
The Dual Role of Small Extracellular Vesicles in Joint Osteoarthritis: Their Global and Non-Coding Regulatory RNA Molecule-Based Pathogenic and Therapeutic Effects
Abstract
:1. Introduction
2. Origin of sEV Therapy: Stem Cell Therapy for Osteoarthritis
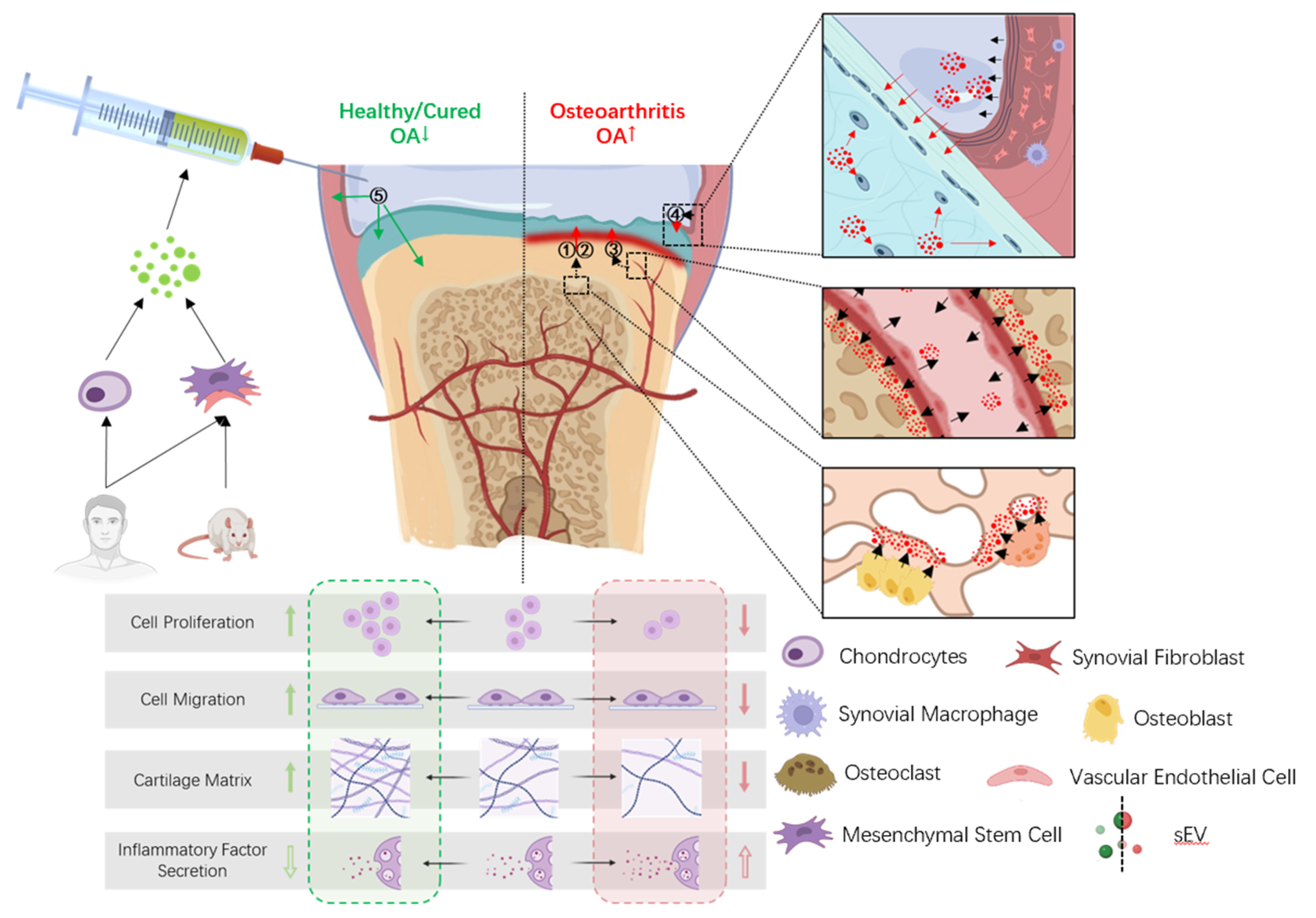
3. Effects of sEVs on Articular Tissues in OA
3.1. Pathogenic sEVs in OA
3.1.1. Effects of Pathogenic sEVS on Chondrocytes
3.1.2. Effects of Pathogenic sEVS on Cartilage Extracellular Matrix
3.1.3. Effects of Pathogenic sEVS on Synovial Tissue
3.2. Therapeutic sEVs in OA
3.2.1. Effects of Therapeutic sEVS on Chondrocytes
3.2.2. Effects of Therapeutic sEVS on Cartilage Extracellular Matrix (ECM)
3.2.3. Effects of Therapeutic sEVS on Subchondral Bone
3.2.4. Effects of Therapeutic sEVS on Synovial Tissue
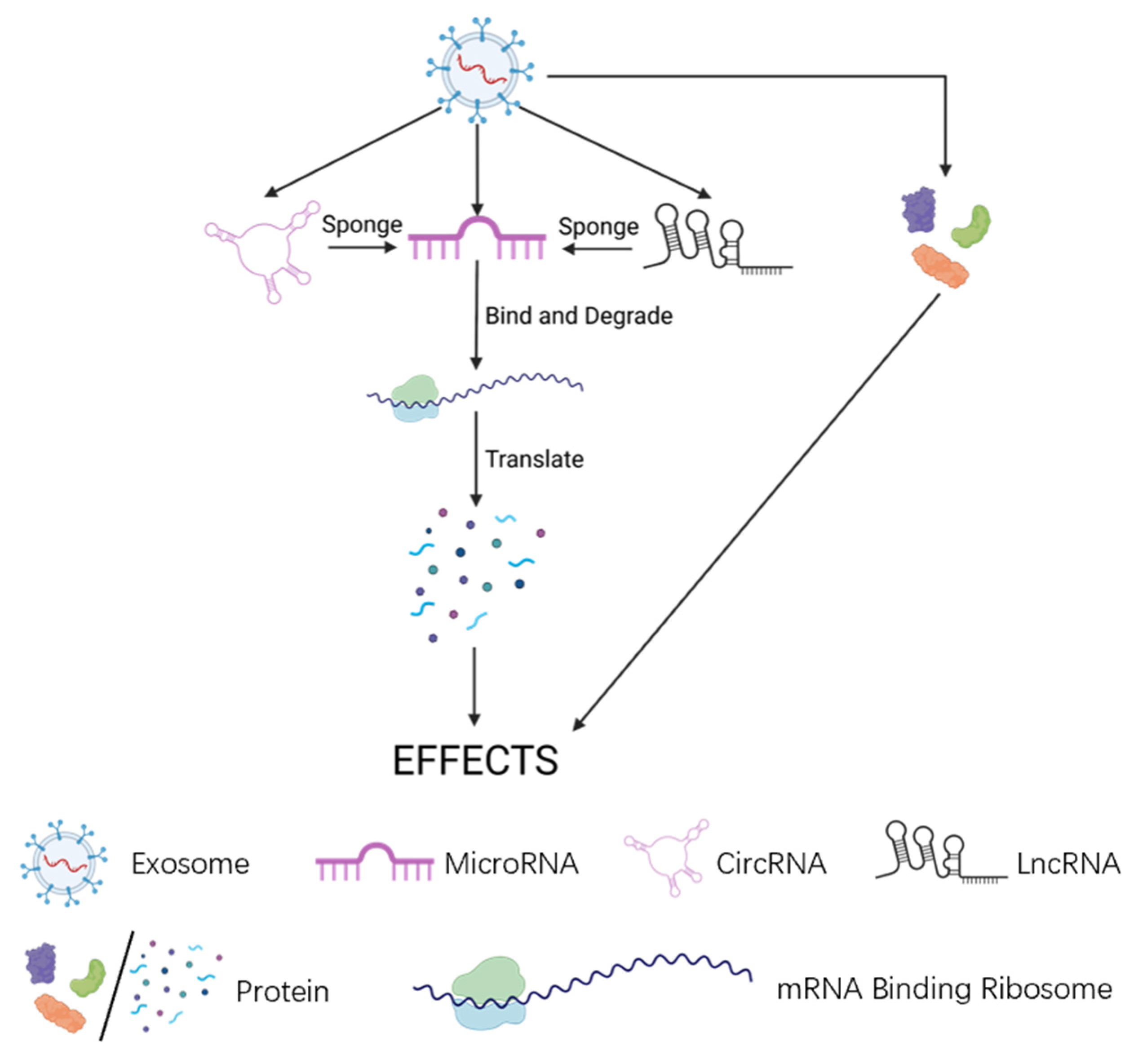
4. Direct Bearers of Sev Effects on OA: Non-coding Regulatory RNA Molecule
4.1. MicroRNAs (miRNAs)

{kind=link}
{kind=link}
| MicroRNA | Origin of sEV | Mechanism | Effects | Species | Ref. |
|---|---|---|---|---|---|
| miR-9-5p | rat BM-MSCs | Negatively regulates SDC1 | Reduces inflammation in OA cartilage (decreases IL-1, IL-6, TNF-α, CRP, NO, MDA, iNOS, COX2, SOD, OCN, MMP13, COMP, AKP) | rat | [15] |
| miR-92a-3p | human BM-MSCs | Negatively regulates WNT5A | Increases cell proliferation and matrix secretion in OA cartilage (increases ACAN, COL2A1, COL9A1, COMP, SOX9; decreases COL10A1, RUNX2, MMP13) | human | [12] |
| miR-95-5p | human BM-MSCs | Negatively regulates HDAC2/8 | Increases cartilage development and cartilage matrix expression in MSCs and chondrocytes (increases ACAN, COL2A1, COL9A1, COMP; decreases COL10A1, MMP13) | human | [79] |
| miR-100-5p | human UC-MSCs | Negatively regulates NOS4 | Reduces ROS production and apoptosis in OA chondrocytes | human | [65] |
| miR-100-5p | human IPFP-MSCs | Negatively regulates mTOR | Increases autophagy and reduces apoptosis in OA chondrocytes; increases secretion in OA ECM (increases COL2; decreases MMP13, ADAMTS5) | human | [21] |
| miR-100-5p | human exfoliated deciduous teeth-SCs | Negatively regulates mTOR | Reduces inflammation in OA chondrocytes (decreases MMP1, MMP9, MMP13, ADAMTS5) | human | [70] |
| miR-124 | MSCs | Negatively regulates ROCK1, which actives TLR9 | Reduces apoptosis in OA chondrocytes | mouse | [102] |
| miR-125a-5p | human BM-MSCs | Negatively regulates E2F2 | Increases migration in OA chondrocytes and increases secretion in OA ECM (increases COL2, ACAN, SOX9; decreases MMP13) | human/mouse | [75] |
| miR-126-3p | rat SFBs | Increases migration and proliferation, reduces apoptosis and inflammation in OA chondrocytes (decreases IL-1β, IL-6, TNF-α); reduces formation of osteophytes and degeneration in cartilage | rat | [48] | |
| miR-127-3p | rat BM-MSCs | Negatively regulates CDH11, which actives Wnt/β-catenin pathway | Increases cell viability and DNA synthesis activity, and reduces apoptosis in OA chondrocytes (increases COL2; decreases MMP13) | rat | [103] |
| miR-129-5p | human S-MSCs | Negatively regulates HMGB1, which up-regulates TLR4 and then actives NF-κB signaling pathway | Reduces inflammatory response and apoptosis in OA chondrocytes (increases COL2; decreases COX2, iNOS, MMP13) | human | [17] |
| miR-135b | rat MSCs | Negatively regulates Sp1 | Increases viability and proliferation in OA chondrocytes | rat | [64] |
| miR-135b | rat BM-MSCs | Negatively regulates MAPK6 | Increases M2 polarization in synovial macrophages; increases repair of OA cartilage (increases ACAN, SOX9); reduces inflammatory factors (IL-1β, PGE2, COX-2, COX-1, NO) in OA serum | rat | [62] |
| miR-136-5p | human BM-MSCs | Negatively regulates ELF3 | Increases migration in chondrocytes; reduces degeneration in cartilage (increases COL2, ACAN, SOX9; decreases MMP13) | human/mouse | [76] |
| miR-140-5p | human DPSCs/human USCs | Negatively regulates VEGFA | Increases proliferation, migration, and reduces apoptosis in OA chondrocytes; increases secretion in OA ECM (increases COL2, ACAN, SOX9) | human/rat | [18,60] |
| miR-140-5p | human S-MSCs | Negatively regulates RalA | Increases secretion in OA ECM (increases SOX9, ACAN, and COL2) | human/rat | [13] |
| miR-143 | MSCs | Negatively regulates ROCK1, which actives NF-κB signaling pathway | Reduces apoptosis in OA chondrocytes | mouse | [102] |
| miR-145 | human ADSCs | inhibits the NF-κB signaling pathway; actives Wnt/β-catenin signaling pathway | Increases chondrogenesis (increases SOX9 in periosteal cells) and collagen deposits in OA cartilage | human | [94] |
| miR-147b | human BM-MSCs | inhibits the degradation of IκBα, then inhibits the NF-κB signaling pathway | Reduces inflammation in OA synovial cells (increases SOCS3, SOCS6; decreases IL-1β, IL-6, monocyte chemoattractant protein-1) | human | [96] |
| miR-155-5p | human S-MSCs | Negatively regulates Runx2 | Increases proliferation, migration, ECM secretion (increases COL2, SOX9) and reduced apoptosis in OA chondrocytes | human/mouse | [66] |
| miR-206 | mouse | Negatively regulates ELF3 | Increases proliferation, differentiation and reduces apoptosis in OA osteoblasts (increases OCN, BMP2, ALP and calcium deposition level; decreases pro-inflammatory mediators) | mouse | [90] |
