
Dimethyloxalylglycine (DMOG), a Hypoxia Mimetic Agent, Does Not Replicate a Rat Pheochromocytoma (PC12) Cell Biological Response to Reduced Oxygen Culture



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
2.1. Cell Culture
2.2. Alternate Oxygen Level Culture Conditions
2.3. DMOG Treatment
2.4. MTT Assays
2.5. Hypoxia-Inducible Factor-1α Subunit Expression Analysis
2.6. Reactive Oxygen Species (ROS) Measurement
2.7. Mitochondrial Burden and Action Potential
2.8. Cell Cycle Analysis
2.9. Differentiation
2.10. Data Analysis
3. Results
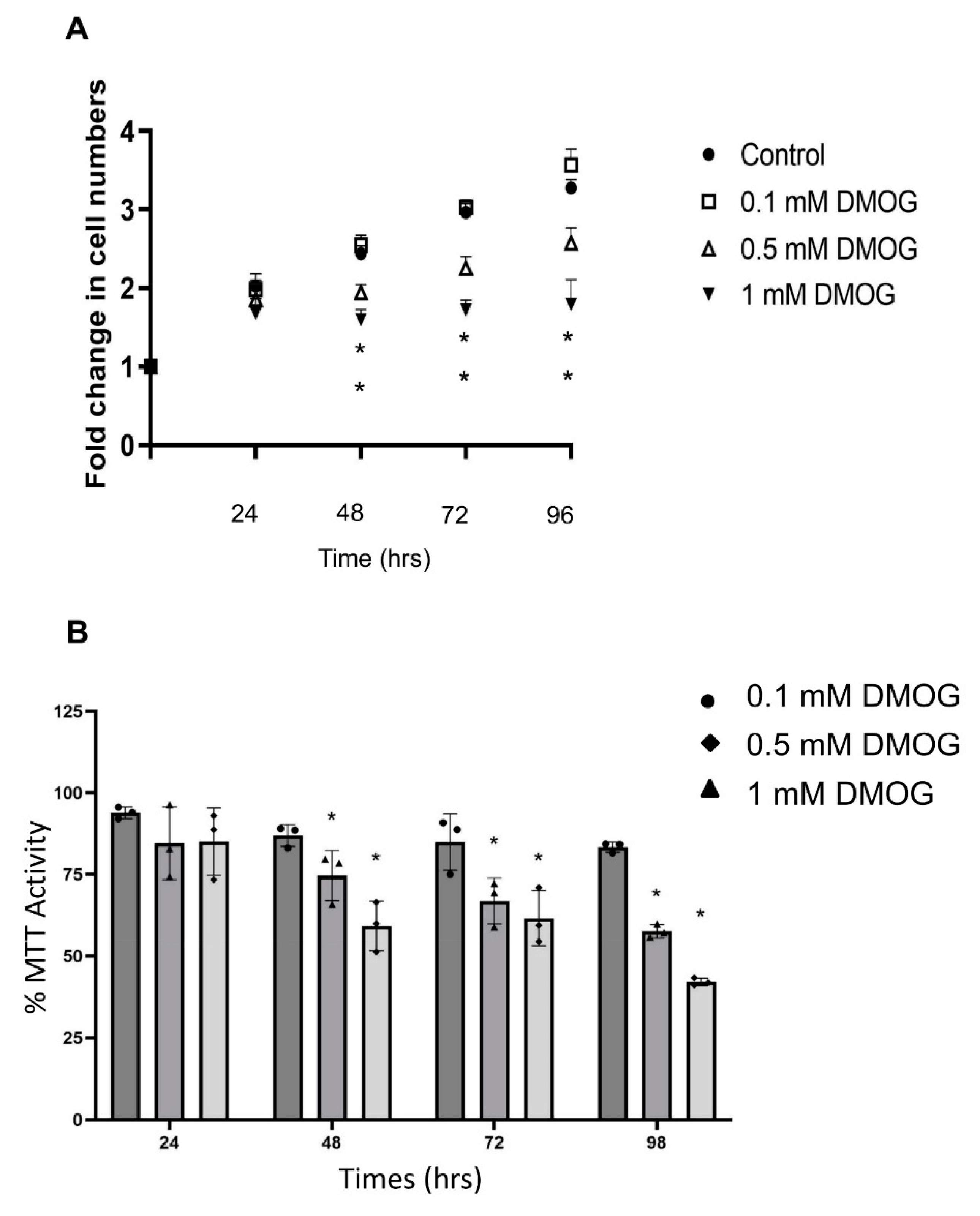
3.1. Effect of Oxygen Levels and DMOG on PC12 Cell Count and Metabolic Activity
3.2. Effect of Oxygen Levels and DMOG on PC12 Cell HIF-1 Expression
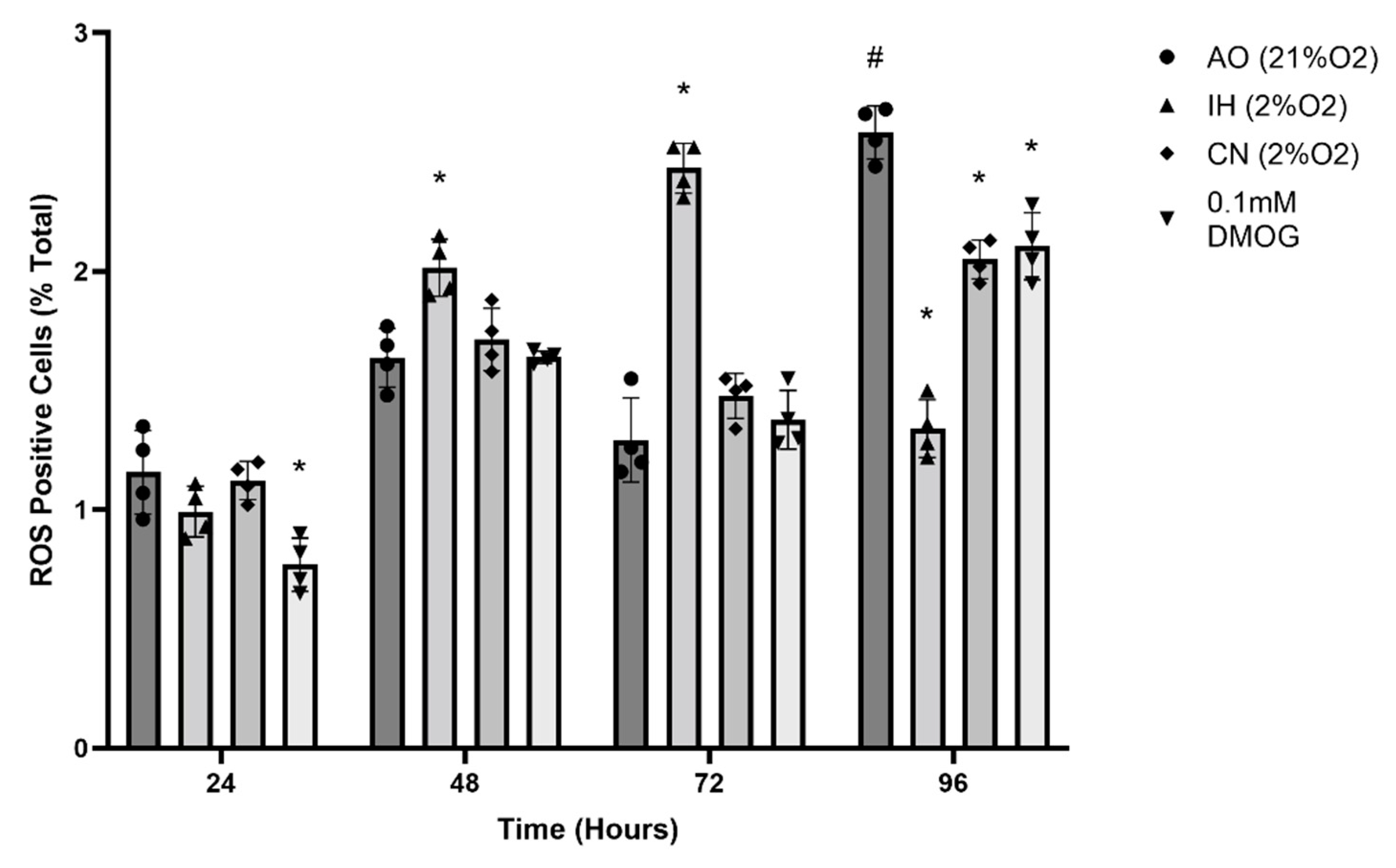
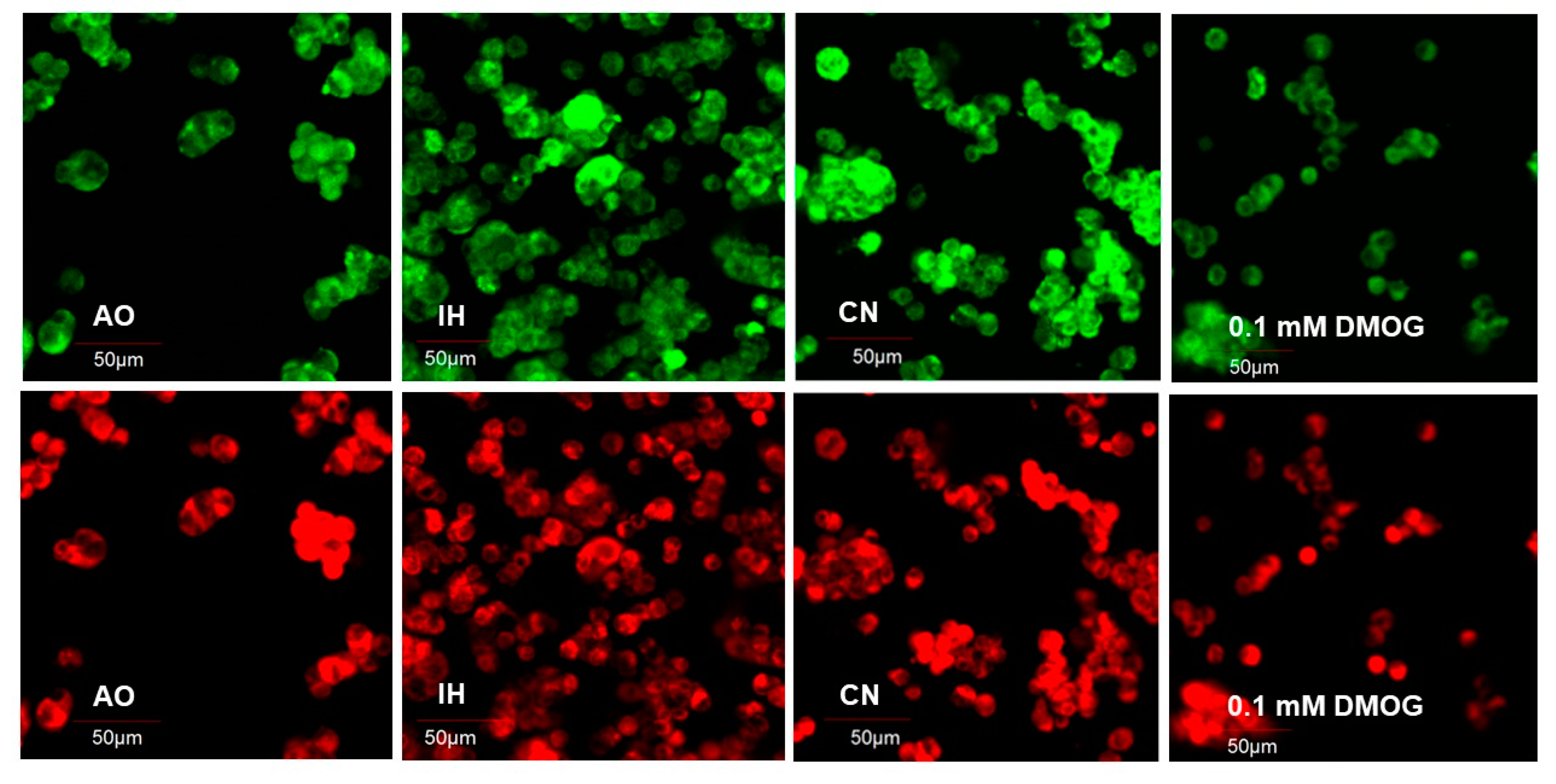
3.3. Effect of Oxygen Levels and DMOG on ROS Production
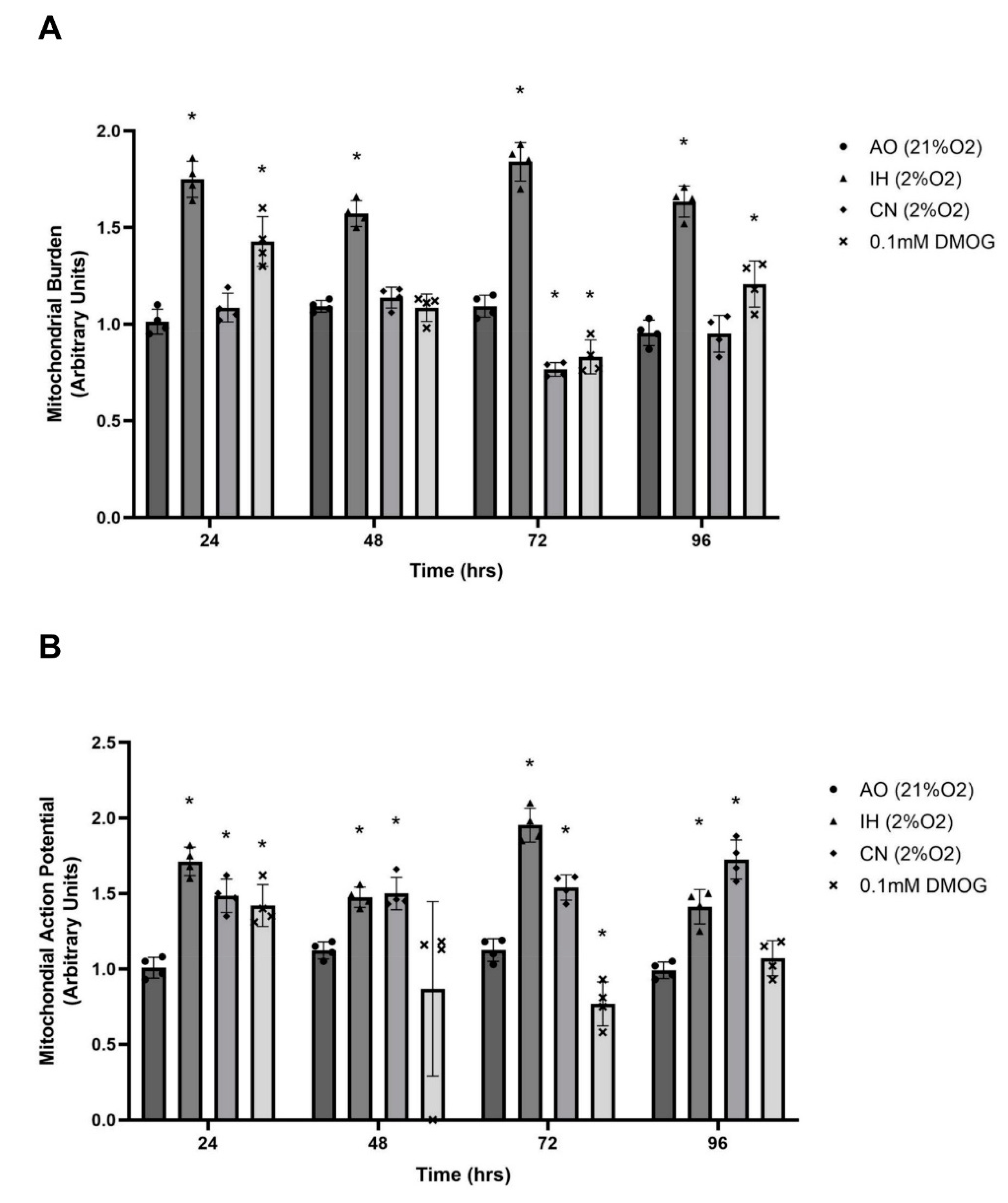
3.4. Effect of Oxygen Levels and DMOG on Mitochondrial Burden and Action Potential
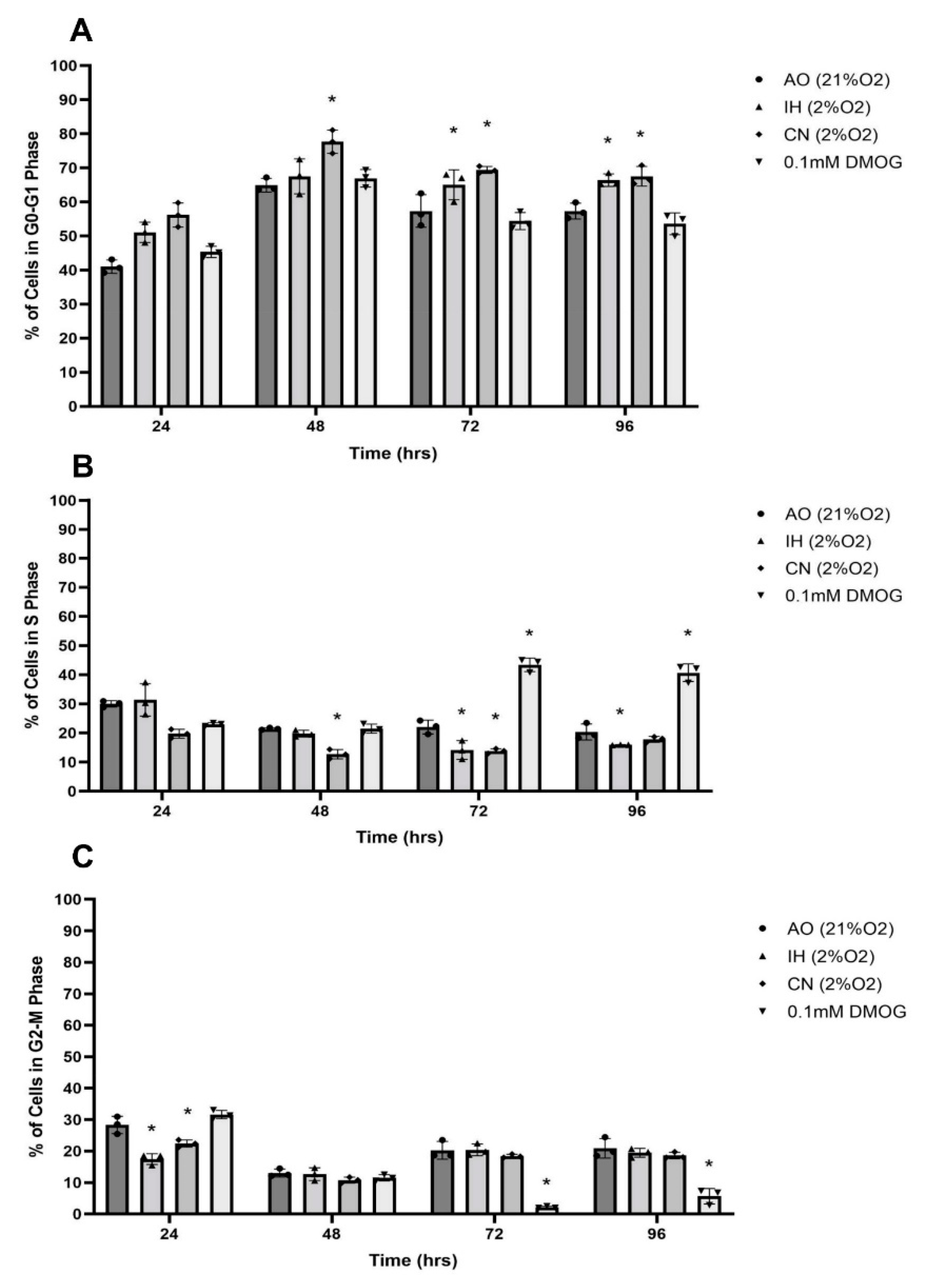
3.5. Effect of Oxygen Levels and DMOG on PC12 Cell Cycle Progression
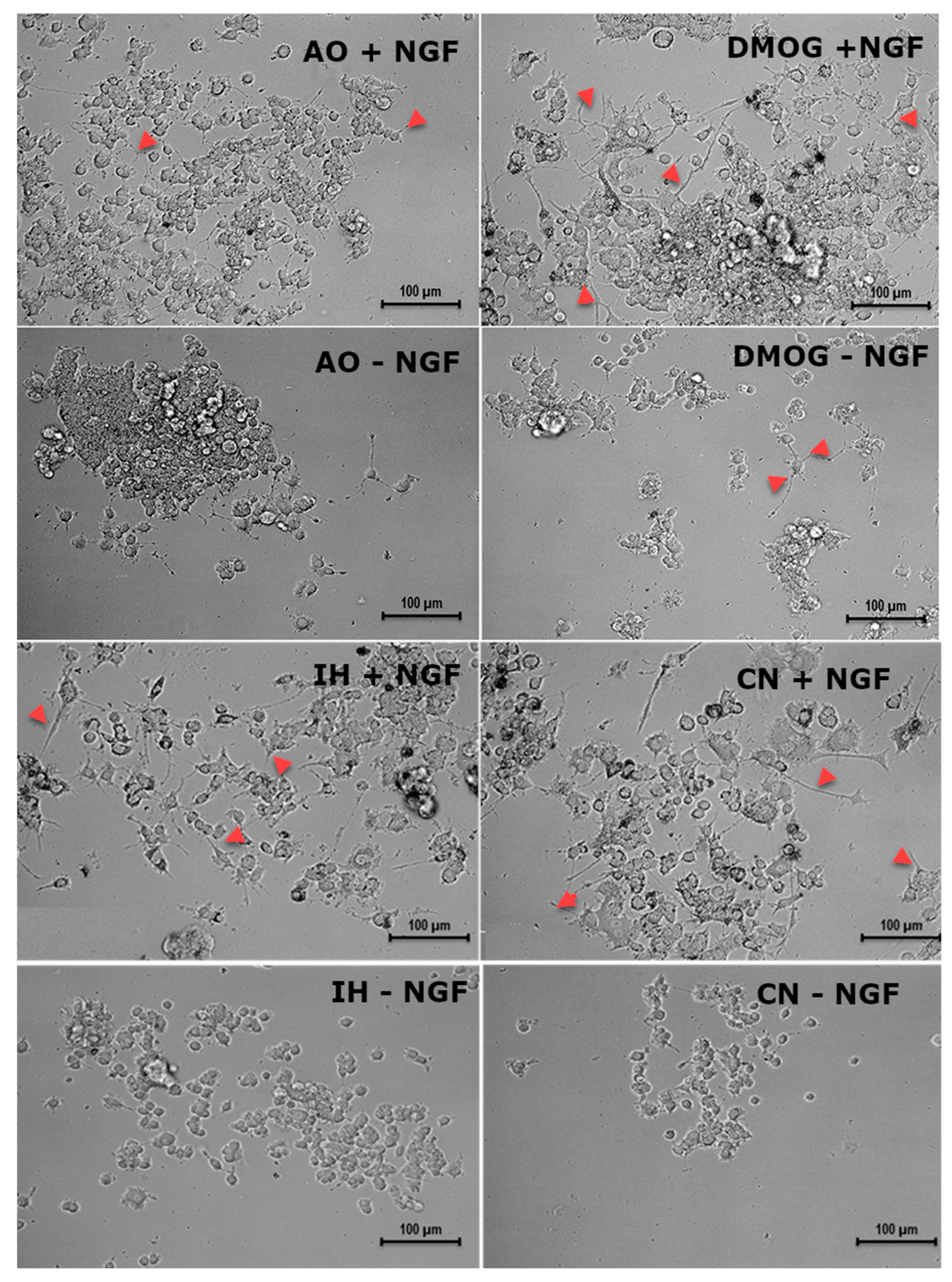
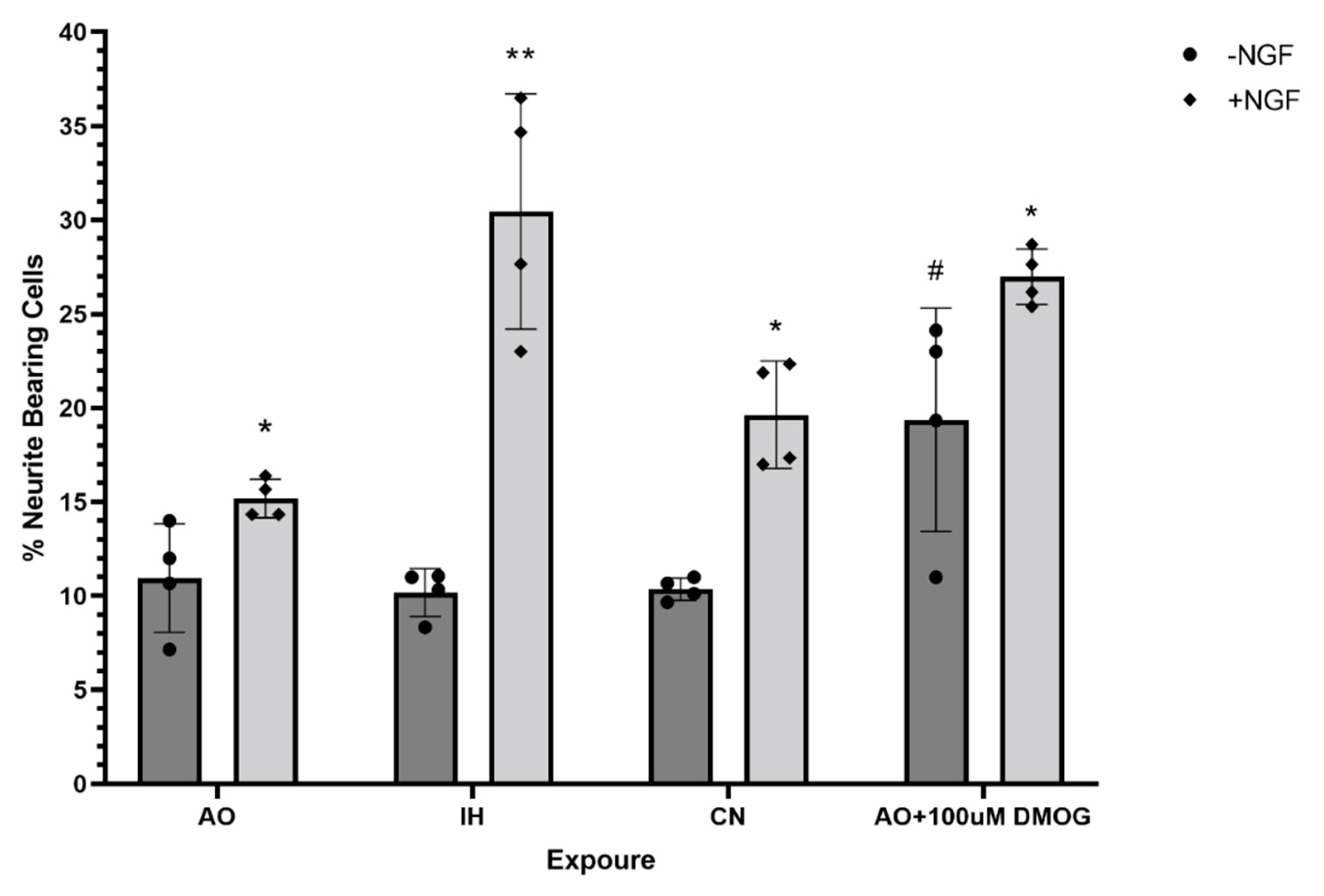
3.6. Effect of Oxygen Levels and DMOG on PC12 Cell Differentiation Capacity
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Wu, L.Y.; He, Y.L.; Zhu, L.L. Possible Role of PHD Inhibitors as Hypoxia-Mimicking Agents in the Maintenance of Neural Stem Cells’ Self-Renewal Properties. Front. Cell Dev. Biol. 2018, 6, 169. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Shen, W.B.; Yang, P.; Dong, D.; Sun, W.; Yang, P. High Glucose Inhibits Neural Stem Cell Differentiation Through Oxidative Stress and Endoplasmic Reticulum Stress. Stem. Cells Dev. 2018, 27, 745–755. [Google Scholar] [CrossRef] [Green Version]
- Dello Sbarba, P.; Cheloni, G. Tissue “hypoxia” and the maintenance of leukemia stem cells. Adv. Exp. Med. Biol. 2019, 1143, 129–145. [Google Scholar]
- Chen, R.L.; Lai, U.H.; Zhu, L.L.; Singh, A.; Ahmed, M.; Forsyth, N.R. Reactive oxygen species (ROS) formation in the brain at different oxygen Levels: Role of hypoxia inducible factors. Front. Cell Dev. Biol. 2018, 6, 132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loiacono, L.A.; Shapiro, D.S. Detection of hypoxia at the cellular level. Crit. Care Clin. 2010, 26, 409–421. [Google Scholar] [CrossRef]
