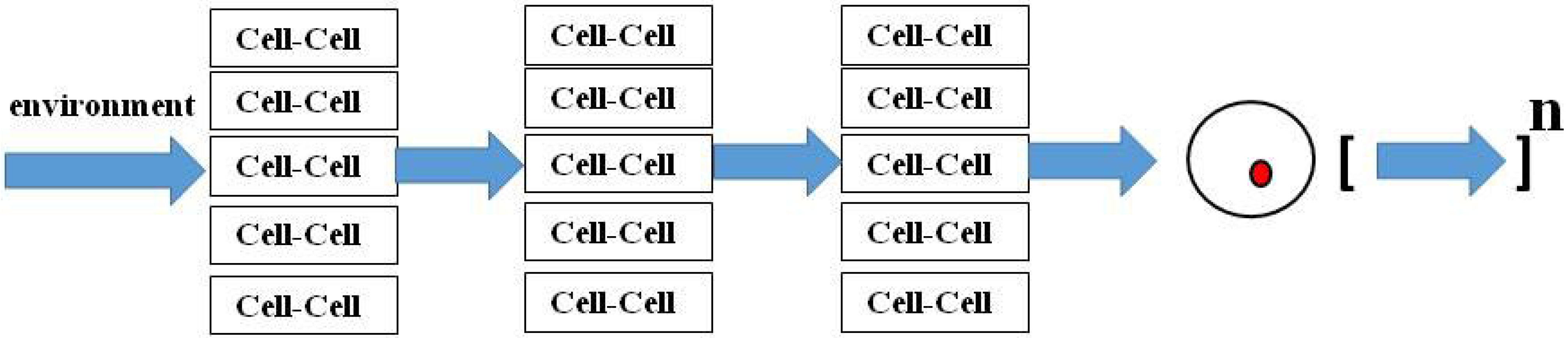
7. Homeostatic Regulation as Downward Causation

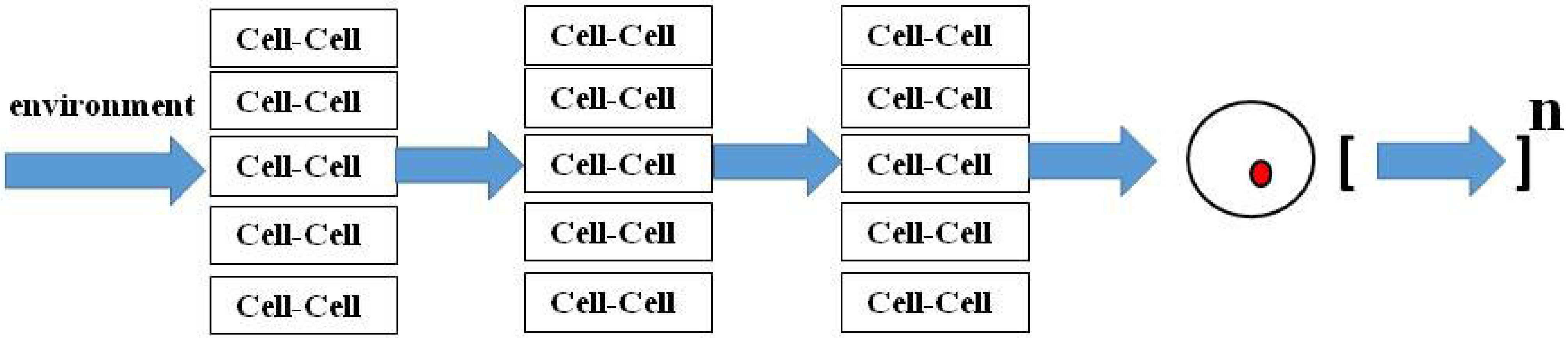
8. Diachronic Growth Factor Signaling Mechanisms are Common to Development, Homeostasis and Regeneration
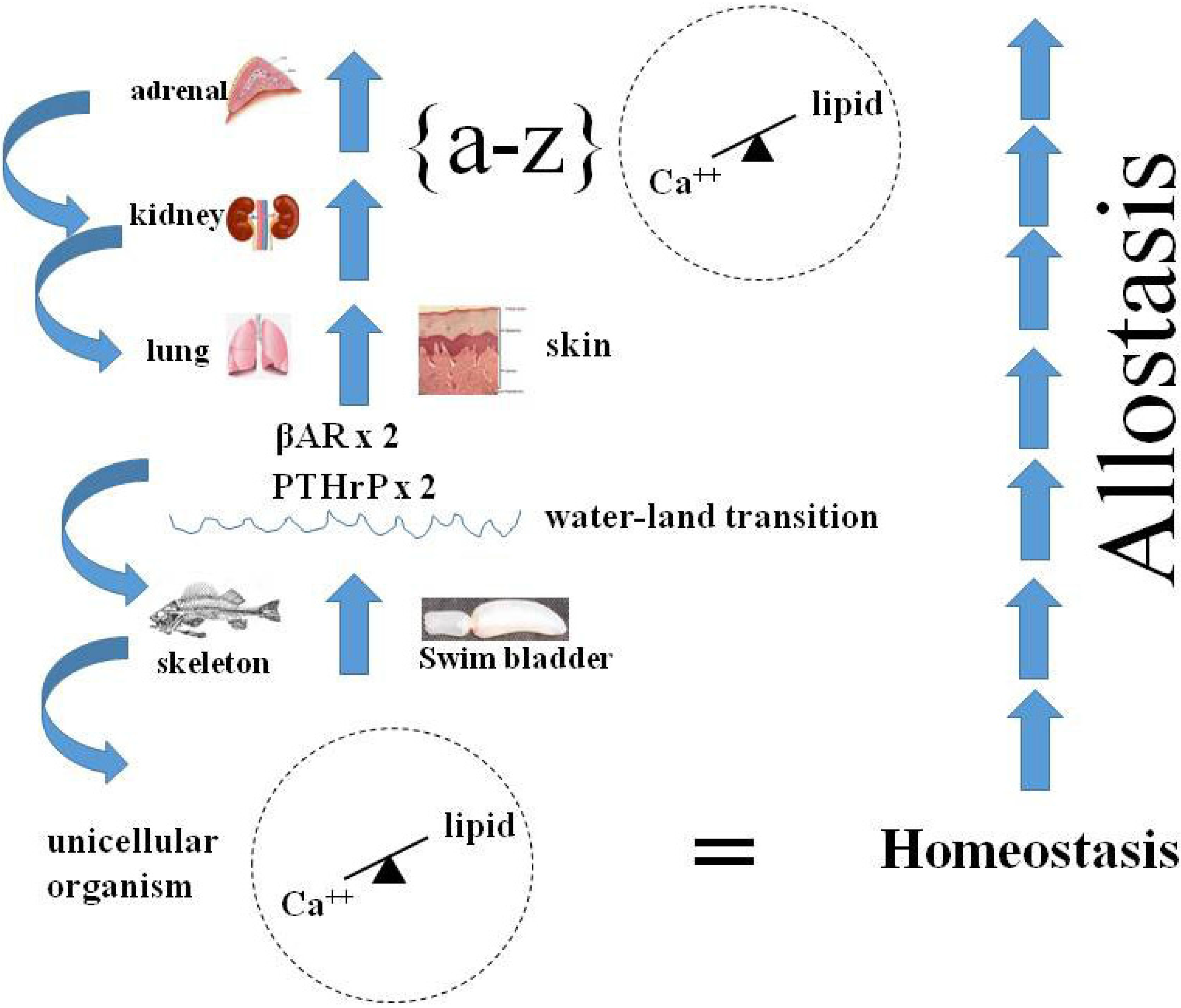
9. Homeostasis as the Agent for Change During the Vertebrate Water-Land Transition-Emergence
10. Homeostasis as the Consequence of Developmental Mechanisms


11. PTHrP and Hypothalamic-Pituitary-Adrenal Regulation of Physiologic Stress
12. The Role of PTHrP Expression in Adrenal Corticoid Synthesis
13. Homeostatic Regulation is Diachronic
14. Allostasis as Integrated Homeostasis
15. Conclusions
Acknowledgements
Conflicts of Interest
References
- Birks, J.B. Rutherford at Manchester; Heywood & Co.: London, UK, 1962. [Google Scholar]
- Torday, J.S.; Rehan, V.K. Evolutionary Biology, Cell-Cell Signaling and Complex Disease; Wiley: Hoboken, NJ, USA, 2012. [Google Scholar]
- Torday, J.S. Evolutionary biology redux. Perspect. Biol. Med. 2013, 56, 455–484. [Google Scholar] [CrossRef] [PubMed]
- Torday, J.S. A central theory of biology. Med. Hypotheses 2015, 85, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Jacob, F. Evolution and tinkering. Science 1977, 196, 1161–1166. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, P.L.; Cardoso, J.C.; Power, D.M.; Canário, A.V. Functional characterization and evolution of PTH/PTHrP receptors: Insights from the chicken. BMC Evol. Biol. 2012. [Google Scholar] [CrossRef] [PubMed]
- Aris-Brosou, S.; Chen, X.; Perry, S.F.; Moon, T.W. Timing of the functional diversification of alpha- and beta-adrenoceptors in fish and other vertebrates. Ann. N. Y. Acad. Sci. 2009, 1163, 343–347. [Google Scholar] [CrossRef] [PubMed]
- Bridgham, J.T.; Carroll, S.M.; Thornton, J.W. Evolution of hormone-receptor complexity by molecular exploitation. Science 2006, 312, 97–101. [Google Scholar] [CrossRef] [PubMed]
- Lane, N.; Allen, J.F.; Martin, W. How did LUCA make a living? Chemiosmosis in the origin of life. Bioessays 2010, 32, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Lane, N.; Martin, W.F. The origin of membrane bioenergetics. Cell 2012, 151, 1406–1416. [Google Scholar] [CrossRef] [PubMed]
- Martin, W.F.; Sousa, F.L.; Lane, N. Energy at life’s origin. Science 2014, 344, 1092–1093. [Google Scholar] [CrossRef] [PubMed]
