
Molecular Mechanism of Oocyte Activation in Mammals: Past, Present, and Future Directions

1
Laboratory of Animal Reproduction, Graduate School of Veterinary Sciences, Azabu University, Sagamihara 252-5201, Japan
2
School of Veterinary Medicine, Azabu University, Sagamihara 252-5201, Japan
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Biomolecules 2024, 14(3), 359; https://doi.org/10.3390/biom14030359
Submission received: 14 February 2024
/
Revised: 11 March 2024
/
Accepted: 14 March 2024
/
Published: 17 March 2024
(This article belongs to the Section Molecular Reproduction)
Abstract
:During mammalian fertilization, repetitive intracellular Ca2+ increases known as Ca2+ oscillations occur. These oscillations are considered crucial for successful fertilization and subsequent embryonic development. Numerous researchers have endeavored to elucidate the factors responsible for inducing Ca2+ oscillations across various mammalian species. Notably, sperm-specific phospholipase C zeta (PLCζ) emerged as a prominent candidate capable of initiating Ca2+ oscillations, particularly in mammals. Genetic mutation of PLCζ in humans results in the absence of Ca2+ oscillations in mouse oocytes. Recent studies further underscored PLCζ’s significance, revealing that sperm from PLCζ-deficient (Plcz1−/−) mice fail to induce Ca2+ oscillations upon intracytoplasmic sperm injection (ICSI). Despite these findings, observations from in vitro fertilization (IVF) experiments using Plcz1−/− sperm revealed some residual intracellular Ca2+ increases and successful oocyte activation, hinting at potential alternative mechanisms. In this review, we introduced the current hypothesis surrounding oocyte activation in mammals, informed by contemporary literature, and probed into the enigmatic mechanisms underlying mammalian fertilization-induced oocyte activation.
1. The Discovery of PLCζ as a Factor Inducing Oocyte Activation
1.1. Essential Events of Mammalian Fertilization
Mammalian fertilization heralds a pivotal moment in the inception of life, which requires the ovulated oocyte and sperm [1]. Throughout this process, the sperm embark on a journey through the vaginal canal and uterus, ultimately reaching the oviductal ampullae, which serves as the primary site of fertilization in most mammalian species [2,3]. Meanwhile, oocytes expelled from the ovary remain arrested at metaphase during the second meiosis (MII), awaiting fertilization [4,5]. Preceding fertilization, sperm undergo a series of physiological and morphological transformations, including capacitation, the acrosome reaction (AR), and hyperactivation [6].
After the physiological and morphological transformations, the sperm trigger “oocyte activation”, which includes the exocytosis of cortical granules (CGs), resumption from the MII arrest, extrusion of the second polar body, and pronuclear formation (PN) [7,8,9]. Failure to induce oocyte activation at the time of fertilization results in the absence of the aforementioned biochemical and morphological changes, thereby inhibiting subsequent embryonic development [10]. Successful fertilization necessitates the fusion of a single sperm with a single oocyte [11]. The entry of two or more sperm into the oocyte leads to polyspermy [11]. Generally, both the zona reaction and a membrane block function prevent polyspermy [3].
Upon penetration of the oocyte, sperm induce the exocytosis of CGs [12]. The released CGs modify the protein structure within the zona pellucida, leading to the zona reaction [12] (Figure 1). Furthermore, physiological alterations to the oocyte’s membrane following sperm penetration serve as a barrier to prevent multiple sperm from entering the oocyte cytoplasm [13]. Polyspermy during fertilization results in the formation of multiple pronuclei and the generation of polyploid embryos, which typically fail to develop or perish shortly after implantation [14]. The arrest at the metaphase II (MII) stage is regulated by the cytostatic factor, which maintains the high activity of the maturation-promoting factor (MPF) consisting of cyclin-dependent protein kinase 1 and cyclin B [10]. Upon sperm entry into the oocyte during fertilization, cyclin B undergoes degradation, leading to the inactivation of MPF [10]. Consequently, chromosome decondensation ensues, facilitating pronuclear formation [15]. The inactivation of MPF requires calcium ions (Ca2+) for oocyte activation, as a rise in Ca2+ levels leads to MPF inactivation through the activation of Ca2+/calmodulin-dependent protein kinase gamma (CamKIIγ) and the degradation of early mitotic inhibitor 2 (Emi2) [16]. Therefore, an increase in Ca2+ ions within the oocyte is considered indispensable for oocyte activation during fertilization [16].
1.2. Ca2+ Oscillations
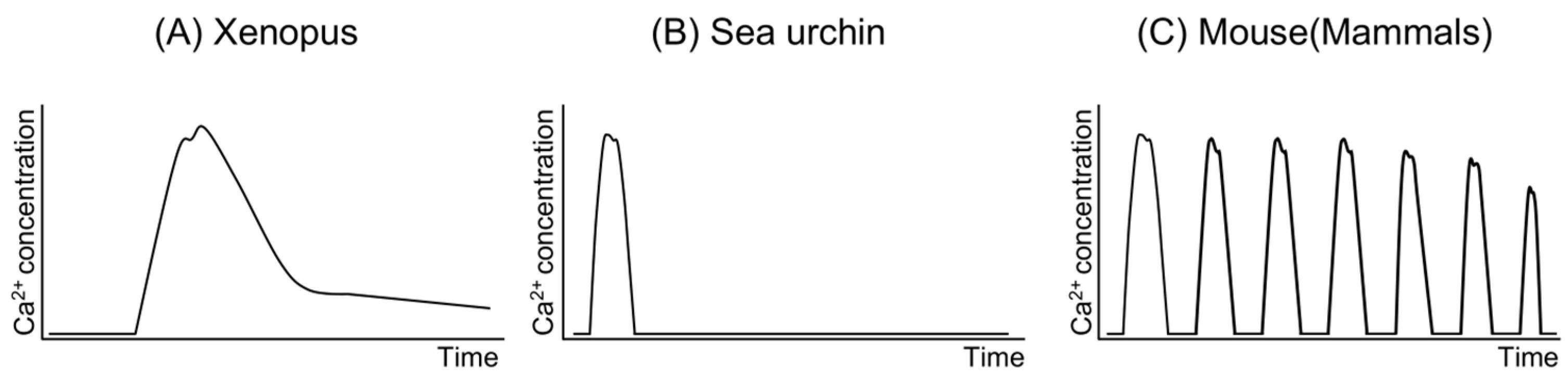
Rises in Ca2+ concentration within the oocyte are observed during fertilization across all species investigated to date [17]. These Ca2+ increases occur as a single wave in some species, such as Xenopus and sea urchins [18,19,20,21], while in others, they manifest as repetitive waves [22,23,24] (Figure 2). In mammals like humans and mice, oocytes exhibit repetitive Ca2+ increases, known as “Ca2+ oscillations”, immediately following sperm–oocyte fusion [22,25,26,27], with variations in duration, amplitude, and frequency observed among species [26]. In mouse oocytes, the typical Ca2+ concentration is approximately 100 nM, peaking at just under 1000 nM, with each increase lasting approximately 0.5 min [25,28]. Ca2+ oscillations persist for about 4–5 h, ceasing around pronuclear formation [29,30,31]. Consequently, Ca2+ oscillations are regarded as a pivotal event for oocyte activation, particularly in mammals. Numerous researchers have endeavored to elucidate the mechanisms underlying Ca2+ oscillations [32,33]. Studies have demonstrated that a reduction in the number of Ca2+ oscillations during fertilization in mice results in aberrant gene expression in blastocysts and abnormal postnatal growth of offspring following embryo transfer of the blastocyst [34]. Nevertheless, whether Ca2+ oscillations are indispensable for mammalian fertilization remains unclear.
The significance of Ca2+ during mammalian fertilization has been underscored through various experiments. For instance, treatment with the Ca2+ chelator 1,2-bis-(O-aminophenoxy)-ethane-N,N,N’,N’,-tetraacetic acid (BAPTA) prevented the elevation of Ca2+ levels in mouse oocytes, resulting in inhibition of CG exocytosis, meiotic resumption, and pronuclear (PN) formation [25]. Conversely, treatments of mouse oocytes with Ca2+ ionophores, ethanol, or electrical stimulation led to an increase in Ca2+ concentration within the oocytes, inducing oocyte activation [35,36,37]. Following these treatments, oocytes were capable of forming PN and subsequently developing into blastocysts. These experiments unequivocally demonstrated that the elevation of Ca2+ levels in the oocyte is adequate to trigger oocyte activation in mammals. However, the activation rate of oocytes was not notably high [23,24], possibly due to the transient nature of the Ca2+ increase rather than the repetitive Ca2+ oscillations characteristic of mammalian fertilization [38]. Thus, these experiments established that in mice, repetitive increases in Ca2+ are indispensable for oocyte activation and subsequent embryonic development.
1.3. The Sperm Factor Theory and the Discovery of PLCζ
Three hypotheses have been proposed regarding the mechanism of Ca2+ oscillations in the oocyte during fertilization (Figure 3). The first hypothesis, known as the “Sperm ligand theory”, suggests that receptors on the oocyte membrane bind to ligands on the sperm, leading to an increase in Ca2+. This process is believed to involve G proteins that activate phospholipase C isoforms (PLCs) within the oocyte [39]. The second hypothesis, termed the “Sperm conduit theory”, posits that extracellular Ca2+ enters the oocyte through the sperm when the sperm membrane fuses with the oocyte membrane [19,40]. However, the success of intracytoplasmic sperm injection (ICSI) has challenged these two hypotheses, as ICSI does not involve the fusion of sperm and oocyte membranes during fertilization.
The third hypothesis, known as the “sperm factor (SF) theory”, is widely considered a plausible explanation for the induction of Ca2+ oscillations. It has been demonstrated that immediately following the fusion of sperm and oocyte, soluble activating factors are released from the sperm into the oocyte cytoplasm [17,41,42,43]. Supporting evidence for the SF theory comes from studies showing that injection of soluble extracts from sperm into mammalian oocytes induces Ca2+ oscillations similar to those observed during fertilization [42,44]. The sperm factor appears to be a protein [42], as evidenced by studies showing that injection of soluble sperm extract activates mouse oocytes, leading to their development at least to the blastocyst stage [43]. Importantly, this sperm factor is not species-specific among mammals [44]. Further supporting this hypothesis, the injection of soluble sperm extracts into oocytes has been shown to elicit a rise in Ca2+ levels similar to that observed during fertilization across various species, including sea urchins [45,46]. Thus, it is proposed that one or more SFs exist in the sperm, and these SF(s) may serve as the trigger for inducing the increase in Ca2+ levels and subsequent oocyte activation. While various candidate factors have been reported, subsequent studies have shown that oscillin and tr-kit, initially identified as SFs, do not fulfill this role [47,48]. However, it was later revealed that sperm extracts contain a sperm-specific phospholipase C (PLC), which exhibits distinctive properties such as higher Ca2+ sensitivity compared to known PLC isoforms [49,50].
