Parasitism of the Invasive Brown Marmorated Stink Bug, Halyomorpha halys (Hemiptera: Pentatomidae), by the Native Parasitoid, Trichopoda pennipes (Diptera: Tachinidae)


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Field Collection
2.2. Oviposition Patterns
2.3. Data Analysis
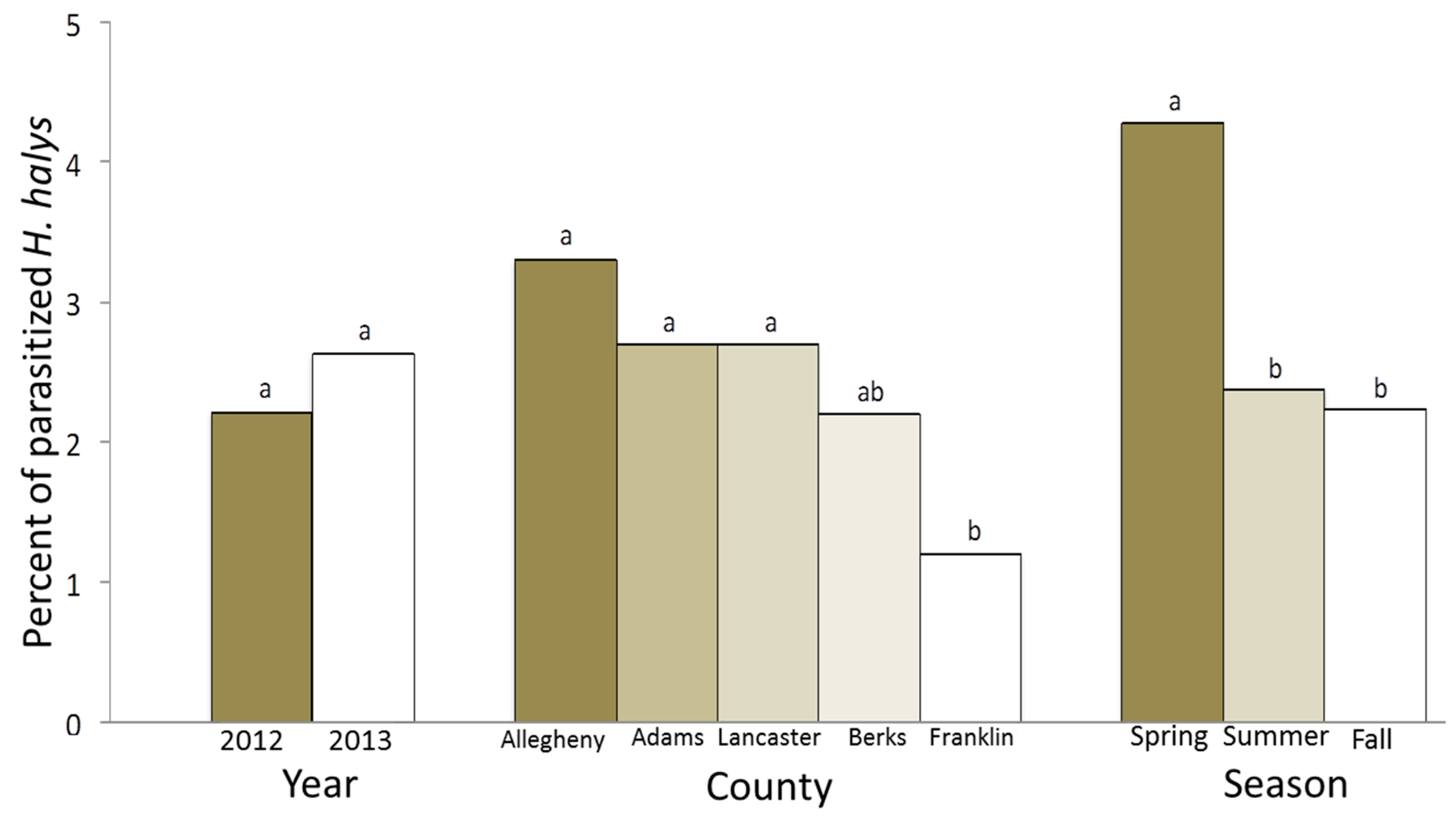
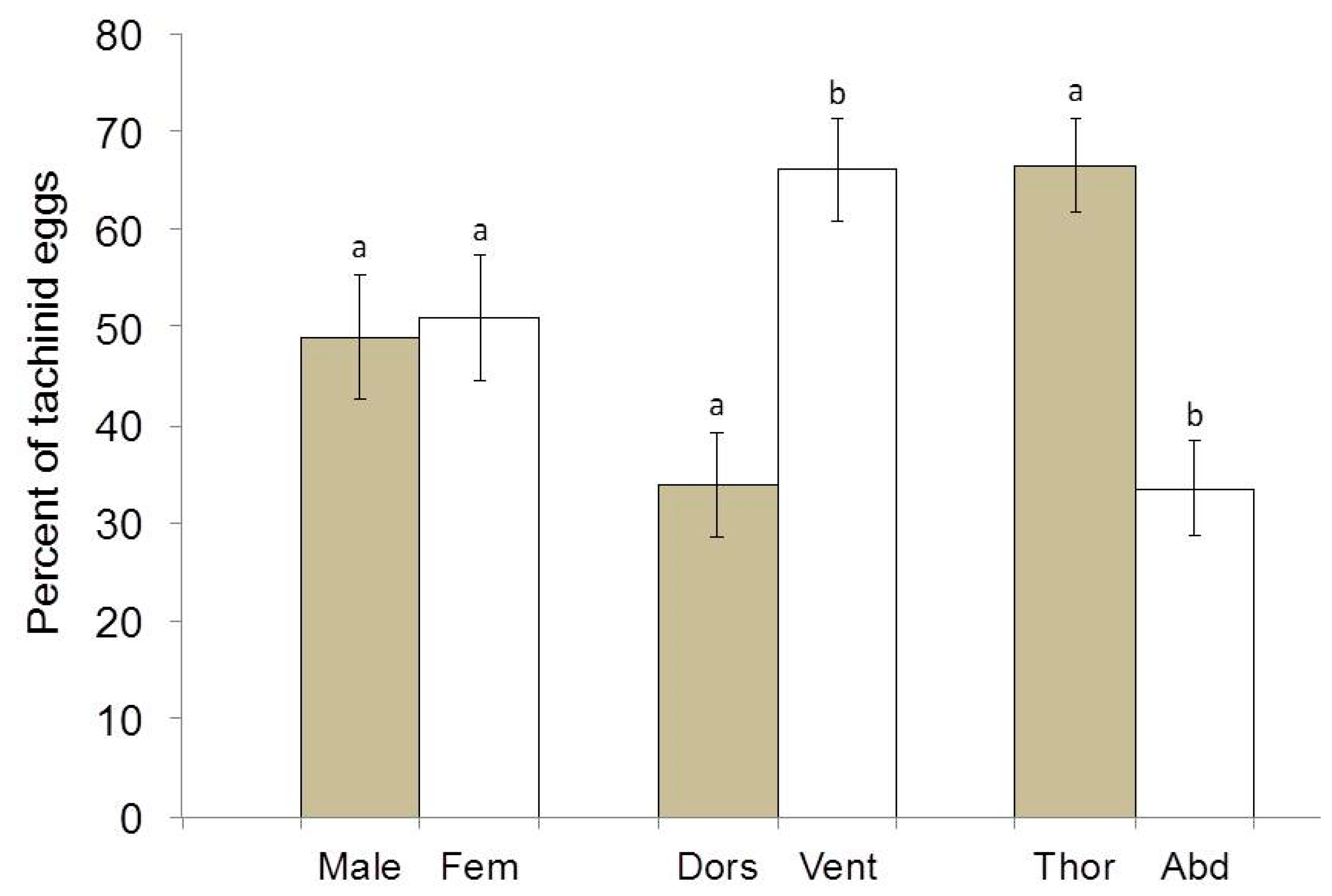
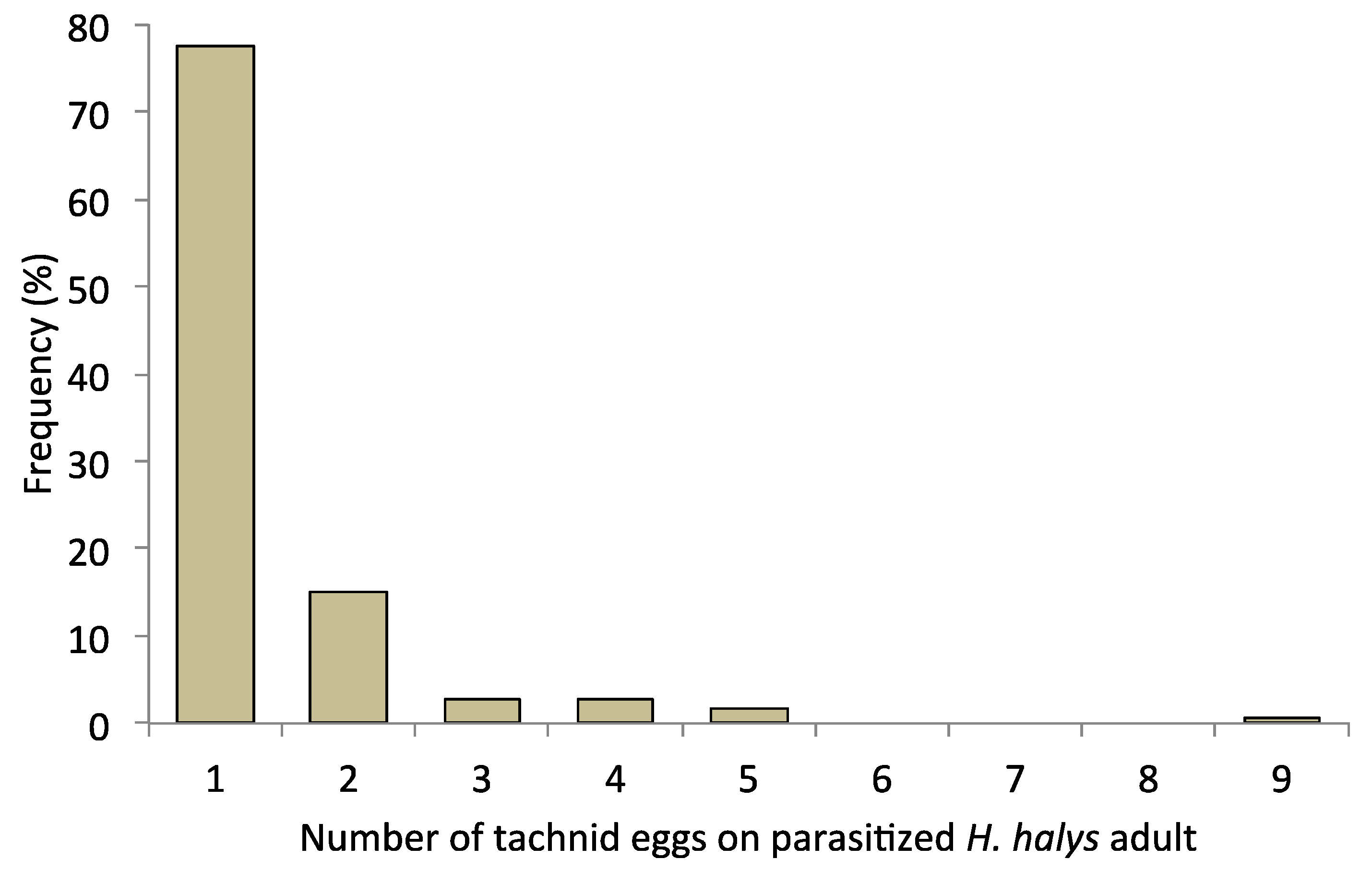
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hoebeke, E.R.; Carter, M.E. Halyomorpha halys (Stahl) (Heteroptera: Pentatomidae): A polyphagous plant pest from Asia newly detected in North America. Proc. Entomol. Soc. Wash. 2003, 105, 225–237. [Google Scholar]
- Haye, T.; Gariepy, T.; Hoelmer, K.; Rossi, J.-P.; Streito, J.-C.; Tassus, X.; Desneux, N. Range expansion of the invasive brown marmorated stinkbug, Halyomorpha halys: An increasing threat to field, fruit and vegetable crops worldwide. J. Pest Sci. 2015, 88, 665–673. [Google Scholar] [CrossRef]
- Leskey, T.C.; Nielsen, A.L. Impact of the invasive brown marmorated stink bug in North American and Europe: History, biology, ecology, and management. Annu. Rev. Entomol. 2018, 63, 599–618. [Google Scholar] [CrossRef] [PubMed]
- Kuhar, T.P.; Kamminga, K.L.; Whalen, J.; Dively, G.P.; Brust, G.; Hooks, C.R.R.; Hamilton, G.; Herbert, D.A. The Pest Potential of Brown Marmorated Stink Bug on Vegetable Crops. Plant Heal. Prog. 2012, 13, 41. [Google Scholar] [CrossRef]
- Nielsen, A.L.; Hamilton, G.C. Seasonal occurrence and impact of Halyomorpha halys (Hemiptera: Pentatomidae) in tree fruit. J. Econ. Entomol. 2009, 102, 1133–1140. [Google Scholar] [CrossRef] [PubMed]
- Leskey, T.C.; Short, B.D.; Butler, B.R.; Wright, S.E. Impact of the invasive brown marmorated stink bug, Halymorpha halys (Stal), in mid-Atlantic tree fruit orchards in the United States: Case studies of commercial management. Psyche A J. Entomol. 2012, 535062, 2012. [Google Scholar] [CrossRef]