- Bhagavan, N.V. Medical Biochemistry; Academic Press: Waltham, MA, USA, 2002. [Google Scholar]
- Reydon, T.A. Generalizations and kinds in natural science: The case of species. Stud. Hist. Philos. Biol. Biomed. Sci. 2006, 37, 230–255. [Google Scholar] [CrossRef] [PubMed]
- Pennazio, S. Homeostasis: A history of biology. Riv. Biol. 2009, 102, 253–271. [Google Scholar] [PubMed]
- Roux, E. The concept of function in modern physiology. J. Physiol. 2014, 592, 2245–2249. [Google Scholar] [CrossRef] [PubMed]
- MacDougall-Shackleton, S.A. The levels of analysis revisited. Philos. Trans. R. Soc. Lond. B 2011, 366, 2076–2085. [Google Scholar] [CrossRef] [PubMed]
- Lwebuga-Mukasa, J.S.; Ingbar, D.H.; Madri, J.A. Repopulation of a human alveolar matrix by adult rat type II pneumocytes in vitro. A novel system for type II pneumocyte culture. Exp. Cell Res. 1986, 162, 423–435. [Google Scholar] [CrossRef]
- Torday, J.S.; Rehan, V.K. Up-regulation of fetal rat lung parathyroid hormone-related protein gene regulatory network down-regulates the Sonic Hedgehog/Wnt/betacatenin gene regulatory network. Pediatr. Res. 2006, 60, 382–388. [Google Scholar] [CrossRef] [PubMed]
- Gleason, C.A.; Devaskar, S. Avery’s Diseases of the Newborn; Saunders: Philadelphia, PA, USA, 2011. [Google Scholar]
- Torday, J.S.; Rehan, V.K.; Hicks, J.W.; Wang, T.; Maina, J.; Weibel, E.R.; Hsia, C.C.; Sommer, R.J.; Perry, S.F. Deconvoluting lung evolution: From phenotypes to gene regulatory networks. Integr. Comp. Biol. 2007, 47, 601–609. [Google Scholar] [CrossRef] [PubMed]
- Sahni, R.; Polin, R.A. Physiologic underpinnings for clinical problems in moderately preterm and late preterm infants. Clin. Perinatol. 2013, 40, 645–663. [Google Scholar] [CrossRef] [PubMed]
- Chilosi, M.; Poletti, V.; Zamò, A.; Lestani, M.; Montagna, L.; Piccoli, P.; Pedron, S.; Bertaso, M.; Scarpa, A.; Murer, B.; et al. Aberrant Wnt/β-catenin pathway activation in idiopathic pulmonary fibrosis. Am. J. Pathol. 2003, 162, 1495–1502. [Google Scholar] [CrossRef]
- Marieb, E.N.; Hoehn, K. Human Anatomy & Physiology; Pearson Benjamin Cummings: San Francisco, CA, USA, 2007. [Google Scholar]
