
Changes in Expression in BMP2 and Two Closely Related Genes in Guinea Pig Retinal Pigment Epithelium during Induction and Recovery from Myopia

Abstract
:1. Introduction
2. Methods
2.1. Animals
2.2. Rigid Gas-Permeable Contact Lense-Induced Myopia Model
2.3. Biometric Data Collection
2.3.1. Refractive Errors and Axial Lengths
2.3.2. Choroidal and Scleral Thicknesses (SD-OCT)
2.4. Retinal Pigment Epithelium (RPE) Gene Expression
2.4.1. RPE Collection
2.4.2. Selection of RPE Genes for Analysis
2.4.3. Quantitative Real-Time PCR (qRT-PCR)
2.5. Statistical Analyses
3. Results
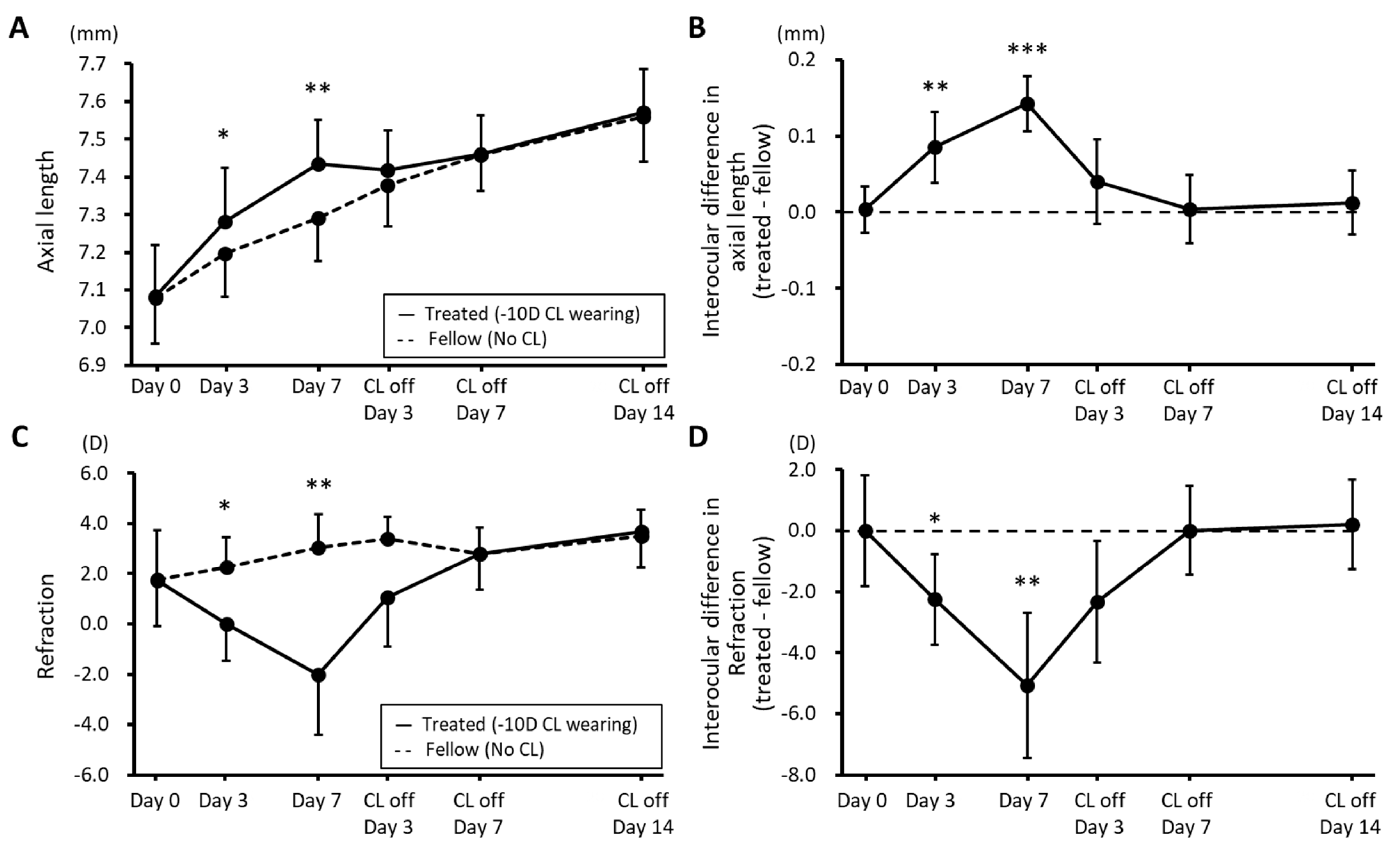
3.1. Changes in Biometric Parameters during Myopia Induction and Recovery
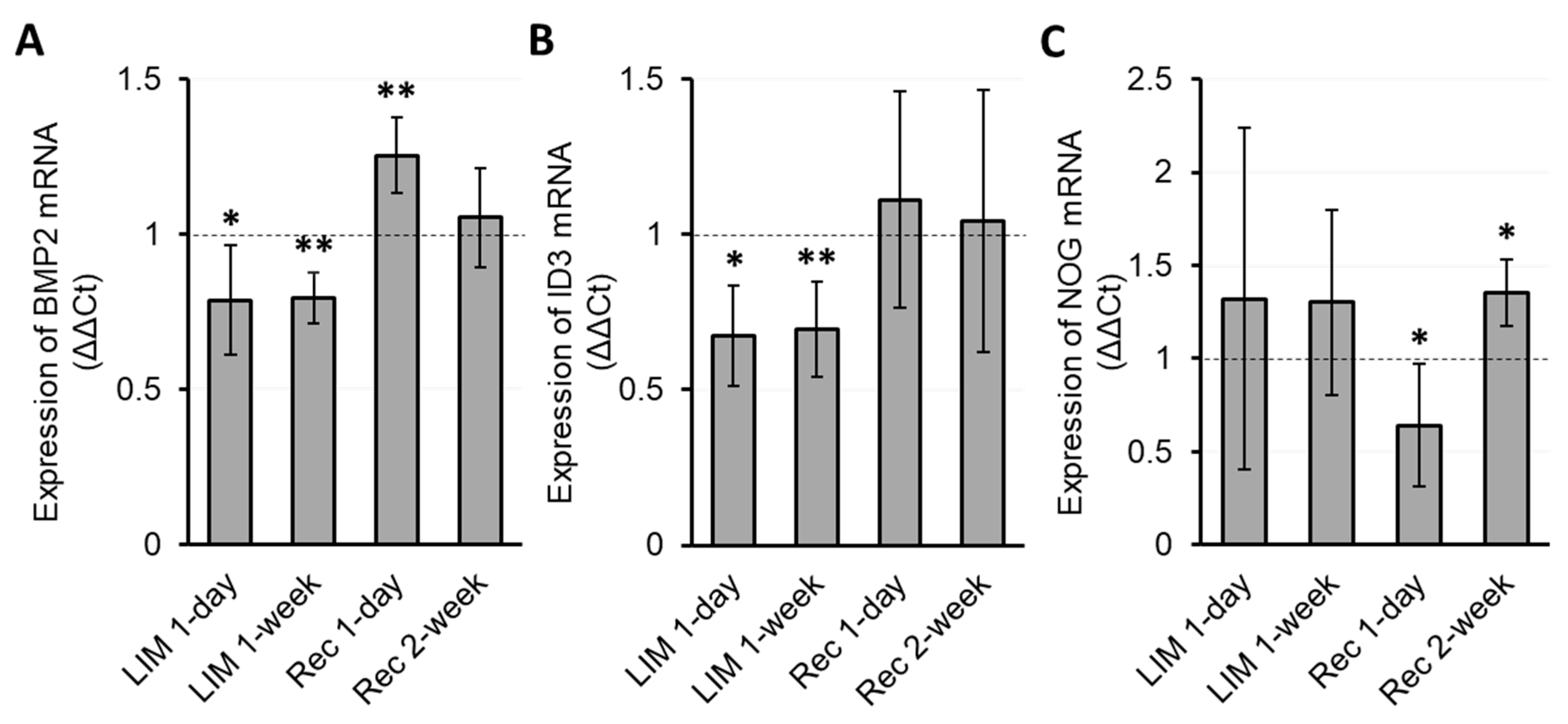
3.2. RPE Bmp2, Id3, and Nog Expression Changes with Myopia Induction and Recovery
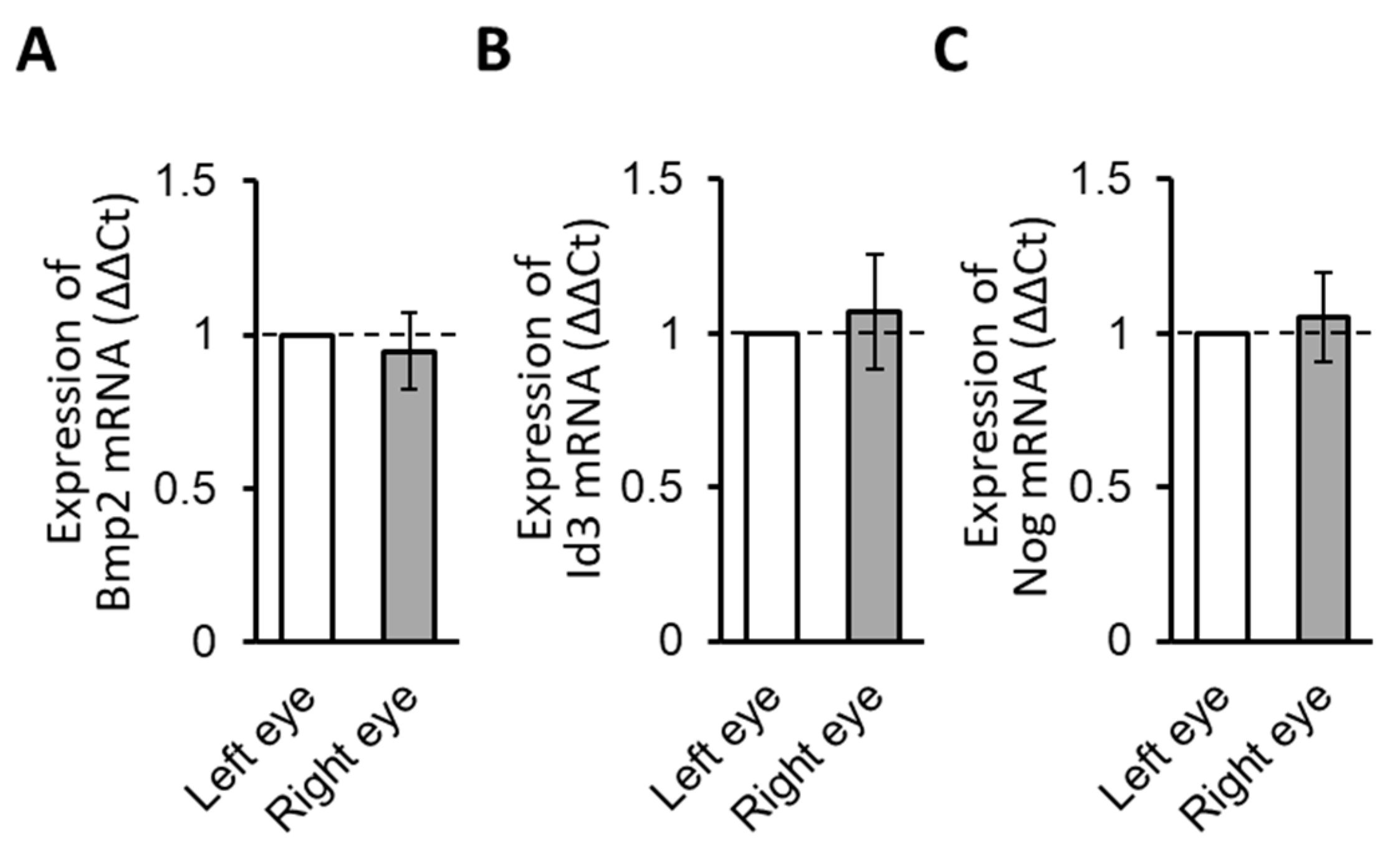
3.2.1. Normal Gene Expression
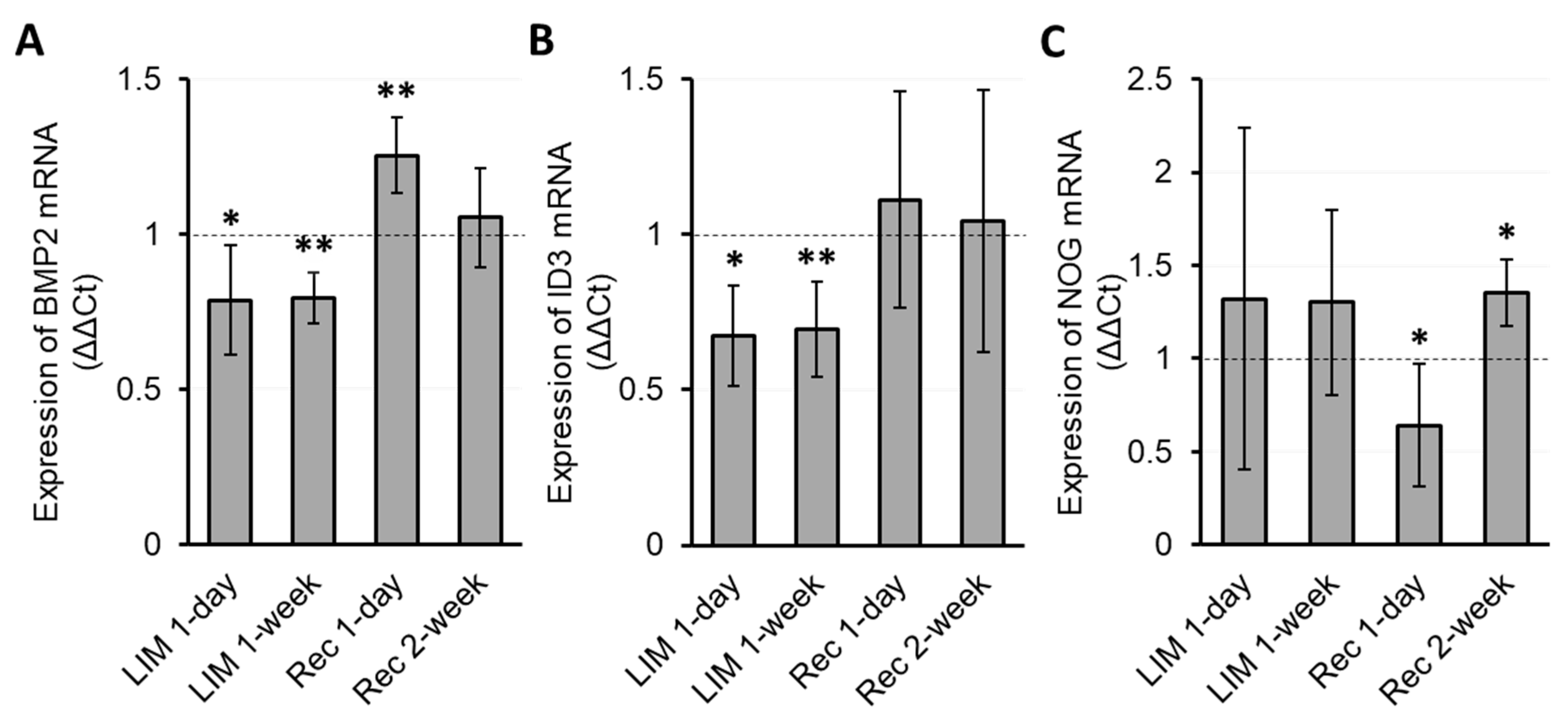
3.2.2. Myopia Induction
3.3. Temporal Relationship between Gene Expression and Biometric Changes
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dolgin, E. The myopia boom. Nature 2015, 519, 276–278. [Google Scholar] [CrossRef] [PubMed]
- Baird, P.N.; Saw, S.-M.; Lanca, C.; Guggenheim, J.A.; Smith III, E.L.; Zhou, X.; Matsui, K.-O.; Wu, P.-C.; Sankaridurg, P.; Chia, A.; et al. Myopia. Nat. Rev. Dis. Prim. 2020, 6, 99. [Google Scholar] [CrossRef] [PubMed]
- Sankaridurg, P.; Tahhan, N.; Kandel, H.; Naduvilath, T.; Zou, H.; Frick, K.D.; Marmamula, S.; Friedman, D.S.; Lamoureux, E.; Keeffe, J.; et al. IMI Impact of Myopia. Investig. Ophthalmol. Vis. Sci. 2021, 62, 2. [Google Scholar] [CrossRef] [PubMed]
- Morgan, I.G.; French, A.N.; Ashby, R.S.; Guo, X.; Ding, X.; He, M.; Rose, K.A. The epidemics of myopia: Aetiology and prevention. Prog. Retin. Eye Res. 2018, 62, 134–149. [Google Scholar] [CrossRef] [PubMed]