| miR-210-5p | human subchondral bone osteoblast | May involve PI3K/AKT/mTOR signaling pathway | Increases hypertrophic, degradative gene expression, and changes cellular aerobic respiration in OA chondrocytes (increases ADAMTS5, COL10, RUNX2, MMP13; decreases SOX9, ACAN, COL2) | human | [52] |
| miR-214-3p | rat SFBs | Increases proliferation, and reduces apoptosis, inflammation (decreases TNF-α, IL-1β) in OA chondrocytes; reduces degeneration of OA cartilage and synovium; reduces osteophytes formation and maintains the subchondral bone structure in OA | rat | [49] | |
| miR-221 | human ADSCs | inhibits the NF-κB signaling pathway; actives Wnt/β-catenin signaling pathway | Increases viability and proliferation of periosteal cells | human | [94] |
| miR-320c | S-MSCs | Negatively regulates ADAM19, which up-regulates β-catenin, MYC and then actives Wnt signaling pathway | Increases proliferation, migration and reduces apoptosis in OA chondrocytes; increases secretion in OA ECM (increases COL2A1, ACAN) | rat | [104,105] |
| miR-320c | human BM-MSCs | Increases proliferation in OA chondrocytes; increases secretion in OA ECM (increases SOX9; decreases MMP13) | human | [80] | |
| miR-326 | rat BM-MSCs | Negatively regulates HDAC3, which negatively regulates STAT1 and then actives NF-κB signaling pathway | Increases proliferation, migration, the expression of chondrogenic specific genes (COL2A1, SOX9, Agg, and Prg4), and reduce inflammation(IL-6, TNF-α), pyroptosis (decreases NLRP3, ASC, GSDMD, Caspase-1, IL-1β, IL-18) in OA chondrocytes | rat | [101] |
| miR-361-5p | human BM-MSCs | Negatively regulates DDX20, which actives NF-κB signaling pathway | Reduces inflammation (decreases IL-18, IL-6, TNF-α) and degradation (decreases iNOS, MMP3, MMP13) in OA cartilage | rat | [81] |
| miR-let-7a-5p | mouse osteoclast | Negatively regulates Smad2 | Increases hypertrophic gene expression in OA chondrocytes (increases COL10A1, RUNX2, MMP13) | mouse | [53] |
4.2. Long Non-Coding RNAs (lncRNAs)
4.3. Circular RNAs (CircRNAs)
5. Limitation and Future Direction
5.1. Limitations in the Mechanism Research for OA Diagnosis and Treatment
5.2. Limitations in sEV Isolation Technology
Author Contributions
Funding
Conflicts of Interest
References
- Arden, N.; Blanco, F.; Cooper, C.; Guermazi, A.; Hayashi, D.; Hunter, D.; Javaid, M.K.; Rannou, F.; Roemer, F.; Reginster, J.Y. Atlas of Osteoarthritis M.; Springer Healthcare Ltd.: London, UK, 2014. [Google Scholar]
- Martel-Pelletier, J.; Barr, A.J.; Cicuttini, F.M.; Conaghan, P.G.; Cooper, C.; Goldring, M.B.; Goldring, S.R.; Jones, G.; Teichtahl, A.J.; Pelletier, J.P. Osteoarthritis. Nat. Rev. Dis. Primers 2016, 2, 16072. [Google Scholar] [CrossRef]
- Mancuso, P.; Raman, S.; Glynn, A.; Barry, F.; Murphy, J.M. Mesenchymal Stem Cell Therapy for Osteoarthritis: The Critical Role of the Cell Secretome. Front. Bioeng. Biotechnol. 2019, 7, 9. [Google Scholar] [CrossRef]
- Ruiz Iban, M.A.; Benavides, J.; Forero, J.P.; Bittelman, S.; Martinez, R.; Mite, M.A.; Diaz Heredia, J.; Ulloa, S.; Lizárraga Ferrand, M.M. Use of strong opioids for chronic pain in osteoarthritis: An insight into the Latin American reality. Expert Rev. Clin. Pharmacol. 2018, 11, 47–59. [Google Scholar] [CrossRef]
- The Institute of Joint Surgery; Orthopedic Branch of the Chinese Medical Association; Osteoarthritis Study Group of the Orthopedic Physician Branch of the Chinese Physician Association; National Clinical Research Center for Geriatric Diseases (Xiangya Hospital). Guideline fors diagnosis and treatment of osteoarthritis in China (2021 edition). Chin. J. Orthop. 2021, 41, 24. [Google Scholar]
- Hong, J.W.; Noh, J.H.; Kim, D.J. The prevalence of and demographic factors associated with radiographic knee osteoarthritis in Korean adults aged ≥ 50 years: The 2010–2013 Korea National Health and Nutrition Examination Survey. PLoS ONE 2020, 15, e0230613. [Google Scholar] [CrossRef] [PubMed]
- Mianehsaz, E.; Mirzaei, H.R.; Mahjoubin-Tehran, M.; Rezaee, A.; Sahebnasagh, R.; Pourhanifeh, M.H.; Mirzaei, H.; Hamblin, M.R. Mesenchymal stem cell-derived exosomes: A new therapeutic approach to osteoarthritis? Stem Cell Res. Ther. 2019, 10, 340. [Google Scholar] [CrossRef] [PubMed]
- Bao, C.; He, C. The role and therapeutic potential of MSC-derived exosomes in osteoarthritis. Arch. Biochem. Biophys. 2021, 710, 109002. [Google Scholar] [CrossRef] [PubMed]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef]
- Lobb, R.J.; Becker, M.; Wen, S.W.; Wong, C.S.; Wiegmans, A.P.; Leimgruber, A.; Möller, A. Optimized exosome isolation protocol for cell culture supernatant and human plasma. J. Extracell. Vesicles 2015, 4, 27031. [Google Scholar] [CrossRef]
- Zhang, Y.; Bi, J.; Huang, J.; Tang, Y.; Du, S.; Li, P. Exosome: A Review of Its Classification, Isolation Techniques, Storage, Diagnostic and Targeted Therapy Applications. Int. J. Nanomed. 2020, 15, 6917–6934. [Google Scholar] [CrossRef]
- Mao, G.; Zhang, Z.; Hu, S.; Zhang, Z.; Chang, Z.; Huang, Z.; Liao, W.; Kang, Y. Exosomes derived from miR-92a-3p-overexpressing human mesenchymal stem cells enhance chondrogenesis and suppress cartilage degradation via targeting WNT5A. Stem Cell Res. Ther. 2018, 9, 247. [Google Scholar] [CrossRef]
