Adenosine 5′-Triphosphate Metabolism in Red Blood Cells as a Potential Biomarker for Post-Exercise Hypotension and a Drug Target for Cardiovascular Protection †

 ,
, 
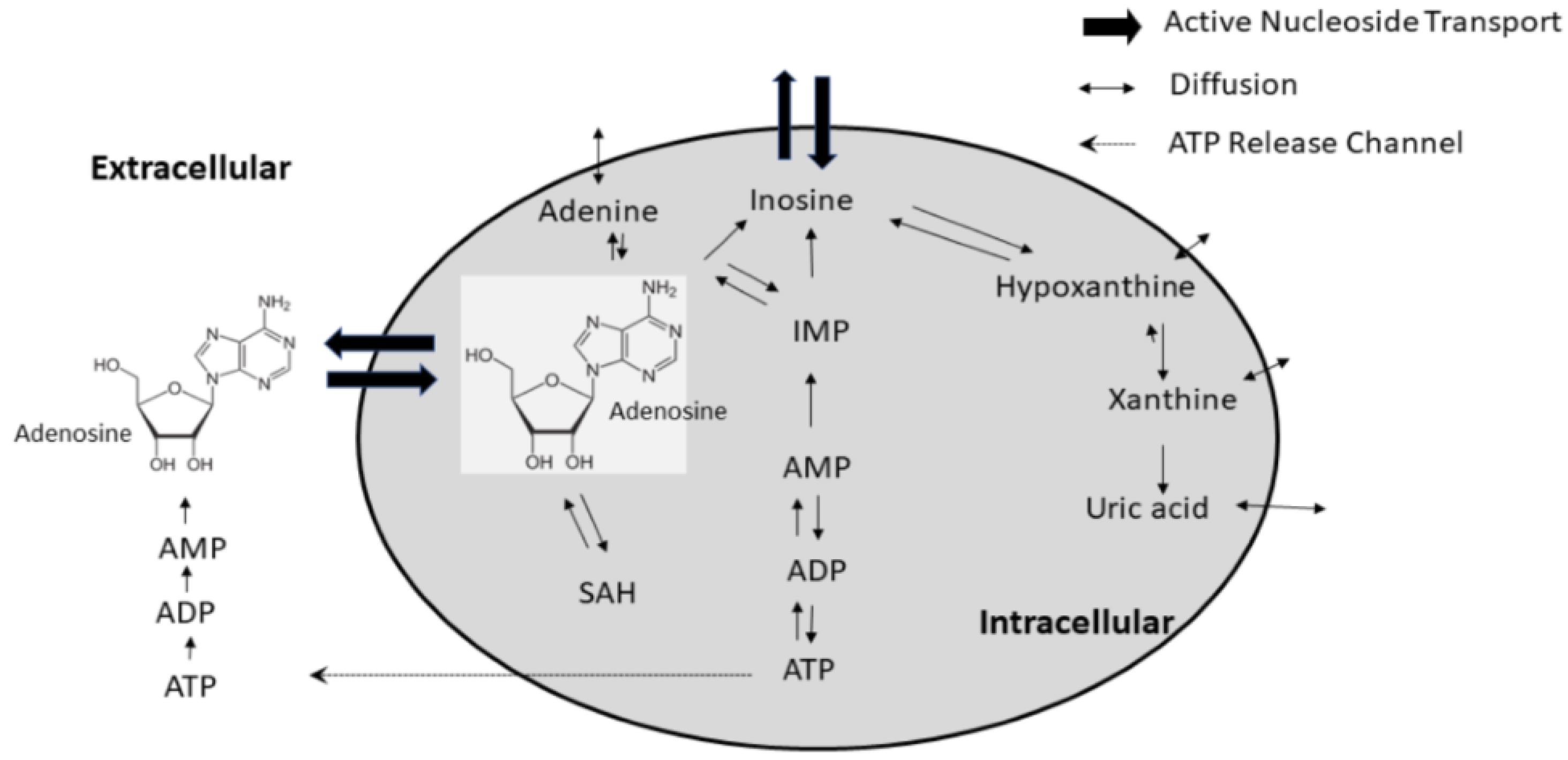{kind=link}
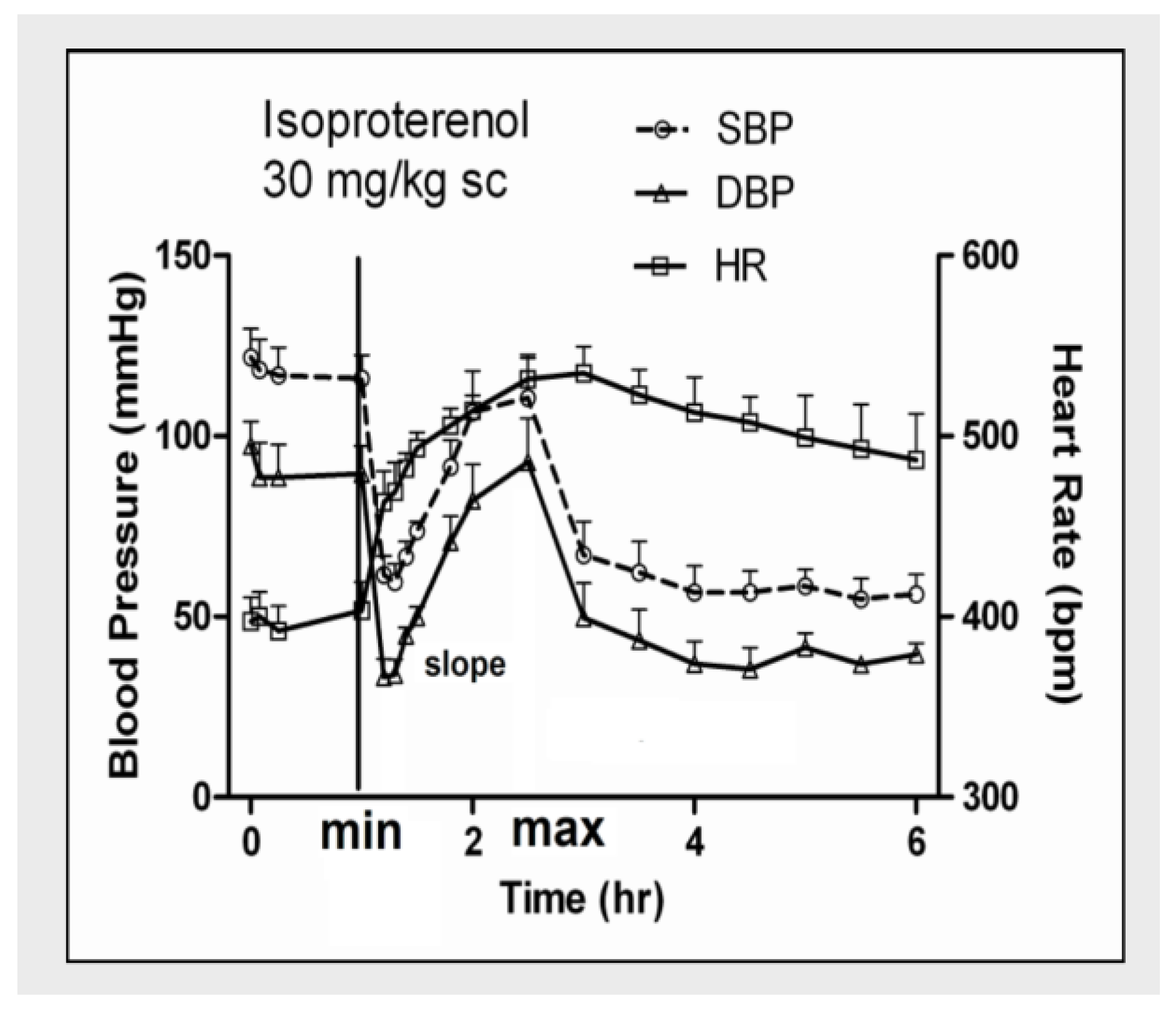{kind=link}
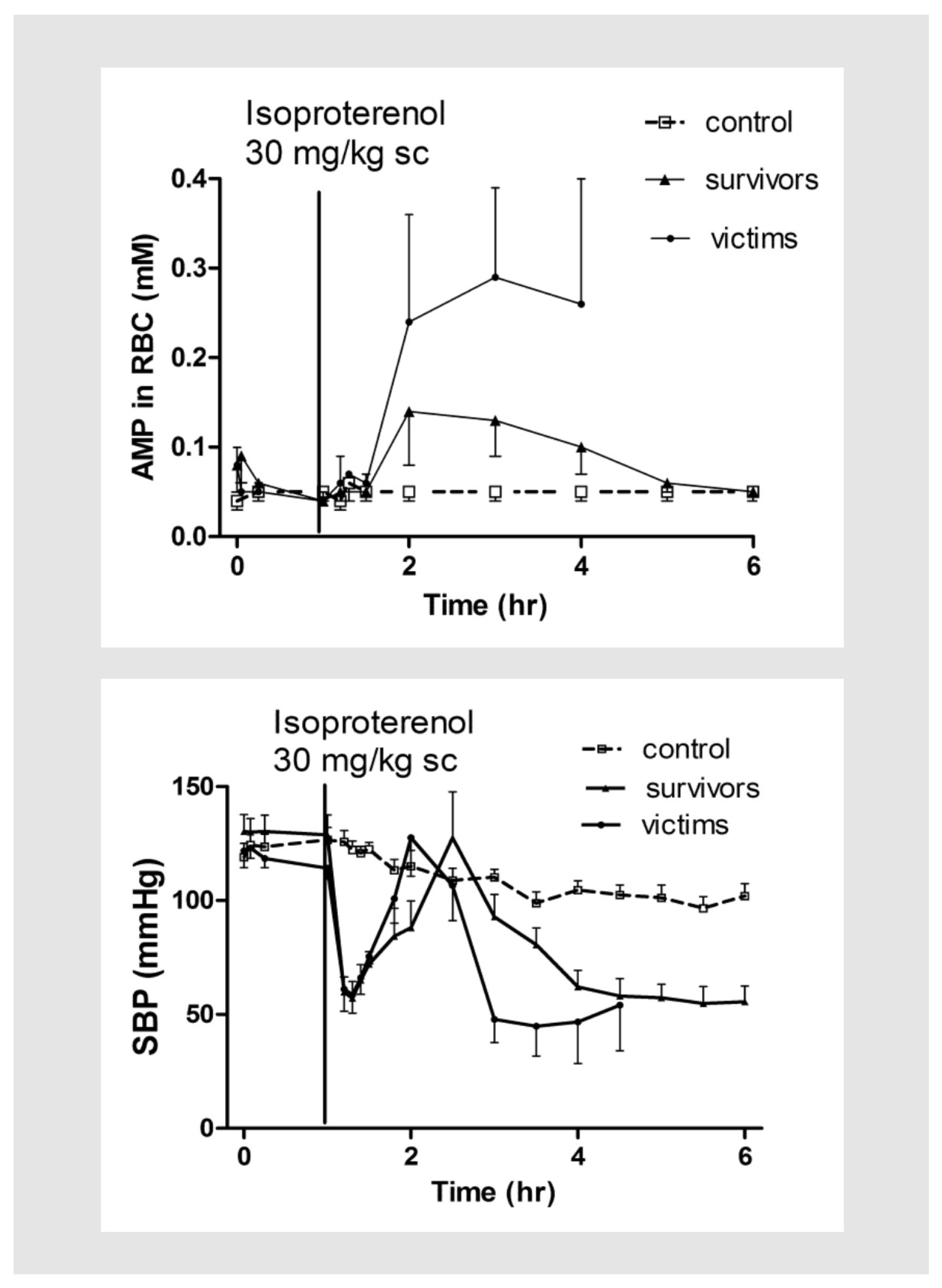{kind=link}
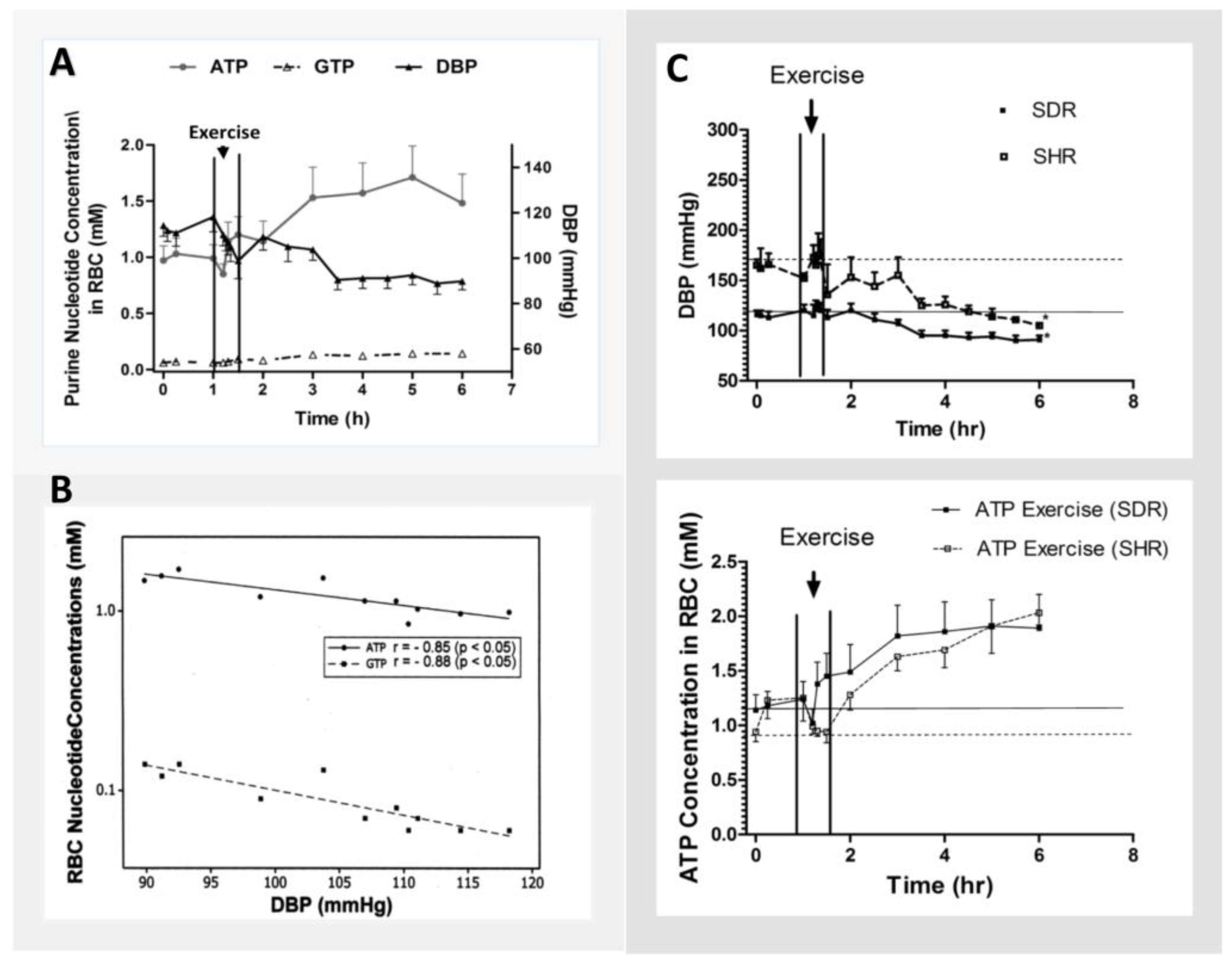{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Biomarker for Cardiovascular Toxicities
3. Biomarker for Post-Exercise Hypotension and Cardiovascular Protection
4. Biomarker and Target for Development of Drug Therapy
5. Challenges, Opportunities, and Future Direction
6. Concluding Remarks
Acknowledgments
Conflicts of Interest
References
- Sollevi, A. Cardiovascular effects of adenosine in man; possible clinical implications. Prog. Neurobiol. 1986, 27, 319–349. [Google Scholar] [CrossRef]
- Olsson, R.; Pearson, J. Cardiovascular purinoceptors. Physiol. Rev. 1990, 70, 761–845. [Google Scholar] [CrossRef] [PubMed]
- Ely, S.W.; Berne, R.M. Protective effects of adenosine in myocardial ischemia. Circulation 1992, 85, 893–904. [Google Scholar] [CrossRef] [PubMed]
- Shiode, N.; Kato, M.; Nakayama, K.; Shinohara, K.; Kurokawa, J.; Yamagata, T.; Matsuura, H.; Kajiyama, G. Effect of adenosine triphosphate on human coronary circulation. Intern. Med. 1998, 37, 818–825. [Google Scholar] [CrossRef] [PubMed]
- Oxhorn, B.C.; Cheek, D.J.; Buxton, I.L. Role of nucleotides and nucleosides in the regulation of cardiac blood flow. AACN Clin. Issues 2000, 11, 241–251. [Google Scholar] [CrossRef] [PubMed]
- Linden, J. Molecular approach to adenosine receptors: Receptor-mediated mechanisms of tissue protection. Annu. Rev. Pharmacol. Toxicol. 2001, 41, 775–787. [Google Scholar] [CrossRef] [PubMed]
- Burnstock, G. Purinergic signaling and vascular cell proliferation and death. Arterioscler. Thromb. Vasc. Biol. 2002, 22, 364–373. [Google Scholar] [CrossRef] [PubMed]
- Moens, A.L.; Claeys, M.J.; Timmermans, J.P.; Vrints, C.J. Myocardial ischemia/reperfusion-injury, a clinical view on a complex pathophysiological process. Int. J. Cardiol. 2005, 100, 179–190. [Google Scholar] [CrossRef] [PubMed]
- Ingwall, J.S. Energy metabolism in heart failure and remodelling. Cardiovasc. Res. 2009, 81, 412–419. [Google Scholar] [CrossRef] [PubMed]
- Cohen, M.V.; Downey, J.M. Adenosine: Trigger and mediator of cardioprotection. Basic Res. Cardiol. 2008, 103, 203–215. [Google Scholar] [CrossRef] [PubMed]
- Berne, R. The role of adenosine in the regulation of coronary blood flow. Circ. Res. 1980, 47, 807–813. [Google Scholar] [CrossRef] [PubMed]
- Jeremias, A.; Filardo, S.D.; Whitbourn, R.J.; Kernoff, R.S.; Yeung, A.C.; Fitzgerald, P.J.; Yock, P.G. Effects of intravenous and intracoronary adenosine 5′-triphosphate as compared with adenosine on coronary flow and pressure dynamics. Circulation 2000, 101, 318–323. [Google Scholar] [CrossRef] [PubMed]