- Wallman, J.; Winawer, J. Homeostasis of eye growth and the question of myopia. Neuron 2004, 43, 447–468. [Google Scholar] [CrossRef] [PubMed]
- Troilo, D.; Nickla, D.L.; Wildsoet, C.F. Choroidal thickness changes during altered eye growth and refractive state a primate. Investig. Ophthalmol. Vis. Sci. 2000, 41, 1249–1258. [Google Scholar]
- Nickla, D.L.; Wallman, J. The multifunctional choroid. Prog. Retin. Eye Res. 2010, 29, 144–168. [Google Scholar] [CrossRef]
- Zhang, Y.; Wildsoet, C.F. RPE and choroid mechanisms underlying ocular growth and myopia. Prog. Mol. Biol. Transl. Sci. 2015, 134, 221–240. [Google Scholar] [CrossRef]
- Troilo, D.; Smith, E.L.; Nickla, D.L.; Ashby, R.; Tkatchenko, A.V.; Ostrin, L.A.; Gawne, T.J.; Pardue, M.T.; Summers, J.A.; Kee, C.S.; et al. Imi—Report on experimental models of emmetropization and myopia. Investig. Ophthalmol. Vis. Sci. 2019, 60, M31–M88. [Google Scholar] [CrossRef]
- Strauss, O. The Retinal Pigment Epithelium in Visual Function. Physiol. Rev. 2005, 85, 845–881. [Google Scholar] [CrossRef]
- Goto, S.; Onishi, A.; Misaki, K.; Yonemura, S.; Sugita, S.; Ito, H.; Ohigashi, Y.; Ema, M.; Sakaguchi, H.; Nishida, K.; et al. Neural retina-specific Aldh1a1 controls dorsal choroidal vascular development via Sox9 expression in retinal pigment epithelial cells. eLife 2018, 7, 225–234. [Google Scholar] [CrossRef] [PubMed]
- Rymer, J.; Wildsoet, C.F. The role of the retinal pigment epithelium in eye growth regulation and myopia: A review. Vis. Neurosci. 2005, 22, 251–261. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, S.I.; Kurihara, T.; Jiang, X.; Miwa, Y.; Lee, D.; Serizawa, N.; Jeong, H.; Mori, K.; Katada, Y.; Kunimi, H.; et al. Scleral PERK and ATF6 as targets of myopic axial elongation of mouse eyes. Nat. Commun. 2022, 13, 5859. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Chen, W.; Zhao, F.; Zhou, Q.; Reinach, P.S.; Deng, L.; Ma, L.; Luo, S.; Srinivasalu, N.; Pan, M.; et al. Scleral hypoxia is a target for myopia control. Proc. Natl. Acad. Sci. USA 2018, 115, E7091–E7100. [Google Scholar] [CrossRef]