- Tao, S.C.; Yuan, T.; Zhang, Y.L.; Yin, W.J.; Guo, S.C.; Zhang, C.Q. Exosomes derived from miR-140-5p-overexpressing human synovial mesenchymal stem cells enhance cartilage tissue regeneration and prevent osteoarthritis of the knee in a rat model. Theranostics 2017, 7, 180–195. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zou, R.; Wang, Z.; Wen, C.; Zhang, F.; Lin, F. Exosomal KLF3-AS1 from hMSCs promoted cartilage repair and chondrocyte proliferation in osteoarthritis. Biochem. J. 2018, 475, 3629–3638. [Google Scholar] [CrossRef] [PubMed]
- Jin, Z.; Ren, J.; Qi, S. Exosomal miR-9-5p secreted by bone marrow-derived mesenchymal stem cells alleviates osteoarthritis by inhibiting syndecan-1. Cell Tissue Res. 2020, 381, 99–114. [Google Scholar] [CrossRef]
- Wang, Y.; He, S.H.; Liang, X.; Zhang, X.X.; Li, S.S.; Li, T.F. ATF4-modified serum exosomes derived from osteoarthritic mice inhibit osteoarthritis by inducing autophagy. IUBMB Life 2021, 73, 146–158. [Google Scholar] [CrossRef]
- Qiu, M.; Liu, D.; Fu, Q. MiR-129-5p shuttled by human synovial mesenchymal stem cell-derived exosomes relieves IL-1β induced osteoarthritis via targeting HMGB1. Life Sci. 2021, 269, 118987. [Google Scholar] [CrossRef]
- Lin, T.; Wu, N.; Wang, L.; Zhang, R.; Pan, R.; Chen, Y.F. Inhibition of chondrocyte apoptosis in a rat model of osteoarthritis by exosomes derived from miR-140-5p-overexpressing human dental pulp stem cells. Int. J. Mol. Med. 2021, 47, 7. [Google Scholar] [CrossRef]
- Jin, Y.; Xu, M.; Zhu, H.; Dong, C.; Ji, J.; Liu, Y.; Deng, A.; Gu, Z. Therapeutic effects of bone marrow mesenchymal stem cells-derived exosomes on osteoarthritis. J. Cell. Mol. Med. 2021, 25, 9281–9294. [Google Scholar] [CrossRef]
- Jiang, S.; Tian, G.; Yang, Z.; Gao, X.; Wang, F.; Li, J.; Tian, Z.; Huang, B.; Wei, F.; Sang, X. Enhancement of acellular cartilage matrix scaffold by Wharton’s jelly mesenchymal stem cell-derived exosomes to promote osteochondral regeneration. Bioact. Mater. 2021, 6, 2711–2728. [Google Scholar] [CrossRef]
- Wu, J.; Kuang, L.; Chen, C.; Yang, J.; Zeng, W.N.; Li, T.; Chen, H.; Huang, S.; Fu, Z.; Li, J.; et al. miR-100-5p-abundant exosomes derived from infrapatellar fat pad MSCs protect articular cartilage and ameliorate gait abnormalities via inhibition of mTOR in osteoarthritis. Biomaterials 2019, 206, 87–100. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Chu, W.C.; Lai, R.C.; Lim, S.K.; Hui, J.H.; Toh, W.S. Exosomes derived from human embryonic mesenchymal stem cells promote osteochondral regeneration. Osteoarthr. Cartil. 2016, 24, 2135–2140. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Wang, Y.; Zhao, B.; Niu, X.; Hu, B.; Li, Q.; Zhang, J.; Ding, J.; Chen, Y.; Wang, Y. Comparison of exosomes secreted by induced pluripotent stem cell-derived mesenchymal stem cells and synovial membrane-derived mesenchymal stem cells for the treatment of osteoarthritis. Stem Cell Res. Ther. 2017, 8, 64. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Chuah, S.J.; Lai, R.C.; Hui, J.H.P.; Lim, S.K.; Toh, W.S. MSC exosomes mediate cartilage repair by enhancing proliferation, attenuating apoptosis and modulating immune reactivity. Biomaterials 2018, 156, 16–27. [Google Scholar] [CrossRef] [PubMed]
- Cosenza, S.; Toupet, K.; Maumus, M.; Luz-Crawford, P.; Blanc-Brude, O.; Jorgensen, C.; Noël, D. Mesenchymal stem cells-derived exosomes are more immunosuppressive than microparticles in inflammatory arthritis. Theranostics 2018, 8, 1399–1410. [Google Scholar] [CrossRef]
- Gao, K.; Zhu, W.; Li, H.; Ma, D.; Liu, W.; Yu, W.; Wang, L.; Cao, Y.; Jiang, Y. Association between cytokines and exosomes in synovial fluid of individuals with knee osteoarthritis. Mod. Rheumatol. 2020, 30, 758–764. [Google Scholar] [CrossRef]
- Nam, Y.; Rim, Y.A.; Lee, J.; Ju, J.H. Current Therapeutic Strategies for Stem Cell-Based Cartilage Regeneration. Stem Cells Int. 2018, 2018, 8490489. [Google Scholar] [CrossRef]
- Caron, M.M.; Emans, P.J.; Coolsen, M.M.; Voss, L.; Surtel, D.A.; Cremers, A.; van Rhijn, L.W.; Welting, T.J. Redifferentiation of dedifferentiated human articular chondrocytes: Comparison of 2D and 3D cultures. Osteoarthr. Cartil. 2012, 20, 1170–1178. [Google Scholar] [CrossRef]
- Kim, Y.S.; Kwon, O.R.; Choi, Y.J.; Suh, D.S.; Heo, D.B.; Koh, Y.G. Comparative Matched-Pair Analysis of the Injection Versus Implantation of Mesenchymal Stem Cells for Knee Osteoarthritis. Am. J. Sports Med. 2015, 43, 2738–2746. [Google Scholar] [CrossRef]
- Wakitani, S.; Okabe, T.; Horibe, S.; Mitsuoka, T.; Saito, M.; Koyama, T.; Nawata, M.; Tensho, K.; Kato, H.; Uematsu, K.; et al. Safety of autologous bone marrow-derived mesenchymal stem cell transplantation for cartilage repair in 41 patients with 45 joints followed for up to 11 years and 5 months. J. Tissue Eng. Regen. Med. 2011, 5, 146–150. [Google Scholar] [CrossRef]
- Wong, K.L.; Lee, K.B.; Tai, B.C.; Law, P.; Lee, E.H.; Hui, J.H. Injectable cultured bone marrow-derived mesenchymal stem cells in varus knees with cartilage defects undergoing high tibial osteotomy: A prospective, randomized controlled clinical trial with 2 years’ follow-up. Arthroscopy 2013, 29, 2020–2028. [Google Scholar] [CrossRef]
- Orozco, L.; Munar, A.; Soler, R.; Alberca, M.; Soler, F.; Huguet, M.; Sentís, J.; Sánchez, A.; García-Sancho, J. Treatment of knee osteoarthritis with autologous mesenchymal stem cells: Two-year follow-up results. Transplantation 2014, 97, e66–e68. [Google Scholar] [CrossRef] [PubMed]