- Besnard, V.; Wert, S.E.; Stahlman, M.T.; Postle, A.D.; Xu, Y.; Ikegami, M.; Whitsett, J.A. Deletion of Scap in alveolar type II cells influences lung lipid homeostasis and identifies a compensatory role for pulmonary lipofibroblasts. J. Biol. Chem. 2009, 284, 4018–4030. [Google Scholar] [CrossRef] [PubMed]
- Torday, J.S.; Powell, F.L.; Farmer, C.G.; Orgeig, S.; Nielsen, H.C.; Hall, A.J. Leptin integrates vertebrate evolution: From oxygen to the blood-gas barrier. Respir. Physiol. Neurobiol. 2010, 173, S37–S42. [Google Scholar] [CrossRef] [PubMed]
- Torday, J.S.; Rehan, V.K. Deconvoluting lung evolution using functional/comparative genomics. Am. J. Respir. Cell Mol. Biol. 2004, 31, 8–12. [Google Scholar] [CrossRef] [PubMed]
- Reid, R.G.B. Evolutionary Biology: The Unfinished Synthesis; Cornell University Press: Ithaca, NY, USA, 1985. [Google Scholar]
- Bernard, C. Introduction à L’étude de la Médecine Expérimentale; Flammarion: Paris, France, 1865. [Google Scholar]
- Cannon, W.B. Organization for physiological homeostasis. Physiol. Rev. 1929, 9, 399–431. [Google Scholar]
- Waddington, C.H. The Strategy of the Genes; George Allen & Unwin: London, UK, 1957. [Google Scholar]
- McEwen, B.S.; Wingfield, J.C. The concept of allostasis in biology and biomedicine. Horm. Behav. 2003, 43, 2–15. [Google Scholar] [CrossRef]
- McEwen, B.S.; Wingfield, J.C. What is in a name? Integrating homeostasis, allostasis and stress. Horm. Behav. 2010, 57, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Rué, P.; Arias, A.M. Cell dynamics and gene expression control in tissue homeostasis and development. Mol. Syst. Biol. 2015. [Google Scholar] [CrossRef] [PubMed]
- Kotas, M.E.; Medzhitov, R. Homeostasis, inflammation, and disease susceptibility. Cell 2015, 160, 816–827. [Google Scholar] [CrossRef] [PubMed]
- Mayer, E.A. Gut feelings: The emerging biology of gut-brain communication. Nat. Rev. Neurosci. 2011, 12, 453–466. [Google Scholar] [CrossRef] [PubMed]
- Kirschner, M.; Gerhart, J.; Mitchison, T. Molecular “vitalism”. Cell 2000, 100, 79–88. [Google Scholar] [CrossRef]
- Fankhauser, G. Maintenance of normal structure in heteroploid salamander larvae, through compensation of changes in cell size by adjustment in cell number and cell shape. J. Exp. Zool. 1945, 100, 445–455. [Google Scholar] [CrossRef] [PubMed]
- Fankhauser, G. Memories of great embryologists. Am. Sci. 1972, 40, 46–55. [Google Scholar]