- Tune, J.D.; Richmond, K.N.; Gorman, M.W.; Feigl, E.O. Control of coronary blood flow during exercise. Exp. Biol. Med. (Maywood) 2002, 227, 238–250. [Google Scholar] [CrossRef] [PubMed]
- Gerlach, E.; Becker, B.F.; Nees, S. Formation of adenosine by vascular endothelium: A homeostatic and antithrombogenic mechanism? In The Topic and Perspectives in Adenosine Research; Gerlach, E., Becker, B.F., Eds.; Springer: New York, NY, USA, 1987; pp. 309–320. [Google Scholar]
- Ashraf, M.; Ahmad, S. Calcium preconditioning elicits a unique protection against the calcium paradox injury in rat heart: Role of adenosine. Circ. Res. 1993, 74, 360–367. [Google Scholar] [CrossRef]
- Kitakaze, M.; Hori, M.; Morioka, T.; Minamino, T.; Takashima, S.; Sato, H.; Shinozaki, Y.; Chujo, M.; Mori, H.; Inoue, M.; et al. Infarct size-Limiting effect of ischemic preconditioning is blunted by inhibition of 5′-nucleotidase activity and attenuation of adenosine release. Circulation 1994, 89, 1237–1246. [Google Scholar] [CrossRef] [PubMed]
- Obata, T. Adenosine production and its role in protection against ischemic and reperfusion injury of the myocardium. Nippon Yakurigaku Zasshi 2002, 119, 273–279. [Google Scholar] [CrossRef] [PubMed]
- Deckert, J.; Gleiter, C. Adenosine—An endogenous neuroprotective metabolite and neuromodulator. J. Neural Transm. Suppl. 1994, 43, 23–31. [Google Scholar] [PubMed]
- Zalewska-Kaszubska, J. [Neuroprotective mechanisms of adenosine action on CNS neurons]. Neurologia i Neurochirurgia Polska 2002, 36, 329–336. [Google Scholar] [PubMed]
- Ribeiro, J.A.; Sebastiao, A.M.; de Mendonca, A. Participation of adenosine receptors in neuroprotection. Drug News Perspect. 2003, 16, 80–86. [Google Scholar] [CrossRef] [PubMed]
- Burnstock, G. Purines and sensory nerves. Handb. Exp. Pharmacol. 2009, 333–392. [Google Scholar] [CrossRef]
- Leonelli, M.; Torrao, A.S.; Britto, L.R. Unconventional neurotransmitters, neurodegeneration and neuroprotection. Braz. J. Med. Biol. Res. 2009, 42, 68–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ralevic, V. Purines as neurotransmitters and neuromodulators in blood vessels. Curr. Vasc. Pharmacol. 2009, 7, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Franco, M.; Perez-Mendez, O.; Martinez, F. Interaction of intrarenal adenosine and angiotensin II in kidney vascular resistance. Curr. Opin. Nephrol. Hypertens. 2009, 18, 63–67. [Google Scholar] [CrossRef] [PubMed]
- Burnstock, G. Purinergic signalling: Past, present and future. Braz. J. Med. Biol. Res. 2009, 42, 3–8. [Google Scholar] [CrossRef] [PubMed]