- He, L.; Frost, M.R.; Siegwart, J.T.; Norton, T.T. Gene expression signatures in tree shrew choroid during lens-induced myopia and recovery. Exp. Eye Res. 2014, 123, 56–71. [Google Scholar] [CrossRef]
- Metlapally, R.; Wildsoet, C.F. Scleral Mechanisms Underlying Ocular Growth and Myopia. In Physiology & Behavior; Elsevier: Amsterdam, The Netherlands, 2015; Volume 134, pp. 241–248. [Google Scholar] [CrossRef]
- Zhou, X.; Pardue, M.T.; Iuvone, P.M.; Qu, J. Dopamine signaling and myopia development: What are the key challenges. Prog. Retin. Eye Res. 2017, 61, 60–71. [Google Scholar] [CrossRef]
- Jiang, X.; Pardue, M.T.; Mori, K.; Ikeda, S.I.; Torii, H.; D’Souza, S.; Lang, R.A.; Kurihara, T.; Tsubota, K. Violet light suppresses lens-induced myopia via neuropsin (OPN5) in mice. Proc. Natl. Acad. Sci. USA 2021, 118, e2018840118. [Google Scholar] [CrossRef]
- Zhang, P.; Zhu, H. Light Signaling and Myopia Development: A Review. Ophthalmol. Ther. 2022, 11, 939–957. [Google Scholar] [CrossRef]
- Zhang, Y.; Wildsoet, C.F. The RPE in Myopia Development. In Retinal Pigment Epithelium in Health and Disease; Springer International Publishing: Cham, Switzerland, 2020; Volume 10, pp. 117–138. ISBN 9783030283834. [Google Scholar]
- Heermann, S.; Schütz, L.; Lemke, S.; Krieglstein, K.; Wittbrodt, J. Eye morphogenesis driven by epithelial flow into the optic cup facilitated by modulation of bone morphogenetic protein. eLife 2015, 4, e05216. [Google Scholar] [CrossRef]
