
Use of Bacterial Extracellular Vesicles for Gene Delivery to Host Cells



{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Culture
2.2. Isolation and Analysis of bEVs
2.3. Cell Viability Analysis after Treatment with bEVs
2.4. Vector and RNA Preparation
2.5. Electroporation of Plasmid and RNA into bEVs
2.6. RNA Immunoprecipitation
2.7. Reverse Transcription-Quantitative PCR (RT-qPCR)
2.8. Confocal Microscopy Analysis
2.9. Cel-miR-39 Precursor RNA Synthesis
2.10. Transmission Electron Microscopy (TEM)
2.11. Statistical Analysis
3. Results
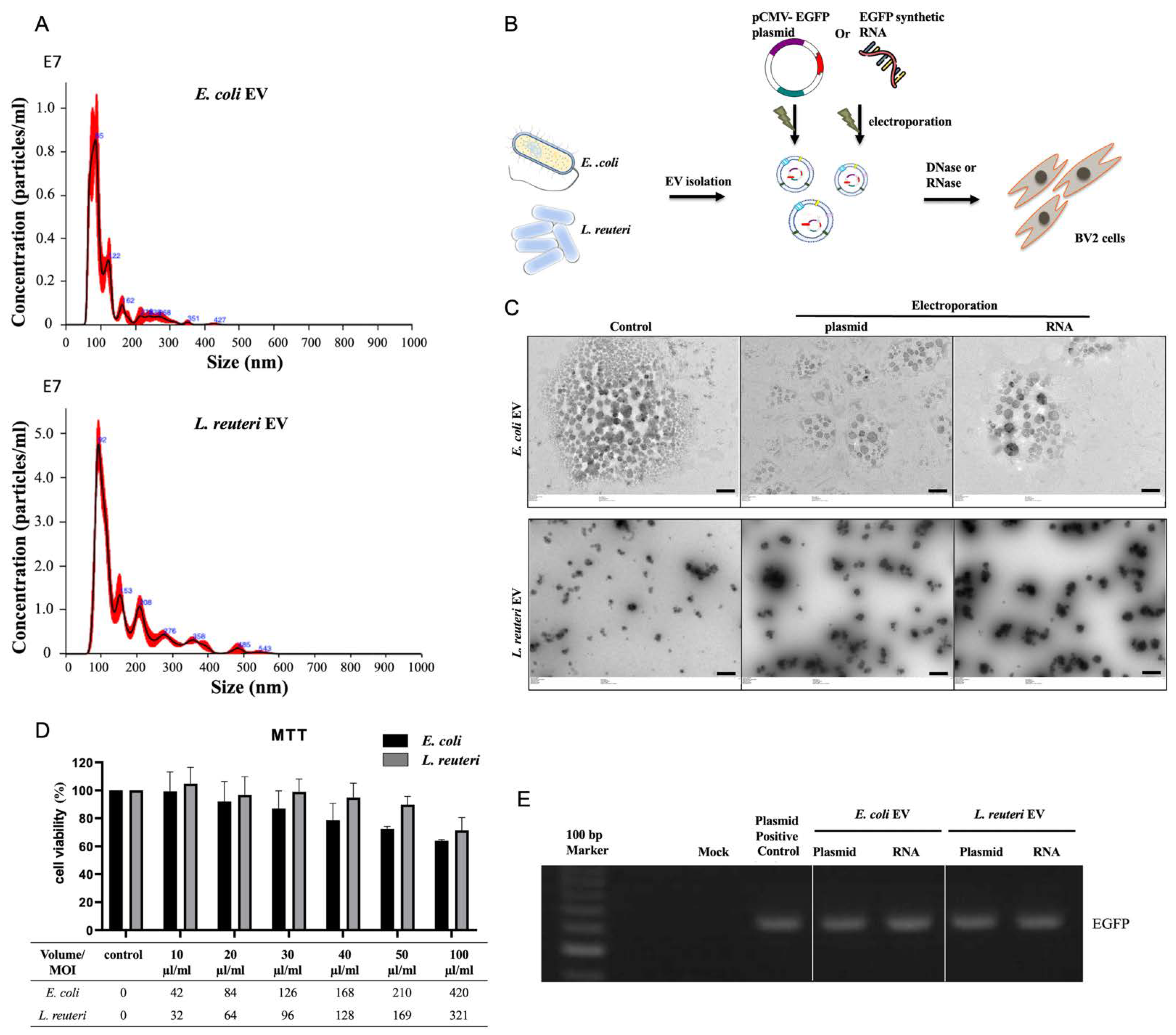
3.1. bEV Analysis and BV2 Cell Viability Assay
3.2. Transfer of EGFP DNA and RNA via bEVs
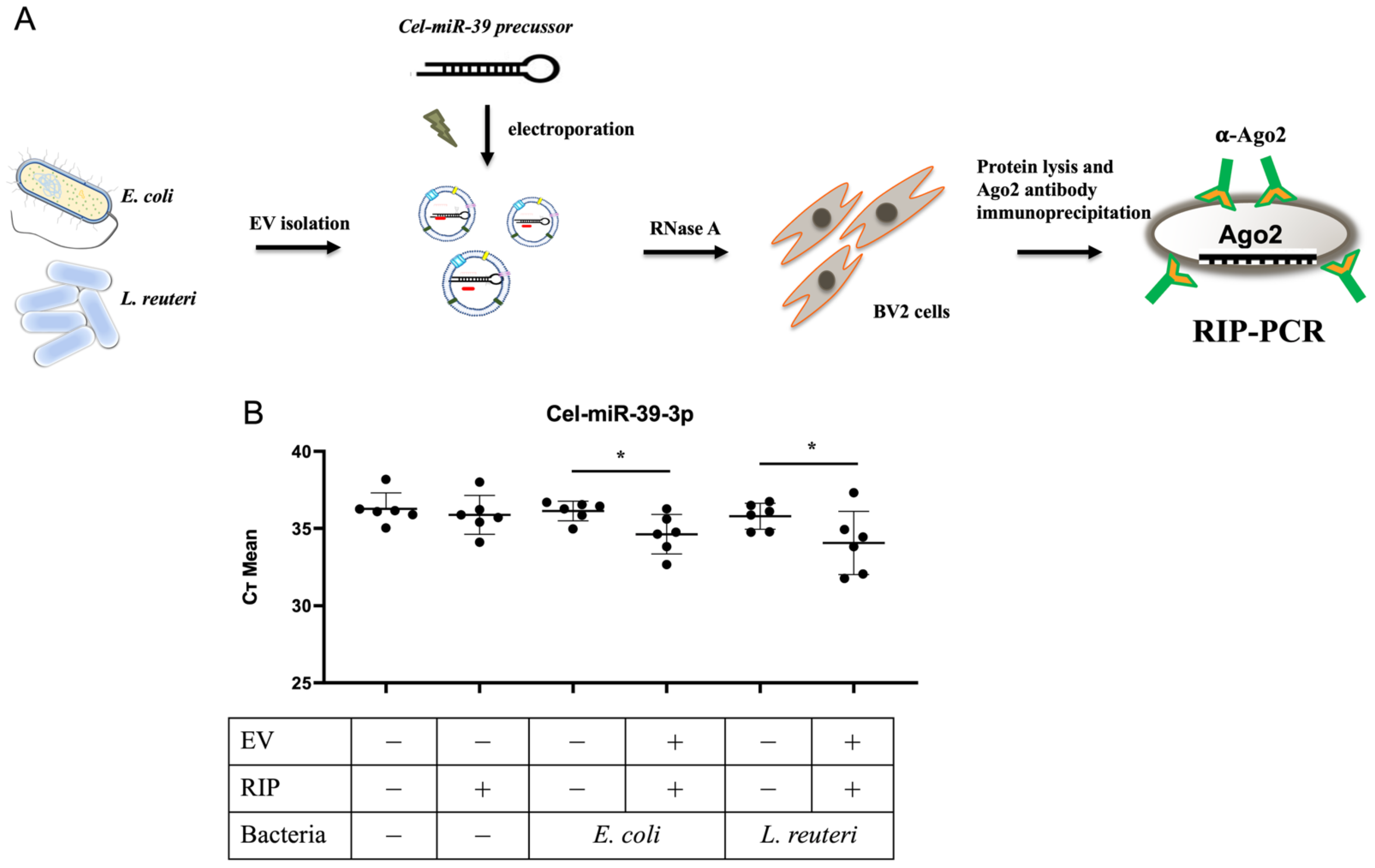
3.3. Exogenous Maturation of Pre-Cel-miR-39 and Incorporation into Ago2 in the Host Cells via bEVs
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gebre, M.S.; Brito, L.A.; Tostanoski, L.H.; Edwards, D.K.; Carfi, A.; Barouch, D.H. Novel Approaches for Vaccine Development. Cell 2021, 184, 1589–1603. [Google Scholar] [CrossRef] [PubMed]
- Kheirollahpour, M.; Mehrabi, M.; Dounighi, N.M.; Mohammadi, M.; Masoudi, A. Nanoparticles and Vaccine Development. Pharm. Nanotechnol. 2020, 8, 6–21. [Google Scholar] [CrossRef] [PubMed]
- Briaud, P.; Carroll, R.K. Extracellular Vesicle Biogenesis and Functions in Gram-Positive Bacteria. Infect. Immun. 2020, 88, e00433-20. [Google Scholar] [CrossRef] [PubMed]
- Kulp, A.; Kuehn, M.J. Biological Functions and Biogenesis of Secreted Bacterial Outer Membrane Vesicles. Annu. Rev. Microbiol. 2010, 64, 163–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, Y.; Yin, S.; Wang, M. Extracellular Vesicles of Bacteria as Potential Targets for Immune Interventions. Hum. Vaccin. Immunother. 2020, 17, 897–903. [Google Scholar] [CrossRef] [PubMed]
- Cheng, K.; Zhao, R.; Li, Y.; Qi, Y.; Wang, Y.; Zhang, Y.; Qin, H.; Qin, Y.; Chen, L.; Li, C.; et al. Bioengineered Bacteria-Derived Outer Membrane Vesicles as a Versatile Antigen Display Platform for Tumor Vaccination via Plug-and-Display Technology. Nat. Commun. 2021, 12, 2041. [Google Scholar] [CrossRef] [PubMed]