- Ahtiainen, K.; Mauno, J.; Ellä, V.; Hagström, J.; Lindqvist, C.; Miettinen, S.; Ylikomi, T.; Kellomäki, M.; Seppänen, R. Autologous adipose stem cells and polylactide discs in the replacement of the rabbit temporomandibular joint disc. J. R. Soc. Interface 2013, 10, 20130287. [Google Scholar] [CrossRef] [PubMed]
- Mäenpää, K.; Ellä, V.; Mauno, J.; Kellomäki, M.; Suuronen, R.; Ylikomi, T.; Miettinen, S. Use of adipose stem cells and polylactide discs for tissue engineering of the temporomandibular joint disc. J. R. Soc. Interface 2010, 7, 177–188. [Google Scholar] [CrossRef] [PubMed]
- Tatullo, M.; Marrelli, M.; Shakesheff, K.M.; White, L.J. Dental pulp stem cells: Function, isolation and applications in regenerative medicine. J. Tissue Eng. Regen. Med. 2015, 9, 1205–1216. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Liao, W.; Su, K.; Jia, J.; Qin, L.; Liu, W.; He, Y.; Zhang, H.; Ou, F.; Zhang, Z.; et al. Suberoylanilide Hydroxamic Acid Attenuates Interleukin-1β-Induced Interleukin-6 Upregulation by Inhibiting the Microtubule Affinity-Regulating Kinase 4/Nuclear Factor-κB Pathway in Synovium-Derived Mesenchymal Stem Cells from the Temporomandibular Joint. Inflammation 2020, 43, 1246–1258. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Sun, Y.; He, Y.; Zhang, H.; Zheng, Y.; Yao, Y.; Zhang, Z. IL-1β impedes the chondrogenic differentiation of synovial fluid mesenchymal stem cells in the human temporomandibular joint. Int. J. Mol. Med. 2017, 39, 317–326. [Google Scholar] [CrossRef]
- Lin, Y.; Umebayashi, M.; Abdallah, M.N.; Dong, G.; Roskies, M.G.; Zhao, Y.F.; Murshed, M.; Zhang, Z.; Tran, S.D. Combination of polyetherketoneketone scaffold and human mesenchymal stem cells from temporomandibular joint synovial fluid enhances bone regeneration. Sci. Rep. 2019, 9, 472. [Google Scholar] [CrossRef]
- Sun, Y.P.; Zheng, Y.H.; Liu, W.J.; Zheng, Y.L.; Zhang, Z.G. Synovium fragment-derived cells exhibit characteristics similar to those of dissociated multipotent cells in synovial fluid of the temporomandibular joint. PLoS ONE 2014, 9, e101896. [Google Scholar] [CrossRef]
- Koyama, N.; Okubo, Y.; Nakao, K.; Osawa, K.; Fujimura, K.; Bessho, K. Pluripotency of mesenchymal cells derived from synovial fluid in patients with temporomandibular joint disorder. Life Sci. 2011, 89, 741–747. [Google Scholar] [CrossRef]
- Zhang, J.; Guo, F.; Mi, J.; Zhang, Z. Periodontal ligament mesenchymal stromal cells increase proliferation and glycosaminoglycans formation of temporomandibular joint derived fibrochondrocytes. BioMed Res. Int. 2014, 2014, 410167. [Google Scholar] [CrossRef]
- Ogasawara, N.; Kano, F.; Hashimoto, N.; Mori, H.; Liu, Y.; Xia, L.; Sakamaki, T.; Hibi, H.; Iwamoto, T.; Tanaka, E.; et al. Factors secreted from dental pulp stem cells show multifaceted benefits for treating experimental temporomandibular joint osteoarthritis. Osteoarthr. Cartil. 2020, 28, 831–841. [Google Scholar] [CrossRef] [PubMed]
- De Almeida, P.E.; Ransohoff, J.D.; Nahid, A.; Wu, J.C. Immunogenicity of pluripotent stem cells and their derivatives. Circ. Res. 2013, 112, 549–561. [Google Scholar] [CrossRef]
- Kalluri, R.; Lebleu, V.S. The biology, function, and biomedical applications of exosomes. Science 2020, 367, eaau6977. [Google Scholar] [CrossRef] [PubMed]
- Murao, A.; Brenner, M.; Aziz, M.; Wang, P. Exosomes in Sepsis. Front. Immunol. 2020, 11, 2140. [Google Scholar] [CrossRef]
- Li, Z.; Huang, Z.; Bai, L. Cell Interplay in Osteoarthritis. Front. Cell Dev. Biol. 2021, 9, 720477. [Google Scholar] [CrossRef]
- Yu, H.; Huang, Y.; Yang, L. Research progress in the use of mesenchymal stem cells and their derived exosomes in the treatment of osteoarthritis. Ageing Res. Rev. 2022, 80, 101684. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Ming, J.; Li, Y.; Li, B.; Deng, M.; Ma, Y.; Chen, Z.; Zhang, Y.; Li, J.; Liu, S. Exosomes derived from miR-126-3p-overexpressing synovial fibroblasts suppress chondrocyte inflammation and cartilage degradation in a rat model of osteoarthritis. Cell Death Discov. 2021, 7, 37. [Google Scholar] [CrossRef]
- Lai, C.; Liao, B.; Peng, S.; Fang, P.; Bao, N.; Zhang, L. Synovial fibroblast-miR-214-3p-derived exosomes inhibit inflammation and degeneration of cartilage tissues of osteoarthritis rats. Mol. Cell. Biochem. 2022, 478, 637–649. [Google Scholar] [CrossRef]
- Tan, F.; Wang, D.; Yuan, Z. The Fibroblast-Like Synoviocyte Derived Exosomal Long Non-coding RNA H19 Alleviates Osteoarthritis Progression Through the miR-106b-5p/TIMP2 Axis. Inflammation 2020, 43, 1498–1509. [Google Scholar] [CrossRef]
- Kato, T.; Miyaki, S.; Ishitobi, H.; Nakamura, Y.; Nakasa, T.; Lotz, M.K.; Ochi, M. Exosomes from IL-1β stimulated synovial fibroblasts induce osteoarthritic changes in articular chondrocytes. Arthritis Res. Ther. 2014, 16, R163. [Google Scholar] [CrossRef]
- Wu, X.; Crawford, R.; Xiao, Y.; Mao, X.; Prasadam, I. Osteoarthritic Subchondral Bone Release Exosomes That Promote Cartilage Degeneration. J. Cells 2021, 10, 251. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.; Dong, R.; Han, X.; Li, J.; Gong, X.; Bai, Y.; Kang, F.; Liang, M.; Zeng, F.; Hou, Z.; et al. Osteoclast-derived exosomal let-7a-5p targets Smad2 to promote the hypertrophic differentiation of chondrocytes. Am. J. Physiol. Cell Physiol. 2020, 319, C21–C33. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.Z.; Zheng, H.L.; Xu, W.N.; Zheng, X.F.; Li, B.; Jiang, L.S.; Jiang, S.D. Vascular endothelial cell-secreted exosomes facilitate osteoarthritis pathogenesis by promoting chondrocyte apoptosis. Aging 2021, 13, 4647–4662. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Wang, H.; Zhao, F.; Liu, M.; Wang, F.; Kang, M.; He, W.; Lv, Z. Exosomal circ-BRWD1 contributes to osteoarthritis development through the modulation of miR-1277/TRAF6 axis. Arthritis Res. Ther. 2021, 23, 159. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.; Shen, K.; Zhou, W.; Wu, H.; Lu, Y. Exosome-mediated circ_0001846 participates in IL-1β-induced chondrocyte cell damage by miR-149-5p-dependent regulation of WNT5B. Clin. Immunol. 2021, 232, 108856. [Google Scholar] [CrossRef] [PubMed]