- Huang, J.; Liu, Y.; Filas, B.; Gunhaga, L.; Beebe, D.C. Negative and positive auto-regulation of BMP expression in early eye development. Dev. Biol. 2015, 407, 256–264. [Google Scholar] [CrossRef]
- Goto, S.; Muroy, S.E.; Zhang, Y.; Saijo, K.; Kolora, S.R.R.; Zhu, Q.; Wildsoet, C.F. Gene Expression Signatures of Contact Lens-Induced Myopia in Guinea Pig Retinal Pigment Epithelium. Investig. Ophthalmol. Vis. Sci. 2022, 63, 25. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, Y.; Wildsoet, C.F. Bidirectional, optical sign-dependent regulation of BMP2 gene expression in chick retinal pigment epithelium. Investig. Ophthalmol. Vis. Sci. 2012, 53, 6072–6080. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Phan, E.; Wildsoet, C.F. Retinal defocus and form-deprivation exposure duration affects RPE BMP gene expression. Sci. Rep. 2019, 9, 7332. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Goto, S.; Singh, S.; Torres, J.A.; Wildsoet, C.F. Daily or less frequent topical 1% atropine slows defocus-induced myopia progression in contact lens-wearing guinea pigs. Transl. Vis. Sci. Technol. 2022, 11, 26. [Google Scholar] [CrossRef] [PubMed]
- Goto, S.; Frost, M.; Wildsoet, C.F. Isolation of Retinal Pigment Epithelial Cells from Guinea Pig Eyes. J. Vis. Exp. 2023. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein–protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef]
- Howlett, M.H.C.; McFadden, S.A. Spectacle lens compensation in the pigmented guinea pig. Vis. Res. 2009, 49, 219–227. [Google Scholar] [CrossRef]
- Wallman, J.; Adams, J.I. Developmental aspects of experimental myopia in chicks: Susceptibility, recovery and relation to emmetropization. Vis. Res. 1987, 27, 1139–1163. [Google Scholar] [CrossRef]
- Zhou, X.; Lu, F.; Xie, R.; Jiang, L.; Wen, J.; Li, Y.; Shi, J.; He, T.; Qu, J. Recovery from axial myopia induced by a monocularly deprived facemask in adolescent (7-week-old) guinea pigs. Vis. Res. 2007, 47, 1103–1111. [Google Scholar] [CrossRef]