A novel sperm-specific PLC named phospholipase C zeta (PLCζ) (gene name: Plcz1) was eventually identified [51], and it is considered to have all of the requirements of an SF for oocyte activation. PLCζ has been detected in mice [51,52], rats [53], humans [53,54], macaque monkeys [54], cows [55], pigs [56,57], horse [58,59], quail [60], and medaka (Japanese rice fish) [53]. Plcz1 is a gene that is widely conserved among animal species, and it is now known that PLCζ hydrolyzes phosphatidylinositol 4,5-bisphosphate (PIP2) to IP3 [51,61], which binds to the receptor of IP3 (i.e., IP3R) on the endoplasmic reticulum (ER) and releases Ca2+ stored in the ER into the oocyte cytoplasm, causing an increase in Ca2+ in the oocyte at fertilization [31,62,63].
Microinjection of a small amount of Plcz1 mRNA (0.02 mg/mL) into mouse oocytes induced Ca2+ oscillations similar to those observed during IVF, and 62% of the embryos reached the morulae/blastocyst state [51]. The amount of PLCζ that can induce Ca2+ oscillations (4–75 fg, i.e., 0.002–0.02 mg/mL cRNA) is in a range that is similar to that of the PLCζ protein in a single sperm (20–50 fg) [51]. On the other hand, when transgenic mice suppressing the expression of PLCζ mRNA were generated, PLCζ protein was reduced in their sperm, resulting in reduced Ca2+ oscillations activity and oocyte activation [64]. These findings suggest that PLCζ plays important roles in oocyte activation and embryonic development during mammalian fertilization.
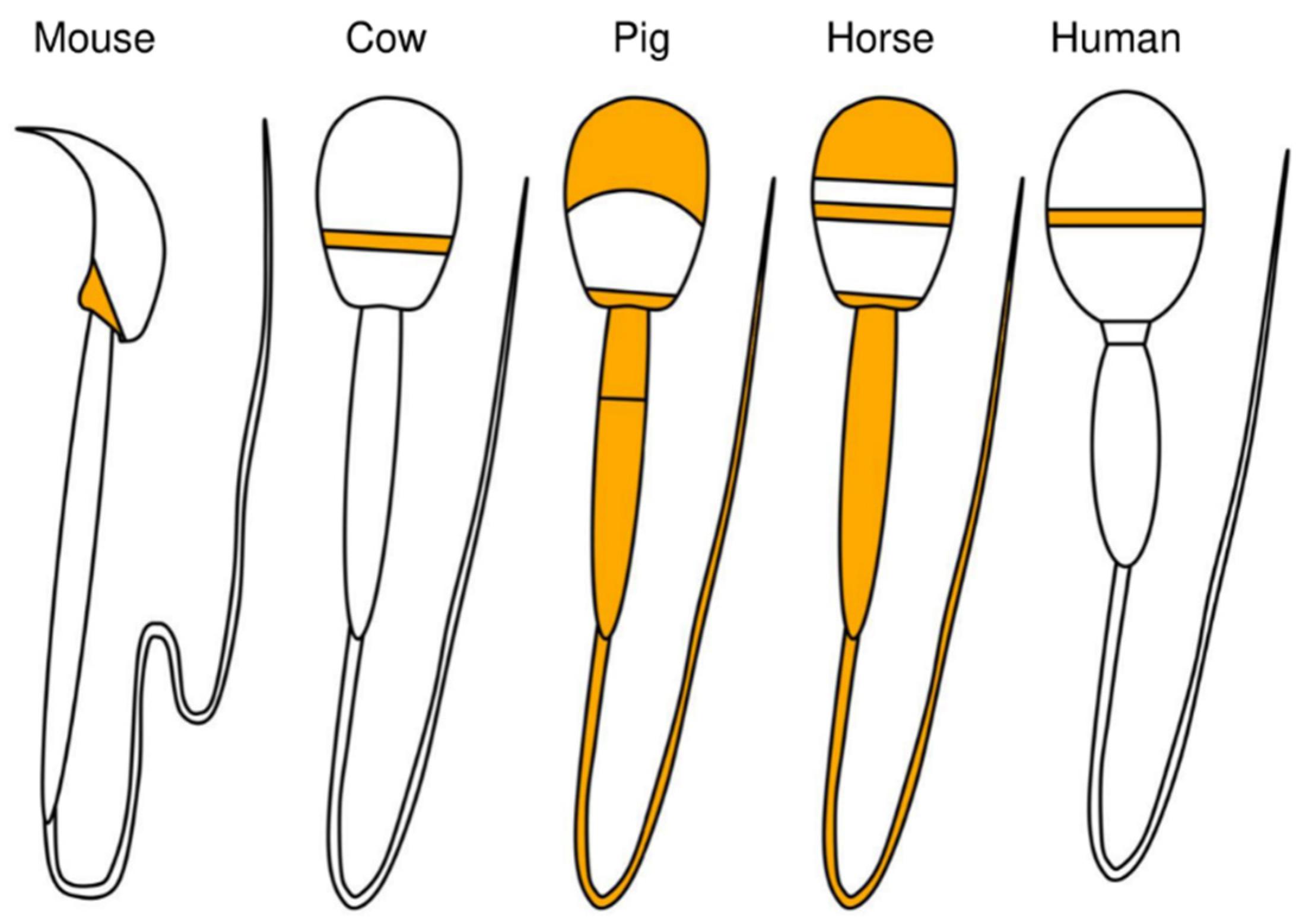
Regarding the localization of PLCζ in mice, PLCζ was shown to be located in the post-acrosomal region of the sperm [65] (Figure 4). In bovine sperm, PLCζ was observed in the equatorial region [65]; in porcine sperm, PLCζ was observed in both acrosomal and post-acrosomal regions and the tail [56,66]; and in horse sperm, PLCζ was observed in the apical body, equatorial region, head–midpiece junction, and tail, indicating differences among animal species [58,59]. It was reported that changes in the localization of PLCζ in sperm are influenced by phenomena such as AR. AR is an exocytotic process that is essential for mammalian fertilization and is critical for the release of acrosomal contents into the oviduct and for the process that allows sperm to interact and fuse with the oocyte [67]. In humans, after fertilization or AR, the proportion of sperm with PLCζ protein localized to the equatorial/acrosomal region decreased, and the proportion with PLCζ localized to the posterior equatorial/acrosomal region increased [68]. It is not clear whether PLCζ protein actually moves to the posterior acrosomal region during fertilization and/or AR. Taken together, the above-described findings indicate that the localization of PLCζ in sperm, which varies among animal species and fertilization events, may be involved in species-specific fertilization mechanisms.
2. The Functions of PLCζ
2.1. The Molecular Structure of PLCs
Six isotypes (PLCβ, PLCγ, PLCδ, PLCε, PLCζ, and PLCη) and 13 subtypes (β1-4, γ1-2, δ1, δ3-4, ε1, ζ1, and η1-2) of PLCs are currently known in mammals (Figure 5) [69,70]. The structures of these PLCs have in common the pleckstrin-homology (PH domain) (except for PLCζ), an EF-hand domain, an X-Y catalytic domain, and a C2 domain [30,51]. The PH domain is composed of seven β-sheets and an α-helix, and the β-sheet of the PH domain binds to PIP2 [71,72]. The β-sheet of the PH domain has been shown to bind to PIP2 [71]. The PH domain is a structure that plays an important role in the binding of PLC to PIP2 and G proteins at the plasma membrane [73]. The EF-hand motif contains a helix–loop–helix structure, which is also present in numerous Ca2+-binding proteins such as myosin, calmodulin, calreticulin, and troponin [74]. After binding with Ca2+, the EF-hand motif alters its conformation and triggers Ca2+-regulatory functions by disclosing the ligand sites on other proteins. This process stabilizes the PLC structure [75]. Irrespective of the concentration of Ca2+, removal of the EF-hand motif from the enzyme diminishes the function of PLC [76]. When Ca2+ binds to the EF-hand motif, it triggers the binding of PLC to PIP2 through the PH domain [72,76]. The catalytic X-Y domain comprises approximately 300 amino acids and is situated on the C-terminus of the EF-hand motif [72]. These domains, also known as triosephosphate isomerase (TIM) barrel domains, are composed of alternating α-helices and β-sheets, forming a βaβaβaβ pattern with a TIM barrel-like structure [72]. The X region, which harbors all catalytic residues, is moderately conserved among members of the PLC family [72,77]. The X and Y catalytic domains, also known as the TIM barrel-like domain, are responsible for the catalytic activity of PLC [69,78]. According to a study using two cyclic inositol 4,5-bisphosphate, the catalytic activity of this domain increases as the concentration of Ca2+ increases from 0.01 μM to 10 μM [78]. The Y domain, which comprises residues 489–606, forms the other half of the TIM barrel-like structure [79]. The eightfold barrel structure is found in enzymes that regulate metabolism, with diverse functions [79]. The Y domain plays a critical role in substrate recognition, regulating PLCs’ preference for PIP2, PIP, and PI [69]. The X-Y domains of PLC isoforms are separated by an X-Y linker sequence [54], which is believed to be essential. The X-Y linker sequence of PLC is negatively charged, maintaining the inactive state of the PLC [51]. Upon stimulation, PLC translocates to the plasma membrane [80]. The negatively charged plasma membrane and X-Y linker sequence repel each other, leading to structural changes in PLC and enabling its binding to PIP2 [80].
The C2 domain, found at the C-terminus, consists of approximately 120 amino acids [81] and is present in more than 40 proteins, including protein kinase C [72]. The eight-stranded antiparallel beta sandwich forms the C2 domain in PLC family members [72]. The presence of multiple binding sites for phospholipids, dependent on Ca2+ in the C2 domain, suggests that the binding sites work synergistically [82]. The EF-hand motif possesses a Ca2+-binding pocket formed by successive stretches of amino acids in the primary sequence [83,84]. Conversely, the Ca2+-binding pockets of the C2 domain reside far apart from each other in the amino acid sequence associated with the C2 domain and are separate from each other [83,84]. Functionally, the EF-hand motif, considered the most prevalent Ca2+-binding motif in proteins, could potentially compete with the C2 domain for Ca2+ binding [85].
Figure 5.