- Semenza, G.L. Oxygen sensory, homeostasis and disease. N. Engl. J. Med. 2011, 365, 537–547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, R.L.; Forsyth, N.R. Development of hypoxia mimetic agents for the clinical use. Front. Cell Dev. Biol. 2019, 6, 134. [Google Scholar]
- Ratcliffe, P.J. HIF-1 and HIF-2: Working alone or together in hypoxia? J. Clin. Investig. 2007, 117, 862–865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keith, B.; Johnson, R.S.; Simon, M.C. HIF1α and HIF2α: Sibling rivalry in hypoxic tumour growth and progression. Nat. Rev. Cancer 2011, 12, 9–22. [Google Scholar] [CrossRef] [Green Version]
- Hu, C.J.; Wang, L.Y.; Chodosh, L.A.; Keith, B.; Simon, M.C. Differential roles of hypoxia-inducible factor 1alpha (HIF-1alpha) and HIF-2alpha in hypoxic gene regulation. Mol. Cell. Biol. 2003, 23, 9361–9374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koh, M.Y.; Powis, G. Passing the baton: The HIF switch. Trends Biochem. Sci. 2012, 37, 364–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, S.L.; Wu, C.; Xiong, Z.F.; Fang, X. Progress on hypoxia-inducible factor-3: Its structure, gene regulation and biological function. Mol. Med. Rep. 2015, 12, 2411–2416. [Google Scholar] [CrossRef] [Green Version]
- Chan, M.C.; Ilott, N.E.; Schödel, J.; Sims, D.; Tumber, A.; Lippl, K.; Mole, D.R.; Pugh, C.W.; Ratcliffe, P.J.; Ponting, C.P.; et al. Tuning the Transcriptional Response to Hypoxia by Inhibiting Hypoxia-inducible Factor (HIF) Prolyl and Asparaginyl Hydroxylases. J. Biol. Chem. 2006, 291, 20661–20673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, C.K.; Jain, S.A.; Bae, O.N.; Majid, A.; Rajanikant, G.K. Hypoxia mimetic agents for ischemic stroke. Front. Cell Dev. Biol. 2018, 6, 175. [Google Scholar] [CrossRef] [PubMed]
- Nagel, S.; Papadakis, M.; Chen, R.; Hoyte, L.C.; Brooks, K.J.; Gallichan, D.; Sibson, N.R.; Pugh, C.; Buchan, A. Neuroprotection by Dimethyloxalylglycine following permanent and transient focal cerebral ischemia in rats. J. Cereb. Blood Flow Metab. 2011, 31, 132–143. [Google Scholar] [CrossRef] [Green Version]