- Cain, B.S.; Meldrum, D.R.; Dinarello, C.A.; Meng, X.; Banerjee, A.; Harken, A.H. Adenosine reduces cardiac TNF-alpha production and human myocardial injury following ischemia-reperfusion. J. Surg. Res. 1998, 76, 117–123. [Google Scholar] [CrossRef] [PubMed]
- De Jong, J.W.; de Jonge, R.; Keijzer, E.; Bradamante, S. The role of adenosine in preconditioning. Pharmacol. Ther. 2000, 87, 141–149. [Google Scholar] [CrossRef]
- Funahashi, M. Effects of ischemic preconditioning on myocardial protective on cardiac surgery: Possibility of ischemic preconditioning and adenosine administration. Ann. Thorac. Cardiovasc. Surg. 2003, 9, 307–313. [Google Scholar] [PubMed]
- Donato, M.; Gelpi, R.J. Adenosine and cardioprotection during reperfusion—An overview. Mol. Cell. Biochem. 2003, 251, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Das, M.; Das, D.K. Molecular mechanism of preconditioning. IUBMB Life 2008, 60, 199–203. [Google Scholar] [CrossRef] [PubMed]
- Reffelmann, T.; Schwarz, E.R.; Skobel, C.E.; Petek, O.; Hanrath, P. [Ischemic preconditioning. Does this animal experiment phenomenon have clinical relevance?]. Med. Klin. 2000, 95, 559–567. [Google Scholar] [CrossRef]
- Light, P.E. Cardiac KATP channels and ischemic preconditioning: Current perspectives. Can. J. Cardiol. 1999, 15, 1123–1130. [Google Scholar] [PubMed]
- McCallion, K.; Harkin, D.W.; Gardiner, K.R. Role of adenosine in immunomodulation: Review of the literature. Crit. Care Med. 2004, 32, 273–277. [Google Scholar] [CrossRef] [PubMed]
- Dal Ben, D.; Antonioli, L.; Lambertucci, C.; Fornai, M.; Blandizzi, C.; Volpini, R. Purinergic Ligands as Potential Therapeutic Tools for the Treatment of Inflammation-Related Intestinal Diseases. Front. Pharmacol. 2018, 9, 212. [Google Scholar] [CrossRef] [PubMed]
- Rossi, A.; Lortet, S. Energy metabolism patterns in mammalian myocardium adapted to chronic physiopathological conditions. Cardiovasc. Res. 1996, 31, 163–171. [Google Scholar] [CrossRef]
- Sommerschild, H.T.; Kirkeboen, K.A. Adenosine and cardioprotection during ischaemia and reperfusion—An overview. Acta Anaesthesiol. Scand. 2000, 44, 1038–1055. [Google Scholar] [CrossRef] [PubMed]
- Porkka-Heiskanen, T.; Kalinchuk, A.; Alanko, L.; Urrila, A.; Stenberg, D. Adenosine, energy metabolism, and sleep. Sci. World J. 2003, 3, 790–798. [Google Scholar] [CrossRef] [PubMed]
- Camici, M.; Garcia-Gil, M.; Tozzi, M.G. The Inside Story of Adenosine. Int. J. Mol. Sci. 2018, 19, 784. [Google Scholar] [CrossRef] [PubMed]
- Bahreyni, A.; Samani, S.S.; Rahmani, F.; Behnam-Rassouli, R.; Khazaei, M.; Ryzhikov, M.; Parizadeh, M.R.; Avan, A.; Hassanian, S.M. Role of adenosine signaling in the pathogenesis of breast cancer. J. Cell. Physiol. 2018, 233, 1836–1843. [Google Scholar] [CrossRef] [PubMed]
- Deussen, A. Metabolic flux rates of adenosine in the heart. Naunyn Schmiedebergs Arch. Pharmacol. 2000, 362, 351–363. [Google Scholar] [CrossRef] [PubMed]
