
Neutrophil Development, Migration, and Function in Teleost Fish

{kind=link}
Abstract
:1. Introduction
2. The Life of a Neutrophil
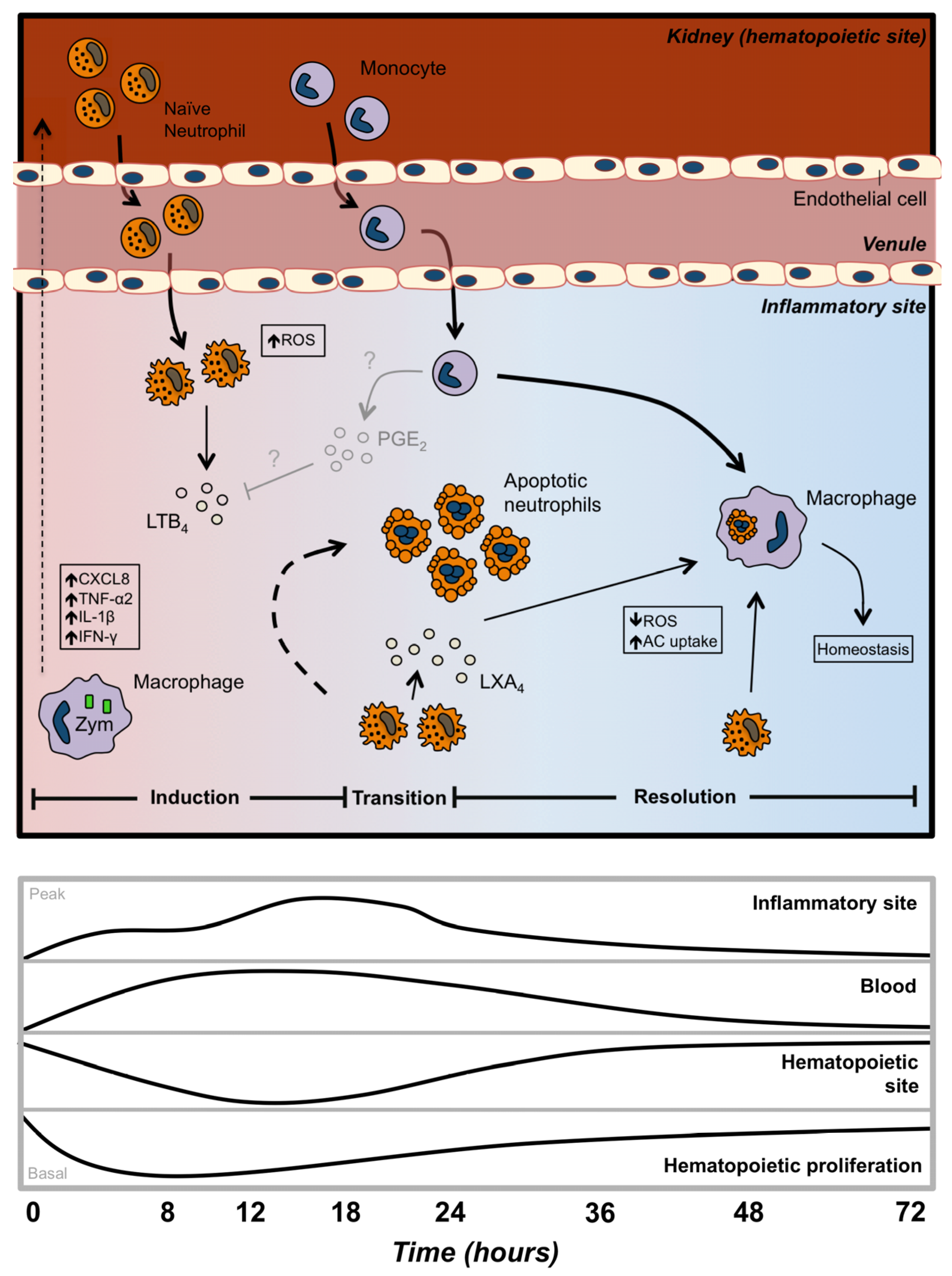
2.1. Neutrophil Development
2.2. Neutrophil Reservoirs
2.3. Neutrophil Migration
3. Neutrophils and the Promotion of Inflammation
3.1. Degranulation
3.2. Neutrophil Extracellular Traps (NETs)
3.3. Respiratory Burst
3.4. Nitric Oxide
4. Neutrophils and the Resolution of Inflammation
4.1. Production of Lipid Mediators
4.2. Death of a Neutrophil
5. Conclusions

Acknowledgments
Author Contributions
Conflicts of Interest
References
- Havixbeck, J.J.; Rieger, A.M.; Wong, M.E.; Hodgkinson, J.W.; Barreda, D.R. Neutrophil contributions to the induction and regulation of the acute inflammatory response in teleost fish. J. Leukoc. Biol. 2015. [Google Scholar] [CrossRef] [PubMed]
- Meseguer, J.; López-Ruiz, A.; Esteban, M.A. Cytochemical characterization of leucocytes from the seawater teleost, gilthead seabream (Sparus aurata L.). Histochemistry 1994, 102, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Flerova, E.A.; Balabanova, L.V. Ultrastructure of granulocytes of teleost fish (Salmoniformes, Cypriniformes, Perciformes). J. Evol. Biochem. Physiol. 2013, 49, 223–233. [Google Scholar] [CrossRef]
- Rieger, A.M.; Konowalchuk, J.D.; Grayfer, L.; Katzenback, B.A.; Havixbeck, J.J.; Kiemele, M.D.; Belosevic, M.; Barreda, D.R. Fish and mammalian phagocytes differentially regulate pro-inflammatory and homeostatic responses in vivo. PLoS ONE 2012, 7, e47070. [Google Scholar] [CrossRef] [PubMed]
- Katzenback, B.A.; Belosevic, M. Isolation and functional characterization of neutrophil-like cells, from goldfish (Carassius auratus L.) kidney. Dev. Comp. Immunol. 2009, 33, 601–611. [Google Scholar] [CrossRef] [PubMed]
- Filho, D.W. Reactive oxygen species, antioxidants and fish mitochondria. Front. Biosci. 2007, 12, 1229–1237. [Google Scholar] [CrossRef]
- Palić, D.; Andreasen, C.B.; Ostojić, J.; Tell, R.M.; Roth, J.A. Zebrafish (Danio rerio) whole kidney assays to measure neutrophil extracellular trap release and degranulation of primary granules. J. Immunol. Methods 2007, 319, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Pijanowski, L.; Golbach, L.; Kolaczkowska, E.; Scheer, M.; Verburg-van Kemenade, B.M.L.; Chadzinska, M. Carp neutrophilic granulocytes form extracellular traps via ROS-dependent and independent pathways. Fish Shellfish Immunol. 2013, 34, 1244–1252. [Google Scholar] [CrossRef] [PubMed]
- Borregaard, N. Neutrophils, from Marrow to Microbes. Immunity 2010, 33, 657–670. [Google Scholar] [CrossRef] [PubMed]
- Prokopowicz, Z.; Marcinkiewicz, J.; Katz, D.R.; Chain, B.M. Neutrophil myeloperoxidase: Soldier and statesman. Arch. Immunol. Ther. Exp. 2012, 60, 43–54. [Google Scholar] [CrossRef] [PubMed]
- Nathan, C. Neutrophils and immunity: Challenges and opportunities. Nat. Rev. Immunol. 2006, 6, 173–182. [Google Scholar] [CrossRef] [PubMed]
- Winterbourn, C.C.; Kettle, A.J. Redox reactions and microbial killing in the neutrophil phagosome. Antioxid. Redox Signal. 2012, 18, 642–660. [Google Scholar] [CrossRef] [PubMed]
- Brinkmann, V.; Reichard, U.; Goosmann, C.; Fauler, B.; Uhlemann, Y.; Weiss, D.S.; Weinrauch, Y.; Zychlinsky, A. Neutrophil extracellular traps kill bacteria. Science 2004, 303, 1532–1535. [Google Scholar] [CrossRef] [PubMed]
- Brinkmann, V.; Zychlinsky, A. Neutrophil extracellular traps: Is immunity the second function of chromatin? J. Cell Biol. 2012, 198, 773–783. [Google Scholar] [CrossRef] [PubMed]
- Rieger, A.M.; Barreda, D.R. Antimicrobial mechanisms of fish leukocytes. Dev. Comp. Immunol. 2011, 35, 1238–1245. [Google Scholar] [CrossRef] [PubMed]
- Secombes, C.J.; Fletcher, T.C. The role of phagocytes in the protective mechanisms of fish. Annu. Rev. Fish Dis. 1992, 2, 53–71. [Google Scholar] [CrossRef]
