Retinoic Acid Signaling Regulates the Metamorphosis of Feather Stars (Crinoidea, Echinodermata): Insight into the Evolution of the Animal Life Cycle


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling and Culture of Larvae
2.2. Immunohistochemistry
2.3. Reagent Treatments
2.4. Statistical Analysis
2.5. Construction of the Phylogenic Trees
3. Results
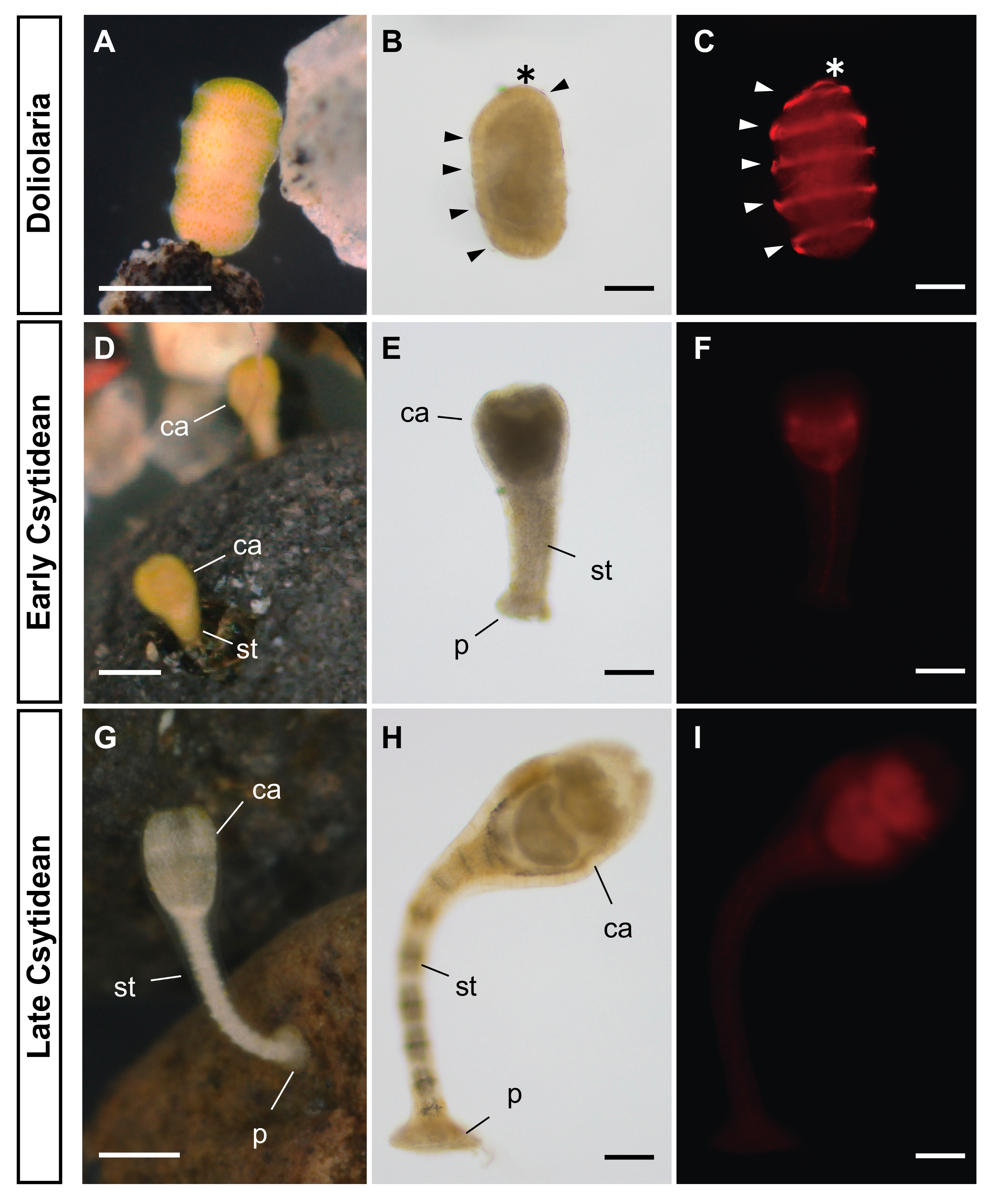
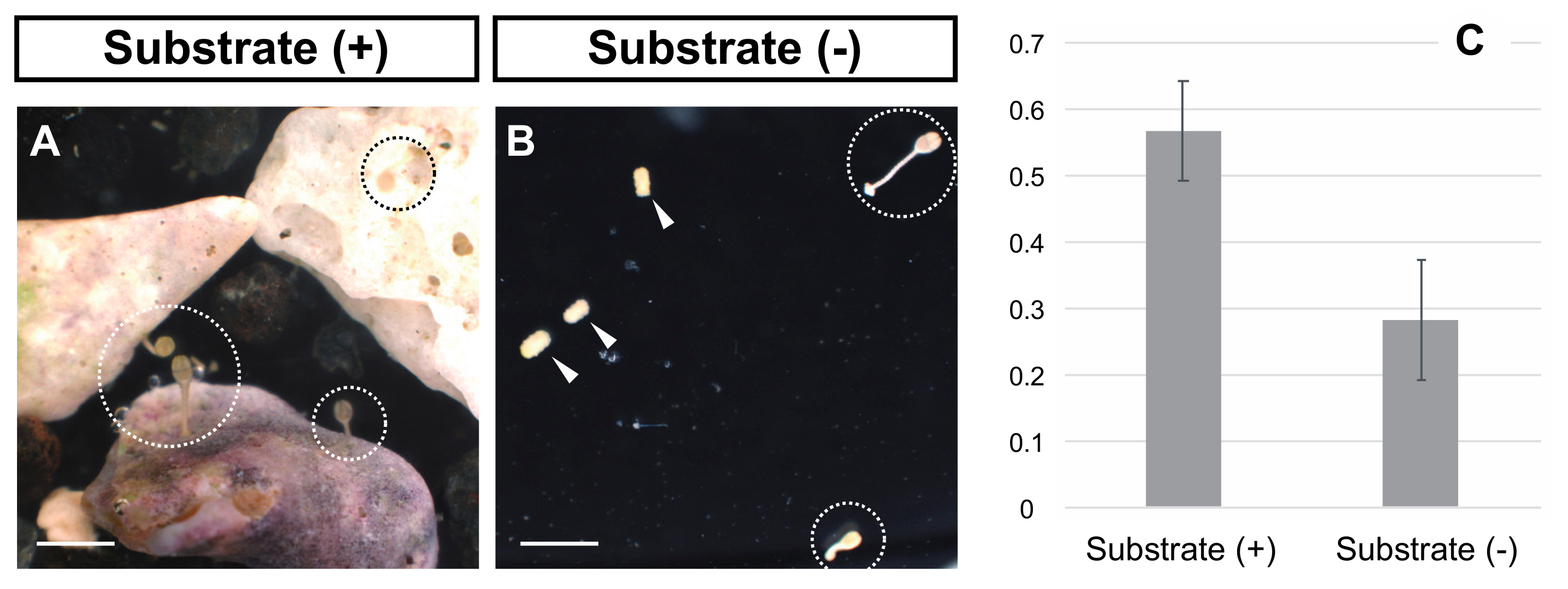
3.1. Incubation with Natural Substrates Stimulated the Metamorphosis of A. serrata
3.2. Exogenous RA Treatment Induced the Metamorphosis of A. serrata
3.3. Endogenous RA Synthesis is Required for the Metamorphosis of A. serrata
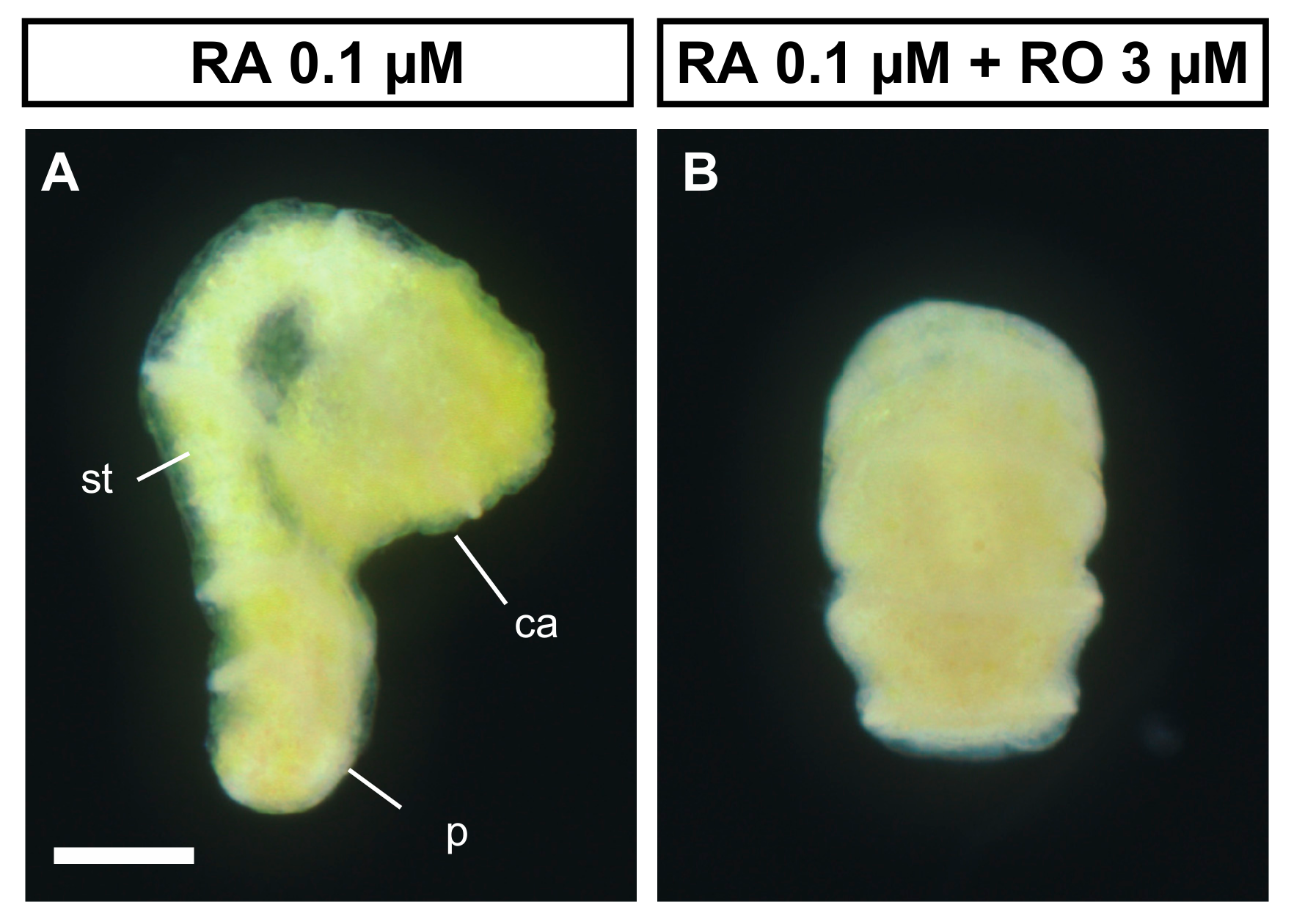
3.4. RA Binding with RAR is Required for the Metamorphosis of A. serrata
4. Discussion
4.1. Metamorphosis Regulation by RA Signaling in the Ancestor of Living Echinoderms
4.2. Life Cycle Evolution from the Viewpoint of RA Signaling
4.3. Insight into the Ancestral Function of RA Signaling
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Jägersten, G. Evolution of the Metazoan Life Cycle; Academic Press: London, UK, 1972. [Google Scholar]
- Jackson, D.; Leys, S.P.; Hinman, V.F.; Woods, R.; Lavin, M.F.; Degnan, B.M. Ecological regulation of development: Induction of marine invertebrate metamorphosis. Int. J. Dev. Biol. 2002, 46, 679–686. [Google Scholar]
- Degnan, S.M.; Degnan, B.M. The initiation of metamorphosis as an ancient polyphenic trait and its role in metazoan life-cycle evolution. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2010, 365, 641–651. [Google Scholar] [CrossRef] [Green Version]
- Marlow, H.; Tosches, M.A.; Tomer, R.; Steinmetz, P.R.; Lauri, A.; Larsson, T.; Arendt, D. Larval body patterning and apical organs are conserved in animal evolution. BMC Biol. 2014, 12, 7. [Google Scholar] [CrossRef] [Green Version]