- Ogle, M.E.; Gu, X.; Espinera, A.R.; Wei, L. Inhibition of prolyl hydroxylases by dimethyloxaloylglycine after stroke reduces ischemic brain injury and requires hypoxia inducible factor-1[alpha]. Neurobiol. Dis. 2012, 45, 733. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.; Wilson, J.W.; Schofiled, C.J.; Chen, R.L. Hypoxia-inducible factor (HIF) prolyl hydroxylase inhibitors induce autophagy and have a protective effect in an in-vitro ischaemia model. Nat. Sci. Rep. 2020, 10, 1597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaudry, D.; Stork, P.J.S.; Lazarovici, P.; Eiden, L.E. Signaling Pathways for PC12 Cell Differentiation: Making the Right Connections. Science 2002, 296, 1648–1649. [Google Scholar] [CrossRef]
- Hu, R.; Cao, Q.; Sun, Z.; Chen, J.; Zheng, Q.; Xiao, F. A novel method of neural differentiation of PC12 cells by using Opti-MEM as a basic induction medium. Int. J. Mol. Med. 2018, 41, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Kay, A.G.; Dale, T.P.; Akram, K.M.; Mohan, P.; Hampson, K.; Maffulli, N.; Spiteri, M.A.; El Haj, A.J.; Forsyth, N.R. BMP2 repression and optimized culture conditions promote human bone marrow-derived mesenchymal stem cell isolation. Regen. Med. 2015, 10, 109–125. [Google Scholar] [CrossRef] [PubMed]
- Luan, Y.; Zhang, X.; Zhang, Y.; Dong, Y. MicroRNA-210 protects PC-12 cells against hypoxia-induced injury by targeting BNIP3. Front. Cell Neurosci. 2017, 11, 285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, M.; Sun, H.; Pang, J.; Guo, X.; Huo, Y.; Wu, X.; Ma, J. Propofol alleviates hypoxia-induced nerve injury in PC-12 cells by up-regulation of microRNA-153. BMC Anesthesiol. 2018, 18, 197. [Google Scholar] [CrossRef]
- Yan, R.; Tian, H.; Du, Z. Quercetin protects PC-12 cells against hypoxia injury by down-regulation of miR-122. Iran. J. Basic Med. Sci. 2019, 22, 391–398. [Google Scholar] [PubMed]