- Mcbrien, N.A.; Lawlor, P.; Gentle, A. Recovery from Axial Myopia in the Tree Shrew. Investig. Ophthalmol. Vis. Sci. 2000, 41, 3713–3719. [Google Scholar]
- Qiao-Grider, Y.; Hung, L.F.; Kee, C.S.; Ramamirtham, R.; Smith, E.L. Recovery from form-deprivation myopia in rhesus monkeys. Investig. Ophthalmol. Vis. Sci. 2004, 45, 3361–3372. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Zhang, G.; Zhou, X.; Xu, R.; Wang, S.; Guan, Z.; Lu, J.; Srinivasalu, N.; Shen, M.; Jin, Z.; et al. Changes in Choroidal Thickness and Choroidal Blood Perfusion in Guinea Pig Myopia. Investig. Ophthalmol. Vis. Sci. 2019, 60, 3074–3083. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Li, S.; Wang, B.; Lin, X.; Wu, Y.; Liu, H.; Qu, X.; Dai, J.; Zhou, X.; Zhou, H. Scleral cross-linking using riboflavin UVA irradiation for the prevention of myopia progression in a guinea pig model: Blocked axial extension and altered scleral microstructure. PLoS ONE 2016, 11, e0165792. [Google Scholar] [CrossRef] [PubMed]
- El-Nimri, N.W.; Yao, M.; Huerta, A.; Hoang, M.; Wildsoet, C.F. Effect of chronic topical latanoprost on the sclera and lamina cribrosa of form-deprived myopic guinea pigs. Exp. Eye Res. 2019, 186, 107740. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, M.; Ito, Y.; Takahashi, A.; Kawano, K.; Terasaki, H. Scleral thickness in highly myopic eyes measured by enhanced depth imaging optical coherence tomography. Eye 2013, 27, 410–417. [Google Scholar] [CrossRef]
- Curtin, B.; Teng, C. Scleral changes in pathological myopia. Trans Am Acad Ophthalmol Otolaryngol. 1958, 62, 777–788. [Google Scholar]
- Zhang, Y.; Azmoun, S.; Hang, A.; Zeng, J.; Eng, E.; Wildsoet, C.F. Retinal defocus and form-deprivation induced regional differential gene expression of bone morphogenetic proteins in chick retinal pigment epithelium. J. Comp. Neurol. 2020, 528, 2864–2873. [Google Scholar] [CrossRef]