- Lai, X.; Song, Y.; Tian, J. CircCDK14 ameliorates interleukin-1β-induced chondrocyte damage by the miR-1183/KLF5 pathway in osteoarthritis. Autoimmunity 2022, 55, 408–417. [Google Scholar] [CrossRef]
- Meng, Y.; Qiu, S.; Sun, L.; Zuo, J. Knockdown of exosome-mediated lnc-PVT1 alleviates lipopolysaccharide-induced osteoarthritis progression by mediating the HMGB1/TLR4/NF-κB pathway via miR-93-5p. Mol. Med. Rep. 2020, 22, 5313–5325. [Google Scholar] [CrossRef]
- Zeng, G.; Deng, G.; Xiao, S.; Li, F. Fibroblast-like Synoviocytes-derived Exosomal PCGEM1 Accelerates IL-1β-induced Apoptosis and Cartilage Matrix Degradation by miR-142-5p/RUNX2 in Chondrocytes. Immunol. Investig. 2022, 51, 1284–1301. [Google Scholar] [CrossRef]
- Liu, Y.; Zeng, Y.; Si, H.B.; Tang, L.; Xie, H.Q.; Shen, B. Exosomes Derived From Human Urine-Derived Stem Cells Overexpressing miR-140-5p Alleviate Knee Osteoarthritis Through Downregulation of VEGFA in a Rat Model. Am. J. Sports Med. 2022, 50, 1088–1105. [Google Scholar] [CrossRef]
- Ni, Z.; Kuang, L.; Chen, H.; Xie, Y.; Zhang, B.; Ouyang, J.; Wu, J.; Zhou, S.; Chen, L.; Su, N.; et al. The exosome-like vesicles from osteoarthritic chondrocyte enhanced mature IL-1β production of macrophages and aggravated synovitis in osteoarthritis. Cell death & disease 2019, 10, 522. [Google Scholar]
- Wang, R.; Xu, B. TGF-β1-modified MSC-derived exosomal miR-135b attenuates cartilage injury via promoting M2 synovial macrophage polarization by targeting MAPK6. Cell Tissue Res. 2021, 384, 113–127. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Lin, L.; Zou, R.; Wen, C.; Wang, Z.; Lin, F. MSC-derived exosomes promote proliferation and inhibit apoptosis of chondrocytes via lncRNA-KLF3-AS1/miR-206/GIT1 axis in osteoarthritis. Cell Cycle 2018, 17, 2411–2422. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Xu, B.; Xu, H. TGF-β1 promoted chondrocyte proliferation by regulating Sp1 through MSC-exosomes derived miR-135b. Cell Cycle 2018, 17, 2756–2765. [Google Scholar] [CrossRef]
- Li, X.; Wang, Y.; Cai, Z.; Zhou, Q.; Li, L.; Fu, P. Exosomes from human umbilical cord mesenchymal stem cells inhibit ROS production and cell apoptosis in human articular chondrocytes via the miR-100-5p/NOX4 axis. Cell Biol. Int. 2021, 45, 2096–2106. [Google Scholar] [CrossRef]
- Wang, Z.; Yan, K.; Ge, G.; Zhang, D.; Bai, J.; Guo, X.; Zhou, J.; Xu, T.; Xu, M.; Long, X.; et al. Exosomes derived from miR-155-5p-overexpressing synovial mesenchymal stem cells prevent osteoarthritis via enhancing proliferation and migration, attenuating apoptosis, and modulating extracellular matrix secretion in chondrocytes. Cell Biol. Toxicol. 2021, 37, 85–96. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.; Chen, M.; Cui, X.; Liu, Y.; Liu, Y.; Deng, S.; Yuan, T.; Fan, Y.; Wang, Q.; Zhang, X. Cell-Free Osteoarthritis Treatment with Sustained-Release of Chondrocyte-Targeting Exosomes from Umbilical Cord-Derived Mesenchymal Stem Cells to Rejuvenate Aging Chondrocytes. ACS nano 2023, 17, 13358–13376. [Google Scholar] [CrossRef]
- Yan, L.; Liu, G.; Wu, X. The umbilical cord mesenchymal stem cell-derived exosomal lncRNA H19 improves osteochondral activity through miR-29b-3p/FoxO3 axis. Clin. Transl. Med. 2021, 11, e255. [Google Scholar] [CrossRef]
- Chang, L.H.; Wu, S.C.; Chen, C.H.; Chen, J.W.; Huang, W.C.; Wu, C.W.; Lin, Y.S.; Chen, Y.J.; Chang, J.K.; Ho, M.L. Exosomes Derived from Hypoxia-Cultured Human Adipose Stem Cells Alleviate Articular Chondrocyte Inflammaging and Post-Traumatic Osteoarthritis Progression. Int. J. Mol. Sci. 2023, 24, 13414. [Google Scholar] [CrossRef]
- Luo, P.; Jiang, C.; Ji, P.; Wang, M.; Xu, J. Exosomes of stem cells from human exfoliated deciduous teeth as an anti-inflammatory agent in temporomandibular joint chondrocytes via miR-100-5p/mTOR J. Stem Cell Res. Ther. 2019, 10, 216. [Google Scholar] [CrossRef]
- Li, Z.; Wang, Y.; Xiang, S.; Zheng, Z.; Bian, Y.; Feng, B.; Weng, X. Chondrocytes-derived exosomal miR-8485 regulated the Wnt/β-catenin pathways to promote chondrogenic differentiation of BMSCs. Biochem. Biophys. Res. Commun. 2020, 523, 506–513. [Google Scholar] [CrossRef]
- Ma, K.; Zhu, B.; Wang, Z.; Cai, P.; He, M.; Ye, D.; Yan, G.; Zheng, L.; Yang, L.; Zhao, J. Articular chondrocyte-derived extracellular vesicles promote cartilage differentiation of human umbilical cord mesenchymal stem cells by activation of autophagy. J. Nanobiotechnol. 2020, 18, 163. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, X.; Chen, J.; Qian, D.; Gao, P.; Qin, T.; Jiang, T.; Yi, J.; Xu, T.; Huang, Y.; et al. Exosomes derived from platelet-rich plasma administration in site mediate cartilage protection in subtalar osteoarthritis. J. Nanobiotechnol. 2022, 20, 56. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Xue, K.; Zhang, X.; Zheng, Z.; Liu, K. Exosomes derived from mature chondrocytes facilitate subcutaneous stable ectopic chondrogenesis of cartilage progenitor cells. Stem Cell Res. Ther. 2018, 9, 318. [Google Scholar] [CrossRef]