- Melling, G.E.; Carollo, E.; Conlon, R.; Simpson, J.C.; Carter, D.R.F. The Challenges and Possibilities of Extracellular Vesicles as Therapeutic Vehicles. Eur. J. Pharm. Biopharm. 2019, 144, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Sartorio, M.G.; Pardue, E.J.; Feldman, M.F.; Haurat, M.F. Bacterial Outer Membrane Vesicles: From Discovery to Applications. Annu. Rev. Microbiol. 2021, 75, 609–630. [Google Scholar] [CrossRef] [PubMed]
- Ñahui Palomino, R.A.; Vanpouille, C.; Costantini, P.E.; Margolis, L. Microbiota–Host Communications: Bacterial Extracellular Vesicles as a Common Language. PLoS Pathog. 2021, 17, e1009508. [Google Scholar] [CrossRef]
- Grande, R.; Celia, C.; Mincione, G.; Stringaro, A.; Di Marzio, L.; Colone, M.; Di Marcantonio, M.C.; Savino, L.; Puca, V.; Santoliquido, R.; et al. Detection and Physicochemical Characterization of Membrane Vesicles (MVs) of Lactobacillus Reuteri DSM 17938. Front. Microbiol. 2017, 8, 1040. [Google Scholar] [CrossRef] [PubMed]
- Ha, J.Y.; Choi, S.-Y.; Lee, J.H.; Hong, S.-H.; Lee, H.-J. Delivery of Periodontopathogenic Extracellular Vesicles to Brain Monocytes and Microglial IL-6 Promotion by RNA Cargo. Front. Mol. Biosci. 2020, 7, 596366. [Google Scholar] [CrossRef] [PubMed]
- Han, E.-C.; Choi, S.-Y.; Lee, Y.; Park, J.-W.; Hong, S.-H.; Lee, H.-J. Extracellular RNAs in Periodontopathogenic Outer Membrane Vesicles Promote TNF-α Production in Human Macrophages and Cross the Blood-Brain Barrier in Mice. FASEB J. 2019, 33, 13412–13422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, S.-Y.; Han, E.-C.; Hong, S.-H.; Kwon, T.-G.; Lee, Y.; Lee, H.-J. Regulating Osteogenic Differentiation by Suppression of Exosomal MicroRNAs. Tissue Eng. Part A 2019, 25, 1146–1154. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, K.; Breyne, K.; Ughetto, S.; Laurent, L.C.; Breakefield, X.O. RNA Delivery by Extracellular Vesicles in Mammalian Cells and Its Applications. Nat. Rev. Mol. Cell Biol. 2020, 21, 585–606. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-J. Exceptional Stories of MicroRNAs. Exp. Biol. Med. 2013, 238, 339–343. [Google Scholar] [CrossRef] [PubMed]