- Campbell, D.T. Studies in the Philosophy of Biology: Reduction and Related Problems; Macmillan: London, UK, 1974. [Google Scholar]
- Noble, D. A theory of biological relativity: No privileged level of causation. Interface Focus 2012, 2, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Rehan, V.K.; Liu, J.; Naeem, E.; Tian, J.; Sakurai, R.; Kwong, K.; Akbari, O.; Torday, J.S. Perinatal nicotine exposure induces asthma in second generation offspring. BMC Med. 2012. [Google Scholar] [CrossRef] [PubMed]
- Waterland, R.A.; Jirtle, R.L. Early nutrition, epigenetic changes at transposons and imprinted genes, and enhanced susceptibility to adult chronic diseases. Nutrition 2004, 20, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Jaenisch, R.; Bird, A. Epigenetic regulation of gene expression: How the genome integrates intrinsic and environmental signals. Nat. Genet. 2003, 33, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Cropley, J.E.; Suter, C.M.; Beckman, K.B.; Martin, D.I. Germ-line epigenetic modification of the murine Avy allele by nutritional supplementation. Proc. Natl. Acad. Sci. USA 2006, 103, 17308–17312. [Google Scholar] [CrossRef] [PubMed]
- Rehan, V.K.; Liu, J.; Sakurai, R.; Torday, J.S. Perinatal nicotine-induced transgenerational asthma. Am. J. Physiol. Lung Cell Mol. Physiol. 2013, 305, L501–L507. [Google Scholar] [CrossRef] [PubMed]
- Darwin, C. On the Origin of Species; John Murray: London, UK, 1859. [Google Scholar]
- Calkins, K.; Devaskar, S.U. Fetal origins of adult disease. Curr. Probl. Pediatr. Adolesc. Health Care 2011, 41, 158–176. [Google Scholar] [CrossRef] [PubMed]
- Warburton, D.; Schwarz, M.; Tefft, D.; Flores-Delgado, G.; Anderson, K.D.; Cardoso, W.V. The molecular basis of lung morphogenesis. Mech. Dev. 2000, 92, 55–81. [Google Scholar] [CrossRef]
- Zheng, W.; Wang, Z.; Collins, J.E.; Andrews, R.M.; Stemple, D.; Gong, Z. Comparative transcriptome analyses indicate molecular homology of zebrafish swimbladder and mammalian lung. PLoS ONE 2011, 6, e24019. [Google Scholar] [CrossRef] [PubMed]
- Grobstein, C. Mechanisms of organogenetic tissue interaction. Natl. Cancer Inst. Monogr. 1967, 26, 279–299. [Google Scholar] [PubMed]
- Berner, R.A.; Vandenbrooks, J.M.; Ward, P.D. Evolution. Oxygen and evolution. Science 2007, 316, 557–558. [Google Scholar] [CrossRef] [PubMed]