- Neumann, N.F.; Stafford, J.L.; Barreda, D.; Ainsworth, A.J.; Belosevic, M. Antimicrobial mechanisms of fish phagocytes and their role in host defense. Dev. Comp. Immunol. 2001, 25, 807–825. [Google Scholar] [CrossRef]
- Plouffe, D.A.; Hanington, P.C.; Walsh, J.G.; Wilson, E.C.; Belosevic, M. Comparison of select innate immune mechanisms of fish and mammals. Xenotransplantation 2005, 12, 266–277. [Google Scholar] [CrossRef] [PubMed]
- Magnadóttir, B. Innate immunity of fish (overview). Fish Shellfish Immunol. 2006, 20, 137–151. [Google Scholar] [CrossRef] [PubMed]
- Robertsen, B. The interferon system of teleost fish. Fish Shellfish Immunol. 2006, 20, 172–191. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Pellitero, P. Fish immunity and parasite infections: From innate immunity to immunoprophylactic prospects. Vet. Immunol. Immunopathol. 2008, 126, 171–198. [Google Scholar] [CrossRef] [PubMed]
- Traver, D.; Paw, B.H.; Poss, K.D.; Penberthy, W.T.; Lin, S.; Zon, L.I. Transplantation and in vivo imaging of multilineage engraftment in zebrafish bloodless mutants. Nat. Immunol. 2003, 4, 1238–1246. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, I.; Kuniyoshi, S.; Saito, K.; Moritomo, T.; Takahashi, T.; Nakanishi, T. Long-term hematopoietic reconstitution by transplantation of kidney hematopoietic stem cells in lethally irradiated clonal ginbuna crucian carp (Carassius auratus langsdorfii). Dev. Comp. Immunol. 2008, 32, 957–965. [Google Scholar] [CrossRef] [PubMed]
- Galloway, J.L.; Zon, L.I. 3 Ontogeny of hematopoiesis: Examining the emergence of hematopoietic cells in the vertebrate embryo. Curr. Top. Dev. Biol. 2003, 53, 139–158. [Google Scholar] [PubMed]
- Davidson, A.J.; Zon, L.I. The “definitive” (and “primitive”) guide to zebrafish hematopoiesis. Oncogene 2004, 23, 7233–7246. [Google Scholar] [CrossRef] [PubMed]
- Guyader, D.L.; Redd, M.J.; Colucci-Guyon, E.; Murayama, E.; Kissa, K.; Briolat, V.; Mordelet, E.; Zapata, A.; Shinomiya, H.; Herbomel, P. Origins and unconventional behavior of neutrophils in developing zebrafish. Blood 2008, 111, 132–141. [Google Scholar] [CrossRef] [PubMed]
- Murayama, E.; Kissa, K.; Zapata, A.; Mordelet, E.; Briolat, V.; Lin, H.-F.; Handin, R.I.; Herbomel, P. Tracing hematopoietic Precursor migration to successive hematopoietic organs during zebrafish development. Immunity 2006, 25, 963–975. [Google Scholar] [CrossRef] [PubMed]
- Willett, C.E.; Cortes, A.; Zuasti, A.; Zapata, A.G. Early hematopoiesis and developing lymphoid organs in the zebrafish. Dev. Dyn. 1999, 214, 323–336. [Google Scholar] [CrossRef]
- Metcalf, D. Hematopoietic cytokines. Blood 2008, 111, 485–491. [Google Scholar] [CrossRef] [PubMed]
- Rieger, M.A.; Hoppe, P.S.; Smejkal, B.M.; Eitelhuber, A.C.; Schroeder, T. Hematopoietic cytokines can instruct lineage choice. Science 2009, 325, 217–218. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Emerson, S.G. Hematopoietic cytokines, transcription factors and lineage commitment. Oncogene 2002, 21, 3295–3313. [Google Scholar] [CrossRef] [PubMed]
- Rosmarin, A.G.; Yang, Z.; Resendes, K.K. Transcriptional regulation in myelopoiesis: Hematopoietic fate choice, myeloid differentiation, and leukemogenesis. Exp. Hematol. 2005, 33, 131–143. [Google Scholar] [CrossRef] [PubMed]
- Lenny, N.; Westendorf, J.J.; Hiebert, S.W. Transcriptional regulation during myelopoiesis. Mol. Biol. Rep. 1997, 24, 157–168. [Google Scholar] [CrossRef] [PubMed]
- Hermans, M.H.A.; van de Geijn, G.-J.; Antonissen, C.; Gits, J.; van Leeuwen, D.; Ward, A.C.; Touw, I.P. Signaling mechanisms coupled to tyrosines in the granulocyte colony-stimulating factor receptor orchestrate G-CSF-induced expansion of myeloid progenitor cells. Blood 2003, 101, 2584–2590. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Wu, H.Y.; Wesselschmidt, R.; Kornaga, T.; Link, D.C. Impaired production and increased apoptosis of neutrophils in granulocyte colony-stimulating factor receptor-deficient mice. Immunity 1996, 5, 491–501. [Google Scholar] [CrossRef]