- Shelton, L.; Troilo, D.; Lerner, M.R.; Gusev, Y.; Brackett, D.J.; Rada, J.S. Microarray analysis of choroid/RPE gene expression in marmoset eyes undergoing changes in ocular growth and refraction. Mol. Vis. 2008, 14, 1465–1479. [Google Scholar]
- Geng, C.; Li, Y.; Guo, F.; Wang, J.; Yue, Y.; Zhou, K.; Wei, R.; Zhang, Y. RNA sequencing analysis of long non-coding RNA expression in ocular posterior poles of guinea pig myopia models. Mol. Vis. 2020, 26, 117–134. [Google Scholar]
- Lamb, T.M.; Knecht, A.K.; Smith, W.C.; Stachel, S.E.; Economides, A.N.; Stahl, N.; Yancopolous, G.D.; Harland, R.M. Neural induction by the secreted polypeptide noggin. Science 1993, 262, 713–718. [Google Scholar] [CrossRef]
- Hirsinger, E.; Duprez, D.; Jouve, C.; Malapert, P.; Cooke, J.; Pourquié, O. Noggin acts downstream of Wnt and Sonic Hedgehog to antagonize BMP4 in avian somite patterning. Development 1997, 124, 4605–4614. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, K.; Takizawa, T.; Ochiai, W.; Yanagisawa, M.; Hisatsune, T.; Nakafuku, M.; Miyazono, K.; Kishimoto, T.; Kageyama, R.; Taga, T. BMP2-mediated alteration in the developmental pathway of fetal mouse brain cells from neurogenesis to astrocytogenesis. Proc. Natl. Acad. Sci. USA 2001, 98, 5868–5873. [Google Scholar] [CrossRef]
- Dudley, A.T.; Lyons, K.M.; Robertson, E.J. A requirement for bone morphogenetic protein-7 during development of the mammalian kidney and eye. Genes Dev. 1995, 9, 2795–2807. [Google Scholar] [CrossRef] [PubMed]
- Hogan, B.L.M. Bone morphogenetic proteins in development. Curr. Opin. Genet. Dev. 1996, 6, 432–438. [Google Scholar] [CrossRef] [PubMed]