- Romer, A.S. The Vertebrate Story; University of Chicago Press: Chicago, IL, USA, 1949. [Google Scholar]
- Rubin, L.P.; Kovacs, C.S.; de Paepe, M.E.; Tsai, S.W.; Torday, J.S.; Kronenberg, H.M. Arrested pulmonary alveolar cytodifferentiation and defective surfactant synthesis in mice missing the gene for parathyroid hormone-related protein. Dev. Dyn. 2004, 230, 278–289. [Google Scholar] [CrossRef] [PubMed]
- Kronenberg, H.M.; Karaplis, A.C.; Lanske, B. Role of parathyroid hormone-related protein in skeletal development. Ann. N. Y. Acad. Sci. 1996, 785, 119–123. [Google Scholar] [CrossRef] [PubMed]
- Bosch, R.J.; Rodríguez-Puyol, D.; Bover, J.; Rodríguez-Puyol, M. Parathyroid hormone-related protein: Roles in the glomerulus. Exp. Nephrol. 1999, 7, 212–216. [Google Scholar] [CrossRef] [PubMed]
- Karaplis, A.C.; Kronenberg, H.M. Physiological roles for parathyroid hormone-related protein: Lessons from gene knockout mice. Vitam. Horm. 1996, 52, 177–193. [Google Scholar] [PubMed]
- Weissmann, G. Evo-Devo and the lungfish: The last gasp of intelligent design. FASEB J. 2007, 21, 623–626. [Google Scholar] [CrossRef] [PubMed]
- Torday, J.S.; Rehan, V.K. The evolutionary continuum from lung development to homeostasis and repair. Am. J. Physiol. Lung Cell Mol. Physiol. 2007, 292, L608–L611. [Google Scholar] [CrossRef] [PubMed]
- Torday, J.S.; Rehan, V.K. A cell-molecular approach predicts vertebrate evolution. Mol. Biol. Evol. 2011, 28, 2973–2981. [Google Scholar]
- De Duve, C. Evolution of the peroxisome. Ann. N. Y. Acad. Sci. 1969, 168, 369–381. [Google Scholar] [CrossRef] [PubMed]
- Clack, J.A. Gaining Ground; Indiana University Press: Bloomington, UK, 2002. [Google Scholar]
- Storr, S.J.; Woolston, C.M.; Zhang, Y.; Martin, S.G. Redox environment, free radical, and oxidative DNA damage. Antioxid. Redox Signal. 2013, 18, 2399–2408. [Google Scholar] [CrossRef] [PubMed]
- Rubin, L.P.; Kifor, O.; Hua, J.; Brown, E.M.; Torday, J.S. Parathyroid hormone (PTH) and PTH-related protein stimulate surfactant phospholipid synthesis in rat fetal lung, apparently by a mesenchymal-epithelial mechanism. Biochim. Biophys. Acta 1994, 1223, 91–100. [Google Scholar] [CrossRef]
- Gao, Y.; Raj, J.U. Parathyroid hormone-related protein-mediated responses in pulmonary arteries and veins of newborn lambs. Am. J. Physiol. Lung Cell Mol. Physiol. 2005, 289, L60–L66. [Google Scholar] [CrossRef] [PubMed]