- Noji, T.; Karasawa, A.; Kusaka, H. Adenosine uptake inhibitors. Eur. J. Pharmacol. 2004, 495, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Fromonot, J.; Deharo, P.; Bruzzese, L.; Cuisset, T.; Quilici, J.; Bonatti, S.; Fenouillet, E.; Mottola, G.; Ruf, J.; Guieu, R. Adenosine plasma level correlates with homocysteine and uric acid concentrations in patients with coronary artery disease. Can. J. Physiol. Pharmacol. 2016, 94, 272–277. [Google Scholar] [CrossRef] [PubMed]
- Yeung, P.; Kolathuru, S.; Agu, R. Effect of Cardiovascular Injury on Catabolism of Adenosine and Adenosine 5-Triphosphate in Systemic Blood in a Freely Moving Rat Model In Vivo. Drug Metab. Lett. 2016, 10, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wang, Q.; Xue, Y.; Chen, J.; Lv, Q. Ticagrelor Compared with Clopidogrel Increased Adenosine and Cyclic Adenosine Monophosphate Plasma Concentration in Acute Coronary Syndrome Patients. Basic Clin. Pharmacol. Toxicol. 2017, 120, 610–614. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Duncker, D.J.; Xu, Y.; Zhang, Y.; Path, G.; Merkle, H.; Hendrich, K.; From, A.H.; Bache, R.J.; Ugurbil, K. Transmural bioenergetic responses of normal myocardium to high workstates. Am. J. Physiol. 1995, 268, H1891–H1905. [Google Scholar] [CrossRef] [PubMed]
- Hori, M.; Kitakaze, M. Adenosine, the heart, and coronary circulation. Hypertension 1991, 18, 565–574. [Google Scholar] [CrossRef] [PubMed]
- Panayiotou, C.; Solaroli, N.; Karlsson, A. The many isoforms of human adenylate kinases. Int. J. Biochem. Cell Biol. 2014, 49, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Schuster, S.; Kenanov, D. Adenine and adenosine salvage pathways in erythrocytes and the role of S-adenosylhomocysteine hydrolase. A theoretical study using elementary flux modes. FEBS J. 2005, 272, 5278–5290. [Google Scholar] [CrossRef] [PubMed]
- Scantland, S.; Tessaro, I.; Macabelli, C.H.; Macaulay, A.D.; Cagnone, G.; Fournier, E.; Luciano, A.M.; Robert, C. The adenosine salvage pathway as an alternative to mitochondrial production of ATP in maturing mammalian oocytes. Biol. Reprod. 2014, 91, 75. [Google Scholar] [CrossRef] [PubMed]
- Yeung, P. Metabolomics and biomarkers for drug discovery (Editorial). Metabolites 2018, 8, 11. [Google Scholar] [CrossRef] [PubMed]
- FDA Biomarkers Definition Working Group. Biomarkers and surrogate endpoints: Preferred definitions and conceptual framework. Clin. Pharmacol. Ther. 2001, 69, 89–95. [Google Scholar]
- DeJong, J.W. Diagnosis of Ischemic Heart Disease with AMP Catabolites; Martinus Nijhoff Publisher: Boston, MA, USA, 1988. [Google Scholar]
- Round, S.; Hsieh, L.; Agarwal, K. Effects of endotoxin injury on endothelial cell adenosine metabolism. J. Lab. Clin. Med. 1994, 123, 309–317. [Google Scholar]