- Rice, K.B.; Berg, C.J.; Bergmann, E.J.; Biddinger, D.J.; Dieckhoff, C.; Dively, G.; Fraser, H.; Gariepy, T.; Hamilton, G.; Hayes, T.; et al. Biology, ecology, and management of brown marmorated stink bug (Hemiptera: Pentatomidae). J. Integr. Pest Manag. 2014, 5, A1–A13. [Google Scholar] [CrossRef]
- Bakken, A.J.; Schoof, S.C.; Bickerton, M.; Kamminga, K.L.; Jenrette, J.C.; Malone, S.; Abney, M.A.; Herbert, D.A.; Reisig, D.; Kuhar, T.P.; et al. Occurrence of Brown Marmorated Stink Bug (Hemiptera: Pentatomidae) on Wild Hosts in Nonmanaged Woodlands and Soybean Fields in North Carolina and Virginia. Environ. Entomol. 2015, 44, 1011–1021. [Google Scholar] [CrossRef] [Green Version]
- Colautti, R.I.; Ricciardi, A.; Grigorovich, I.A.; MacIsaac, H.J. Is invasion success explained by the enemy release hypothesis? Ecol. Lett. 2004, 7, 721–733. [Google Scholar] [CrossRef]
- Lockwood, J.A. The ethics of biological control: Understanding the moral implications of our most powerful ecological technology. Agric. Hum. Values 1996, 13, 2–19. [Google Scholar] [CrossRef]
- Lockwood, J.A. The ethics of “classical” biological control and the value of place. In Balancing Nature: Assessing the Impact of Importing Non-Native Biological Control Agents (An International Perspective); Lockwood, J.A., Howarth, F.G., Purcell, M.F., Eds.; Thomas Say Publications in Entomology, Entomological Society of America: Lanham, MD, USA, 2001; pp. 100–119. [Google Scholar]
- Lockwood, J.A.; Purcell, M.F.; Howarth, F.G. The value of place and the place of values. In Balancing Nature: Assessing the Impact of Importing Non-Native Biological Control Agents (An International Perspective); Lockwood, J.A., Howarth, F.G., Purcell, M.F., Eds.; Thomas Say Publications in Entomology, Entomological Society of America: Lanham, MD, USA, 2001; pp. 1–2. [Google Scholar]
