1. Introduction
Recent studies have shown strong antioxidant activity of astaxanthin [
1] with benefits to the immune system, cardiac muscles, and to treatments of cancer and skin aging [
1,
2,
3,
4]. The green alga
Haematococcus pluvialis is the most widely used source for natural astaxanthin at a commercial scale and has been reported to accumulate astaxanthin up to 5.9% (w/w) of the dry biomass [
5,
6,
7].
Due to its life history,
H. pluvialis is usually grown in two-phase cultivation, in which the cells are grown by cell multiplication in the “green phase” and are stimulated to accumulate astaxanthin in the successive “red phase” (e.g., [
6,
8]). The two-phase cultivation method is technically demanding and is characterized by high energy consumption and relatively long cultivation times [
9,
10].
Two-phase suspended cultivation of
H. pluvialis is the predominant cultivation system at both exploratory and commercial scales. In suspended cultivation, a maximum light intensity of 150 µmol photons m
−2 s
−1 should not be exceeded to maintain cell growth and divisions and environmental parameters such as temperature, CO
2, and pH need to be closely monitored [
6,
8]. As the needed biomass is reached, the second phase, known as the stressed or red phase is switched on to stimulate astaxanthin accumulation [
6,
8]. Suspended cultivation of
Haematococcus is performed in open ponds/raceways or in closed tubular or flat-plate photobioreactors (PBR). Open pond cultivation is usually used only for the stressed phase with short cultivation time (4–6 days) to minimize contamination and apply stress conditions [
8]. The closed PBR can minimize contamination and control growth parameters better, but has the drawbacks of high assembly and maintenance costs (on average, a tenfold increase in costs) [
11,
12,
13]. Suspended cultivation usually yields low biomass density (0.05–0.06% of the total cultivation liquid) due to the ineffective light and carbon dioxide distribution caused by the low surface-to-volume ratio of the cultivation systems and demands additional effort for the harvest of the algae [
14]. Flat or tubular closed systems increase productivity by increasing the efficiency of light usage and limiting contamination, but there are other limitations, especially in the adhesion of cells to the inner PBR surfaces of many microalgae including
H. pluvialis [
15].
Porous substrate photobioreactors (PSBR) such as the vertical Twin-Layer system can be used to cultivate various algal species including
H. pluvialis (for a review, see Podola et al. [
16]). PSBRs are a recently developed technique for microalgal cultivation in immobilized systems that has also shown considerable promise for application in
Haematococcus [
7,
17,
18,
19,
20,
21]. The Twin-Layer system is a modular PSBR. The prototype (large scale) TL-PSBR is capable to operate up to eight Twin-Layer modules, the area for growth of microalgae on both sides is 2 × 0.67 m
2 for each module. A tube-type TL-PSBR for small-scale holding of 12 Twin-Layer modules of 10 × 90 cm was used to monitor growth kinetics of
H. pluvialis [
15,
22,
23]. Each TL module consists of a vertically orientated source layer (e.g., glass fiber nonwoven), through which a flow of medium is established to feed nutrient. Onto both sides of the source layer, the substrate layers (e.g., printing paper or filter paper that has small porosity) are applied by self-adhesion of hydrogen bonds. Microalgae will be immobilized on the substrate layers and will develop a biofilm [
15,
22,
23].
The concurrent production of biomass and astaxanthin of
H. pluvialis in one-phase cultivation at high light intensity with CO
2 supplementation using a vertical Twin-Layer photobioreactor yielded biomass productivities of up to 19.4 g m
−2 d
−1 and final biomass of 213 g dry weight m
−2 growth area after 16 days of cultivation [
23]. In comparison with two-phase cultivation using the same photobioreactor system, one-phase cultivation yielded a similar astaxanthin output in just half of the cultivation time [
23]. In one-phase cultivation on TL-PSBR, growth and astaxanthin accumulation of
H. pluvialis happens simultaneously but in different cell layers of the biofilm. Cells on the surface are exposed to direct light and change into red akinetes, whereas green cells in the lower layers divide by spore formation [
23]. Hence, the thickness of the biofilm and high light intensity play an important role in one-phase cultivation. However, in modular vertical TL-PSBRs high intensity and uniformity of incident light is difficult to achieve due to mutual shading by adjacent modules, although the vertical arrangement of the TL modules saves space and enhances areal productivity.