- Darras, S.; Gerhart, J.; Terasaki, M.; Kirschner, M.; Lowe, C.J. β-catenin specifies the endomesoderm and defines the posterior organizer of the hemichordate Saccoglossus kowalevskii. Development 2011, 138, 959–970. [Google Scholar] [CrossRef] [Green Version]
- Marlow, H.; Matus, D.Q.; Martindale, M.Q. Ectopic activation of the canonical wnt signaling pathway affects ectodermal patterning along the primary axis during larval development in the anthozoan Nematostella vectensis. Dev. Biol. 2013, 380, 324–334. [Google Scholar] [CrossRef] [Green Version]
- Range, R.C.; Angerer, R.C.; Angerer, L.M. Integration of canonical and noncanonical Wnt signaling pathways patterns the neuroectoderm along the anterior-posterior axis of sea urchin embryos. PLoS Biol. 2013, 11, e1001467. [Google Scholar] [CrossRef] [Green Version]
- Raff, R.A. Origins of the other metazoan body plans: The evolution of larval forms. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2008, 363, 1473–1479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuchs, B.; Wang, W.; Graspeuntner, S.; Li, Y.; Insua, S.; Herbst, E.M.; Dirksen, P.; Bohm, A.M.; Hemmrich, G.; Sommer, F.; et al. Regulation of polyp-to-jellyfish transition in Aurelia aurita. Curr. Biol. 2014, 24, 263–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamakawa, S.; Morino, Y.; Honda, M.; Wada, H. The role of retinoic acid signaling in starfish metamorphosis. EvoDevo 2018, 9, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murabe, N.; Hatoyama, H.; Komatsu, M.; Kaneko, H.; Nakajima, Y. Adhesive papillae on the brachiolar arms of brachiolaria larvae in two starfishes, Asterina pectinifera and Asterias amurensis, are sensors for metamorphic inducing factor(s). Dev. Growth Differ. 2007, 49, 647–656. [Google Scholar] [CrossRef] [PubMed]