- Yeung, P.; Dauphinee, J.; Simonson, K.; Gouzoules, T. RBC concentrations of ATP as potential in vivo biomarkers for cardiovascular safety of anti-hypertensive agents in rats. Clin. Pharmacol. Ther. 2009, 85 (Suppl. 1), S70. [Google Scholar]
- Yeung, P.; Ding, L.; Casley, W. HPLC assay with UV detection for determination of RBC purine nucleotides concentrations and application for biomarker study in vivo. J. Pharm. Biomed. Anal. 2008, 47, 377–382. [Google Scholar] [CrossRef] [PubMed]
- Klein, L.C.; Yeung, P.K.; Berman, J.N. Cladribine inhibits a diltiazem-induced increase in red blood cell purine nucleotide concentrations in a zebrafish model. Biomarkers 2009, 14, 554–559. [Google Scholar] [CrossRef] [PubMed]
- Yeung, P.; Dauphinee, J.; Simonson, K.; Gouzoules, T. Anti-Ischemia Drugs have no Effect on the In Vivo Metabolism of ATP by RBC in Normotensive Restrained Rats. Open Drug Metab. J. 2011, 5, 1–6. [Google Scholar] [CrossRef]
- Yeung, P.; Xu, Z.; Seeto, D. Diltiazem Reduces Mortality and Breakdown of ATP in Red Blood Cell Induced by Isoproterenol in a Freely Moving Rat Model in Vivo. Metabolites 2014, 4, 775–789. [Google Scholar] [CrossRef] [PubMed]
- Dudzinska, W.; Lubkowska, A.; Dolegowska, B.; Safranow, K.; Jakubowska, K. Adenine, guanine and pyridine nucleotides in blood during physical exercise and restitution in healthy subjects. Eur. J. Appl. Physiol. 2010, 110, 1155–1162. [Google Scholar] [CrossRef] [PubMed]
- Yeung, P.; Dauphinee, J.; Marcoux, T. Effect of acute exercise on cardiovascular hemodynamic and red blood cell concentrations of purine nucleotides in hypertensive compared with normotensives rats. Ther. Adv. Cardiovasc. Dis. 2013, 7, 63–74. [Google Scholar] [CrossRef] [PubMed]
- Ferdinandy, P.; Schulz, R.; Baxter, G.F. Interaction of cardiovascular risk factors with myocardial ischemia/reperfusion injury, preconditioning, and postconditioning. Pharmacol. Rev. 2007, 59, 418–458. [Google Scholar] [CrossRef] [PubMed]
- Yeung, P.K.; Tinkel, J.; Seeto, D. A Pilot Study to Assess Adenosine 5′-triphosphate Metabolism in Red Blood Cells as a Drug Target for Potential Cardiovascular Protection. Cardiovasc. Hematol. Disord. Drug Targets 2016, 15, 224–232. [Google Scholar] [CrossRef] [PubMed]
- Yeung, P.; Seeto, D. A study of the effect of isoproterenol on red blood cell concentrations of adenine nucleotides in a freely moving rat model in vivo. Cardiovasc. Pharmacol. (Open Access) 2013, 2, 102. [Google Scholar] [CrossRef]
- Yeung, P. Mohammadizadeha, Akhoundia, and Pulinilkunnilb, Assessing cardiovascular toxicity of doxorubicin and isoproterenol by hemodynamics and in vivo catabolism of adenosine 5′-triphosphate. In Proceedings of the Annual Meeting and Symposium of the CSPS/CC-CRS/CSP, Toronto, ON, Canada, 22–25 May 2018. [Google Scholar]