A horizontal (low-angled) TL-PSBR was developed for research purposes in Vietnam starting in 2016, as it is easier to construct and operate than a vertical TL system and for high light applications such as the production of astaxanthin, only slightly less productive (aerial productivity) than a vertical system. Furthermore, harvesting the biomass is technically less complicated in low-angled TL-systems compared to vertical systems [
16]. In Vietnam, the angled Twin-Layer photobioreactor system for immobilized cultivation of microalgae provides significant advantages over traditional suspended cultivation such as saving of water, energy and cultivation time [
24]. This study investigates several factors affecting biomass and astaxanthin accumulation of
H. pluvialis cultivated in a small-scale angled TL-PSBR: initial biomass (inoculum) density, algae storage time before immobilization and influence of different light intensities. Furthermore, a larger-scale angled TL-PSBR system for
H. pluvialis was designed and operated to probe upscaling astaxanthin production to pilot scale.
2. Materials and Methods
2.1. Algal Strain and Culture Maintenance
The
H. pluvialis strain CCAC 0125 was supplied from the Culture Collection of Algae at the University of Cologne, Germany (
http://www.ccac.uni-koeln.de/). The strain was maintained in 100 mL Erlenmeyer flasks, with 50 mL of modified BG-11 medium [
23] and transferred every 3–4 weeks. Cultures were grown at 20 °C, with a light intensity of 30 µmol photons m
−2 s
−1 provided by fluorescent lamps (Philips Essential energy saver 18W E27 Cool day light lamp, Philips Lighting GmbH, Hamburg, Germany), using a 14/10 h light/dark cycle.
2.2. Experimental Designs
The experiments were carried out on small scale (0.1 × 0.5 = 0.05 m
2 growth area) and large scale (0.5 m
2 × 4 growth area) angled TL-PSBRs [
24]. The non-sterile culture medium was stored in 40 L plastic chambers.
2.2.1. Experiments with the Small-Scale TL-PSBR
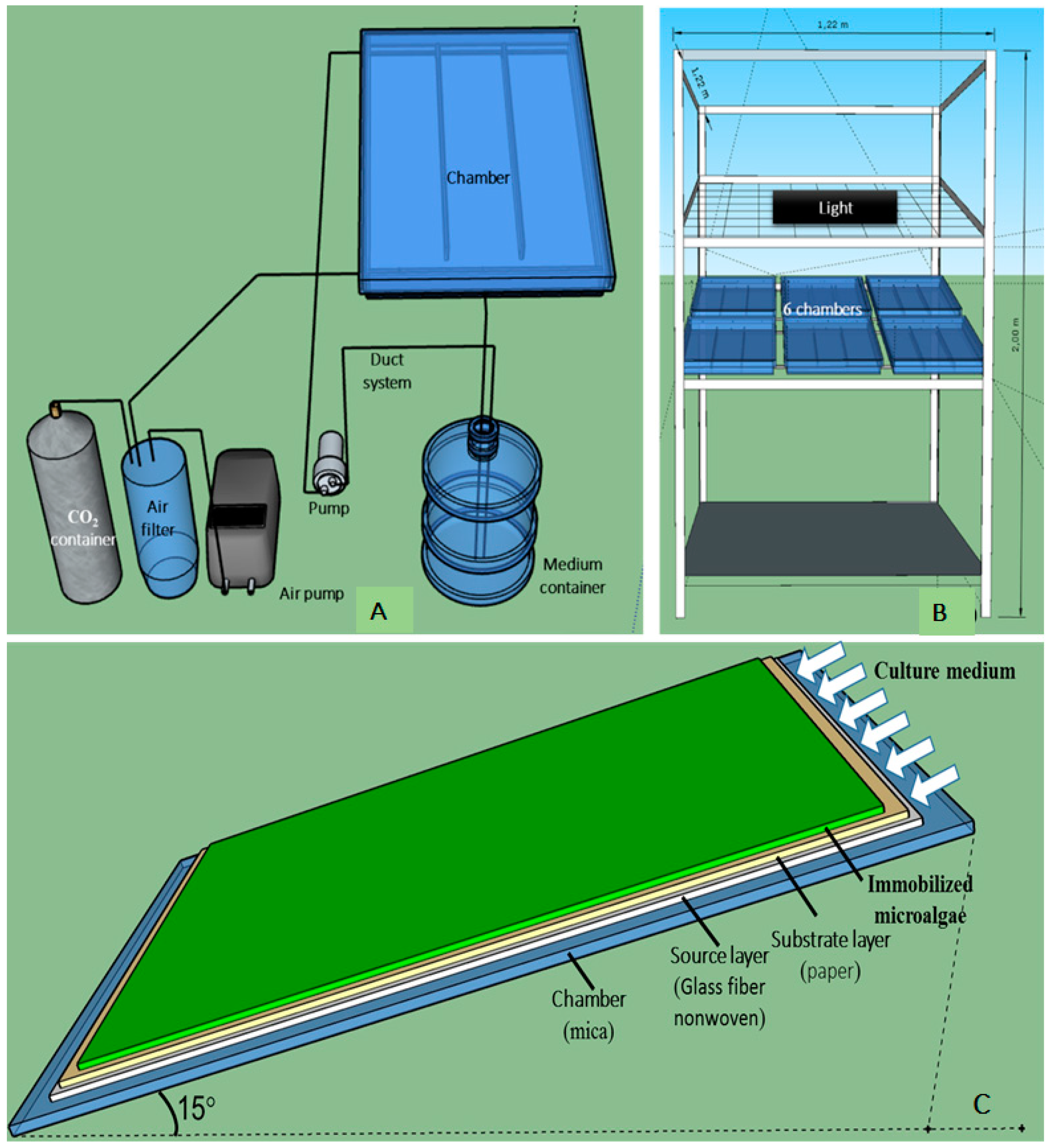
The small-scale angled TL-PSBR for
H. pluvialis immobilized cultivation includes small chambers with source layers, substrate layer (paper) and immobilized algae inside, nutrient circulation system, air circulation system, light supply system and steel frame (
Figure 1).