- Howarth, F.G. Environmental issues concerning the importation of nonindigenous biological control agents. In Balancing Nature: Assessing the Impact of Importing Non-Native Biological Control Agents (An International Perspective); Lockwood, J.A., Howarth, F.G., Purcell, M.F., Eds.; Thomas Say Publications in Entomology, Entomological Society of America: Lanham, MD, USA, 2001; pp. 70–99. [Google Scholar]
- Carlson, N.O.; Sarnelle, O.; Strayer, D.L. Native predators and exotic prey—An acquired taste? Front. Ecol. Environ. 2009, 7, 525–532. [Google Scholar] [CrossRef]
- Messing, R.H.; Purcell, M.F. Regulatory constraints to the practice of biological control in Hawaii. In Balancing Nature: Assessing the Impact of Importing Non-Native Biological Control Agents (An International Perspective); Lockwood, J.A., Howarth, F.G., Purcell, M.F., Eds.; Thomas Say Publications in Entomology, Entomological Society of America: Lanham, MD, USA, 2001; pp. 3–14. [Google Scholar]
- Allen, H.W. Parasites of the Oriental Fruit Moth in the Eastern United States; Agricultural Research Service, US Department of Agriculture: Washington, DC, USA, 1962; p. 1265.
- Stacconi, M.V.R.; Grassi, A.; Dalton, D.T.; Miller, B.; Ouantar, M.; Loni, A.; Ioriatti, C.; Walton, V.M.; Anfora, G. First field records of Pachycrepoideus vindemiae as a parasitoid of Drosphila suzukii in European and Oregon small fruit production areas. Entomologia 2013, 1, 11–16. [Google Scholar]
- Biddinger, D.; Butler, B.; Joshi, N.K. Biological control of mites in Pennsylvania and Maryland apple orchards. Pa. Fruit News 2014, 91, 41–51. [Google Scholar]