- Xia, Q.; Wang, Q.; Lin, F.; Wang, J. miR-125a-5p-abundant exosomes derived from mesenchymal stem cells suppress chondrocyte degeneration via targeting E2F2 in traumatic osteoarthritis. Bioengineered 2021, 12, 11225–11238. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Shi, Y.; Xue, P.; Ma, X.; Li, J.; Zhang, J. Mesenchymal stem cell-derived exosomal microRNA-136-5p inhibits chondrocyte degeneration in traumatic osteoarthritis by targeting ELF3. Arthritis Res. Ther. 2020, 22, 256. [Google Scholar] [CrossRef] [PubMed]
- He, L.; He, T.; Xing, J.; Zhou, Q.; Fan, L.; Liu, C.; Chen, Y.; Wu, D.; Tian, Z.; Liu, B.; et al. Bone marrow mesenchymal stem cell-derived exosomes protect cartilage damage and relieve knee osteoarthritis pain in a rat model of osteoarthritis. Stem Cell Res. Ther. 2020, 11, 276. [Google Scholar] [CrossRef]
- Cosenza, S.; Ruiz, M.; Toupet, K.; Jorgensen, C.; Noël, D. Mesenchymal stem cells derived exosomes and microparticles protect cartilage and bone from degradation in osteoarthritis. Sci. Rep. 2017, 7, 16214. [Google Scholar] [CrossRef]
- Mao, G.; Hu, S.; Zhang, Z.; Wu, P.; Zhao, X.; Lin, R.; Liao, W.; Kang, Y. Exosomal miR-95-5p regulates chondrogenesis and cartilage degradation via histone deacetylase 2/8. J. Cell. Mol. Med. 2018, 22, 5354–5366. [Google Scholar] [CrossRef]
- Sun, H.; Hu, S.; Zhang, Z.; Lun, J.; Liao, W.; Zhang, Z. Expression of exosomal microRNAs during chondrogenic differentiation of human bone mesenchymal stem cells. J. Cell. Biochem. 2019, 120, 171–181. [Google Scholar] [CrossRef]
- Tao, Y.; Zhou, J.; Wang, Z.; Tao, H.; Bai, J.; Ge, G.; Li, W.; Zhang, W.; Hao, Y.; Yang, X.; et al. Human bone mesenchymal stem cells-derived exosomal miRNA-361-5p alleviates osteoarthritis by downregulating DDX20 and inactivating the NF-κB signaling pathway. Bioorganic Chem. 2021, 113, 104978. [Google Scholar] [CrossRef]
- Mao, G.; Xu, Y.; Long, D.; Sun, H.; Li, H.; Xin, R.; Zhang, Z.; Li, Z.; Yang, Z.; Kang, Y. Exosome-transported circRNA_0001236 enhances chondrogenesis and suppress cartilage degradation via the miR-3677-3p/Sox9 axis. Stem Cell Res. Ther. 2021, 12, 389. [Google Scholar] [CrossRef] [PubMed]
- Liao, Q.; Li, B.J.; Li, Y.; Xiao, Y.; Zeng, H.; Liu, J.M.; Yuan, L.X.; Liu, G. Low-intensity pulsed ultrasound promotes osteoarthritic cartilage regeneration by BMSC-derived exosomes via modulating the NF-κB signaling pathway. Int. Immunopharmacol. 2021, 97, 107824. [Google Scholar] [CrossRef]
- Xia, P.; Wang, Q.; Song, J.; Wang, X.; Wang, X.; Lin, Q.; Cheng, K.; Chen, A.; Li, X. Low-Intensity Pulsed Ultrasound Enhances the Efficacy of Bone Marrow-Derived MSCs in Osteoarthritis Cartilage Repair by Regulating Autophagy-Mediated Exosome Release. Cartilage 2022, 13, 19476035221093060. [Google Scholar] [CrossRef] [PubMed]
- Shao, L.T.; Luo, L.; Qiu, J.H.; Deng, D.Y.B. PTH (1-34) enhances the therapeutic effect of bone marrow mesenchymal stem cell-derived exosomes by inhibiting proinflammatory cytokines expression on OA chondrocyte repair in vitro. Arthritis Res. Ther. 2022, 24, 96. [Google Scholar] [CrossRef]
- Wang, Y.; Yu, D.; Liu, Z.; Zhou, F.; Dai, J.; Wu, B.; Zhou, J.; Heng, B.C.; Zou, X.H.; Ouyang, H.; et al. Exosomes from embryonic mesenchymal stem cells alleviate osteoarthritis through balancing synthesis and degradation of cartilage extracellular matrix. Stem Cell Res. Ther. 2017, 8, 189. [Google Scholar] [CrossRef] [PubMed]
- Tofiño-Vian, M.; Guillén, M.I.; Pérez Del Caz, M.D.; Silvestre, A.; Alcaraz, M.J. Microvesicles from Human Adipose Tissue-Derived Mesenchymal Stem Cells as a New Protective Strategy in Osteoarthritic Chondrocytes. Cell. Physiol. Biochem. 2018, 47, 11–25. [Google Scholar] [CrossRef]
- Huang, C.Y.; Vesvoranan, O.; Yin, X.; Montoya, A.; Londono, V.; Sawatari, Y.; Garcia-Godoy, F. Anti-Inflammatory Effects of Conditioned Medium of Periodontal Ligament-Derived Stem Cells on Chondrocytes, Synoviocytes, and Meniscus Cells. Stem Cells Dev. 2021, 30, 537–547. [Google Scholar] [CrossRef]
- Bai, J.; Zhang, Y.; Zheng, X.; Huang, M.; Cheng, W.; Shan, H.; Gao, X.; Zhang, M.; Sheng, L.; Dai, J.; et al. LncRNA MM2P-induced, exosome-mediated transfer of Sox9 from monocyte-derived cells modulates primary chondrocytes. Cell Death Dis. 2020, 11, 763. [Google Scholar] [CrossRef]
- Huang, Y.; Zhang, X.; Zhan, J.; Yan, Z.; Chen, D.; Xue, X.; Pan, X. Bone marrow mesenchymal stem cell-derived exosomal miR-206 promotes osteoblast proliferation and differentiation in osteoarthritis by reducing Elf3. J. Cell. Mol. Med. 2021, 25, 7734–7745. [Google Scholar] [CrossRef]
- Yang, Q.; Yao, Y.; Zhao, D.; Zou, H.; Lai, C.; Xiang, G.; Wang, G.; Luo, L.; Shi, Y.; Li, Y.; et al. LncRNA H19 secreted by umbilical cord blood mesenchymal stem cells through microRNA-29a-3p/FOS axis for central sensitization of pain in advanced osteoarthritis. Am. J. Transl. Res. 2021, 13, 1245–1256. [Google Scholar]
- Li, J.; Ding, Z.; Li, Y.; Wang, W.; Wang, J.; Yu, H.; Liu, A.; Miao, J.; Chen, S.; Wu, T.; et al. BMSCs-Derived Exosomes Ameliorate Pain Via Abrogation of Aberrant Nerve Invasion in Subchondral Bone in Lumbar Facet Joint Osteoarthritis. J. Orthop. Res. Off. Publ. Orthop. Res. Soc. 2020, 38, 670–679. [Google Scholar] [CrossRef]