Schematic diagram of PLC domains in mammals. Different color indicates different domain.

2.2. Features of PLCζ
PLCζ is the smallest PLC of all PLCs studied to date and maintains high hydrolytic ability (Figure 5). Its unique structure and properties distinguish it from other members of the PLC family, which we will discuss later in this chapter. One notable feature of PLCζ is its absence of a PH domain, distinguishing it from all other members of the PLC family [51]. Although the PH domain enables PLCs to link with components of cell membranes, including PIP2 and G proteins [71], PLCζ moves into the cytoplasm and binds with PIP2 situated within tiny cellular sacs, thereby enhancing its remarkable ability to break down PIP2 molecules after the fusion of sperm and oocyte [86]. PLCs, except PLCζ, perform membrane targeting of PIP2 through the PH domain; PLCζ lacks the PH domain, and membrane targeting of PIP2 is performed by the EF-hand domain. This was demonstrated by the fact that the introduction of an EF-hand domain mutant PLCζ protein into mouse oocytes reduced PIP2 hydrolysis without affecting Ca2+ sensitivity [87].
In addition, compared to other PLCs, PLCζ has high activity even at very low Ca2+ concentrations [30]. Ca2+ sensitivity is largely attributed to the EF-hand domain, which comprises four motifs located at the N-terminus of PLCζ and is primarily responsible for Ca2+ regulation [88,89]. The deletion of EF1, EF2, and EF1-2 had no effect on Ca2+ sensitivity, but the EF1-3 deletion in PLCζ mutants required 12 times more Ca2+ than PLCζ to degrade PIP2 as much as PLCζ degrades PIP2; in addition, a marked decrease in Ca2+ sensitivity was observed [90]. Thus, the characteristic Ca2+ sensitivity of PLCζ is likely to involve the structural determinant EF1-3. However, even the Ca2+ sensitivity of ΔEF1-3 PLCζ mutants was observed to be 15-fold higher than that of PLCδ1, suggesting that EF4 of PLCζ may also be involved in the high Ca2+ sensitivity of PLCζ [90]. EF1 and EF2 of PLCζ contain Ca2+-binding loop sequences that are homologous to the Ca2+-binding site [76]. PLCζ mutants in which Asp (an amino acid residue that is present in the Ca2+-binding loop sequence) was mutated to Arg did not show reduced Ca2+ sensitivity [90]. This result is consistent with the above-described finding that the ΔEF1, ΔEF2, and ΔEF1-2 PLCζ mutants do not reduce Ca2+ sensitivity, indicating that the EF1 and EF2 Ca2+-binding loop sequences of PLCζ are not involved in the Ca2+ sensitivity of PLCζ.
An injection of PLCζ mutant cRNA lacking one or both pairs of the EF-hand domains of PLCζ into mouse oocytes under the same conditions as those used for wildtype (WT) PLCζ significantly reduced Ca2+ oscillations [89]. In the evaluation of the rate of pronuclear formation when ΔEF1-3 PLCζ cRNA was injected into the oocyte, WT PLCζ showed a high rate of pronucleus formation at 0.002 mg/mL, but ΔEF1-3 PLCζ could hardly induce pronuclear formation. However, when 1 mg/mL of ΔEF1-3 PLCζ cRNA was injected, the rate of PN formation was similar to that of WT PLCζ [91]. These results indicate that a reduction in the EF-hand motif reduces Ca2+ sensitivity, but not completely. Thus, the EF-hand domain of PLCζ is responsible for membrane targeting of PIP2 instead of the PH domain. Although a single EF-hand motif does not cause the loss of Ca2+ sensitivity, the characteristic high Ca2+ sensitivity of PLCζ is regulated by the EF-hand domain, which consists of four EF-hand motifs.
The TIM barrel domain of PLCζ, consisting of the X and Y catalytic domains, is located between the EF-hand domain and the C2 domain and is the part that is actually responsible for enzyme activity [51]. Several cases of X-Y catalytic domain mutations have been reported in human male infertility; the X and Y catalytic domains of PLCζ are similar to sequences of other PLCs, and most of the sequences are conserved [51]. Saunders et al. [51] reported that replacing Aspartic Acid at position 210 (D210) (which is involved in the catalytic reaction at the active site residue of PLCδ1 and is conserved in the X catalytic domain of PLCζ [92,93]) with arginine (D210R) resulted in the loss of Ca2+ vibration-inducing activity. Aspartic Acid at position 210 corresponds to Aspartic Acid at position 343 in PLCδ1, and like PLCδ1, this amino acid residue is involved in the Ca2+-binding site that is responsible for enzyme activity in PLCζ [92,94]. In addition, the replacement of both the K299 and K301 of PLCζ with glutamic acid (K299A or K301A) resulted in a loss of Ca2+ vibration-inducing activity [95]. These amino acid residues correspond to K432 and K434 in PLCδ1 and are similar to the sites that interact with the substrate PIP2 [96,97], and it is possible that these mutations render interaction with PIP2 impossible. PLCζ is known to move around the nucleus when PN formation occurs after the induction of Ca2+ oscillations [98], but the base substitutions in D210R, K299A, and K301A inhibited the nuclear migration of PLCζ after PN formation. This indicates that the X catalytic domain of PLCζ has a role not only in enzymatic activity but also in nuclear translocation.
PLCζ has a long X-Y linker, i.e., an unstructured region mediated by the X and Y catalytic domains, which is one of the characteristic structures of PLCζ [69] and has been shown to play several roles [80,95,99]. The X-Y linker regions of PLCβ, PLCγ, PLCδ, and PLCη are negatively charged, whereas the linker of PLCζ is positively charged [80]. The positively charged X-Y linker sequence of PLCζ facilitates binding to the negatively charged PIP2 and is one of the factors responsible for the very high hydrolytic resolution of PLCζ [80]. In mouse experiments, the deletion of the X-Y linker of PLCζ markedly reduced the ability of PLCζ to induce Ca2+ oscillations [95]. In addition, when three of the lysines in the X-Y linker sequence were replaced with alanine (K374A, K375A, and K377A) in mice, the positive charge of the linker portion decreased, and these mutations have been shown to reduce the effectiveness of the PIP2 interaction and Ca2+ oscillations-inducing activity in vitro [99]. The X-Y linker also contains a nuclear localization sequence (NLS) for nuclear migration, which is thought to prevent long-lasting and excessive Ca2+ oscillations after pronuclear formation by inducing PLCζ into the nucleus [95]. The X-Y linker, together with the EF-hand domain, is thus considered essential for efficiently promoting PIP2 binding and maintaining the high hydrolytic resolution of PLC [80,99]. However, various functions are thought to be retained in different mammalian species, such as porcine PLCζ, which retains its biological activity even after proteolytic cleavage of the X-Y linker [100]. To elucidate the essential role of the X-Y linker of PLCζ in mammals, further studies are required.
The C2 domain of PLCζ is located at the C-terminus and is involved in the targeting of proteins to the plasma membrane. PLCs except PLCζ bind to PIP2 on the plasma membrane, but PLCζ showed minimal loss of PIP2 on the oocyte membrane even after Ca2+ oscillations, and depletion of the oocyte membrane PIP2 pool had no effect on Ca2+ oscillations. The C2 domain of PLCζ is largely responsible for this. PLCs other than PLCζ are targeted to PIP2 by the PH domain [78]. The C2 domain binds to phospholipid-containing membranes and exhibits different phospholipid selectivity [101], suggesting that PLCζ may target and specifically hydrolyze PIP2 in the plasma membrane by its C2 domain instead of its PH domain [99]. In vitro experiments have also shown that the C2 domain of PLCζ interacts with PI(3)P and PI(5)P in a Ca2+-independent manner and inhibits their binding to PI(4,5)P2 [90]. This interaction with PI(3)P and PI(5)P may play an important role in the inhibition of enzyme activity by PLCζ before fertilization [90]. Deletion of the C2 domain from PLCζ results in a loss of Ca2+ oscillations-inducing activity even though the Ca2+ sensitivity is not affected [102]. In addition, replacing the C2 domain of PLCζ with the C2 domain of PLCδ1 did not cause a release of Ca2+ [102]. This experiment also suggested that the C2 domain of PLCζ binds to PIP2 for hydrolysis. However, in the membrane-spotted arrays performed by Nomikos et al. [99], the binding of PIP2 to the C2 domain of PLCζ was very weak, while PIP2 showed strong binding to the X-Y linker. It is thus suggested that the binding of PLCζ to PIP2 is carried out mainly by the X-Y linker, although the C2 domain is also responsible.
Deletion of the C2 domain of PLCζ results in a loss of Ca2+ oscillations induction activity [102], which plays an important role in the induction of Ca2+ oscillations by PLCζ but may retain an important unknown function other than PIP2 binding. It was reported that PLCζ mutant proteins lacking the C2 domain of PLCζ or the 37 amino acid residues in the C-terminal region adjacent to the C2 domain (PLCζ 611–647) were completely unable to hydrolyze PIP2 at all in the presence of Ca2+ and completely lose PLC activity [90]. The C-terminal region (as well as the C2 domain) is essential for the induction of Ca2+ oscillations in PLC activity; the C2 domain of PLCζ is known to bind to the N-terminal EF-hand domain [95]. The binding of the C2 domain to the EF-hand domain exposes the X-Y domain of PLCζ, making the catalytic activity of PIP2 more efficient [95]. For these reasons, the C2 domain plays an important role in the regulation of the enzymatic activity of PLCζ at the appropriate time and in the high enzymatic activity due to the conservation of the C2 terminus.
2.3. Mutations and Manipulations of PLCζ
Several studies have used transgenic (Tg) and knockout mice to clarify the functions of PLCζ in mammalian fertilization. In 2005, researchers generated Plcz1-RNAi Tg mice that express Plcz1 short hairpin (sh)RNA to inhibit PLCζ function [64]. The sperm derived from the Plcz1-RNAi male mice exhibited neither morphological nor motility abnormalities, but the number of Ca2+ oscillations was greatly decreased when the sperm were used for IVF [64]. When in vivo fertilized oocytes were collected from the oviduct and cultured after mating Plcz1-RNAi male mice with WT female mice, the fusion of sperm and oocyte and the extrusion of the second polar body were similar to those of the controls, but oocyte activation did not occur, and the blastocyst development rate was reduced (control: 99% vs. Plcz1-RNAi Tg: 65%) [64]. The fertility of the Plcz1-RNAi male mice was also decreased (7.0 ± 0.6 pups/litter) compared to the controls (12.0 ± 1.3 pups/litter) [64]. In addition, offspring derived from Plcz1-RNAi male mice were not obtained [64]. These findings indicate that PLCζ plays a critical role in inducing Ca2+ oscillations and further embryonic development, at least in mice.