- Abram, P.K.; Hoelmer, K.A.; Acebes-Doria, A.; Andrews, H.; Beers, E.H.; Bergh, J.C.; Bessin, R.; Biddinger, D.; Botch, P.; Buffington, M.L.; et al. Indigenous arthropod natural enemies of the invasive brown marmorated stink bug in North America and Europe. J. Pest Sci. 2017, 90, 1009–1020. [Google Scholar] [CrossRef]
- Aldrich, J.R.; Khrimian, A.; Zhang, A.; Shearer, P.W. Bug pheromones (Hemiptera: Heteroptera) and tachinid fly host-finding. Denisia 2006, 19, 1015–1031. [Google Scholar]
- Pickett, C.H.; Schoenig, S.E.; Hoffman, M.P. Establishment of the squash bug parasitoid Trichopoda pennipes Fabr. (Diptera: Tachinidae), in northern California. Pan-Pac. Entomol. 1996, 72, 220–226. [Google Scholar]
- Shahjahan, M.; Beardsley, J.W. Egg viability and larval penetration in Trichopoda pennipes pilipes Fabricius (Diptera: Tachinidae). Proc. Hawaii. Entomol. Soc. 1975, 21, 133–136. [Google Scholar]
- Todd, J.W.; Lewis, W.J. Incidence and oviposition patterns of Trichopoda pennipes (F), a parasite of the southern green stink bug, Nezara viridula (L.). J. Ga. Entomol. Soc. 1976, 11, 50–54. [Google Scholar]
- Worthley, H.N. The biology of Trichopoda pennipes Fab. (Diptera, Tachinidae), a parasite of the common squash bug. Psyche A J. Entomol. 1924, 31, 7–16. [Google Scholar] [CrossRef]
- Herlihy, M.V.; Talamas, E.J.; Weber, D.C. Attack and Success of Native and Exotic Parasitoids on Eggs of Halyomorpha halys in Three Maryland Habitats. PLoS ONE 2016, 11, e0150275. [Google Scholar] [CrossRef]
- Moraglio, S.T.; Tortorici, F.; Pansa, M.G.; Castelli, G.; Pontini, M.; Scovero, S.; Visentin, S.; Tavella, L. A 3-year survey on parasitism of Halyomorpha halys by egg parasitoids in northern Italy. J. Pest Sci. 2019. [Google Scholar] [CrossRef]