- Yeung, P.; Tinkel, J. Complementary Effect of Exercise in Cardiovascular Medicine (Editorial). Cardiovasc. Pharmacol. Open Access 2013, 2, e119. [Google Scholar]
- MacDonald, J.R. Potential causes, mechanisms, and implications of post exercise hypotension. J. Hum. Hypertens. 2002, 16, 225–236. [Google Scholar] [CrossRef] [PubMed]
- Yellon, D.M.; Hausenloy, D.J. Myocardial reperfusion injury. N. Engl. J. Med. 2007, 357, 1121–1135. [Google Scholar] [CrossRef] [PubMed]
- Di Francescomarino, S.; Sciartilli, A.; Di Valerio, V.; Di Baldassarre, A.; Gallina, S. The effect of physical exercise on endothelial function. Sports Med. 2009, 39, 797–812. [Google Scholar] [CrossRef] [PubMed]
- Kavazis, A.N. Exercise preconditioning of the myocardium. Sports Med. 2009, 39, 923–935. [Google Scholar] [CrossRef] [PubMed]
- Gorman, M.W.; Rooke, G.A.; Savage, M.V.; Jayasekara, M.P.; Jacobson, K.A.; Feigl, E.O. Adenine nucleotide control of coronary blood flow during exercise. Am. J. Physiol. Heart Circ. Physiol. 2010, 299, H1981–H1989. [Google Scholar] [CrossRef] [PubMed]
- Yeung, P.K.; Dauphinee, J.; Gouzoules, T.; Simonson, K.; Schindler, C. Exercise improves hemodynamic profiles and increases red blood cell concentrations of purine nucleotides in a rodent model. Ther. Adv. Cardiovasc. Dis. 2010, 4, 341–347. [Google Scholar] [CrossRef] [PubMed]
- Jimenez, S.K.; Jassal, D.S.; Kardami, E.; Cattini, P.A. A single bout of exercise promotes sustained left ventricular function improvement after isoproterenol-induced injury in mice. J. Physiol. Sci. 2011, 61, 331–336. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.J.; Pan, S.S.; Zhuang, T.; Wang, F.J. Exercise preconditioning initiates late cardioprotection against isoproterenol-induced myocardial injury in rats independent of protein kinase C. J. Physiol. Sci. 2011, 61, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Crimi, E.; Ignarro, L.J.; Cacciatore, F.; Napoli, C. Mechanisms by which exercise training benefits patients with heart failure. Nat. Rev. Cardiol. 2009, 6, 292–300. [Google Scholar] [CrossRef] [PubMed]
- Blair, S.N.; Kampert, J.B.; Kohl, H.W., 3rd; Barlow, C.E.; Macera, C.A.; Paffenbarger, R.S., Jr.; Gibbons, L.W. Influences of cardiorespiratory fitness and other precursors on cardiovascular disease and all-cause mortality in men and women. JAMA 1996, 276, 205–210. [Google Scholar] [CrossRef] [PubMed]
- Scott, J.M.; Adams, S.C.; Koelwyn, G.J.; Jones, L.W. Cardiovascular Late Effects and Exercise Treatment in Breast Cancer: Current Evidence and Future Directions. Can. J. Cardiol. 2016, 32, 881–890. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.J.; Wu, P.T.; Middlekauff, H.R.; Nguyen, K.L. Aerobic exercise in anthracycline-induced cardiotoxicity: A systematic review of current evidence and future directions. Am. J. Physiol. Heart Circ. Physiol. 2017, 312, H213–H222. [Google Scholar] [CrossRef] [PubMed]