In 2007, Plcz1-Tg mice with a CAG promoter were generated for PLCζ overexpression [103]. The litter size was dramatically decreased in some strains derived from Plcz1-Tg male mice after mating with female mice (WT: 7.91 ± 0.29, Tg: 0.85 ± 0.348) [103]. When germinal vesicle (GV)-stage oocytes were collected from the ovaries of Plcz1-Tg female mice, the oocytes underwent spontaneous germinal vesicle breakdown (GVBD) and developed to the MII stage [103]. During the progression to the MII stage, increases in Ca2+ were observed in the oocytes [103]. The oocytes derived from PLCζ mice after mating with WT female mice showed segregation of metaphase chromosomes and the completion of meiosis, resulting in the induction of parthenogenetic activation and development to the blastocyst stage [103]. On the other hand, the ovaries of female mice mated with Plcz1-Tg males exhibited tumorigenesis [103]. From the results described above, it is apparent that the optimal timing and the optimal amount of PLCζ activity are essential for oocyte activation and further embryonic development.
Although the importance of PLCζ was demonstrated in the two aforementioned studies, it remains unclear whether PLCζ is indispensable for mammalian fertilization. Nearly two decades after the discovery of PLCζ in 2002, the generation of PLCζ gene-deficient (Plcz1−/−) mice has been reported by two different research groups [32,33]. Histological analysis of testes from Plcz1−/− male mice demonstrated that the loss of Plcz1−/− had no adverse effect on spermatogenesis [32]. Epididymal sperm from Plcz1−/− male mice also showed normal viability, motility, and acrosome reaction [32]. In summary, it has been shown that a loss of Plcz1 had no apparent effect on spermatogenesis or the ability of sperm to bind and fuse with oocytes.
They also investigated whether the application of ICSI with Plcz1−/− sperm could induce Ca2+ oscillations in the oocyte; their findings revealed that Ca2+ oscillations in the oocyte did not occur at all in the embryos that were fertilized with Plcz1−/− sperm [32,33]. These results suggest that PLCζ regulates Ca2+ oscillations in the oocyte during fertilization. The researchers also performed IVF with Plcz1−/− sperm. Unexpectedly, oocytes fertilized with Plcz1−/− sperm showed fertility that was similar to that of WT oocytes [32,33]. Plcz1−/− sperm-derived fertilized oocytes also exhibited a high rate of polyspermy [32,33]. In addition, fertilized oocytes from the oviducts of WT female mice mated with male Plcz1−/− mice were obtained, and the rate of polyspermy was assessed. The results revealed that, similar to IVF, the fertilized oocytes derived from Plcz1−/− male mice had an increased rate of polyspermy [32,33]. An increase in the number of pronuclei was observed in some oocytes fertilized by Plcz1−/− sperm, suggesting delayed fertilization and oocyte activation by Plcz1−/− sperm. The blastocyst development rate of oocytes fertilized by Plcz1−/− sperm was significantly decreased (WT 90.2%, Plcz1−/− 32.2%) [33]. These results suggest that the suppression of polyspermy is not working properly due to Plcz1 deficiency. It could have been proposed that (i) Juno, expressed on the oocyte surface, is indispensable for preventing polyspermy [104], and (ii) PLCζ may have a role in the mechanism underlying the blocking of polyspermy. Those authors also observed an improvement in the blastocyst development rate of fertilized oocytes derived from Plcz1−/− sperm by injection of Plcz1 cRNA or mRNA after ICSI [32,33]. This indicates that Plcz1 also plays an important role in embryonic development.
The number of pups per litter produced by the Plcz1−/− male mice was significantly lower than that of the WT, but the Plcz1−/− male mice were still fertile (WT 7.8 ± 0.8 vs. Plcz1−/− 3.2 ± 1.2~4.2 ± 0.6 pups/litter [32]; WT 8.9 ± 0.26 vs. Plcz1−/− 2.3 ± 0.50 pups/litter [33]). It was thus demonstrated that embryos derived from Plcz1−/− male mice were able to develop to term without Ca2+ oscillations by PLCζ, although defects such as oocyte activation failure (OAF) and polyspermy reduced the number of pups per litter. Nozawa et al. [33] also performed IVF with the use of various sperm concentrations (2, 10, and 50 × 103 sperm/mL) to further investigate the fertilization and oocyte activation potential of Plcz1−/− sperm. In Plcz1−/− sperm, activation failure was more pronounced at the lowest sperm concentration (WT 0% and Plcz1−/− 12.4% at 2 × 105/mL), and the polyspermy rate increased to about 80% at the highest sperm concentration (WT 7.6% and Plcz1−/− 82.4% at 50 × 103/mL) [33]. When the intracellular Ca2+ concentration was monitored during IVF using live imaging, all single-sperm fertilized eggs with Plcz1−/− sperm showed rise(s) in intracellular Ca2+ regardless of PN formation. However, these fertilized oocytes had a significantly reduced number of Ca2+ spikes (WT 12.0 ± 5.68 spikes vs. Plcz1−/− 2.75 ± 0.65 spikes) [33]. As the number of sperm fusing increased, the number of Ca2+ spikes increased, and more fertilized oocytes resumed the cell cycle. These results suggest that sperm have PLCζ-independent oocyte activation ability. However, a single sperm is insufficient for oocyte activation to cause resumption from the MII arrest, and incomplete oocyte activation may lead to polyspermy.
A previous study introduced two mutations into mice, i.e., H435P (a mutation corresponding to human H398P) and D210R (an enzymatic dead mutation), in order to investigate the mechanism of infertility caused by point mutations in Plcz1 reported in humans [33]. An immunoblot analysis of sperm proteins revealed that the WT and D210R PLCζ mutation exhibited an approximately 74 kDa signal, whereas an approx. 20 kDa signal was detected only in the sperm of the H435P PLCζ mice used in the experiment [33], suggesting instability of the H435P protein in vivo. These results indicated that H435P PLCζ seems to be unstable in vivo. The spermatogenesis, fertility, IVF, and ICSI results obtained with homozygous male mice with the D210R or H435P PLCζ mutation were comparable to those of Plcz1−/− male mice [33]. ICSI with these mouse sperm and the microinjection of mouse and human Plcz1 mRNA resulted in Ca2+ oscillations in the oocyte and 2PN formation [33]. These embryos transplanted into pseudopregnant mice yielded normal litters.
As described above, the importance of PLCζ in oocyte activation was demonstrated using Plcz1−/− mice by two research groups, and both groups also demonstrated that Plcz1−/− sperm caused polyspermy due to failure of the zona reaction [32,33], suggesting a new role for PLCζ. Although polyspermy has been reported in male infertility after IVF, the cause of this phenomenon has been unknown; the present results indicate a possible abnormality of PLCζ in the sperm of these individuals.
2.4. Human PLCζ
Currently, in infertility treatment, if there is no pronucleation in multiple cycles of ICSI or ROSI, it is considered to be a failure of oocyte activation [105]. In humans, male infertility has been reported due to PLC-zeta abnormalities. The abnormalities of PLCζ are thought to be one of the causes of the failure of oocyte activation [106].
Therefore, PLCζ has recently gained attention as an infertility diagnostic tool [107]. Artificial oocyte activation was the only treatment for oocyte activation failure. As mentioned above, artificial oocyte activation has been used when multiple cycles of in vitro fertilization or ICSI have failed to achieve fertilization or when low fertilization rates have been observed, and there are no tools to diagnose sperm–oocyte activation failure. By performing a fluorescence analysis of PLCζ and observing its fluorescence level and localization pattern, it is possible to predict the gene expression level and localization of PLCζ and indirectly diagnose whether artificial oocyte activation can improve sperm fertilization rates in male infertility patients. This measurement can avoid unnecessary IVF and reduce the consumption of unwanted oocytes.
Infertility caused by mutations in the PLCζ gene has also been reported in humans, and various phenotypic abnormalities have been reported with mutations. The mutation (H233L or H398P) of PLCζ found in infertile male humans altered the localization of PLCζ, although these mutations did not affect sperm morphology [104,108]. The microinjection of these mutant Plcz1 cRNAs into oocytes also resulted in a marked reduction in the number and amplitude of Ca2+ oscillations [101]. PLCζ (H233) is located in exon 6 and encodes the X catalytic domain, while H389 is located in exon 11 and encodes the Y catalytic domain [109]. The X-Y catalytic domain is the domain responsible for enzyme activity. An injection of patient sperm into mouse eggs reduced Ca2+ oscillations. This was the first finding that base substitutions in the X-Y catalytic domain in human sperm altered the localization of PLCζ in the spermatozoa [109]. They then expressed H233L and H398P PLCζ with the use of human embryonic kidney (HEK)293T cells, injected the cells into oocytes, and measured their cytoplasmic localization. They then observed that the expression levels were decreased, suggesting that genetic mutations due to base substitutions were responsible for the instability of the proteins [110].
The sperm of two infertile brothers reported by Escoffer et al. in 2016 both carried the missense mutation I489P PLCζ, and their sperm showed abnormal localization in the oocyte after ICSI and a significant decrease in Ca2+ oscillations in the oocytes [111]. PLCζ I489 is located in exon 13 and encodes the C2 domain. In addition, embryos fertilized via ICSI using the brothers’ sperm failed to develop to the two-cell stage. These events suggest that PLCζ in human spermatozoa also acts in the same way to induce Ca2+ oscillations by the hydrolysis of PIP2 localized in the plasma membrane, a feature of the specific phospholipid selection of the C2 domain performed using mice [111].
All previous Plcz1 mutations have been single missense mutations, but compound heterozygous mutations in Plcz1 were identified in an infertile male patient [112]. This patient was found to have a P420L substitution due to a base substitution in 1259C>T and an M578T residue substitution due to a base substitution in 1733T>C. Sequencing of the parental DNA of the patient revealed that the father was a carrier of 1733T>C (M578T), and the mother was a carrier of Plcz1 1259C>T (P420L).
Artificial oocyte activation procedures such as Ca2+ ionophores are performed when oocyte activation failure is observed [113]. Mutations of Plcz1 in human sperm reduce oocyte activation, indicating male infertility, and fertility was restored in most of these individuals after ICSI by an injection of Plcz1 RNA or an activation treatment such as Ca2+ ionophore and SrCl2 [113,114]. However, in 2019, Torra-Massana et al. reported that out of 12 patients with genetic mutations in Plcz1 (residue substitutions at R197H and H233L in the X catalytic domain and a deletion mutation of two bases of V326K in the X-Y linker), the sperm from three patients did not undergo mouse oocytes activation after receiving an injection of Plcz1 cRNA. This finding suggests that mutant Plcz1 may also impact the enzymatic activity of normal PLCζ [115].