Each chamber is 0.37 × 0.52 m in size divided into three compartments. Each compartment is about 0.12 × 0.52 m in size and contains three layers: the source layer (nonwoven fiberglass, 0.11 × 0.51 m), substrate layers (Kraft paper 70 g m
−2, 0.11 × 0.51 m) and biofilm (immobilized algae on the substrate layer, the growth area, 0.1 × 0.5 m) (
Figure 1C). A small-scale TL-PSBR has six small chambers (
Figure 1B) that share the same nutrient, air and light supply systems. Therefore, a small system with 18 growth areas (0.05 m
2 for each) for microalgae was divided into different treatments for each experiment.
Three experiments with the small-scale TL-PSBR were carried out. The most effective outcome from each preceding experiment would be applied for the next experiment.
Influence of algal storage time before immobilization: the concentrated algae were stored at 4 °C in the dark over 24, 72, 120 and 168 h after centrifugation, respectively. Initial algae biomass for inoculation of the TL-PSBR was 5 g dry weight m−2, the light intensity was adjusted to reach the range of 400–600 µmol photons m−2 s−1 over the growth area.
The optimal initial algal density: the initial biomass densities of 2.5, 5, 7.5 and 10 g dry weight m−2 were tested. Algae were immobilized immediately after centrifugation or were stored for 24 h at 4 °C. The light intensity was set at 400–600 µmol photons m−2 s–1.
Influence of light intensity on algal growth and astaxanthin accumulation: The experiment used sodium high-pressure lamps (PHILIPS) as a light source with the intensity of 300–1000 µmol photon m−2 s−1. Each point on the cultivation chambers received different light intensity and was considered to be one experiment for one value point of this parameter. The initial biomass density was 7.5 g dry weight m−2.
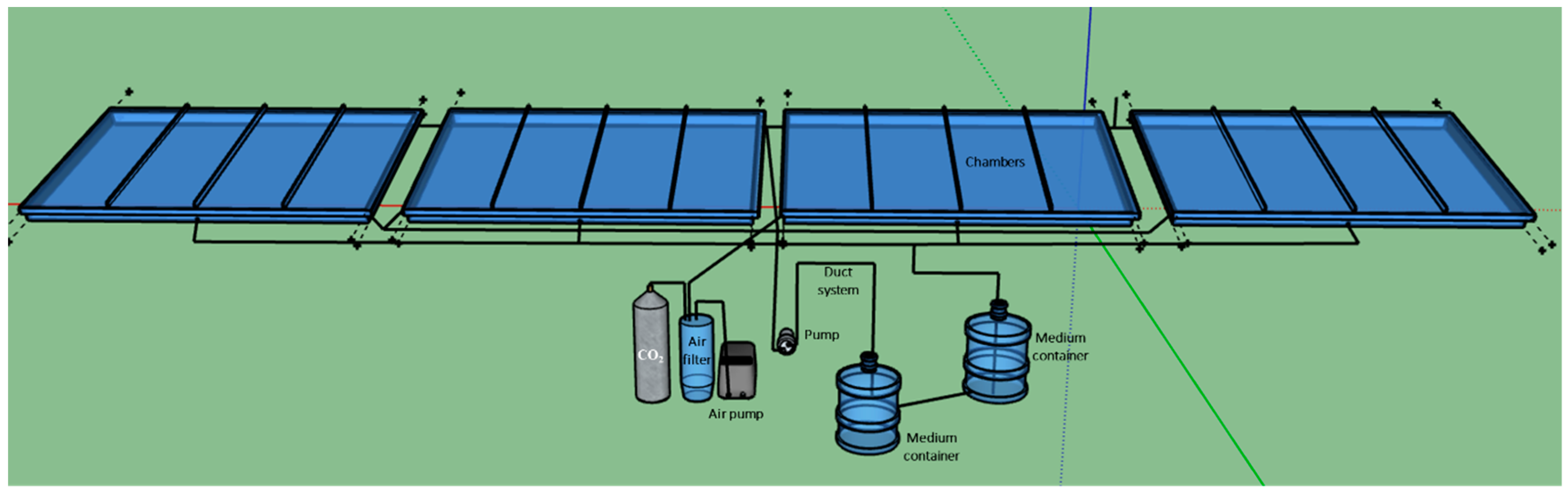
2.2.2. Experiment with the Large Scale TL-PSBR
The large-scale angled TL-PSBR uses the same components of the small system but has four larger chambers assembled in the same nutrient, air and light supply systems, each chamber provides 1 × 0.5 = 0.5 m
2 area for algae growth (
Figure 2). Thus, each large-scale TL-PSBR will have 0.5 × 4 = 2 m
2 growth area. Two large TL-PSBRs with two different lighting systems were set up and operated at the same time to perform the experiment.