- Wang, R.; Xu, B. TGFβ1-modified MSC-derived exosome attenuates osteoarthritis by inhibiting PDGF-BB secretion and H-type vessel activity in the subchondral bone. Acta Histochem. 2022, 124, 151933. [Google Scholar] [CrossRef]
- Zhao, C.; Chen, J.Y.; Peng, W.M.; Yuan, B.; Bi, Q.; Xu, Y.J. Exosomes from adipose-derived stem cells promote chondrogenesis and suppress inflammation by upregulating miR-145 and miR-221. Mol. Med. Rep. 2020, 21, 1881–1889. [Google Scholar] [CrossRef] [PubMed]
- Jin, Z.; Ren, J.; Qi, S. Human bone mesenchymal stem cells-derived exosomes overexpressing microRNA-26a-5p alleviate osteoarthritis via down-regulation of PTGS2. Int. Immunopharmacol. 2020, 78, 105946. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Shin, D.I.; Choi, B.H.; Min, B.H. Exosomes from IL-1β-Primed Mesenchymal Stem Cells Inhibited IL-1β- and TNF-α-Mediated Inflammatory Responses in Osteoarthritic SW982 Cells. Tissue Eng. Regen. Med. 2021, 18, 525–536. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Rong, Y.; Luo, C.; Cui, W. Bone marrow mesenchymal stem cell-derived exosomes prevent osteoarthritis by regulating synovial macrophage polarization. Aging 2020, 12, 25138–25152. [Google Scholar] [CrossRef]
- Liang, C.; Wu, S.; Xia, G.; Huang, J.; Wen, Z.; Zhang, W.; Cao, X. Engineered M2a macrophages for the treatment of osteoarthritis. Front. Immunol. FRONT IMMUNOL 2022, 13, 1054938. [Google Scholar] [CrossRef]
- Foo, J.B.; Looi, Q.H.; How, C.W.; Lee, S.H.; Al-Masawa, M.E.; Chong, P.P.; Law, J.X. Mesenchymal Stem Cell-Derived Exosomes and MicroRNAs in Cartilage Regeneration: Biogenesis, Efficacy, miRNA Enrichment and Delivery. Pharmaceuticals 2021, 14, 1093. [Google Scholar] [CrossRef]
- Yao, Q.; Wu, X.; Tao, C.; Gong, W.; Chen, M.; Qu, M.; Zhong, Y.; He, T.; Chen, S.; Xiao, G. Osteoarthritis: Pathogenic signaling pathways and therapeutic targets. Signal Transduct. Target. Ther. 2023, 8, 56. [Google Scholar] [CrossRef]
- Xu, H.; Xu, B. BMSC-Derived Exosomes Ameliorate Osteoarthritis by Inhibiting Pyroptosis of Cartilage via Delivering miR-326 Targeting HDAC3 and STAT1//NF-κB p65 to Chondrocytes. Mediat. Inflamm. 2021, 2021, 9972805. [Google Scholar] [CrossRef]
- Qiu, B.; Xu, X.; Yi, P.; Hao, Y. Curcumin reinforces MSC-derived exosomes in attenuating osteoarthritis via modulating the miR-124/NF-kB and miR-143/ROCK1/TLR9 signalling pathways. J. Cell. Mol. Med. 2020, 24, 10855–10865. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.; Li, L.; Fang, X.; Zang, M. Exosome-Encapsulated microRNA-127-3p Released from Bone Marrow-Derived Mesenchymal Stem Cells Alleviates Osteoarthritis Through Regulating CDH11-Mediated Wnt/β-Catenin Pathway. J. Pain Res. 2021, 14, 297–310. [Google Scholar] [CrossRef] [PubMed]
- Kong, R.; Gao, J.; Zhang, J.; Ji, L.; Yu, Y.; Zhang, L.; Zhao, D. Synovial mesenchymal stem cell-derived exosomal miR-320c enhances chondrogenesis by targeting ADAM19. Future Med. Chem. 2022, 14, 81–96. [Google Scholar] [CrossRef]
- Kong, R.; Zhang, J.; Ji, L.; Yu, Y.; Gao, J.; Zhao, D. Synovial mesenchymal stem cell-derived exosomal microRNA-320c facilitates cartilage damage repair by targeting ADAM19-dependent Wnt signalling in osteoarthritis rats. Inflammopharmacology 2023, 31, 915–926. [Google Scholar] [CrossRef]
- Bridges, M.C.; Daulagala, A.C.; Kourtidis, A. LNCcation: lncRNA localization and function. J. Cell Biol. 2021, 220, e202009045. [Google Scholar] [CrossRef] [PubMed]
- Xie, F.; Liu, Y.L.; Chen, X.Y.; Li, Q.; Zhong, J.; Dai, B.Y.; Shao, X.F.; Wu, G.B. Role of MicroRNA, LncRNA, and Exosomes in the Progression of Osteoarthritis: A Review of Recent Literature. Orthop. Surg. 2020, 12, 708–716. [Google Scholar] [CrossRef]
- Ali, S.A.; Peffers, M.J.; Ormseth, M.J.; Jurisica, I.; Kapoor, M. The non-coding RNA interactome in joint health and disease. Nat. Rev. Rheumatol. 2021, 17, 692–705. [Google Scholar] [CrossRef]
- Liu, Z.; Cao, J.; Zhang, L.; Li, J.; Yan, T.; Zhou, P.; Zhang, S. Knockdown of circ-PRKCH alleviates IL-1β-treated chondrocyte cell phenotypic changes through modulating miR-502-5p/ADAMTS5 axis. Autoimmunity 2022, 55, 179–191. [Google Scholar] [CrossRef]
- Chen, T.S.; Arslan, F.; Yin, Y.; Tan, S.S.; Lai, R.C.; Choo, A.B.; Padmanabhan, J.; Lee, C.N.; de Kleijn, D.P.; Lim, S.K. Enabling a robust scalable manufacturing process for therapeutic exosomes through oncogenic immortalization of human ESC-derived MSCs. J. Transl. Med. 2011, 9, 47. [Google Scholar] [CrossRef]
- Piñeiro-Ramil, M.; Castro-Viñuelas, R.; Sanjurjo-Rodríguez, C.; Rodríguez-Fernández, S.; Hermida-Gómez, T.; Blanco-García, F.J.; Fuentes-Boquete, I.; Díaz-Prado, S. Immortalizing Mesenchymal Stromal Cells from Aged Donors While Keeping Their Essential Features. Stem Cells Int. 2020, 2020, 5726947. [Google Scholar] [CrossRef]
- Albanese, M.; Chen, Y.A.; Hüls, C.; Gärtner, K.; Tagawa, T.; Mejias-Perez, E.; Keppler, O.T.; Göbel, C.; Zeidler, R.; Shein, M.; et al. MicroRNAs are minor constituents of extracellular vesicles that are rarely delivered to target cells. PLoS Genet. 2021, 17, e1009951. [Google Scholar] [CrossRef] [PubMed]
- Chevillet, J.R.; Kang, Q.; Ruf, I.K.; Briggs, H.A.; Vojtech, L.N.; Hughes, S.M.; Cheng, H.H.; Arroyo, J.D.; Meredith, E.K.; Gallichotte, E.N.; et al. Quantitative and stoichiometric analysis of the microRNA content of exosomes. Proc. Natl. Acad. Sci. USA 2014, 111, 14888–14893. [Google Scholar] [CrossRef] [PubMed]