- Handberg-Thorsager, M.; Gutierrez-Mazariegos, J.; Arold, S.T.; Kumar Nadendla, E.; Bertucci, P.Y.; Germain, P.; Tomancak, P.; Pierzchalski, K.; Jones, J.W.; Albalat, R.; et al. The ancestral retinoic acid receptor was a low-affinity sensor triggering neuronal differentiation. Sci. Adv. 2018, 4, eaao1261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Telford, M.J.; Lowe, C.J.; Cameron, C.B.; Ortega-Martinez, O.; Aronowicz, J.; Oliveri, P.; Copley, R.R. Phylogenomic analysis of echinoderm class relationships supports Asterozoa. Proc. Biol. Sci. 2014, 281. [Google Scholar] [CrossRef] [PubMed]
- Hart, M.W. Life history evolution and comparative developmental biology of echinoderms. Evol. Dev. 2002, 4, 62–71. [Google Scholar] [CrossRef] [PubMed]
- Hyman, L.H. The Invertebrates. IV. Echinodermata; McGraw-Hill Book Company, Inc.: New York, NY, USA, 1955. [Google Scholar]
- McEdward, L.R.; Miner, B.G. Larval and life-cycle patterns in echinoderms. Can. J. Zool. 2001, 79, 1125–1170. [Google Scholar] [CrossRef]
- Raff, R.A.; Byrne, M. The active evolutionary lives of echinoderm larvae. Heredity 2006, 97, 244–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heyland, A.; Schuh, N.; Rast, J. Sea Urchin Larvae as a Model for Postembryonic Development. In Marine Organisms as Model Systems in Biology and Medicine; Kloc, M., Kubiak, J.Z., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 137–161. [Google Scholar] [CrossRef]
- Sutherby, J.; Giardini, J.-L.; Nguyen, J.; Wessel, G.; Leguia, M.; Heyland, A. Histamine is a modulator of metamorphic competence in Strongylocentrotus purpuratus (Echinodermata: Echinoidea). BMC Evol. Biol. 2012, 12, 14. [Google Scholar] [CrossRef] [Green Version]
- Chino, Y.; Saito, M.; Yamasu, K.; Suyemitsu, T.; Ishihara, K. Formation of the adult rudiment of sea urchins is influenced by thyroid hormones. Dev. Biol. 1994, 161, 1–11. [Google Scholar] [CrossRef]