- Ding, H.; Gao, Y.S.; Wang, Y.; Hu, C.; Sun, Y.; Zhang, C. Dimethyloxaloylglycine increases the bone healing capacity of adipose-derived stem cells by promoting osteogenic differentiation and angiogenic potential. Stem. Cells Dev. 2014, 23, 990–1000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menon, A.; Creo, P.; Piccoli, M.; Bergante, S.; Conforti, E.; Banfi, G.; Randelli, P.; Anastasia, L. Chemical activation of the hypoxia-inducible factor reversibly reduces tendon stem cell proliferation, inhibits their differentiation, and maintains cell undifferentiation. Stem. Cells Int. 2018, 2018, 9468085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rose, N.R.; McDonough, M.A.; King, O.N.; Kawamura, A.; Schofield, C.J. Inhibition of 2-oxoglutarate dependent oxygenases. Chem. Soc. Rev. 2011, 40, 4364–4397. [Google Scholar] [CrossRef] [PubMed]
- Zhdanov, A.V.; Okkelman, I.A.; Collins, F.W.; Melgar, S.; Papkovsky, D.B. A novel effect of DMOG on cell metabolism: Direct inhibition of mitochondrial function precedes HIF target gene expression. Biochim. Biophys. Acta-Bioenerg. 2015, 1847, 1254–1266. [Google Scholar] [CrossRef] [Green Version]
- Duran, R.V.; MacKenzie, E.D.; Boulahbel, H.; Frezza, C.; Heiserich, L.; Tardito, S.; Bussolati, O.; Rocha, S.; Hall, M.N.; Gottlieb, E. HIF-independent role of prolyl hydroxylases in the cellular response to amino acids. Oncogene. 2013, 32, 4549–4556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, L.E.; Arany, Z.; Livingston, D.M.; Bunn, H.F. Activation of hypoxia-inducible transcription factor depends primarily upon redox-sensitive stabilization of its alpha subunit. J. Biol. Chem. 1996, 271, 32253–32259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uehlinger, F.D.; Barkema, H.W.; O’Handley, R.M.; Parenteau, M.; Parrington, L.J.; Vanleeuwen, J.A.; Dixon, B.R. Comparison of flow cytometry and immunofluorescence microscopy for the detection of Giardia duodenalis in bovine fecal samples. J. Vet. Diagn. Investig. 2008, 20, 178–185. [Google Scholar] [CrossRef] [Green Version]
- Junga, K.M.; Baea, I.H.; Kimb, B.H.; Kima, W.K.; Chungc, J.H.; Parka, Y.H.; Lima, K.M. Comparison of flow cytometry and immunohistochemistry in non-radioisotopic murine lymph node assay using bromodeoxyuridine. Toxicol. Lett. 2010, 192, 229–237. [Google Scholar]