Currently, 1–3% of infertile male patients are reported to have reduced oocyte activation ability [116], and sperm from these individuals fail to activate oocytes and produce pregnancy even with ICSI. Since there have been many reports of severely reduced oocyte activation ability due to Plcz1 mutations [105,117], further research on Plcz1 mutations in human sperm could improve fertility treatments for infertile couples with Plcz1 gene mutations. Table 1 and Figure 6 described the major Plcz1 mutations and their phenotypes reported in humans to date [118,119,120].

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Summary of human Plcz1 variants.
| Mutation (Protein) | Mutation (PLCZ1 Coding Sequence) | Gene Location | Protein Location | Type of Mutation | Phenotype | Authors | Date |
|---|---|---|---|---|---|---|---|
| p.I120M | c.360 C>G | Exon 4 | EF-hand | Missense | OAF | Torra-Massana et al. | 2019 [115] |
| p.C196 * | c.588 C>A | Exon 6 | X | Missense | OAF | Dai et al. | 2020 [118] |
| p.R197H | c.590 G>A | Exon 6 | X | Missense | OAF | Torra-Massana et al. | 2019 [115] |
| p.L224P | c.671 T>C | Exon 6 | X | Missense | OAF | Torra-Massana et al. | 2019 [115] |
| p.H233L | c.698 A>T | Exon 6 | X | Missense | OAF | Kashir et al. | 2011 [110] |
| p.L246F | c.736 C>T | Exon 7 | X | Missense | OAF | Dai et al. | 2020 [118] |
| p.V326K fs | c.972-973 (AG) deletion | Exon 6 | X-Y linker | Frameshift | OAF | Torra-Massana et al. | 2019 [115] |
| p.S350P | c.1048 T>C | Exon 10 | Y | Missense | OAF | Dai et al. | 2020 [118] |
| p.H398P | c.1193 C>A | Exon 11 | Y | Missense | OAF, protein instability | Heytens et al. | 2009 [108] |
| p.R412fs | c.1234 (A) del | Exon 11 | Y | Frameshift | OAF | Mu et al. | 2020 [119] |
| p.I489P | c.1465 A>T | Exon 13 | C2 | Missense | OAF | Escoffier et al. | 2016 [111] |
| p.S500L | c.1499 C>T | Exon13 | C2 | Missense | OAF | Torra-Massana et al. | 2019 [115] |
| p.R553P | c.1658 G>C | Exon 14 | C2 | Missense | OAF, protein instability | Yuan et al. | 2020 [120] |
| p.P420L | c.1259 C>T (compound) | Exon 12 | Y | Missense | OAF | Yuan et al. | 2020 [112] |
| p.M578T | c.1733 T>C (compound) | Exon 14 | C2 | Missense | OAF | Yuan et al. | 2020 [112] |
The asterisk indicates the stop codon.
Figure 6.
Summary of PLCζ mutations identified by the scientific literature. Schematic diagram of PLCζ domains with the mutation. The positions of the variants identified reported in the literature. Pink indicates compound heterozygous mutations.
Figure 6.
Summary of PLCζ mutations identified by the scientific literature. Schematic diagram of PLCζ domains with the mutation. The positions of the variants identified reported in the literature. Pink indicates compound heterozygous mutations.

3. Future Directions for a Better Understanding of Oocyte Activation
3.1. Another Candidate Factor(s) Inducing Oocyte Activation
Although PLCζ has been considered a potent SF, it remains uncertain whether it is the sole factor and whether it is essential for Ca2+ oscillations and oocyte activation. Several research groups have reported possible alternative sperm factor candidates. In 2007, Oko’s group discovered a protein in the post-acrosomal sheath region of the sperm head’s perinuclear theca (PT), and the protein was extractable under alkaline conditions [121]. This protein shares sequence homology with the N-terminal region of the WW domain-binding protein 2 (WBP2), and its C-terminal region is substantially enriched in proline [121]. Post-acrosomal sheath WW domain-binding protein (PAWP) is the designated name for the protein, which is also known as WBP2NL [121]. The impact of introducing recombinant human PAWP or alkaline PT extract into MII-arrested oocytes from porcine, cows, macaques, and Xenopus was examined [121], and the results revealed a high rate of PN formation in the oocytes after either injection, indicating that PAWP has potential as a SF for oocyte activation.
The ability of PAWP to induce Ca2+ increases during oocyte activation is a crucial factor in assessing its candidacy as a sperm factor. A study by Aarabi et al. (2009) [122] showed that Xenopus oocytes injected with recombinant human PAWP resulted in an intracellular release of Ca2+. Their subsequent research established that introducing human PAWP protein through an injection elicits Ca2+ oscillations and PN formation in mammalian oocytes (both mouse and human) [123]. These results strongly suggested that PAWP is a sperm factor, like PLCζ. Another SF candidate is extra-mitochondrial citrate synthase (eCS), which was first identified as a sperm factor in Cynops pyrrhogaster [124]. In the mouse, eCS was localized in the sperm head, and an injection of eCs mRNA induced Ca2+ oscillations in the oocyte [125], suggesting that eCS also appears to meet the requirements as an SF. Subsequent studies using gene-deficient mice have shown that PAWP and eCS are not essential for Ca2+ oscillations and oocyte activation [125,126].
However, the possibility that there may be factors involved in oocyte activation beyond PLCζ cannot be completely ruled out. Indeed, results obtained in studies of Plcz1−/− mice suggest that there may be other unknown mechanisms that induce the rise in intracellular Ca2+ because a few oscillations of intracellular Ca2+ were still observed in the oocytes when Plcz1−/− sperm were applied for IVF but not for ICSI, suggesting that the underlying mechanism requires the membrane fusion between sperm and oocyte [33]. As mentioned above, there is no doubt that the sperm factor theory is currently the main pathway for the rise(s) in Ca2+ during fertilization, but the other two theories may need to be reconsidered.
In addition, the results from the study using Plcz1−/− have led us to the possibility of the presence of not only “sperm factor” but “Oocyte factor” for membrane fusion-mediated Ca2+ oscillations. This is because Ca2+ oscillations require specific PIP2 conditions: as described above, PLCs except for PLCζ target and hydrolyze PIP2 on the plasma membrane, whereas PLCζ hydrolysis PIP2 in the oocyte cytoplasm [86]. Furthermore, PIP2 fused to PLCζ-formed clusters in the oocyte cytoplasm, and Ca2+ oscillations decreased as the distance between PIP2 in the oocyte cytoplasm increased [127]. It was also found that the PLCζ protein did not show Ca2+ oscillations when injected into hamster ovary cells [128]. These results suggest a possibility that the Ca2+ oscillations that occur during fertilization may require currently uncharacterized factor(s) on the oocyte side and that “other factor(s)” is present besides PLCζ on the sperm side.
Based on the RNA sequence in germ cells, it may be informative to explore the possibility of PLCs other than PLCζ that are highly expressed in sperm and oocytes and that could be candidate factors inducing the rise in Ca2+. In mouse oocytes, PLCε1 is highly expressed compared to other PLCs, and therefore, we used mice lacking the Plce1 gene (Plce1−/−), which were previously produced by another group [129,130]. In our preliminary study, we observed that the female Plce1−/− mice did not show any impaired fertility. In addition, the oocytes retrieved from female Plce1−/− mice were successfully fertilized after IVF, and these embryos developed to the blastocyst stage, as did those from control mice. Although we did not measure the levels of intracellular Ca2+ in Plce1−/− oocytes during fertilization, even the Plcz1−/− sperm and Plce1−/− oocytes were also successfully fertilized. These findings suggest that (i) PLCε1 in the oocytes is not involved in oocyte activation, and (ii) PLCε1 can be ruled out as the candidate factor.
PLCδ4 is one of the most abundantly expressed PLCs in spermatocytes [131], and PLCδ4-deficient (Plcd4−/−) male mice produced a low number of small litters or were sterile [131]. With the use of IVF, they also showed that insemination with Plcd4−/− sperm resulted in significantly fewer oocytes becoming activated and that the Ca2+ transients associated with fertilization were absent or delayed [131]. There is no doubt that PLCζ induces Ca2+ oscillations during mammalian fertilization [32,33], but nonetheless, PLCζ and PLCδ4 may cooperate to increase the fusion of membrane-dependent rises in intracellular Ca2+ and then initiate Ca2+ oscillations in oocytes during fertilization. Results from PLCζ and PLCδ4 double gene-deficient (Plcz1−/−Plcd4−/−) mice may provide answers to the questions described above (Figure 7).
3.2. Another Divalent Ion for Oocyte Activation
The fertilization process is widely acknowledged to be dependent on the increase in intracellular Ca2+ ions. However, recent investigations have illuminated alternative pathways for inducing meiotic resumption from the MII arrest. Notably, these pathways include the utilization of a Zn2+ chelator, TPEN, and the suppression of meiotic resumption through Zn2+ ionophore overload [132,133,134]. In mouse models, TPEN treatment alone was observed to effectively activate MII-arrested oocytes that had been injected with “inactivated” sperm heads, resulting in successful live births post-embryo transfer [133]. Intriguingly, this activation occurred in the absence of intracellular Ca2+ oscillations, challenging the established notion that full-term development is reliant on the release of Ca2+ during MII exit, as suggested by Suzuki et al. (2010) [133]. These findings collectively imply that while the elevation of intracellular Ca2+ ions in the oocyte is traditionally considered crucial for oocyte activation, the depletion of Zn2+ ions within the oocyte can also act as a trigger for activation.