- Villunger, A.; Scott, C.; Bouillet, P.; Strasser, A. Essential role for the BH3-only protein Bim but redundant roles for Bax, Bcl-2, and Bcl-w in the control of granulocyte survival. Blood 2003, 101, 2393–2400. [Google Scholar] [CrossRef] [PubMed]
- Liongue, C.; Hall, C.J.; O’Connell, B.A.; Crosier, P.; Ward, A.C. Zebrafish granulocyte colony-stimulating factor receptor signaling promotes myelopoiesis and myeloid cell migration. Blood 2009, 113, 2535–2546. [Google Scholar] [CrossRef] [PubMed]
- Shivdasani, R.A.; Mayer, E.L.; Orkin, S.H. Absence of blood formation in mice lacking the T-cell leukaemia oncoprotein tal-1/SCL. Nature 1995, 373, 432–434. [Google Scholar] [CrossRef] [PubMed]
- Robb, L.; Lyons, I.; Li, R.; Hartley, L.; Köntgen, F.; Harvey, R.P.; Metcalf, D.; Begley, C.G. Absence of yolk sac hematopoiesis from mice with a targeted disruption of the scl gene. Proc. Natl. Acad. Sci. 1995, 92, 7075–7079. [Google Scholar] [CrossRef] [PubMed]
- Rey Vázquez, G.; Guerrero, G.A. Characterization of blood cells and hematological parameters in Cichlasoma dimerus (Teleostei, Perciformes). Tissue Cell 2007, 39, 151–160. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, Y.; Maita, M.; Okamoto, N. Rainbow trout neutrophils are responsible for non-specific cytotoxicity. Fish Shellfish Immunol. 2002, 14, 243–252. [Google Scholar] [CrossRef]
- Hamdani, S.H.; McMillan, D.N.; Pettersen, E.F.; Wergeland, H.; Endresen, C.; Ellis, A.E.; Secombes, C.J. Isolation of rainbow trout neutrophils with an anti-granulocyte monoclonal antibody. Vet. Immunol. Immunopathol. 1998, 63, 369–380. [Google Scholar] [CrossRef]
- Scharsack, J.P.; Steinhagen, D.; Kleczka, C.; Schmidt, J.O.; Körting, W.; Michael, R.D.; Leibold, W.; Schuberth, H.J. Head kidney neutrophils of carp (Cyprinus carpio L.) are functionally modulated by the haemoflagellate Trypanoplasma borreli. Fish Shellfish Immunol. 2003, 14, 389–403. [Google Scholar] [CrossRef] [PubMed]
- Woo, P.T.K.; Leatherland, J.F.; Bruno, D.W. Fish Diseases and Disorders; CABI International: Wallingford, UK, 2011. [Google Scholar]
- McDonald, B.; Pittman, K.; Menezes, G.B.; Hirota, S.A.; Slaba, I.; Waterhouse, C.C.M.; Beck, P.L.; Muruve, D.A.; Kubes, P. Intravascular danger signals guide neutrophils to sites of sterile inflammation. Science 2010, 330, 362–366. [Google Scholar] [CrossRef] [PubMed]
- Niethammer, P.; Grabher, C.; Look, A.T.; Mitchison, T.J. A tissue-scale gradient of hydrogen peroxide mediates rapid wound detection in zebrafish. Nature 2009, 459, 996–999. [Google Scholar] [CrossRef] [PubMed]
- Renshaw, S.A.; Loynes, C.A.; Trushell, D.M.I.; Elworthy, S.; Ingham, P.W.; Whyte, M.K.B. A transgenic zebrafish model of neutrophilic inflammation. Blood 2006, 108, 3976–3978. [Google Scholar] [CrossRef] [PubMed]
- Mathias, J.R.; Perrin, B.J.; Liu, T.-X.; Kanki, J.; Look, A.T.; Huttenlocher, A. Resolution of inflammation by retrograde chemotaxis of neutrophils in transgenic zebrafish. J. Leukoc. Biol. 2006, 80, 1281–1288. [Google Scholar] [CrossRef] [PubMed]
- Lieschke, G.J.; Trede, N.S. Fish immunology. Curr. Biol. 2009, 19, R678–R682. [Google Scholar] [CrossRef] [PubMed]
- Van der Aa, L.M.; Chadzinska, M.; Tijhaar, E.; Boudinot, P.; Verburg-van Kemenade, B.M.L. CXCL8 chemokines in teleost fish: Two lineages with distinct expression profiles during early phases of inflammation. PLoS ONE 2010, 5. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Xu, Q.; Wang, T.; Collet, B.; Corripio-Miyar, Y.; Bird, S.; Xie, P.; Nie, P.; Secombes, C.J.; Zou, J. Phylogenetic analysis of vertebrate CXC chemokines reveals novel lineage specific groups in teleost fish. Dev. Comp. Immunol. 2013, 41, 137–152. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, S.; Reyes-Aldasoro, C.C.; Candel, S.; Renshaw, S.A.; Mulero, V.; Calado, A. Cxcl8 (IL-8) mediates neutrophil recruitment and behavior in the zebrafish inflammatory response. J. Immunol. 2013, 190, 4349–4359. [Google Scholar] [CrossRef] [PubMed]