- Ramberger, M.; Peschl, P.; Schanda, K.; Irschick, R.; Höftberger, R.; Deisenhammer, F.; Rostásy, K.; Berger, T.; Dalmau, J.; Reindl, M. Comparison of Diagnostic Accuracy of Microscopy and Flow Cytometry in Evaluating N-Methyl-D-Aspartate Receptor Antibodies in Serum Using a Live Cell-Based Assay. PLoS ONE 2015, 10, e0122037. [Google Scholar] [CrossRef]
- Tran, D.N.; Smith, S.A.; Brown, D.A.; Parker, A.J.; Joseph, J.E.; Armstrong, N.; Sewell, W.A. Polychromatic flow cytometry is more sensitive than microscopy in detecting small monoclonal plasma cell populations. Cytom. Part B Clin. Cytom. 2017, 92, 136–144. [Google Scholar] [CrossRef] [Green Version]
- Clambey, E.T.; McNamee, E.N.; Westrich, J.A.; Glover, L.E.; Campbell, E.L.; Jedlicka, P.; de Zoeten, E.F.; Cambier, J.C.; Stenmark, K.R.; Eltzschig, H.K.; et al. Hypoxia-inducible factor-1 alpha–dependent induction of FoxP3 drives regulatory T-cell abundance and function during inflammatory hypoxia of the mucosa. Proc. Natl. Acad. Sci USA 2012, 109, E2784–E2793. [Google Scholar] [CrossRef] [Green Version]
- Huang, R.Y.; Yan, X. HIF1A Is a Critical Downstream Mediator for Hemophagocytic Lymphohistiocytosis. Haematologica 2017, 102, 1956–1968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.; Li, D.; Zhang, X.; Wan, Q.; Zhang, W.; Zheng, M.; Liu, Y.C. The E3 ligase VHL promotes group 2 innate lymphoid cell maturation and function via inhibition of glycolysis and induction of interleukin 33 receptor. Immunity 2018, 48, 258–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tahara, E.B.; Navarete, F.D.; Kowaltowski, A.J. Tissue-, substrate-, and site-specific characteristics of mitochondrial reactive oxygen species generation. Free Radic. Biol. Med. 2009, 46, 1283–1297. [Google Scholar] [CrossRef] [PubMed]
- Meo, S.D.; Reed, T.T.; Venditti, P.; Victor, V.M. Role of ROS and RNS sources in physiological and pathological conditions. Oxidative Med. Cell. Longev. 2016, 2016, 1245049. [Google Scholar]
- Turovsky, E.A.; Varlamova, E.G.; Plotnikov, E.Y. Mechanisms Underlying the Protective Effect of the Peroxiredoxin-6 Are Mediated via the Protection of Astrocytes during Ischemia/Reoxygenation. Int. J. Mol. Sci. 2021, 22, 8805. [Google Scholar] [CrossRef]