When mature mouse oocytes, which are abundant in Zn2+, undergo fertilization, there is a transient release of Zn2+ into the extracellular milieu. This coordinated series of events has been termed the “Zn2+ spark” [135,136]. The occurrence of the Zn2+ spark was reported to coincide closely with the initial elevation of Ca2+ levels. Following its discovery in mice, the phenomenon of the Zn2+ spark has been observed in human and bovine oocytes subsequent to fertilization and oocyte activation in mice [135], humans [137,138], and cattle [139], suggesting a high degree of conservation across several mammalian species. Recently, the presence of the Zn2+ spark during fertilization was also confirmed in amphibian oocytes [140]. Despite the widespread occurrence of Zn2+ sparks, their exact role in fertilization remains incompletely understood. Bernhardt et al. proposed a comprehensive model suggesting that Zn2+ plays a crucial role in modulating the concentration-dependent regulation of meiosis through its interaction with Emi2, a Zn2+-binding protein and a key component of the cytostatic factor (CSF). According to their model, Zn2+ sparks facilitate the rapid and efficient inactivation of Emi2 [132]. It is noteworthy that the inactivation of Emi2 is known to occur through a Ca2+-dependent mechanism [141], and Zn2+ sparks are observed to be absent in the absence of Ca2+ chelation [137]. Furthermore, subsequent studies have provided additional insights into the effects of elevated Zn2+ levels on sperm function. One such study demonstrated that increased Zn2+ concentrations hinder the forward motility of sperm, thereby impeding their ability to traverse through the zona matrix and ultimately leading to the prevention of polyspermy [142]. These findings underscore the intricate interplay between Zn2+ signaling, Ca2+ dynamics, and sperm function during fertilization, highlighting the need for further investigation into the molecular mechanisms underlying these processes.
While further investigations are essential to comprehensively elucidate the intricacies of this mechanism, the significance of the Zn2+ spark during fertilization is widely acknowledged within the scientific community. Prior to the occurrence of a Zn2+ spark, there must be a substantial buildup of Zn2+ ions within the oocyte. Studies have consistently demonstrated that immature mouse oocytes exhibit an incapacity to generate a Zn2+ spark, thereby emphasizing the pivotal role of acute Zn2+ accumulation during meiotic maturation [137,143]. Moreover, the presence of Zn2+ accumulation has been well documented in MII oocytes [144]. The dynamic regulation of Zn2+ influx and efflux is governed by various Zn2+ transporters, such as ZIP1-14 and ZnT1-9. Notably, in mouse oocytes, the expression of ZIP6 and ZIP10 has been highlighted as significant [145]. Hence, it is hypothesized that Zn2+ influx in oocytes is intricately modulated by these ZIP proteins. By providing this detailed explanation, we aim to offer a more comprehensive understanding of the intricate processes surrounding the phenomenon of the Zn2+ spark during fertilization, shedding light on its role in reproductive biology and potential implications for assisted reproductive technologies.
Zn2+ spark profiles suggest that zygotes progressing into blastocysts release higher levels of Zn2+ compared to those that fail to develop further. This phenomenon correlates with increased rates of embryo development and a greater total cell number at the blastocyst [146]. Therefore, the levels of Zn2+ ions detected at specific time points of Zn2+ sparks could potentially serve as an early biomarker for assessing the quality of embryos in mouse models. These findings indicate that both the elevation of intracellular Ca2+ ions and the release of Zn2+ ions from the oocyte are essential for oocyte activation during fertilization. Furthermore, it is suggested that one or more sperm factors must be capable of inducing both of these events.
3.3. Conclusions
As previously discussed, the fertilization process in mammals involves complicated mechanisms, not all of which have been fully elucidated. Our proposed hypothesis for oocyte activation is shown in Figure 7. While the importance of the elevation of intracellular Ca2+ during oocyte activation is undeniable, it is believed that there are two distinct mechanisms of Ca2+ elevation within the oocyte: one involving unknown Ca2+ elevation through the fusion of sperm and oocyte membranes and the other involving Ca2+ oscillations mediated by PLCζ. Furthermore, further research is needed to elucidate the role of Zn2+ ion-dependent mechanisms, such as Zn2+ spark, in oocyte activation. The clarification of these Ca2+ and Zn2+-dependent oocyte activation mechanisms is expected not only to contribute to knowledge in the field of reproductive biology but also to facilitate research on the treatment of human infertility and the efficient reproduction of livestock and endangered wild animals.
Figure 7.
Proposed hypothesis of the mechanism underlying Ca2+ oscillations during mammalian fertilization.
Figure 7.
Proposed hypothesis of the mechanism underlying Ca2+ oscillations during mammalian fertilization.

Author Contributions
Conceptualization, J.I.; investigation, H.S., S.T., Y.K. and A.K.; validation, A.K. and J.I.; data curation, A.K. and J.I.; writing—original draft preparation, H.S., S.T. and J.I.; writing—review and editing, H.S., S.T., A.K., Y.K. and J.I.; visualization; H.S., S.T. and J.I.; supervision; J.I. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by a Grant-in-Aid for Scientific Research from the Japan Society for the Promotion of Science (21H02384 and 20H05373 to J.I.). This study was also funded by the Center for Diversity, Equity & Inclusion, Azabu University (H.S.)
Acknowledgments
We thank the members of the Laboratory of Animal Reproduction for technical help. We also thank M. Ikawa (Osaka University) and T. Kataoka (Kobe University) for providing Plcz1−/− mice (RBRC10014) and Plce1−/− mice (RBRC02716), respectively. RBRC10014 and RBRC02716 were provided by the RIKEN BRC through the National Bio-Resource Project of the MEXT, Japan.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Zafar, M.I.; Lu, S.; Li, H. Sperm-Oocyte Interplay: An Overview of Spermatozoon’s Role in Oocyte Activation and Current Perspectives in Diagnosis and Fertility Treatment. Cell Biosci. 2021, 11, 4. [Google Scholar] [CrossRef]
- Satouh, Y.; Ikawa, M. New Insights into the Molecular Events of Mammalian Fertilization. Trends Biochem. Sci. 2018, 43, 818–828. [Google Scholar] [CrossRef]
- Evans, J.P. Preventing Polyspermy in Mammalian Eggs—Contributions of the Membrane Block and Other Mechanisms. Mol. Reprod. Dev. 2020, 87, 341–349. [Google Scholar] [CrossRef]
- Jones, K.T. Mammalian Egg Activation: From Ca2+ Spiking to Cell Cycle Progression. Reproduction 2005, 130, 813–823. [Google Scholar] [CrossRef]
- Madgwick, S.; Jones, K.T. How Eggs Arrest at Metaphase II: MPF Stabilisation plus APC/C Inhibition Equals Cytostatic Factor. Cell Div. 2007, 2, 4. [Google Scholar] [CrossRef]
- Tosti, E.; Ménézo, Y. Gamete Activation: Basic Knowledge and Clinical Applications. Hum. Reprod. Update 2016, 22, 420–439. [Google Scholar] [CrossRef] [PubMed]
- Schultz, R.M.; Kopf, G.S. Molecular Basis of Mammalian Egg Activation. Curr. Top. Dev. Biol. 1995, 30, 21–62. [Google Scholar] [CrossRef] [PubMed]
- Ducibella, T.; Fissore, R. The Roles of Ca2+, Downstream Protein Kinases, and Oscillatory Signaling in Regulating Fertilization and the Activation of Development. Dev. Biol. 2008, 315, 257–279. [Google Scholar] [CrossRef] [PubMed]
- Ito, J.; Parrington, J.; Fissore, R.A. PLCζ and Its Role as a Trigger of Development in Vertebrates. Mol. Reprod. Dev. 2011, 78, 846–853. [Google Scholar] [CrossRef] [PubMed]
- Jones, K.T. Intracellular Calcium in the Fertilization and Development of Mammalian Eggs. Clin. Exp. Pharmacol. Physiol. 2007, 34, 1084–1089. [Google Scholar] [CrossRef]
- Kang, I.; Koo, M.; Yoon, H.; Park, B.S.; Jun, J.H.; Lee, J. Ovastacin: An Oolemma Protein That Cleaves the Zona Pellucida to Prevent Polyspermy. Clin. Exp. Reprod. Med. 2023, 50, 154–159. [Google Scholar] [CrossRef]
- Liu, M. The Biology and Dynamics of Mammalian Cortical Granules. Reprod. Biol. Endocrinol. 2011, 9, 149. [Google Scholar] [CrossRef] [PubMed]
- Xia, P.; Wang, Z.; Yang, Z.; Tan, J.; Qin, P. Ultrastructural Study of Polyspermy during Early Embryo Development in Pigs, Observed by Scanning Electron Microscope and Transmission Electron Microscope. Cell Tissue Res. 2001, 303, 271–275. [Google Scholar] [CrossRef]