In large-scale TL-PSBRs, the experiment was set up to compare the efficiency for lighting of two different luminaires. The two luminaires used to compare energy efficiency in this experiment are: luminair 250W (light intensity of 300–1150 µM photons m−2 s−1, provided by ten PHILIPS 250W sodium high-pressure lamps) and luminair 400W (light intensity of 300–1300 µmol photon m−2 s−1, provided by eight PHILIPS 400W sodium high-pressure lamps).
Some experimental conditions were established based on the results obtained with the small TL-PSBR, in which the initial dry biomass density was 7.5 g m−2, and algae were immobilized after centrifugation within 24 h.
2.3. Preparation of H. pluvialis Biomass for Experiments on Angled TL-PSBRs
Suspended cultivation before immobilization: The culture medium used was modified BG-11 [
23]. Suspended cultivation of microalgae was done in 500 mL (culture time is about 10 days) and 2000 mL (culture time is about 14 days) Erlenmeyer flasks at 23 ± 2 °C, illuminated by a fluorescent lamp at 40–60 µmol photons m
−2 s
−1, using a 14/10 h light/dark cycle. However, the large-scale angled TL-PSBRs required a larger amount of initial algal biomass, so the volume of suspended algae was increased to 10 L in PE bags (with 6 L of modified BG-11 medium) (
Figure 3). Algae from 2000 mL flasks will be transplanted through PE bags with the initial algal density of about 1.5 x 10
4 cells mL
−1, and a culture period of about 16 days. The medium was aerated with ambient air during the cultivation period, illuminated by fluorescent lamps at 40–60 µmol photons m
−2 s
−1 with 14/10 h light/dark cycle, at 23 ± 2 °C.
Centrifugation to concentrate algae in suspension: Algae in 10 L PE bags after 20 days of culture consisted to 85% of flagellate cells and were harvested from suspension by low-speed centrifugation (800 × g for 5 min) with a large bench centrifuge (ROTANTA 460 RC, Hettich GmbHo. KG, Tuttlingen, Germany). Under these conditions, no damage was observed to the flagellate cells. After centrifugation, the concentrated algal suspension was used to determine the amount of dry biomass (mL).
The dry biomass of the concentrated algal suspension was determined by filtering 1 mL of the suspension onto a filter paper (0.4 µm pore size, Whatman GmbH, Dassel, Germany) with a predetermined dry weight of W1 and dried the whole set in 2 h at 105 °C. The dried filter paper with algae was weighed, and the drying process repeated until an unchanged total weight of W2 was achieved. The dry biomass in 1 mL concentrated algae liquid was calculated as D = W2 – W1.
2.4. Algae Immobilization on Angled TL-PSBRs
The inoculation of algae on the substrate layer was tested with several methods. However, using a brush to apply the concentrated algal suspension x showed many advantages. The required volume V (mL) of algal suspension immobilized onto a surface area S was calculated as V (mL) = (M × S)/D with M as the initial immobilized dry biomass density (g m−2). The concentrated algal suspension was immobilized by a soft brush onto substrate layer on 0.05 and 0.5 m2 TL-PSBRs, respectively.
2.5. Culture Conditions after Algal Immobilization
Immobilized cultivation time was 10 days. A nutrient-replete BG-11 medium was used during days 1–7 by replacing 20 L (on small-scale TL-PSBR) or 40 L (on large scale TL-PSBR) of medium every 2 or 3 days. Twenty or forty litres of BG-11 medium without N and P (excludes NaNO
3 and K
2HPO
4 × 3H
2O in modified BG-11 medium) [
18] were used on days 8, 9 and 10 for astaxanthin induction. The culture medium was supplemented with CO
2 1% (v/v) as a carbon source and also as a means for pH adjustment. The temperature was kept in 23–26 °C, pH = 6.5–8. The electrical conductivity of the medium was kept in 1800–2000 µS cm
−1 by adding filtered water after every 24 h.
The following variables were monitored: Dry algae biomass productivity (g m−2 d−1) was calculated based on equation Mp = (Ma-M)/10 (Ma: dry biomass after 10 days, M: initial immobilized dry biomass); astaxanthin productivity (mg m−2 d−1) was calculated based on equation Ap = (Ma × 1000 × %A)/10, %A is the astaxathin content in the dry biomass.
Measurement of immobilized dry biomass: after 10 days of cultivation, the thin algal layer and substrate layer (Kraft paper 70 g/m2, Vietnam) were harvested and dried for 2 hours at 105 °C, and cooled in a desiccator in 30 min. The dried product was weighed and the drying process repeated until an unchanged total weight of m2 was obtained. The dry biomass was calculated as Ma = m2 − m1 (g), m1 is the weight of dry paper without algae.