- Oehlers, S.H.B.; Flores, M.V.; Hall, C.J.; O’Toole, R.; Swift, S.; Crosier, K.E.; Crosier, P.S. Expression of zebrafish cxcl8 (interleukin-8) and its receptors during development and in response to immune stimulation. Dev. Comp. Immunol. 2010, 34, 352–359. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, S.; Lopez-Muñoz, A.; Martínez-Navarro, F.J.; Galindo-Villegas, J.; Mulero, V.; Calado, Â. Cxcl8-l1 and Cxcl8-l2 are required in the zebrafish defense against Salmonella Typhimurium. Dev. Comp. Immunol. 2015, 49, 44–48. [Google Scholar] [CrossRef] [PubMed]
- Yoo, S.K.; Deng, Q.; Cavnar, P.J.; Wu, Y.I.; Hahn, K.M.; Huttenlocher, A. Differential regulation of protrusion and polarity by PI(3)K during neutrophil motility in live zebrafish. Dev. Cell 2010, 18, 226–236. [Google Scholar] [CrossRef] [PubMed]
- Buckley, C.D.; Ross, E.A.; McGettrick, H.M.; Osborne, C.E.; Haworth, O.; Schmutz, C.; Stone, P.C.W.; Salmon, M.; Matharu, N.M.; Vohra, R.K.; et al. Identification of a phenotypically and functionally distinct population of long-lived neutrophils in a model of reverse endothelial migration. J. Leukoc. Biol. 2006, 79, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Forlenza, M.; Scharsack, J.P.; Kachamakova, N.M.; Taverne-Thiele, A.J.; Rombout, J.H.W.M.; Wiegertjes, G.F. Differential contribution of neutrophilic granulocytes and macrophages to nitrosative stress in a host-parasite animal model. Mol. Immunol. 2008, 45, 3178–3189. [Google Scholar] [CrossRef] [PubMed]
- Mayumi, M.; Takeda, Y.; Hoshiko, M.; Serada, K.; Murata, M.; Moritomo, T.; Takizawa, F.; Kobayashi, I.; Araki, K.; Nakanishi, T.; et al. Characterization of teleost phagocyte NADPH oxidase: Molecular cloning and expression analysis of carp (Cyprinus carpio) phagocyte NADPH oxidase. Mol. Immunol. 2008, 45, 1720–1731. [Google Scholar] [CrossRef] [PubMed]
- Moritomo, T.; Serata, K.; Teshirogi, K.; Aikawa, H.; Inoue, Y.; Itou, T.; Nakanishi, T. Flow cytometric analysis of the neutrophil respiratory burst of ayu, Plecoglossus altivelis: Comparison with other fresh water fish. Fish Shellfish Immunol. 2003, 15, 29–38. [Google Scholar] [CrossRef]
- Leiro, J.; Iglesias, R.; Paramá, A.; Sanmartı́n, M.L.; Ubeira, F.M. Effect of Tetramicra brevifilum (Microspora) infection on respiratory-burst responses of turbot (Scophthalmus maximus L.) phagocytes. Fish Shellfish Immunol. 2001, 11, 639–652. [Google Scholar] [CrossRef] [PubMed]
- Cuesta, A.; Meseguer, J.; Esteban, M.Á. The antimicrobial peptide hepcidin exerts an important role in the innate immunity against bacteria in the bony fish gilthead seabream. Mol. Immunol. 2008, 45, 2333–2342. [Google Scholar] [CrossRef] [PubMed]
- Mulero, I.; Noga, E.J.; Meseguer, J.; García-Ayala, A.; Mulero, V. The antimicrobial peptides piscidins are stored in the granules of professional phagocytic granulocytes of fish and are delivered to the bacteria-containing phagosome upon phagocytosis. Dev. Comp. Immunol. 2008, 32, 1531–1538. [Google Scholar] [CrossRef] [PubMed]
- Morel, F.; Doussiere, J.; Vignais, P.V. The superoxide-generating oxidase of phagocytic cells. Eur. J. Biol. 1991, 201, 523–546. [Google Scholar] [CrossRef]
- Palić, D.; Andreasen, C.B.; Menzel, B.W.; Roth, J.A. A rapid, direct assay to measure degranulation of primary granules in neutrophils from kidney of fathead minnow (Pimephales promelas Rafinesque, 1820). Fish Shellfish Immunol. 2005, 19, 217–227. [Google Scholar] [CrossRef] [PubMed]