- Stuart, J.A.; Fonseca, J.; Moradi, F.; Cunningham, C.; Seliman, B.; Worsfold, C.R.; Dolan, S.; Abando, J.; Maddalena, L.A. How Supraphysiological Oxygen Levels in Standard Cell Culture Affect Oxygen-Consuming Reactions. Oxidative Med. Cell. Longev. 2018, 2018, 8238459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bell, E.L.; Klimova, T.A.; Eisenbart, J.; Moraes, C.T.; Murphy, M.P.; Budinger, G.S.; Chandel, N.S. The Q o site of the mitochondrial complex III is required for the transduction of hypoxic signaling via reactive oxygen species production. J. Cell Biol. 2007, 177, 1029–1036. [Google Scholar] [CrossRef] [Green Version]
- Abramov, A.Y.; Scorziello, A.; Duchen, M.R. Three distinct mechanisms generate oxygen free radicals in neurons and contribute to cell death during anoxia and reoxygenation. J. Neurosci. 2007, 27, 1129–1138. [Google Scholar] [CrossRef]
- Turovskaya, M.V.; Gaidin, S.G.; Mal’tseva, V.N.; Zinchenko, V.P.; Turovsky, E.A. Taxifolin protects neurons against ischemic injury in vitro via the activation of antioxidant systems and signal transduction pathways of GABAergic neurons. Mol. Cell. Neurosci. 2019, 96, 10–24. [Google Scholar] [CrossRef] [PubMed]
- Agani, F.H.; Pichiule, P.; Chavez, J.C.; LaManna, J.C. The role of mitochondria in the regulation of hypoxia-inducible factor 1 expression during hypoxia. J. Biol. Chem. 2000, 275, 35863–35867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Granger, D.N.; Kvietys, P.R. Reperfusion injury and reactive oxygen species: The evolution of a concept. Redox Biol. 2015, 6, 524–551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wheaton, W.W.; Chandel, N.S. Hypoxia regulates cellular metabolism. Am. J. Physiol. Cell Physiol. 2011, 300, C385–C393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, T.I.; Chiu, H.W.; Pan, Y.C.; Hsu, S.T.; Lin, J.H.; Yang, K.T. Intermittent hypoxia-induced protein phosphatase 2A activation reduces PC12 cell proliferation and differentiation. J. Biomed. Sci. 2014, 21, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bigarella, C.L.; Liang, R.; Ghaffari, S. Stem cells and the impact of ROS signalling. Development 2014, 141, 4206–4218. [Google Scholar] [CrossRef] [Green Version]