- Horii, T.; Yamamoto, M.; Morita, S.; Kimura, M.; Nagao, Y.; Hatada, I. P53 Suppresses Tetraploid Development in Mice. Sci. Rep. 2015, 5, 8907. [Google Scholar] [CrossRef]
- Zhou, C.; Homer, H.A. The Oocyte Spindle Midzone Pauses Cdk1 Inactivation during Fertilization to Enable Male Pronuclear Formation and Embryo Development. Cell. Rep. 2022, 39, 110789. [Google Scholar] [CrossRef] [PubMed]
- Backs, J.; Stein, P.; Backs, T.; Duncan, F.E.; Grueter, C.E.; McAnally, J.; Qi, X.; Schultz, R.M.; Olson, E.N. The γ Isoform of CaM Kinase II Controls Mouse Egg Activation by Regulating Cell Cycle Resumption. Proc. Natl. Acad. Sci. USA 2010, 107, 81–86. [Google Scholar] [CrossRef]
- Stricker, S.A. Comparative Biology of Calcium Signaling during Fertilization and Egg Activation in Animals. Dev. Biol. 1999, 211, 157–176. [Google Scholar] [CrossRef]
- Steinhardt, R.A.; Alderton, J.M. Calmodulin Confers Calcium Sensitivity on Secretory Exocytosis. Nature 1982, 295, 154–155. [Google Scholar] [CrossRef]
- Jaffe, L.F. Sources of Calcium in Egg Activation: A Review and Hypothesis. Dev. Biol. 1983, 99, 265–276. [Google Scholar] [CrossRef]
- Nuccitelli, R. How Do Sperm Activate Eggs? Curr. Top. Dev. Biol. 1991, 25, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Whitaker, M.; Swann, K. Lighting the Fuse at Fertilization. Development 1993, 117, 1–12. [Google Scholar] [CrossRef]
- Miyazaki, S.; Shirakawa, H.; Nakada, K.; Honda, Y. Essential Role of the Inositol 1,4,5-Trisphosphate Receptor/Ca2+ Release Channel in Ca2+ Waves and Ca2+ oscillations at Fertilization of Mammalian Eggs. Dev. Biol. 1993, 158, 62–78. [Google Scholar] [CrossRef] [PubMed]
- Swann, K.; Ozil, J.P. Dynamics of the Calcium Signal That Triggers Mammalian Egg Activation. Int. Rev. Cytol. 1994, 152, 183–222. [Google Scholar] [CrossRef] [PubMed]
- Jones, K.T. Ca2+ oscillations in the Activation of the Egg and Development of the Embryo in Mammals. Int. J. Dev. Biol. 1998, 42, 1–10. [Google Scholar] [PubMed]
- Kline, D.; Kline, J.T. Repetitive Calcium Transients and the Role of Calcium in Exocytosis and Cell Cycle Activation in the Mouse Egg. Dev. Biol. 1992, 149, 80–89. [Google Scholar] [CrossRef]
- Sanders, J.R.; Swann, K. Molecular Triggers of Egg Activation at Fertilization in Mammals. Reproduction 2016, 152, R41–R50. [Google Scholar] [CrossRef]
- Swann, K.; Lai, F.A. Egg Activation at Fertilization by a Soluble Sperm Protein. Physiol. Rev. 2016, 96, 127–149. [Google Scholar] [CrossRef]
- Raffaello, A.; Mammucari, C.; Gherardi, G.; Rizzuto, R. Calcium at the Center of Cell Signaling: Interplay between Endoplasmic Reticulum, Mitochondria, and Lysosomes. Trends Biochem. Sci. 2016, 41, 1035–1049. [Google Scholar] [CrossRef]
- Jones, K.T.; Carroll, J.; Merriman, J.A.; Whittingham, D.G.; Kono, T. Repetitive Sperm-Induced Ca2+ Transients in Mouse Oocytes Are Cell Cycle Dependent. Development 1995, 121, 3259–3266. [Google Scholar] [CrossRef]
- Swann, K.; Saunders, C.M.; Rogers, N.T.; Lai, F.A. PLCζ(Zeta): A Sperm Protein That Triggers Ca2+ oscillations and Egg Activation in Mammals. Semin. Cell Dev. Biol. 2006, 17, 264–273. [Google Scholar] [CrossRef]
- Lee, B.; Vermassen, E.; Yoon, S.Y.; Vanderheyden, V.; Ito, J.; Alfandari, D.; De Smedt, H.; Parys, J.B.; Fissore, R.A. Phosphorylation of IP3R1 and the Regulation of [Ca2+]i Responses at Fertilization: A Role for the MAP Kinase Pathway. Development 2006, 133, 4355–4365. [Google Scholar] [CrossRef]
- Hachem, A.; Godwin, J.; Ruas, M.; Lee, H.C.; Buitrago, M.F.; Ardestani, G.; Bassett, A.; Fox, S.; Navarrete, F.; De Sutter, P.; et al. Plcζ Is the Physiological Trigger of the Ca2+ oscillations That Induce Embryogenesis in Mammals but Conception Can Occur in Its Absence. Development 2017, 144, 2914–2924. [Google Scholar] [CrossRef]
- Nozawa, K.; Satouh, Y.; Fujimoto, T.; Oji, A.; Ikawa, M. Sperm-Borne Phospholipase C Zeta-1 Ensures Monospermic Fertilization in Mice. Sci. Rep. 2018, 8, 1315. [Google Scholar] [CrossRef]
- Ozil, J.P.; Banrezes, B.; Tóth, S.; Pan, H.; Schultz, R.M. Ca2+ Oscillatory Pattern in Fertilized Mouse Eggs Affects Gene Expression and Development to Term. Dev. Biol. 2006, 300, 534–544. [Google Scholar] [CrossRef]
- Nikiforaki, D.; Vanden Meerschaut, F.; De Roo, C.; Lu, Y.; Ferrer-Buitrago, M.; De Sutter, P.; Heindryckx, B. Effect of Two Assisted Oocyte Activation Protocols Used to Overcome Fertilization Failure on the Activation Potential and Calcium Releasing Pattern. Fertil. Steril. 2016, 105, 798–806.e2. [Google Scholar] [CrossRef]
- Rickords, L.F.; White, K.L. Electroporation of Inositol 1,4,5-triphosphate Induces Repetitive Calcium oscillations in Murine Oocytes. J. Exp. Zool. 1993, 265, 178–184. [Google Scholar] [CrossRef]
- Ducibella, T.; Huneau, D.; Angelichio, E.; Xu, Z.; Schultz, R.M.; Kopf, G.S.; Fissore, R.; Madoux, S.; Ozil, J.-P. Egg-to-Embryo Transition Is Driven by Differential Responses to Ca2+ oscillations Number. Dev. Biol. 2002, 250, 280–291. [Google Scholar] [CrossRef]
- Ozil, J.P.; Swann, K. Stimulation of Repetitive Calcium Transients in Mouse Eggs. J. Physiol. 1995, 483, 331–346. [Google Scholar] [CrossRef] [PubMed]
- Ramadan, W.M.; Kashir, J.; Jones, C.; Coward, K. Oocyte Activation and Phospholipase C Zeta (PLCζ): Diagnostic and Therapeutic Implications for Assisted Reproductive Technology. Cell Commun. Signal. 2012, 10, 12. [Google Scholar] [CrossRef] [PubMed]
- Swann, K.; Parrington, J. Mechanism of Ca2+ Release at Fertilization in Mammals. J. Exp. Zool. 1999, 285, 267–275. [Google Scholar] [CrossRef]
- Dale, B.; Santella, L. Sperm-Oocyte Interaction in the Sea-Urchin. J. Cell Sci. 1985, 74, 153–167. [Google Scholar] [CrossRef]
- Swann, K. A Cytosolic Sperm Factor Stimulates Repetitive Calcium Increases and Mimics Fertilization in Hamster Eggs. Development 1990, 110, 1295–1302. [Google Scholar] [CrossRef] [PubMed]
- Fissore, R.A.; Gordo, A.C.; Wu, H. Activation of Development in Mammals: Is There a Role for a Sperm Cytosolic Factor? Theriogenology 1998, 49, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Homa, S.T.; Swann, K. Fertilization and Early Embryology: A Cytosolic Sperm Factor Triggers Calcium oscillations and Membrane Hyperpolarizations in Human Oocytes. Hum. Reprod. 1994, 9, 2356–2361. [Google Scholar] [CrossRef] [PubMed]
- Dale, B.; DeFelice, L.J.; Ehrenstein, G. Injection of a Soluble Sperm Fraction into Sea-Urchin Eggs Triggers the Cortical Reaction. Experientia 1985, 41, 1068–1070. [Google Scholar] [CrossRef]
- Stice, S.L.; Robl, J.M. Activation of Mammalian Oocytes by a Factor Obtained from Rabbit Sperm. Mol. Reprod. Dev. 1990, 25, 272–280. [Google Scholar] [CrossRef] [PubMed]
- Parrington, J.; Anthony Lai, F.; Swann, K. The Soluble Mammalian Sperm Factor Protein That Triggers Ca2+ oscillations in Eggs: Evidence for Expression of MRNA(s) Coding for Sperm Factor Protein(s) in Spermatogenic Cells. Biol. Cell 2000, 92, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Sette, C.; Bevilacqua, A.; Bianchini, A.; Mangia, F.; Geremia, R.; Rossi, P. Parthenogenetic Activation of Mouse Eggs by Microinjection of a Truncated C-Kit Tyrosine Kinase Present in Spermatozoa. Development 1997, 124, 2267–2274. [Google Scholar] [CrossRef] [PubMed]
- Rice, A.; Parrington, J.; Jones, K.T.; Swann, K. Mammalian Sperm Contain a Ca2+-Sensitive Phospholipase C Activity That Can Generate InsP3 from PIP2 Associated with Intracellular Organelles. Dev. Biol. 2000, 228, 125–135. [Google Scholar] [CrossRef]
- Wu, H.; Smyth, J.; Luzzi, V.; Fukami, K.; Takenawa, T.; Black, S.L.; Allbritton, N.L.; Fissore, R.A. Sperm Factor Induces Intracellular Free Calcium oscillations by Stimulating the Phosphoinositide Pathway. Biol. Reprod. 2001, 64, 1338–1349. [Google Scholar] [CrossRef]
- Saunders, C.M.; Larman, M.G.; Parrington, J.; Cox, L.J.; Royse, J.; Blayney, L.M.; Swann, K.; Lai, F.A. PLCζ: A Sperm-Specific Trigger of Ca2+ oscillations in Eggs and Embryo Development. Development 2002, 129, 3533–3544. [Google Scholar] [CrossRef] [PubMed]
- Ito, M.; Shikano, T.; Kuroda, K.; Miyazaki, S. Relationship between Nuclear Sequestration of PLCζ and Termination of PLCζ-Induced Ca2+ oscillations in Mouse Eggs. Cell Calcium 2008, 44, 400–410. [Google Scholar] [CrossRef] [PubMed]
- Ito, M.; Shikano, T.; Oda, S.; Horiguchi, T.; Tanimoto, S.; Awaji, T.; Mitani, H.; Miyazaki, S. Difference in Ca2+ oscillations-Inducing Activity and Nuclear Translocation Ability of PLCZ1, an Egg-Activating Sperm Factor Candidate, between Mouse, Rat, Human, and Medaka Fish. Biol. Reprod. 2008, 78, 1081–1090. [Google Scholar] [CrossRef] [PubMed]
- Cox, L. Sperm Phospholipase Czeta from Humans and Cynomolgus Monkeys Triggers Ca2+ oscillations, Activation and Development of Mouse Oocytes. Reproduction 2002, 124, 611–623. [Google Scholar] [CrossRef] [PubMed]