- Wernersson, S.; Reimer, J.M.; Poorafshar, M.; Karlson, U.; Wermenstam, N.; Bengtén, E.; Wilson, M.; Pilström, L.; Hellman, L. Granzyme-like sequences in bony fish shed light on the emergence of hematopoietic serine proteases during vertebrate evolution. Dev. Comp. Immunol. 2006, 30, 901–918. [Google Scholar] [CrossRef] [PubMed]
- Urban, C.F.; Ermert, D.; Schmid, M.; Abu-Abed, U.; Goosmann, C.; Nacken, W.; Brinkmann, V.; Jungblut, P.R.; Zychlinsky, A. Neutrophil extracellular traps contain calprotectin, a cytosolic protein complex involved in host defense against candida albicans. PLoS Pathog. 2009, 5. [Google Scholar] [CrossRef] [PubMed]
- Palić, D.; Ostojić, J.; Andreasen, C.B.; Roth, J.A. Fish cast NETs: Neutrophil extracellular traps are released from fish neutrophils. Dev. Comp. Immunol. 2007, 31, 805–816. [Google Scholar] [CrossRef] [PubMed]
- Baldridge, C.W.; Gerard, R.W. The extra respiration of phagocytosis. Am. J. Physiol. 1933, 103, 235–236. [Google Scholar]
- Babior, B.M.; Kipnes, R.S.; Curnutte, J.T. Biological Defense Mechanisms. The production by leukocytes of superoxide, a potential bactericidal agent. J. Clin. Investig. 1973, 52, 741–744. [Google Scholar] [CrossRef] [PubMed]
- Briggs, R.; Drath, D.B.; Karnovsky, M.L.; Karnovsky, M.J. Localization of NADH oxidase on the surface of human polymorphonuclear leukocytes by a new cytochemical method. J. Cell Biol. 1975, 67, 566–586. [Google Scholar] [CrossRef] [PubMed]
- Boltaña, S.; Doñate, C.; Goetz, F.W.; MacKenzie, S.; Balasch, J.C. Characterization and expression of NADPH oxidase in LPS-, poly(I:C)- and zymosan-stimulated trout (Oncorhynchus mykiss W.) macrophages. Fish Shellfish Immunol. 2009, 26, 651–661. [Google Scholar] [CrossRef] [PubMed]
- Inoue, Y.; Suenaga, Y.; Yoshiura, Y.; Moritomo, T.; Ototake, M.; Nakanishi, T. Molecular cloning and sequencing of Japanese pufferfish (Takifugu rubripes) NADPH oxidase cDNAs. Dev. Comp. Immunol. 2004, 28, 911–925. [Google Scholar] [CrossRef] [PubMed]
- Olavarría, V.H.; Gallardo, L.; Figueroa, J.E.; Mulero, V. Lipopolysaccharide primes the respiratory burst of Atlantic salmon SHK-1 cells through protein kinase C-mediated phosphorylation of p47phox. Dev. Comp. Immunol. 2010, 34, 1242–1253. [Google Scholar] [CrossRef] [PubMed]
- Kawahara, T.; Lambeth, J.D. Molecular evolution of Phox-related regulatory subunits for NADPH oxidase enzymes. BMC Evol. Biol. 2007, 7. [Google Scholar] [CrossRef] [PubMed]
- Kawahara, T.; Quinn, M.T.; Lambeth, J.D. Molecular evolution of the reactive oxygen-generating NADPH oxidase (Nox/Duox) family of enzymes. BMC Evol. Biol. 2007, 7. [Google Scholar] [CrossRef] [PubMed]
- Nathan, C.F. Respiratory burst in adherent human neutrophils: Triggering by colony-stimulating factors CSF-GM and CSF-G. Blood 1989, 73, 301–306. [Google Scholar] [PubMed]
- Little, R.; White, M.R.; Hartshorn, K.L. Interferon-α enhances neutrophil respiratory burst responses to stimulation with influenza a virus and FMLP. J. Infect. Dis. 1994, 170, 802–810. [Google Scholar] [CrossRef] [PubMed]
- Dewald, B.; Baggiolini, M. Activation of NADPH oxidase in human neutrophils. Synergism between fMLP and the neutrophil products PAF and LTB4. Biochem. Biophys. Res. Commun. 1985, 128, 297–304. [Google Scholar] [CrossRef]
- Grayfer, L.; Belosevic, M. Molecular characterization, expression and functional analysis of goldfish (Carassius aurutus L.) interferon gamma. Dev. Comp. Immunol. 2009, 33, 235–246. [Google Scholar] [CrossRef] [PubMed]
- Grayfer, L.; Walsh, J.G.; Belosevic, M. Characterization and functional analysis of goldfish (Carassius auratus L.) tumor necrosis factor-alpha. Dev. Comp. Immunol. 2008, 32, 532–543. [Google Scholar] [CrossRef] [PubMed]