- Sena, L.A.; Chandel, N.S. Physiological roles of mitochondrial reactive oxygen species. Mol. Cell. 2012, 48, 158–167. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, E.; Sato, M. Anaerobic respiration sustains mitochondrial membrane potential in a prolyl hydroxylase pathway-activated cancer cell line in a hypoxic microenvironment. Am. J. Physiol. Cell Physiol. 2014, 306, C334–C342. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Mu, J.; Chen, L.; Feng, J.; Hu, J.; Li, L.; Zhou, J.-M.; Zuo, J. S nitrosylation positively regulates ascorbate peroxidase activity during plant stress responses. Plant physiol. 2015, 167, 1604–1615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, Y.; Agani, F.H. Oligomycin inhibits HIF-1alpha expression in hypoxic tumor cells. Am. J. Physiol. Cell Physiol. 2005, 288, C1023–C1029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hagen, T. Oxygen versus Reactive Oxygen in the Regulation of HIF-1α: The Balance Tips. Biochem. Res. Int. 2012, 2012, 436981. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Diez, M.; Santamaría, G.; Ortega, Á.D.; Cuezva, J.M. Biogenesis and Dynamics of Mitochondria during the Cell Cycle: Significance of 3′UTRs’. PLoS ONE 2006, 1, e107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Li, Z.; Lu, Y.; Du, R.; Katiyar, S.; Yang, J.; Fu, M.; Leader, J.E.; Quong, A.; Novikoff, P.M.; et al. Cyclin D1 repression of nuclear respiratory factor 1 integrates nuclear DNA synthesis and mitochondrial function. Proc. Natl. Acad. Sci USA 2016, 103, 11567–11572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avramovich-Tirosh, Y.; Amit, T.; Bar-Am, O.; Zheng, H.; Fridkin, M.; Youdim, M.B. Therapeutic targets and potential of the novel brain- permeable multifunctional iron chelator monoamine oxidase inhibitor drug, M-30, for the treatment of Alzheimer’s disease. J. Neurochem. 2007, 100, 490–502. [Google Scholar] [CrossRef] [PubMed]
- Zheng, K.; He, Z.; Kitazato, K.; Wang, Y. Selective Autophagy Regulates Cell Cycle in Cancer Therapy. Theranostics 2019, 9, 104–125. [Google Scholar] [CrossRef]