Measurement of astaxanthin content in the dry biomass (%A): Astaxanthin was determined spectrophotometrically according to Li et al. [
25]. All processes were carried out in the dark. A sample of 1 mg of freeze-dried biomass was placed with 0.5 mL acetone 90% in a 2 mL tube with screw cap to limit the vaporization, incubated in water bath at 70 °C for 5 min with intervals for vortexing, and then ground using a glass pestle. The mixture was centrifuged at 4000×
g for 5 min and the supernatant retrieved. Extraction with acetone was repeated until the pellet turned white. The extract was supplied with acetone (90%) to reach a volume of 3 mL in a 5 mL sealed centrifuge tube with screw cap until spectrophotometric analysis.
The OD value of the extracted pigments was measured at a wavelength of 530 nm with Ultrospec 2100 Pro Spectrophotometer (Amersham Biosciences). Astaxanthin content was calculated based on the calibration curve equation of y = 0.0577x + 0.0131 (established with astaxanthin ≥ 98 % (HPLC), Sigma-Aldrich, St. Louis, MO, USA) (
Figure 4), in which y was the OD value measured and x was the astaxanthin content (µg mL
−1). Astaxanthin content in biomass was calculated in percentage (A%), and subsequent estimation of astaxanthin production per biofilm surface (in mg m
−2 d
−1).
2.6. Data Analysis
The data were analyzed using Microsoft Excel 2016. Statistical analysis and resulting charts were made using R program version 3.4.2. The presented values were averaged value of more than three replicates with corresponding standard deviation.
3. Results
3.1. Dry Biomass Growth and Astaxanthin Accumulation of Immobilized H. pluvialis Cultivated in an Angled TL-Photobioreactor at Different Initial Biomass Density
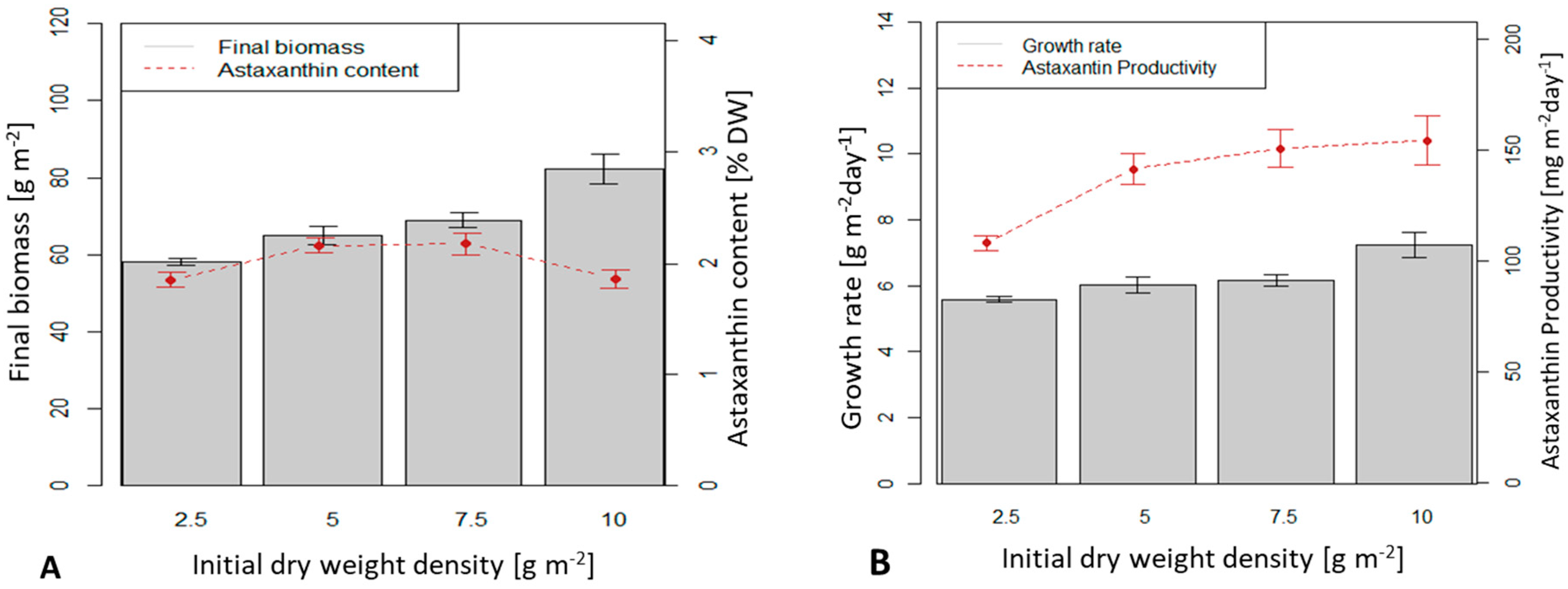
At an initial biomass density of 2.5, 5 and 7.5 g m
−2, the biomass productivities were 5.6, 6 and 6.2 g m
−2 d
−1, respectively (
Figure 5B). These values were not significantly different from each other. The highest average dry biomass productivity reached 7.23 g m
−2 d
−1 at an initial biomass density of 10 g m
−2 and was significantly different from the other three values (
p < 0.05).