- Bishop, C.D.; Brandhorst, B.P. Development of nitric oxide synthase-defined neurons in the sea urchin larval ciliary band and evidence for a chemosensory function during metamorphosis. Dev. Dyn. 2007, 236, 1535–1546. [Google Scholar] [CrossRef]
- Bishop, C.D.; Brandhorst, B.P. NO/cGMP signaling and HSP90 activity represses metamorphosis in the sea urchin Lytechinus pictus. Biol. Bull. 2001, 201, 394–404. [Google Scholar] [CrossRef]
- The R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]
- Koga, H.; Fujitani, H.; Morino, Y.; Miyamoto, N.; Tsuchimoto, J.; Shibata, T.F.; Nozawa, M.; Shigenobu, S.; Ogura, A.; Tachibana, K.; et al. Experimental Approach Reveals the Role of alx1 in the Evolution of the Echinoderm Larval Skeleton. PLoS ONE 2016, 11, e0149067. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capella-Gutiérrez, S.; Silla-Martínez, J.M.; Gabaldón, T. trimAl: A tool for automated alignment trimming in large-scale phylogenetic analyses. Bioinformatics 2009, 25, 1972–1973. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Barbaglio, A.; Turchi, C.; Melone, G.; Benedetto, C.; Martinello, T.; Patruno, M.; Biggiogero, M.; Wilkie, I.; Candia, D. Larval development in the feather star Antedon mediterranea. Invertebr. Reprod. Dev. 2012, 56, 124–137. [Google Scholar] [CrossRef]
- Amemiya, S.; Omori, A.; Tsurugaya, T.; Hibino, T.; Yamaguchi, M.; Kuraishi, R.; Kiyomoto, M.; Minokawa, T. Early stalked stages in ontogeny of the living isocrinid sea lily Metacrinus rotundus. Acta Zool. 2016, 97, 102–116. [Google Scholar] [CrossRef]
- Haig, J.A.; Rouse, G.W. Larval development of the featherstar Aporometra wilsoni (Echinodermata: Crinoidea). Invertebr. Biol. 2008, 127, 460–469. [Google Scholar] [CrossRef]
- Mladenov, P.V.; Chia, F.S. Development, settling behaviour, metamorphosis and pentacrinoid feeding and growth of the feather star Florometra serratissima. Mar. Biol. 1983, 73, 309–323. [Google Scholar] [CrossRef]