- Morrison, W.R., III; Mathews, C.R.; Leskey, T.C. Frequency, efficiency, and physical characteristics of predation by generalist predators of brown marmorated stink bug (Hemiptera: Pentatomidae) eggs. Biol Control 2016, 97, 120–130. [Google Scholar] [CrossRef] [Green Version]
- Biddinger, D.J.; Joshi, N.K. First report of native Astata unicolor (Hymenoptera: Crabronide) predation on the nymphs and adults of the invasive brown marmorated stink bug, Halyomorpha halys (Hemiptera: Pentatomidae). Fla. Entomol. 2017, 100, 809–812. [Google Scholar] [CrossRef]
- Biddinger, D.; Surcica, A.; Joshi, N.K. A native predator utilizing the invasive brown marmorated stink bug, Halyomorpha halys (Hemiptera: Pentatomidae) as a food source. Biocontrol Sci. Technol. 2017, 27, 903–907. [Google Scholar] [CrossRef]
- Morrison, W.R., III; Bryant, A.N.; Poling, B.; Quinn, N.F.; Leskey, T.C. Predation of Halyomorpha halys (Hemiptera: Pentatomidae) from web-building spiders associated with anthropogenic dwellings. J. Insect Behav. 2017, 30, 70–85. [Google Scholar] [CrossRef]
- Arnaud, J. Host-Parasite Catalog of North American Tachnidae (Diptera); United States Department of Agriculture, Science and Education Administration: Washington, DC, USA, 1978.
- Harris, V.E.; Todd, J.W. longevity and reproduction of the southern green stink bug, nezara viridula, as affected by parasitization by trichopoda pennipes. Entomol. Exp. Et Appl. 1982, 31, 409–412. [Google Scholar] [CrossRef]
- De Salles, L.A.B. Effect of Trichopoda pennipes parasitization on Nezara viridula. Pesq. Agropec. Bras. 1992, 27, 981–986. [Google Scholar]
- Decker, K.B.; Yeargan, K.V. Seasonal phenology and natural enemies of the squash bug (Hemiptera: Coreidae) in Kentucky. Environ. Entomol. 2014, 37, 670–678. [Google Scholar] [CrossRef]
- Booth, C.L. Biology of Largus californicus (Hemiptera: Largidae). Southwest. Nat. 1990, 35, 15. [Google Scholar] [CrossRef]