Final astaxanthin content in the dry biomass reached the highest value of 2.17 % at initial biomass densities of 5 and 7.5 g m−2, respectively. However, the astaxanthin content at lower (2.5 g m−2) or higher (10 g m−2) initial biomass densities was lower (1.87 %) and the difference was statistically significant (p < 0.05).
Except for low initial biomass densities (2.5 g m−2), astaxanthin productivities at initial biomass densities of 5, 7.5 and 10 g m−2 were similar: 141 (5 g m−2), 151 and 154 mg.m−2 d-−1 (at 7.5 and 10 g m−2).
3.2. Influence of Algal Storage Time after Centrifugation of a Suspension Culture on Biomass Growth and Astaxanthin Accumulation in a TL-PSBR
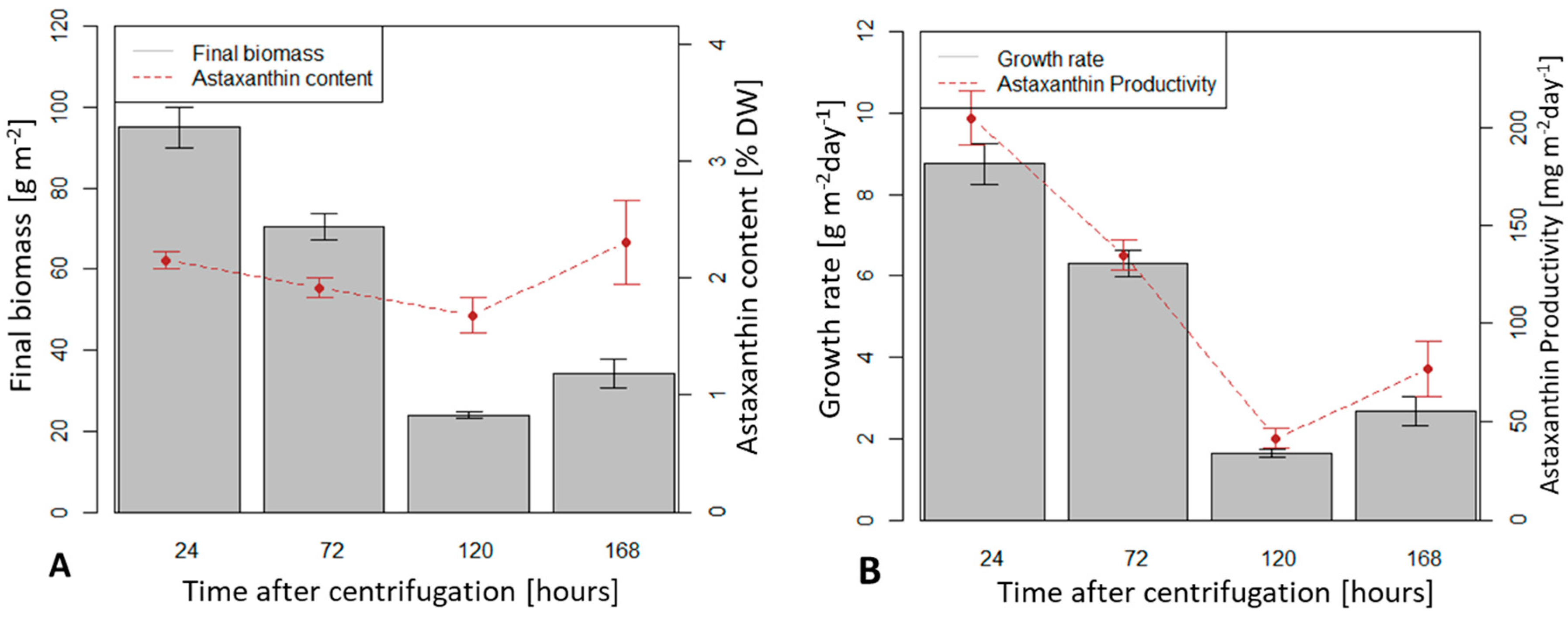
After a storage time of 24 hours before immobilization and at an initial biomass density of 5 g m−2, the highest value of average dry biomass productivity after 10 days of cultivation was achieved (8.75 g m−2 d−1). At an initial biomass density of 5 g m−2, storage times of 72, 120 and 168 h yielded average productivity of 6.3, 1.7 and 2.7 g m−2 d−1, respectively. Biomass productivities at storage time of 24, 72 and 120 h were significantly different (p < 0.05) while productivities at storage times of 120 and 168 h were not (p > 0.05).
Average astaxanthin content reached 2.15 % dry biomass at storage time of 24 h and an initial density of 5 g.m
−2. It decreased with increased time storage until 168 h when it was higher again (1.92, 1.68 and 2.30 % at 72, 120, 168 h storage time, respectively). However, there was no significant difference between these values. Storage time did not influence the final astaxanthin content (% in dry weight) but only the final biomass content (
Figure 6A). Consequently, the storage time of 24 h yielded the highest astaxanthin productivity (205 mg m
−2 d
−1) measured, astaxanthin productivity decreased significantly at longer storage times (
Figure 6B).