- West, J.B.; Mathieu-Costello, O. Structure, strength, failure, and remodeling of the pulmonary blood-gas barrier. Annu. Rev. Physiol. 1999, 61, 543–572. [Google Scholar] [CrossRef] [PubMed]
- Kawashima, M.; Takahashi, T.; Yanai, H.; Ogawa, H.; Yasuoka, T. Direct action of parathyroid hormone-related peptide to enhance corticosterone production stimulated by adrenocorticotropic hormone in adrenocortical cells of hens. Poult. Sci. 2005, 84, 1463–1469. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, H.; Takahashi, T.; Oomatsu, Y.; Nakagawa-Mizuyachi, K.; Kawashima, M. Parathyroid hormone-related peptide directly increases adrenocorticotropic hormone secretion from the anterior pituitary in hens. Poult. Sci. 2011, 90, 175–180. [Google Scholar] [CrossRef] [PubMed]
- Kvetnansky, R.; Lu, X.; Ziegler, M.G. Stress-triggered changes in peripheral catecholaminergic systems. Adv. Pharmacol. 2013, 68, 359–397. [Google Scholar] [PubMed]
- Wurtman, R.J. Stress and the adrenocortical control of epinephrine synthesis. Metabolism 2002, 51, 11–14. [Google Scholar] [CrossRef] [PubMed]
- Park, S.I.; Lee, C.; Sadler, W.D.; Koh, A.J.; Jones, J.; Seo, J.W.; Soki, F.N.; Cho, S.W.; Daignault, S.D.; McCauley, L.K. Parathyroid hormone-related protein drives a CD11b+Gr1+ cell-mediated positive feedback loop to support prostate cancer growth. Cancer Res. 2013, 73, 6574–6583. [Google Scholar] [CrossRef] [PubMed]
- Lawson, E.E.; Brown, E.R.; Torday, J.S.; Madansky, D.L.; Taeusch, H.W., Jr. The effect of epinephrine on tracheal fluid flow and surfactant efflux in fetal sheep. Am. Rev. Respir. Dis. 1978, 118, 1023–1026. [Google Scholar] [PubMed]
- Rohrer, D.K.; Desai, K.H.; Jasper, J.R.; Stevens, M.E.; Regula, D.P., Jr.; Barsh, G.S.; Bernstein, D.; Kobilka, B.K. Targeted disruption of the mouse beta1-adrenergic receptor gene: Developmental and cardiovascular effects. Proc. Natl. Acad. Sci. USA 1996, 93, 7375–7380. [Google Scholar] [CrossRef] [PubMed]
- Simões-Costa, M.S.; Vasconcelos, M.; Sampaio, A.C.; Cravo, R.M.; Linhares, V.L.; Hochgreb, T.; Yan, C.Y.; Davidson, B.; Xavier-Neto, J. The evolutionary origin of cardiac chambers. Dev. Biol. 2005, 277, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Smith, H.W. From Fish to Philosopher; Little Brown: Boston, MA, USA, 1953. [Google Scholar]