- Sakuta, H.; Takahashi, H.; Shintani, T.; Etani, K.; Aoshima, A.; Noda, M. Role of bone morphogenic protein 2 in retinal patterning and retinotectal projection. J. Neurosci. 2006, 26, 10868–10878. [Google Scholar] [CrossRef]
- Feldkaemper, M.; Schaeffel, F. An updated view on the role of dopamine in myopia. Exp. Eye Res. 2013, 114, 106–119. [Google Scholar] [CrossRef]
- Li, H.H.; Sun, Y.L.; Cui, D.M.; Wu, J.; Zeng, J.W. Effect of dopamine on bone morphogenesis protein-2 expression in human retinal pigment epithelium. Int. J. Ophthalmol. 2017, 10, 1370–1373. [Google Scholar] [CrossRef]
- Yan, Z.; Liu, Y.; Abraham, H.; Eileen, P.; Christine, F.W. Differential gene expression of BMP2 and BMP receptors in chick retina & choroid induced by imposed optical defocus. Vis. Neurosci. 2016, 33, E015. [Google Scholar] [CrossRef]
- Li, H.; Wu, J.; Cui, D.; Zeng, J. Retinal and choroidal expression of BMP-2 in lens-induced myopia and recovery from myopia in Guinea pigs. Mol. Med. Rep. 2016, 13, 2671–2676. [Google Scholar] [CrossRef]
- Jiang, L.; Garcia, M.B.; Hammond, D.; Dahanayake, D.; Wildsoet, C.F. Strain-dependent differences in sensitivity to myopia-inducing stimuli in guinea pigs and role of choroid. Investig. Ophthalmol. Vis. Sci. 2019, 60, 1226. [Google Scholar] [CrossRef]
- Xiao, H.; Fan, Z.Y.; Tian, X.D.; Xu, Y.C. Comparison of form-deprived myopia and lens-induced myopia in guinea pigs. Int. J. Ophthalmol. 2014, 7, 245–250. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Cui, D.; Zhao, F.; Huo, L.; Hu, J.; Zeng, J. BMP-2 Is involved in scleral remodeling in myopia development. PLoS ONE 2015, 10, e0125219. [Google Scholar] [CrossRef] [PubMed]
- Harrison, J.M.; Irving, R. Visual and Nonvisual Auditory Systems in Mammals: Anatomical evidence indicates two kinds of auditory pathways and suggests two kinds of hearing in mammals. Science 1966, 154, 738–743. [Google Scholar] [CrossRef] [PubMed]