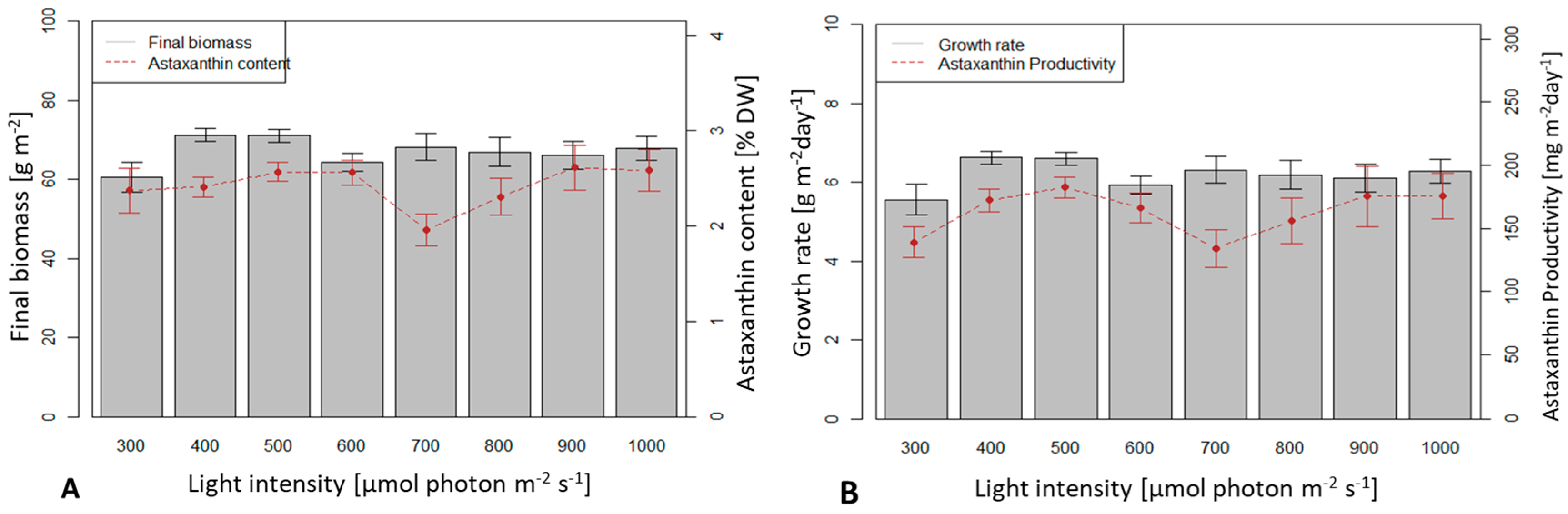
3.3. Influence of Light Intensity on Biomass Growth and Astaxanthin Accumulation of H. pluvialis in an Angled TL-PSBR
The effect of different light intensities was studied in the range of 300–1000 µmol photons m−2 s−1 in 8 experiments.
A light intensity of 400 µmol photons m−2 s−1 yielded the highest average dry biomass productivity (6.6 g m−2 d−1) and 300 µmol photons m−2 s−1 yielded the lowest productivity (5.56 g m−2 d−1). However, the productivity values in the whole range of 300–1000 µmol photons m−2 s−1 were not significantly different (p > 0.05).
Intensities of 500 and 600 µmol photons m
−2 s
−1 yielded highest astaxanthin content (2.56 %) and 700 µM photons m
−2 s
−1 yielded lowest value (1.96 %), the difference, however, was not statistically significant (
p > 0.05) (
Figure 7).
The highest average astaxanthin productivity reached 183 mg m-2 d-1 at 500 µmol photons m−2 s−1 but it was not significantly different from the value at other light intensities.
In general, with a total of 140 h of illumination in 10 days, the dry biomass productivity per each mole of photons decreased with increased light intensity, therefore the highest light intensity only yielded the productivity of 0.12 g m
−2 mol photons
−1. The intensities of 300 and 400 µmol photons m
−2 s
−1 yielded 0.37 and 0.33 g m
−2 mol photons
−1, respectively (
p > 0.05), which was significantly higher than at the other light intensities (
p < 0.05). The light intensity of 500 µmol photons m
−2 s
−1 yielded 0.262 g m
−2 mole photons
−1, higher than the value of 600–1000 µmol photons m
−2 s
−1 (
p < 0.05) (
Figure 8).
Astaxanthin productivity per each mole of photons decreased with increased light intensity. At the intensities of 300, 400 and 500 µmol photons m
−2 s
−1 the productivities were 9.2, 8.5, 7.2 mg m
−2 mole photons
−1, respectively (
p > 0.05) (
Figure 8). These values were significantly higher than at other intensities (p < 0.05). The lowest astaxanthin productivity per mole of photons was 3.5 mg m
−2 mole photons
−1 at 1000 µmol photons m
−2 s
−1.