- Ortmann, B.; Druker, J.; Rocha, S. Cell cycle progression in response to oxygen levels. Cell. Mol. Life Sci. 2014, 71, 3569–3582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gordan, J.D.; Bertovrt, J.A.; Hu, C.J.; Diehl, J.A.; Simon, M.C. HIF-2α promotes hypoxic cell proliferation by enhancing c-Myc transcriptional activity. Cancer Cell 2007, 11, 335–347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gordan, J.D.; Thompson, C.B.; Simon, M.C. HIF and c-Myc: Sibling rivals for control of cancer cell metabolism and proliferation. Cancer Cell 2007, 12, 108–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medina, P.P.; Slack, F.J. microRNAs and cancer: An overview. Cell Cycle 2008, 7, 2485–2492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammond, E.M.; Dorie, M.J.; Giaccia, A.J. ATR/ATM targets are phosphorylated by ATR in response to hypoxia and ATM in response to reoxygenation. J. Biol. Chem. 2003, 278, 12207–12213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Genetos, D.C.; Cheung, W.K.; Decaris, M.L.; Leach, J.K. Oxygen tension modulates neurite outgrowth in PC12 cells through a mechanism involving HIF and VEGF. J. Mol. Neurosci. 2010, 40, 360–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Driscoll, C.M.; Gorman, A.M. Hypoxia induces neurite outgrowth in PC12 cells that is mediated through adenosine A2A receptors. Neuroscience 2005, 131, 321–329. [Google Scholar] [CrossRef] [PubMed]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, R.; Ahmed, M.A.; Forsyth, N.R. Dimethyloxalylglycine (DMOG), a Hypoxia Mimetic Agent, Does Not Replicate a Rat Pheochromocytoma (PC12) Cell Biological Response to Reduced Oxygen Culture. Biomolecules 2022, 12, 541. https://doi.org/10.3390/biom12040541
Chen R, Ahmed MA, Forsyth NR. Dimethyloxalylglycine (DMOG), a Hypoxia Mimetic Agent, Does Not Replicate a Rat Pheochromocytoma (PC12) Cell Biological Response to Reduced Oxygen Culture. Biomolecules. 2022; 12(4):541. https://doi.org/10.3390/biom12040541
Chicago/Turabian StyleChen, RuoLi, Mohammad Alkataan Ahmed, and Nicholas Robert Forsyth. 2022. "Dimethyloxalylglycine (DMOG), a Hypoxia Mimetic Agent, Does Not Replicate a Rat Pheochromocytoma (PC12) Cell Biological Response to Reduced Oxygen Culture" Biomolecules 12, no. 4: 541. https://doi.org/10.3390/biom12040541