- Pearce, C. Induction of settlement and metamorphosis in echinoderms. In Recent Advances in Marine Biotechnology; Fingerman, M., Nagabhushanam, R., Eds.; Oxford and IBH Publishing Co: New Delhi, India, 1997; pp. 283–342. [Google Scholar]
- Rhinn, M.; Dolle, P. Retinoic acid signalling during development. Development 2012, 139, 843–858. [Google Scholar] [CrossRef] [Green Version]
- Marlétaz, F.; Holland, L.Z.; Laudet, V.; Schubert, M. Retinoic acid signaling and the evolution of chordates. Int. J. Biol. Sci. 2006, 2, 38. [Google Scholar] [CrossRef]
- Gutierrez-Mazariegos, J.; Schubert, M.; Laudet, V. Evolution of retinoic acid receptors and retinoic acid signaling. Subcell. Biocchem. 2014, 70, 55–73. [Google Scholar] [CrossRef]
- Amemiya, S.; Hibino, T.; Nakano, H.; Yamaguchi, M.; Kuraishi, R.; Kiyomoto, M. Development of ciliary bands in larvae of the living isocrinid sea lily Metacrinus rotundus. Acta Zool. 2015, 96, 36–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakano, H.; Hibino, T.; Oji, T.; Hara, Y.; Amemiya, S. Larval stages of a living sea lily (stalked crinoid echinoderm). Nature 2003, 421, 158–160. [Google Scholar] [CrossRef] [PubMed]
- Cañestro, C.; Postlethwait, J.H.; Gonzàlez-Duarte, R.; Albalat, R. Is retinoic acid genetic machinery a chordate innovation? Evol. Dev. 2006, 8, 394–406. [Google Scholar] [CrossRef] [PubMed]
- Albalat, R. The retinoic acid machinery in invertebrates: Ancestral elements and vertebrate innovations. Mol. Cell. Endocrinol. 2009, 313, 23–35. [Google Scholar] [CrossRef]
- Smirnov, A.V. Sea cucumbers symmetry (Echinodermata: Holothuroidea). Paleontol. J. 2014, 48, 1215–1236. [Google Scholar] [CrossRef]
- Kayal, E.; Bentlage, B.; Sabrina Pankey, M.; Ohdera, A.H.; Medina, M.; Plachetzki, D.C.; Collins, A.G.; Ryan, J.F. Phylogenomics provides a robust topology of the major cnidarian lineages and insights on the origins of key organismal traits. BMC Evol. Biol. 2018, 18, 68. [Google Scholar] [CrossRef] [Green Version]