- Biddinger, D.J.; Leslie, T.W.; Joshi, N.K. Reduced-risk pest management programs for eastern U.S. peach orchards: Effects on arthropod predators, parasitoids, and select pests. J. Econ. Entomol. 2014, 107, 1084–1091. [Google Scholar] [CrossRef]
- Beard, R.L. The biology of Anasa tristis DeGeer with particular reference to the tachinid parasite, Trichopoda pennipes Fabr. Bull. Conn. Agric. Exp. Stn. 1940, 440, 595–680. [Google Scholar]
- Blaauw, B.R.; Polk, D.; Nielsen, A.L. IPM-CPR for peaches: Incorporating behaviorally-based methods to manage Halyomporpha halys and key pests in peach. Pest Manag. Sci. 2015, 71, 1513–1522. [Google Scholar] [CrossRef] [PubMed]
- Tooker, J.F.; Hauser, M.; Hanks, L.M. Floral Host Plants of Syrphidae and Tachinidae (Diptera) of Central Illinois. Ann. Entomol. Soc. Am. 2006, 99, 96–112. [Google Scholar] [CrossRef]
- Wratten, S.D.; Landis, D.A.; Gurr, G.M. Habitat Management to Conserve Natural Enemies of Arthropod Pests in Agriculture. Annu. Rev. Entomol. 2000, 45, 175–201. [Google Scholar]
- Langellotto, G.A.; Denno, R.F. Responses of invertebrate natural enemies to complex-structured habitats: A meta-analytical synthesis. Oecologia 2004, 139, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, F.; Booij, C.; Tscharntke, T. Sustainable pest regulation in agricultural landscapes: A review on landscape composition, biodiversity and natural pest control. Proc. R. Soc. B Boil. Sci. 2006, 273, 1715–1727. [Google Scholar] [CrossRef] [PubMed]
- Heimpel, G.E.; Jervis, M.A. Does floral nectar improve biological control by parasitoids. In Plant-Provided Food for Carnivorous Insects; Wackers, F.L., van Rijn, P.C.J., Bruin, J., Eds.; Cambridge University Press: Cambridge, MA, USA, 2005; pp. 267–304. [Google Scholar]