- Prasad, D.S.; Das, B.C. Physical inactivity: A cardiovascular risk factor. Indian J. Med. Sci. 2009, 63, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Hunter, D.J.; Reddy, K.S. Noncommunicable Diseases: New England Journal of Medicine. N. Engl. J. Med. 2013, 369, 1336–1343. [Google Scholar] [CrossRef] [PubMed]
- La Gerche, A. The Potential Cardiotoxic Effects of Exercise. Can. J. Cardiol. 2016, 32, 421–428. [Google Scholar] [CrossRef] [PubMed]
- Ellsworth, M.L. The red blood cell as an oxygen sensor: What is the evidence? Acta Physiol. Scand. 2000, 168, 551–559. [Google Scholar] [CrossRef] [PubMed]
- Jensen, F.B. The dual roles of red blood cells in tissue oxygen delivery: Oxygen carriers and regulators of local blood flow. J. Exp. Biol. 2009, 212, 3387–3393. [Google Scholar] [CrossRef] [PubMed]
- Helms, C.C.; Gladwin, M.T.; Kim-Shapiro, D.B. Erythrocytes and Vascular Function: Oxygen and Nitric Oxide. Front. Physiol. 2018, 9, 125. [Google Scholar] [CrossRef] [PubMed]
- Bergfeld, G.R.; Forrester, T. Release of ATP from human erythrocytes in response to a brief period of hypoxia and hypercapnia. Cardiovasc. Res. 1992, 26, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Barneo, J.; Nurse, C.A.; Nilsson, G.E.; Buck, L.T.; Gassmann, M.; Bogdanova, A.Y. First aid kit for hypoxic survival: Sensors and strategies. Physiol. Biochem. Zool. 2010, 83, 753–763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frohlich, J.; Al-Sarraf, A. Cardiovascular risk and atherosclerosis prevention. Cardiovasc. Pathol. 2013, 22, 16–18. [Google Scholar] [CrossRef] [PubMed]
- Brook, R.D.; Appel, L.J.; Rubenfire, M.; Ogedegbe, G.; Bisognano, J.D.; Elliott, W.J.; Fuchs, F.D.; Hughes, J.W.; Lackland, D.T.; Staffileno, B.A.; et al. Beyond medications and diet: Alternative approaches to lowering blood pressure: A scientific statement from the American heart association. Hypertension 2013, 61, 1360–1383. [Google Scholar] [CrossRef] [PubMed]
- Marongiu, E.; Crisafulli, A. Cardioprotection acquired through exercise: The role of ischemic preconditioning. Curr. Cardiol. Rev. 2014, 10, 336–348. [Google Scholar] [CrossRef] [PubMed]
- Antunes, B.M.; Rossi, F.E.; Cholewa, J.M.; Lira, F.S. Regular Physical Activity and Vascular Aging. Curr. Pharm. Des. 2016, 22, 3715–3729. [Google Scholar] [CrossRef] [PubMed]
- Yeung, P. ATP Metabolism as Biomarker Target for Cardiovascular Protection (Editorial). Cardiol. Pharmacol. 2013, 2, e118. [Google Scholar] [CrossRef]
- Yeung, P.K.F.; Mosher, S.J.; MacRae, D.A.; Klassen, G.A. Effect of diltiazem and its metabolites on the uptake of adenosine in blood: An in-vitro investigation. J. Pharm. Pharmacol. 1991, 43, 685–689. [Google Scholar] [CrossRef] [PubMed]