- Zou, J.; Carrington, A.; Collet, B.; Dijkstra, J.M.; Yoshiura, Y.; Bols, N.; Secombes, C. Identification and Bioactivities of IFN-γ in Rainbow Trout Oncorhynchus mykiss: The First Th1-Type Cytokine Characterized Functionally in Fish. J. Immunol. 2005, 175, 2484–2494. [Google Scholar] [CrossRef] [PubMed]
- Arts, J.A.J.; Tijhaar, E.J.; Chadzinska, M.; Savelkoul, H.F.J.; Verburg-van Kemenade, B.M.L. Functional analysis of carp interferon-γ: Evolutionary conservation of classical phagocyte activation. Fish Shellfish Immunol. 2010, 29, 793–802. [Google Scholar] [CrossRef] [PubMed]
- Sepulcre, M.P.; López-Castejón, G.; Meseguer, J.; Mulero, V. The activation of gilthead seabream professional phagocytes by different PAMPs underlines the behavioural diversity of the main innate immune cells of bony fish. Mol. Immunol. 2007, 44, 2009–2016. [Google Scholar] [CrossRef] [PubMed]
- Sepulcre, M.P.; López-Muñoz, A.; Angosto, D.; García-Alcazar, A.; Meseguer, J.; Mulero, V. TLR agonists extend the functional lifespan of professional phagocytic granulocytes in the bony fish gilthead seabream and direct precursor differentiation towards the production of granulocytes. Mol. Immunol. 2011, 48, 846–859. [Google Scholar] [CrossRef] [PubMed]
- Kemenade, B.; Groeneveld, A.; Rens, B.; Rombout, J. Characterization of macrophages and neutrophilic granulocytes from the pronephros of carp (Cyprinus carpio). J. Exp. Biol. 1994, 187, 143–158. [Google Scholar] [PubMed]
- Chaves-Pozo, E.; Muñoz, P.; López-Muñoz, A.; Pelegrín, P.; Ayala, A.G.; Mulero, V.; Meseguer, J. Early innate immune response and redistribution of inflammatory cells in the bony fish gilthead seabream experimentally infected with Vibrio anguillarum. Cell Tissue Res. 2005, 320, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Serada, K.; Moritomo, T.; Teshirogi, K.; Itou, T.; Shibashi, T.; Inoue, Y.; Nakanishi, T. Comparison of respiratory burst activity of inflammatory neutrophils in ayu (Plecoglossus altivelis) and carp (Cyprinus carpio). Fish Shellfish Immunol. 2005, 19, 363–373. [Google Scholar] [CrossRef] [PubMed]
- Hopper, R.A.; Garthwaite, J. Tonic and phasic nitric oxide signals in hippocampal long-term potentiation. J. Neurosci. 2006, 26, 11513–11521. [Google Scholar] [CrossRef] [PubMed]
- Napoli, C.; Paolisso, G.; Casamassimi, A.; Al-Omran, M.; Barbieri, M.; Sommese, L.; Infante, T.; Ignarro, L.J. Effects of nitric oxide on cell proliferation: Novel insights. J. Am. Coll. Cardiol. 2013, 62, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Paniagua, O.A.; Bryant, M.B.; Panza, J.A. Role of endothelial nitric oxide in shear stress-induced vasodilation of human microvasculature diminished activity in hypertensive and Hypercholesterolemic patients. Circulation 2001, 103, 1752–1758. [Google Scholar] [CrossRef] [PubMed]
- Clementi, E. Role of nitric oxide and its intracellular signalling pathways in the control of Ca2 homeostasis. Biochem. Pharmacol. 1998, 55, 713–718. [Google Scholar] [CrossRef]
- MacMicking, J.; Xie, Q.; Nathan, C. Nitric oxide and macrophage function. Annu. Rev. Immunol. 1997, 15, 323–350. [Google Scholar] [CrossRef] [PubMed]
- Knowles, R.G.; Moncada, S. Nitric oxide synthases in mammals. Biochem. J. 1994, 298, 249–258. [Google Scholar] [CrossRef] [PubMed]
- Laing, K.J.; Grabowski, P.S.; Belosevic, M.; Secombes, C.J. A partial sequence for nitric oxide synthase from a goldfish (Carassius auratus) macrophage cell line. Immunol. Cell Biol. 1996, 74, 374–379. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Ward, M.; Grabowski, P.; Secombes, C.J. Molecular cloning, gene organization and expression of rainbow trout (Oncorhynchus mykiss) inducible nitric oxide synthase (iNOS) gene. Biochem. J. 2001, 358, 747–755. [Google Scholar] [CrossRef] [PubMed]