- Hart, P.D.; Bakris, G.L. Should beta-blockers be used to control hypertension in people with chronic kidney disease? Semin. Nephrol. 2007, 27, 555–564. [Google Scholar] [CrossRef] [PubMed]
- Cerny, L.; Torday, J.S.; Rehan, V.K. Prevention and treatment of bronchopulmonary dysplasia: Contemporary status and future outlook. Lung 2008, 186, 75–89. [Google Scholar] [CrossRef] [PubMed]
- Rehan, V.K.; Torday, J.S. Lower parathyroid hormone-related protein content of tracheal aspirates in very low birth weight infants who develop bronchopulmonary dysplasia. Pediatr. Res. 2006, 60, 216–220. [Google Scholar] [CrossRef] [PubMed]
- Wong, D.L. Epinephrine biosynthesis: Hormonal and neural control during stress. Cell Mol. Neurobiol. 2006, 26, 891–900. [Google Scholar] [CrossRef] [PubMed]
- Kolbert, E. The Sixth Extinction; Henry Holt and Company: New York, NY, USA, 2014. [Google Scholar]
- Mayr, E. Cause and effect in biology. Science 1961, 134, 1501–1506. [Google Scholar] [CrossRef] [PubMed]
- Longo, S.; Bollani, L.; Decembrino, L.; di Comite, A.; Angelini, M.; Stronati, M. Short-term and long-term sequelae in intrauterine growth retardation (IUGR). J. Matern. Fetal Neonatal. Med. 2013, 26, 222–225. [Google Scholar] [CrossRef] [PubMed]
- Salam, R.A.; Das, J.K.; Bhutta, Z.A. Impact of intrauterine growth restriction on long-term health. Curr. Opin. Clin. Nutr. Metab. Care 2014, 17, 249–254. [Google Scholar] [CrossRef] [PubMed]
- Ross, M.G.; Desai, M. Developmental programming of offspring obesity, adipogenesis, and appetite. Clin. Obstet. Gynecol. 2013, 56, 529–536. [Google Scholar] [CrossRef] [PubMed]
- Duhem, P.; Wiener, P.P. The Aim and Structure of Physical Theory; Princeton University Press: Princeton, NJ, USA, 1954. [Google Scholar]
- Quine, W.O.V. Word and Object; MIT Press: Cambridge, UK, 2015. [Google Scholar]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Torday, J.S. Homeostasis as the Mechanism of Evolution. Biology 2015, 4, 573-590. https://doi.org/10.3390/biology4030573
Torday JS. Homeostasis as the Mechanism of Evolution. Biology. 2015; 4(3):573-590. https://doi.org/10.3390/biology4030573
Chicago/Turabian StyleTorday, John S. 2015. "Homeostasis as the Mechanism of Evolution" Biology 4, no. 3: 573-590. https://doi.org/10.3390/biology4030573