- Kolathuru, S.; Tillman, P.; Yeung, P. Effect of Dipyridamole on Red Blood Cell Concentrations of Purine Nucleotides After Acute Myocardial Infarction in a Freely Moving Rat Model (A-51). JPPS 2015, 18, 89. [Google Scholar]
- Dunston, C.R.; Griffiths, H.R.; Lambert, P.A.; Staddon, S.; Vernallis, A.B. Proteomic analysis of the anti-inflammatory action of minocycline. Proteomics 2011, 11, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Tillman, P.; Agu, R.; Yeung, P. Coenzyme Q10 lowers blood pressure and heart rate and increases red blood cell concentrations of ATP in a freely moving rat model (A13). JPPS 2014, 17, 67S. [Google Scholar]
- Yeung, P.; Rippke, J.; Borowiak, M.; Zhou, J.; Islam, S.; Lehmann, C. Effect of tigecycline on ATP concentrations in red blood cell in rats (A-112). JPPS 2015, 18, 128. [Google Scholar]
- Feng, J.; Yeung, P. A simple high performance liquid chromatography assay for simultaneous measurement of adenosine, guanosine, and the oxypurine metabolites in plasma. Ther. Drug Monit. 2000, 22, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Gewirtz, H.; Brown, P.; Most, A.S. Measurement of plasma adenosine concentration: Methodological and physiological considerations. Proc. Soc. Exp. Biol. Med. 1987, 185, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Yeung, P.; Feng, J.; Buckley, S. High performance liquid chromatography assay of adenosine, adenine nucleotides, and their purine metabolites in plasma and erythrocytes. Pharm. Res. 1995, 12, S51. [Google Scholar]
- Zhang, Y.; Geiger, J.; Lautt, W. Improved high-pressure liquid chromatographic-fluorometric assay for measurement of adenosine in plasma. Gastrointest. Liver Physiol. 1991, 23, G658–G664. [Google Scholar] [CrossRef] [PubMed]






© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yeung, P.K.; Kolathuru, S.S.; Mohammadizadeh, S.; Akhoundi, F.; Linderfield, B. Adenosine 5′-Triphosphate Metabolism in Red Blood Cells as a Potential Biomarker for Post-Exercise Hypotension and a Drug Target for Cardiovascular Protection. Metabolites 2018, 8, 30. https://doi.org/10.3390/metabo8020030
Yeung PK, Kolathuru SS, Mohammadizadeh S, Akhoundi F, Linderfield B. Adenosine 5′-Triphosphate Metabolism in Red Blood Cells as a Potential Biomarker for Post-Exercise Hypotension and a Drug Target for Cardiovascular Protection. Metabolites. 2018; 8(2):30. https://doi.org/10.3390/metabo8020030
Chicago/Turabian StyleYeung, Pollen K., Shyam Sundar Kolathuru, Sheyda Mohammadizadeh, Fatemeh Akhoundi, and Brett Linderfield. 2018. "Adenosine 5′-Triphosphate Metabolism in Red Blood Cells as a Potential Biomarker for Post-Exercise Hypotension and a Drug Target for Cardiovascular Protection" Metabolites 8, no. 2: 30. https://doi.org/10.3390/metabo8020030