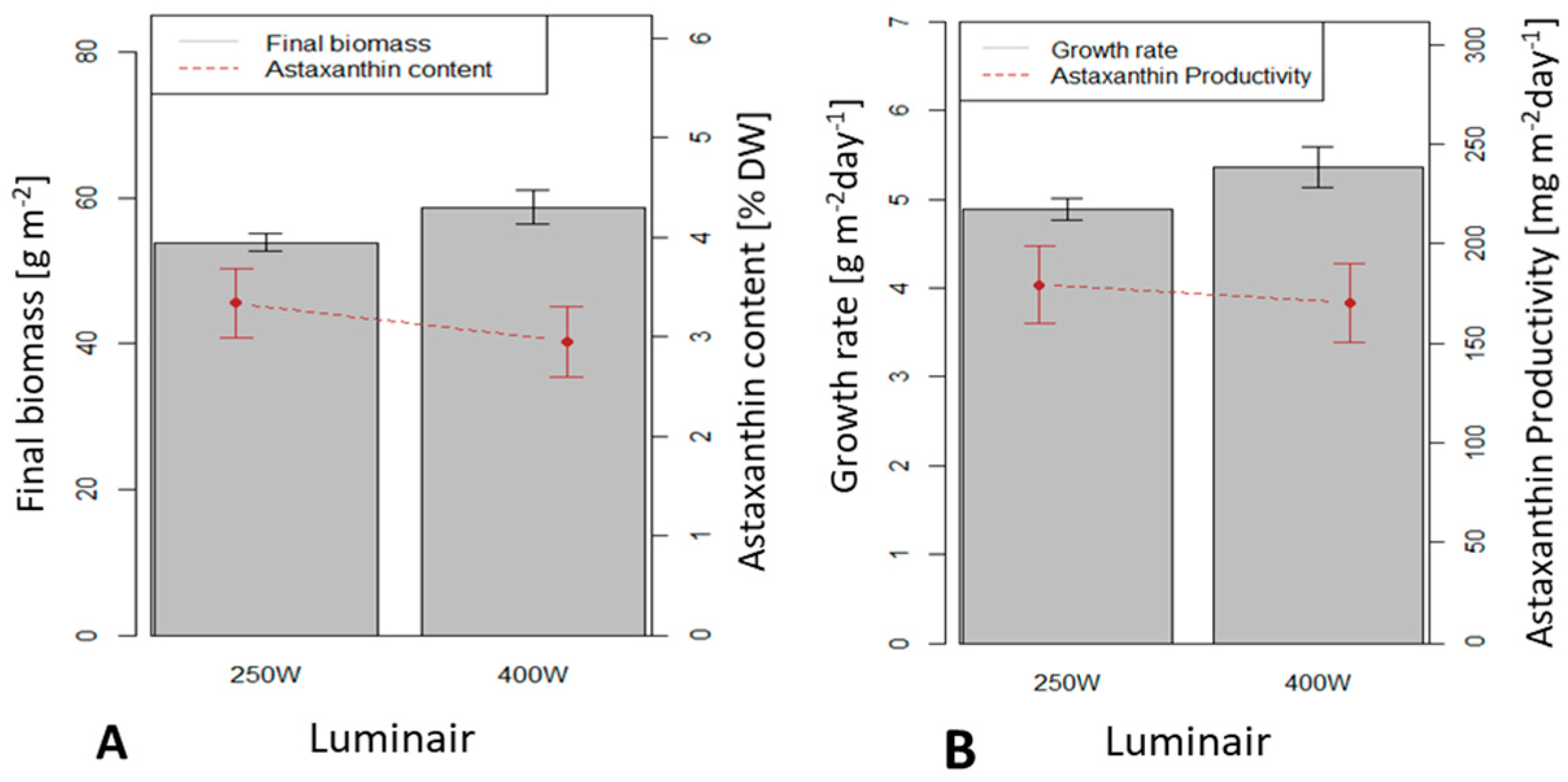
3.4. Influence of the Light Source on H. pluvialis Immobilized Cultivation in a 0.5 m2 x 4 (Large Scale) Twin-Layer Photobioreactor
This study investigated the illumination efficiency of two different luminaires: luminair 250 W and luminair 400 W. The luminair 400 W yielded average dry biomass productivity of 5.4 g m
−2 d
−1, significantly higher than the productivity under luminair 250 W (4.8 g m
−2 d
−1) (
p < 0.05). Astaxanthin contents in biomass under luminair 250 W and luminair 400 W (3.34 and 3.13 %, respectively) were not significantly different (
p > 0.05) (
Figure 9).
Astaxanthin productivities using luminair 250 W and 400 W (179.4 and 180.8 mg m−2 d−1, respectively) were not significantly different (p > 0.05). The luminair 400 W yielded higher dry biomass productivity but lower astaxanthin content than the other; therefore astaxanthin productivities were similar under both luminaires.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}