- Roefs, M.T.; Bauzá-Martinez, J.; van de Wakker, S.I.; Qin, J.; Olijve, W.T.; Tuinte, R.; Rozeboom, M.; Snijders Blok, C.; Mol, E.A.; Wu, W.; et al. Cardiac progenitor cell-derived extracellular vesicles promote angiogenesis through both associated- and co-isolated proteins. Commun. Biol. 2023, 6, 800. [Google Scholar] [CrossRef] [PubMed]
- Yin, B.; Ni, J.; Witherel, C.E.; Yang, M.; Burdick, J.A.; Wen, C.; Wong, S.H.D. Harnessing Tissue-derived Extracellular Vesicles for Osteoarthritis Theranostics. Theranostics 2022, 12, 207–231. [Google Scholar] [CrossRef]
- Simonsen, J.B. What Are We Looking At? Extracellular Vesicles, Lipoproteins, or Both? Circ. Res. 2017, 121, 920–922. [Google Scholar] [CrossRef]
- Vickers, K.C.; Palmisano, B.T.; Shoucri, B.M.; Shamburek, R.D.; Remaley, A.T. MicroRNAs are transported in plasma and delivered to recipient cells by high-density lipoproteins. Nat. Cell Biol. 2011, 13, 423–433. [Google Scholar] [CrossRef]
- Witwer, K.W.; Halushka, M.K. Toward the promise of microRNAs - Enhancing reproducibility and rigor in microRNA research. RNA Biol. 2016, 13, 1103–1116. [Google Scholar] [CrossRef]
- Mayya, V.K.; Duchaine, T.F. On the availability of microRNA-induced silencing complexes, saturation of microRNA-binding sites and stoichiometry. Nucleic Acids Res. 2015, 43, 7556–7565. [Google Scholar] [CrossRef]
| LncRNA | Origin of sEV | Mechanism | Effects | Species | Ref. |
|---|---|---|---|---|---|
| KLF3-AS1 | human MSCs | Sponges miR- 206, which negatively regulates GIT1 | Increases proliferation and reduces apoptosis, hypertrophy (decreases MMP13, Runx2) in OA chondrocytes, and increases secretion in OA ECM (increases COL2A1, ACAN) | mouse | [63] |
| PCGEM1 | human SFBs | Sponges miR-142-5p, which negatively regulates RUNX2 | Increases apoptosis in OA chondrocytes; increases degradation in OA ECM (increases MMP13; decreases ACAN, COL2A1) | human | [59] |
| PVT1 | human CCs | Sponges miR-93-5p, which negatively regulates HMGB1 and then negatively regulates TLR4 to inhibit NF-κB signaling pathway | Increases apoptosis (increases Bax and cleaved caspase-3; decreases Bcl-2), inflammation responses (increases IL-6, IL-1β, TNF-α) and reduces viability in OA chondrocytes; increases collagen degradation in OA ECM(increases MMP13; decreases ACAN) | human | [58] |
| MEG-3 | human BM-MSCs | May involve miR-206/GIT1, miR-92a-3p/Wnt5a, miR-93/TGFBR2, miR-16/SMAD7 axis | Reduces senescence and apoptosis in OA chondrocytes | rat | [19] |
| H19 | human UC-MSCs | Sponges miR-29a-3p, which negatively regulates FOS | Reduces pain and central sensitization in advanced OA | rat | [91] |
| human UC-MSCs | Sponges miR-29a-3p, which negatively regulates FOXO3 | Increases migration and reduces senescence and apoptosis in OA chondrocytes; increases secretion in OA ECM (increases COL2A1, ACAN) | human/rat | [68] | |
| rat SFBs | Sponges miR-106b-5p, which negatively regulates TIMP2 | Increases proliferation and migration in OA chondrocytes; reduces degeneration in OA cartilage (increases COL2A1, ACAN; decreases MMP13, ADAMTS5) | human | [50] |
| CircRNA | Origin of sEV | Mechanism | Effects | Species | Ref. |
|---|---|---|---|---|---|
| circ_0001236 | human BM-MSCs | Sponges miR-3677-3p, which negatively regulates Sox9 | Regulates degradation and repair in OA ECM (increases COL2A1 and SOX9; decreases MMP13) | mouse | [82] |
| circ_0001846 | human CCs | Sponges miR-149-5p, which negatively regulates WNT5B | Reduces cell viability, invasion and migration, and increases apoptosis, inflammatory cytokines production in OA chondrocytes; and increases degradation in ECM | human | [56] |
| circBRWD1 | human CCs | Sponges miR-1277, which negatively regulates TRAF6 | Reduces cell viability and proliferation and increases apoptosis (increases Bax; decreases CyclinD1), inflammation (increases IL-6, IL-8) and ECM degradation (increases MMP13; decreases ACAN) in OA chondrocytes | human | [55] |
| circCDK14 | human CCs | Sponges miR-1183, which negatively regulates KLF5 | Increases proliferation, and reduces apoptosis in OA chondrocytes; reduces degradation in OA ECM | human | [57] |
| circPRKCH | human CCs | Sponges miR-502-5p, which negatively regulates ADAMTS5 | Reduces proliferation, migration, and increases apoptosis, inflammatory response to promote phenotypic changes in OA chondrocytes | human | [109] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Z.; Bi, R.; Zhu, S. The Dual Role of Small Extracellular Vesicles in Joint Osteoarthritis: Their Global and Non-Coding Regulatory RNA Molecule-Based Pathogenic and Therapeutic Effects. Biomolecules 2023, 13, 1606. https://doi.org/10.3390/biom13111606
Li Z, Bi R, Zhu S. The Dual Role of Small Extracellular Vesicles in Joint Osteoarthritis: Their Global and Non-Coding Regulatory RNA Molecule-Based Pathogenic and Therapeutic Effects. Biomolecules. 2023; 13(11):1606. https://doi.org/10.3390/biom13111606
Chicago/Turabian StyleLi, Zhi, Ruiye Bi, and Songsong Zhu. 2023. "The Dual Role of Small Extracellular Vesicles in Joint Osteoarthritis: Their Global and Non-Coding Regulatory RNA Molecule-Based Pathogenic and Therapeutic Effects" Biomolecules 13, no. 11: 1606. https://doi.org/10.3390/biom13111606