- Saeij, J.P.J.; Stet, R.J.M.; Groeneveld, A.; Kemenade, L.B.M.V.; van Muiswinkel, W.B.; Wiegertjes, G.F. Molecular and functional characterization of a fish inducible-type nitric oxide synthase. Immunogenetics 2000, 51, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Barroso, J.B.; Carreras, A.; Esteban, F.J.; Peinado, M.A.; Martínez-Lara, E.; Valderrama, R.; Jiménez, A.; Rodrigo, J.; Lupiáñez, J.A. Molecular and kinetic characterization and cell type location of inducible nitric oxide synthase in fish. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2000, 279, R650–R656. [Google Scholar] [PubMed]
- Tafalla, C.; Figueras, A.; Novoa, B. Role of nitric oxide on the replication of viral haemorrhagic septicemia virus (VHSV), a fish rhabdovirus. Vet. Immunol. Immunopathol. 1999, 72, 249–256. [Google Scholar] [CrossRef]
- CamposPrez, J.J.; Ellis, A.E.; Secombes, C.J. Toxicity of nitric oxide and peroxynitrite to bacterial pathogens of fish. Dis. Aquat. Organ. 2000, 43, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Wiegertjes, G.F.; Forlenza, M. Nitrosative stress during infection-induced inflammation in fish: Lessons from a host-parasite infection model. Curr. Pharm. Des. 2010, 16, 4194–4202. [Google Scholar] [CrossRef] [PubMed]
- Saeij, J.P.J.; van Muiswinkel, W.B.; van de Meent, M.; Amaral, C.; Wiegertjes, G.F. Different capacities of carp leukocytes to encounter nitric oxide-mediated stress: A role for the intracellular reduced glutathione pool. Dev. Comp. Immunol. 2003, 27, 555–568. [Google Scholar] [CrossRef]
- Serhan, C.N.; Chiang, N.; van Dyke, T.E. Resolving inflammation: Dual anti-inflammatory and pro-resolution lipid mediators. Nat. Rev. Immunol. 2008, 8, 349–361. [Google Scholar] [CrossRef] [PubMed]
- Serhan, C.N.; Savill, J. Resolution of inflammation: The beginning programs the end. Nat. Immunol. 2005, 6, 1191–1197. [Google Scholar] [CrossRef] [PubMed]
- Serhan, C.N. Pro-resolving lipid mediators are leads for resolution physiology. Nature 2014, 510, 92–101. [Google Scholar] [CrossRef] [PubMed]
- Levy, B.D.; Clish, C.B.; Schmidt, B.; Gronert, K.; Serhan, C.N. Lipid mediator class switching during acute inflammation: Signals in resolution. Nat. Immunol. 2001, 2, 612–619. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.; Gronert, K.; Devchand, P.R.; Moussignac, R.L.; Serhan, C.N. Novel docosatrienes and 17S-resolvins generated from docosahexaenoic acid in murine brain, human blood, and glial cells autacoids in anti-inflammation. J. Biol. Chem. 2003, 278, 14677–14687. [Google Scholar] [CrossRef] [PubMed]
- Serhan, C.N.; Clish, C.B.; Brannon, J.; Colgan, S.P.; Chiang, N.; Gronert, K. Novel functional sets of lipid-derived mediators with antiinflammatory actions generated from omega-3 fatty acids via cyclooxygenase 2-nonsteroidal antiinflammatory drugs and transcellular processing. J. Exp. Med. 2000, 192, 1197–1204. [Google Scholar] [CrossRef] [PubMed]
- Schwab, J.M.; Chiang, N.; Arita, M.; Serhan, C.N. Resolvin E1 and protectin D1 activate inflammation-resolution programs. Nature 2007, 447, 869–874. [Google Scholar] [CrossRef] [PubMed]
- Canny, G.; Levy, O.; Furuta, G.T.; Narravula-Alipati, S.; Sisson, R.B.; Serhan, C.N.; Colgan, S.P. Lipid mediator-induced expression of bactericidal/permeability-increasing protein (BPI) in human mucosal epithelia. Proc. Natl. Acad. Sci. USA 2002, 99, 3902–3907. [Google Scholar] [CrossRef] [PubMed]
- Serhan, C.N.; Hong, S.; Gronert, K.; Colgan, S.P.; Devchand, P.R.; Mirick, G.; Moussignac, R.L. Resolvins. J. Exp. Med. 2002, 196, 1025–1037. [Google Scholar] [CrossRef] [PubMed]
- Savill, J.; Haslett, C. Granulocyte clearance by apoptosis in the resolution ofinflammation. Semin. Cell Biol. 1995, 6, 385–393. [Google Scholar] [CrossRef]
- Savill, J.; Dransfield, I.; Hogg, N.; Haslett, C. Vitronectin receptor-mediated phagocytosis of cells undergoing apoptosis. Nature 1990, 343, 170–173. [Google Scholar] [CrossRef] [PubMed]