- Pennati, R.; Dell’Anna, A.; Zega, G.; De Bernardi, F.; Piraino, S. Retinoic acid influences antero-posterior positioning of peptidergic neurons in the planula larva of the hydrozoan Clava multicornis. Mar. Ecol. 2013, 34, 143–152. [Google Scholar] [CrossRef]
- Fischer, A.H.L.; Henrich, T.; Arendt, D. The normal development of Platynereis dumerilii (Nereididae, Annelida). Front. Zool. 2010, 7, 31. [Google Scholar] [CrossRef] [Green Version]
- Cloney, R.A. Ascidian Larvae and the Events of Metamorphosis. Am. Zool. 1982, 22, 817–826. [Google Scholar] [CrossRef] [Green Version]
- Karaiskou, A.; Swalla, B.J.; Sasakura, Y.; Chambon, J.P. Metamorphosis in solitary ascidians. Genesis 2015, 53, 34–47. [Google Scholar] [CrossRef]
- Röttinger, E.; Lowe, C.J. Evolutionary crossroads in developmental biology: Hemichordates. Development 2012, 139, 2463–2475. [Google Scholar] [CrossRef] [PubMed] [Green Version]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yamakawa, S.; Morino, Y.; Kohtsuka, H.; Wada, H. Retinoic Acid Signaling Regulates the Metamorphosis of Feather Stars (Crinoidea, Echinodermata): Insight into the Evolution of the Animal Life Cycle. Biomolecules 2020, 10, 37. https://doi.org/10.3390/biom10010037
Yamakawa S, Morino Y, Kohtsuka H, Wada H. Retinoic Acid Signaling Regulates the Metamorphosis of Feather Stars (Crinoidea, Echinodermata): Insight into the Evolution of the Animal Life Cycle. Biomolecules. 2020; 10(1):37. https://doi.org/10.3390/biom10010037
Chicago/Turabian StyleYamakawa, Shumpei, Yoshiaki Morino, Hisanori Kohtsuka, and Hiroshi Wada. 2020. "Retinoic Acid Signaling Regulates the Metamorphosis of Feather Stars (Crinoidea, Echinodermata): Insight into the Evolution of the Animal Life Cycle" Biomolecules 10, no. 1: 37. https://doi.org/10.3390/biom10010037