- Kahn, M.E.; Barany, F.; Smith, H.O. Transformasomes: Specialized Membranous Structures That Protect DNA during Haemophilus Transformation. Proc. Natl. Acad. Sci. USA 1983, 80, 6927–6931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bittel, M.; Reichert, P.; Sarfati, I.; Dressel, A.; Leikam, S.; Uderhardt, S.; Stolzer, I.; Phu, T.A.; Ng, M.; Vu, N.K.; et al. Visualizing Transfer of Microbial Biomolecules by Outer Membrane Vesicles in Microbe-host-communication in Vivo. J. Extracell. Vesicles 2021, 10, e12159. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-J. Microbe-Host Communication by Small RNAs in Extracellular Vesicles: Vehicles for Transkingdom RNA Transportation. Int. J. Mol. Sci. 2019, 20, 1487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stanton, B.A. Extracellular Vesicles and Host-Pathogen Interactions: A Review of Inter-Kingdom Signaling by Small Noncoding RNA. Genes 2021, 12, 1010. [Google Scholar] [CrossRef] [PubMed]
- Pardi, N.; Hogan, M.J.; Weissman, D. Recent Advances in MRNA Vaccine Technology. Curr. Opin. Immunol. 2020, 65, 14–20. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, S.-I.; Ha, J.Y.; Choi, S.-Y.; Hong, S.-H.; Lee, H.-J. Use of Bacterial Extracellular Vesicles for Gene Delivery to Host Cells. Biomolecules 2022, 12, 1171. https://doi.org/10.3390/biom12091171
Kim S-I, Ha JY, Choi S-Y, Hong S-H, Lee H-J. Use of Bacterial Extracellular Vesicles for Gene Delivery to Host Cells. Biomolecules. 2022; 12(9):1171. https://doi.org/10.3390/biom12091171
Chicago/Turabian StyleKim, Su-Im, Jae Yeong Ha, Song-Yi Choi, Su-Hyung Hong, and Heon-Jin Lee. 2022. "Use of Bacterial Extracellular Vesicles for Gene Delivery to Host Cells" Biomolecules 12, no. 9: 1171. https://doi.org/10.3390/biom12091171