- Hughes, A. A quantitative analysis of the cat retinal ganglion cell topography. J. Comp. Neurol. 1975, 163, 107–128. [Google Scholar] [CrossRef]
- Peichl, L. Diversity of Mammalian Photoreceptor Properties: Adaptations to Habitat and Lifestyle? Anat. Rec. A Discov. Mol. Cell. Evol. Biol. 2005, 287, 1001–1012. [Google Scholar] [CrossRef]
- Zaghloul, K.A.; Boahen, K.; Demb, J.B. Different circuits for ON and OFF retinal ganglion cells cause different contrast sensitivities. J. Neurosci. 2003, 23, 2645–2654. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Measurement Timepoints | |||||
|---|---|---|---|---|---|
| Parameters | 1–3 Days LIM | 1 Week LIM | 1–3 Days Rec | 1 Week Rec | 2 Weeks Rec |
| Axial length | ↑ | ↑↑ | ↓ | → | → |
| Choroidal thickness | → | ↓ | ↑ | ↑↑ | → |
| BMP2 expression | ↓ | ↓ | ↑ | N/A | → |
| ID3 expression | ↓ | ↓ | → | N/A | → |
| NOG expression | → | → | ↓ | N/A | ↑ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Goto, S.; Zhang, Y.; Vyas, S.A.; Zhu, Q.; Wildsoet, C.F. Changes in Expression in BMP2 and Two Closely Related Genes in Guinea Pig Retinal Pigment Epithelium during Induction and Recovery from Myopia. Biomolecules 2023, 13, 1373. https://doi.org/10.3390/biom13091373
Goto S, Zhang Y, Vyas SA, Zhu Q, Wildsoet CF. Changes in Expression in BMP2 and Two Closely Related Genes in Guinea Pig Retinal Pigment Epithelium during Induction and Recovery from Myopia. Biomolecules. 2023; 13(9):1373. https://doi.org/10.3390/biom13091373
Chicago/Turabian StyleGoto, So, Yan Zhang, Sonal Aswin Vyas, Qiurong Zhu, and Christine F. Wildsoet. 2023. "Changes in Expression in BMP2 and Two Closely Related Genes in Guinea Pig Retinal Pigment Epithelium during Induction and Recovery from Myopia" Biomolecules 13, no. 9: 1373. https://doi.org/10.3390/biom13091373