- Biddinger, D.; Rajotte, E.G.; Joshi, N.K. Integrating pollinator health into tree fruit IPM—A case study of Pennsylvania apple production. In The Pollination of Cultivated Plants: A Compendium for Practitioners, 2nd ed.; FAO: Rome, Italy, 2018; Volume 1, pp. 69–83. [Google Scholar]
- Harris, V.E.; Todd, J.W. Male mediated aggregation of male, female and 5th instar southern green stink bugs and concomitant attraction of a tachinid parasite, Trichopoda pennipes. Entomol. Exp. Appl. 1980, 27, 117–126. [Google Scholar] [CrossRef]
- Mitchell, W.C.; Mau, R.F.L. Response of the female southern green stink bug and its parasite, Trichopoda pennipes, to male stink bug pheromones. J. Econ. Entomol. 1971, 64, 856–859. [Google Scholar] [CrossRef]
- Aldrich, J.R. Chemistry and biological activity of pentatomid sex pheromones. In Biologically Active Natural Products: Potential Use in Agriculture; Cutler, H.G., Ed.; American Chemical Society: Washington, DC, USA, 1988; pp. 417–431. [Google Scholar]



© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Joshi, N.K.; Leslie, T.W.; Biddinger, D.J. Parasitism of the Invasive Brown Marmorated Stink Bug, Halyomorpha halys (Hemiptera: Pentatomidae), by the Native Parasitoid, Trichopoda pennipes (Diptera: Tachinidae). Biology 2019, 8, 66. https://doi.org/10.3390/biology8030066
Joshi NK, Leslie TW, Biddinger DJ. Parasitism of the Invasive Brown Marmorated Stink Bug, Halyomorpha halys (Hemiptera: Pentatomidae), by the Native Parasitoid, Trichopoda pennipes (Diptera: Tachinidae). Biology. 2019; 8(3):66. https://doi.org/10.3390/biology8030066
Chicago/Turabian StyleJoshi, Neelendra K., Timothy W. Leslie, and David J. Biddinger. 2019. "Parasitism of the Invasive Brown Marmorated Stink Bug, Halyomorpha halys (Hemiptera: Pentatomidae), by the Native Parasitoid, Trichopoda pennipes (Diptera: Tachinidae)" Biology 8, no. 3: 66. https://doi.org/10.3390/biology8030066