- Simon, H.U. Neutrophil apoptosis pathways and their modifications in inflammation. Immunol. Rev. 2003, 193, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.R.; Loison, F. Constitutive neutrophil apoptosis: Mechanisms and regulation. Am. J. Hematol. 2008, 83, 288–295. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.; Whyte, M.K.; Haslett, C. Inhibition of apoptosis and prolongation of neutrophil functional longevity by inflammatory mediators. J. Leukoc. Biol. 1993, 54, 283–288. [Google Scholar] [PubMed]
- Colotta, F.; Re, F.; Polentarutti, N.; Sozzani, S.; Mantovani, A. Modulation of granulocyte survival and programmed cell death by cytokines and bacterial products. Blood 1992, 80, 2012–2020. [Google Scholar] [PubMed]
- Renshaw, S.A.; Parmar, J.S.; Singleton, V.; Rowe, S.J.; Dockrell, D.H.; Dower, S.K.; Bingle, C.D.; Chilvers, E.R.; Whyte, M.K.B. Acceleration of human neutrophil apoptosis by TRAIL. J. Immunol. 2003, 170, 1027–1033. [Google Scholar] [CrossRef] [PubMed]
- Kebir, D.; El Filep, J.G. Role of Neutrophil Apoptosis in the Resolution of Inflammation. Sci. World J. 2010, 10, 1731–1748. [Google Scholar] [CrossRef] [PubMed]
- Fox, S.; Leitch, A.E.; Duffin, R.; Haslett, C.; Rossi, A.G. Neutrophil apoptosis: Relevance to the innate immune response and inflammatory disease. J. Innate Immun. 2010, 2, 216–227. [Google Scholar] [CrossRef] [PubMed]
- Daigle, I.; Yousefi, S.; Colonna, M.; Green, D.R.; Simon, H.U. Death receptors bind SHP-1 and block cytokine-induced anti-apoptotic signaling in neutrophils. Nat. Med. 2002, 8, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, S.; Thomas, G.; Harvey, K.; Cottell, D.; Reville, K.; Berlasconi, G.; Petasis, N.A.; Erwig, L.; Rees, A.J.; Savill, J.; et al. Lipoxins, aspirin-triggered epi-lipoxins, lipoxin stable analogues, and the resolution of inflammation: Stimulation of macrophage phagocytosis of apoptotic neutrophils in vivo. J. Am. Soc. Nephrol. 2002, 13, 2497–2507. [Google Scholar] [CrossRef] [PubMed]
- Fadok, V.A.; Bratton, D.L.; Konowal, A.; Freed, P.W.; Westcott, J.Y.; Henson, P.M. Macrophages that have ingested apoptotic cells in vitro inhibit proinflammatory cytokine production through autocrine/paracrine mechanisms involving TGF-beta, PGE2, and PAF. J. Clin. Investig. 1998, 101, 890–898. [Google Scholar] [CrossRef] [PubMed]
- Maderna, P.; Godson, C. Phagocytosis of apoptotic cells and the resolution of inflammation. Biochim. Biophys. Acta 2003, 1639, 141–151. [Google Scholar] [CrossRef] [PubMed]
- Voll, R.E.; Herrmann, M.; Roth, E.A.; Stach, C.; Kalden, J.R.; Girkontaite, I. Immunosuppressive effects of apoptotic cells. Nature 1997, 390, 350–351. [Google Scholar] [CrossRef] [PubMed]
- Stark, M.A.; Huo, Y.; Burcin, T.L.; Morris, M.A.; Olson, T.S.; Ley, K. Phagocytosis of apoptotic neutrophils regulates granulopoiesis via IL-23 and IL-17. Immunity 2005, 22, 285–294. [Google Scholar] [CrossRef] [PubMed]
- Serinkan, B.F.; Gambelli, F.; Potapovich, A.I.; Babu, H.; Giuseppe, M.D.; Ortiz, L.A.; Fabisiak, J.P.; Kagan, V.E. Apoptotic cells quench reactive oxygen and nitrogen species and modulate TNF-α/TGF-β1 balance in activated macrophages: Involvement of phosphatidylserine-dependent and -independent pathways. Cell Death Differ. 2005, 12, 1141–1144. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Havixbeck, J.J.; Barreda, D.R. Neutrophil Development, Migration, and Function in Teleost Fish. Biology 2015, 4, 715-734. https://doi.org/10.3390/biology4040715
Havixbeck JJ, Barreda DR. Neutrophil Development, Migration, and Function in Teleost Fish. Biology. 2015; 4(4):715-734. https://doi.org/10.3390/biology4040715
Chicago/Turabian StyleHavixbeck, Jeffrey J., and Daniel R. Barreda. 2015. "Neutrophil Development, Migration, and Function in Teleost Fish" Biology 4, no. 4: 715-734. https://doi.org/10.3390/biology4040715