- Cooney, M.A.; Malcuit, C.; Cheon, B.; Holland, M.K.; Fissore, R.A.; D’Cruz, N.T. Species-Specific Differences in the Activity and Nuclear Localization of Murine and Bovine Phospholipase C Zeta 1. Biol. Reprod. 2010, 83, 92–101. [Google Scholar] [CrossRef] [PubMed]
- Yoneda, A.; Kashima, M.; Yoshida, S.; Terada, K.; Nakagawa, S.; Sakamoto, A.; Hayakawa, K.; Suzuki, K.; Ueda, J.; Watanabe, T. Molecular Cloning, Testicular Postnatal Expression, and Oocyte-Activating Potential of Porcine Phospholipase Cζ. Reproduction 2006, 132, 393–401. [Google Scholar] [CrossRef] [PubMed]
- Nakai, M.; Suzuki, S.I.; Ito, J.; Fuchimoto, D.I.; Sembon, S.; Noguchi, J.; Onishi, A.; Kashiwazaki, N.; Kikuchi, K. Efficient Pig ICSI Using Percoll-Selected Spermatozoa; Evidence for the Essential Role of Phospholipase C-ζ in ICSI Success. J. Reprod. Dev. 2016, 62, 639–643. [Google Scholar] [CrossRef]
- Bedford-Guaus, S.J.; McPartlin, L.A.; Xie, J.; Westmiller, S.L.; Buffone, M.G.; Roberson, M.S. Molecular Cloning and Characterization of Phospholipase C Zeta in Equine Sperm and Testis Reveals Species-Specific Differences in Expression of Catalytically Active Protein. Biol. Reprod. 2011, 85, 78–88. [Google Scholar] [CrossRef]
- Sato, K.; Wakai, T.; Seita, Y.; Takizawa, A.; Fissore, R.A.; Ito, J.; Kashiwazaki, N. Molecular Characteristics of Horse Phospholipase C Zeta (PLCζ). Anim. Sci. J. 2013, 84, 359–368. [Google Scholar] [CrossRef]
- Mizushima, S.; Takagi, S.; Ono, T.; Atsumi, Y.; Tsukada, A.; Saito, N.; Shimada, K. Phospholipase Cζ MRNA Expression and Its Potency during Spermatogenesis for Activation of Quail Oocyte as a Sperm Factor. Mol. Reprod. Dev. 2009, 76, 1200–1207. [Google Scholar] [CrossRef]
- Rebecchi, M.J.; Pentyala, S.N. Structure, Function, and Control of Phosphoinositide-Specific Phospholipase C. Physiol. Rev. 2000, 80, 1291–1335. [Google Scholar] [CrossRef]
- Ito, J.; Yoon, S.Y.; Lee, B.; Vanderheyden, V.; Vermassen, E.; Wojcikiewicz, R.; Alfandari, D.; De Smedt, H.; Parys, J.B.; Fissore, R.A. Inositol 1,4,5-Trisphosphate Receptor 1, a Widespread Ca2+ Channel, Is a Novel Substrate of Polo-like Kinase 1 in Eggs. Dev. Biol. 2008, 320, 402–413. [Google Scholar] [CrossRef]
- Miyazaki, S.; Yuzaki, M.; Nakada, K.; Shirakawa, H.; Nakanishi, S.; Nakade, S.; Mikoshiba, K. Block of Ca2+ Wave and Ca2+ oscillations by Antibody to the Inositol 1,4,5-Trisphosphate Receptor in Fertilized Hamster Eggs. Science 1992, 257, 251–255. [Google Scholar] [CrossRef]
- Knott, J.G.; Kurokawa, M.; Fissore, R.A.; Schultz, R.M.; Williams, C.J. Transgenic RNA Interference Reveals Role for Mouse Sperm Phospholipase Cζ in Triggering Ca2+ oscillations during Fertilization. Biol. Reprod. 2005, 72, 992–996. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.Y.; Fissore, R.A. Release of Phospholipase C ζ and [Ca2+]i oscillations-Inducing Activity during Mammalian Fertilization. Reproduction 2007, 134, 695–704. [Google Scholar] [CrossRef] [PubMed]
- Nakai, M.; Ito, J.; Sato, K.I.; Noguchi, J.; Kaneko, H.; Kashiwazaki, N.; Kikuchi, K. Pre-Treatment of Sperm Reduces Success of ICSI in the Pig. Reproduction 2011, 142, 285–293. [Google Scholar] [CrossRef] [PubMed]
- Aldana, A.; Carneiro, J.; Martínez-Mekler, G.; Darszon, A. Discrete Dynamic Model of the Mammalian Sperm Acrosome Reaction: The Influence of Acrosomal PH and Physiological Heterogeneity. Front. Physiol. 2021, 12, 682790. [Google Scholar] [CrossRef] [PubMed]
- Grasa, P.; Coward, K.; Young, C.; Parrington, J. The Pattern of Localization of the Putative Oocyte Activation Factor, Phospholipase Cζ, in Uncapacitated, Capacitated, and Ionophore-Treated Human Spermatozoa. Hum. Reprod. 2008, 23, 2513–2522. [Google Scholar] [CrossRef] [PubMed]
- Williams, R.L. Mammalian Phosphoinositide-Specific Phospholipase C. Biochim. Biophys. Acta—Mol. Cell Biol. Lipids 1999, 1441, 255–267. [Google Scholar] [CrossRef] [PubMed]
- Fukami, K.; Inanobe, S.; Kanemaru, K.; Nakamura, Y. Phospholipase C Is a Key Enzyme Regulating Intracellular Calcium and Modulating the Phosphoinositide Balance. Prog. Lipid Res. 2010, 49, 429–437. [Google Scholar] [CrossRef] [PubMed]
- Ni, T.; Kalli, A.C.; Naughton, F.B.; Yates, L.A.; Naneh, O.; Kozorog, M.; Anderluh, G.; Sansom, M.S.P.; Gilbert, R.J.C. Structure and Lipid-Binding Properties of the Kindlin-3 Pleckstrin Homology Domain. Biochem. J. 2017, 474, 539–556. [Google Scholar] [CrossRef]
- Essen, L.O.; Perisic, O.; Cheung, R.; Katan, M.; Williams, R.L. Crystal Structure of a Mammalian Phosphoinositide-Specific Phospholipase Cδ. Nature 1996, 380, 595–602. [Google Scholar] [CrossRef]
- Fujii, M.; Ohtsubo, M.; Ogawa, T.; Kamata, H.; Hirata, H.; Yagisawa, H. Real-Time Visualization of PH Domain-Dependent Translocation of Phospholipase C-δ1 in Renal Epithelial Cells (MDCK): Response to Hypo-Osmotic Stress. Biochem. Biophys. Res. Commun. 1999, 254, 284–291. [Google Scholar] [CrossRef]
- George, S.E.; Su, Z.; Fan, D.; Wang, S.; David Johnson, J. The Fourth EF-Hand of Calmodulin and Its Helix-Loop-Helix Components: Impact on Calcium Binding and Enzyme Activation. Biochemistry 1996, 35, 8307–8313. [Google Scholar] [CrossRef]
- Rhee, S.G.; Choi, K.D. Regulation of Inositol Phospholipid-Specific Phospholipase C Isozymes. J. Biol. Chem. 1992, 267, 12393–12396. [Google Scholar] [CrossRef] [PubMed]
- Otterhag, L.; Sommarin, M.; Pical, C. N-Terminal EF-Hand-like Domain Is Required for Phosphoinositide-Specific Phospholipase C Activity in Arabidopsis Thaliana. FEBS Lett. 2001, 497, 165–170. [Google Scholar] [CrossRef] [PubMed]
- Bunney, T.D.; Katan, M. PLC Regulation: Emerging Pictures for Molecular Mechanisms. Trends Biochem. Sci. 2011, 36, 88–96. [Google Scholar] [CrossRef]
- Ellis, M.V.; Katan, S.U.; Katan, M. Mutations within a Highly Conserved Sequence Present in the X Region of Phosphoinositide-Specific Phospholipase C-δ1. Biochem. J. 1995, 307, 69–75. [Google Scholar] [CrossRef]
- Nagano, N.; Orengo, C.A.; Thornton, J.M. One Fold with Many Functions: The Evolutionary Relationships between TIM Barrel Families Based on Their Sequences, Structures and Functions. J. Mol. Biol. 2002, 321, 741–765. [Google Scholar] [CrossRef]
- Hicks, S.N.; Jezyk, M.R.; Gershburg, S.; Seifert, J.P.; Harden, T.K.; Sondek, J. General and Versatile Autoinhibition of PLC Isozymes. Mol. Cell 2008, 31, 383–394. [Google Scholar] [CrossRef]
- Van Huizen, R.; Miller, K.; Chen, D.M.; Li, Y.; Lai, Z.C.; Raab, R.W.; Stark, W.S.; Shortridge, R.D.; Li, M. Two Distantly Positioned PDZ Domains Mediate Multivalent INAD-Phospholipase C Interactions Essential for G Protein-Coupled Signaling. EMBO J. 1998, 17, 2285–2297. [Google Scholar] [CrossRef]
- Essen, L.O.; Perisic, O.; Lynch, D.E.; Katan, M.; Williams, R.L. A Ternary Metal Binding Site in the C2 Domain of Phosphoinositide- Specific Phospholipase C-δ1. Biochemistry 1997, 36, 2753–2762. [Google Scholar] [CrossRef]
- Kawasaki, H.; Nakayama, S.; Kretsinger, R.H. Classification and Evolution of EF-Hand Proteins. BioMetals 1998, 11, 277–295. [Google Scholar] [CrossRef]
- Kim, Y.; Welch, J.T.; Lindstrom, K.M.; Franklin, S.J. Chimeric HTH Motifs Based on EF-Hands. J. Biol. Inorg. Chem. 2001, 6, 173–181. [Google Scholar] [CrossRef]
- Bill, C.A.; Vines, C.M. Phospholipase C. Adv. Exp. Med. Biol. 2020, 1131, 215–242. [Google Scholar] [CrossRef]
- Yu, A.; Nomikos, M.; Theodoridou, M.; Nounesis, G.; Lai, F.A.; Swann, K. PLCζ Causes Ca2+ oscillations in Mouse Eggs by Targeting Intracellular and Not Plasma Membrane PI(4,5)P2. Mol. Biol. Cell 2012, 23, 371–380. [Google Scholar] [CrossRef]
- Nomikos, M.; Sanders, J.R.; Parthimos, D.; Buntwal, L.; Calver, B.L.; Stamatiadis, P.; Smith, A.; Clue, M.; Sideratou, Z.; Swann, K.; et al. Essential Role of the EF-Hand Domain in Targeting Sperm Phospholipase Cζ to Membrane Phosphatidylinositol 4,5-Bisphosphate (PIP2). J. Biol. Chem. 2005, 290, 29519–29530. [Google Scholar] [CrossRef]
- Kouchi, Z.; Fukami, K.; Shikano, T.; Oda, S.; Nakamura, Y.; Takenawa, T.; Miyazaki, S. Recombinant Phospholipase Cζ Has High Ca2+ Sensitivity and Induces Ca2+ oscillations in Mouse Eggs. J. Biol. Chem. 2004, 279, 10408–10412. [Google Scholar] [CrossRef] [PubMed]
- Nomikos, M.; Blayney, L.M.; Larman, M.G.; Campbell, K.; Rossbach, A.; Saunders, C.M.; Swann, K.; Lai, F.A. Role of Phospholipase C-ζ Domains in Ca2+-Dependent Phosphatidylinositol 4,5-Bisphosphate Hydrolysis and Cytoplasmic Ca2+ oscillations. J. Biol. Chem. 2005, 280, 31011–31018. [Google Scholar] [CrossRef]
- Kouchi, Z.; Shikano, T.; Nakamura, Y.; Shirakawa, H.; Fukami, K.; Miyazaki, S. The Role of EF-Hand Domains and C2 Domain in Regulation of Enzymatic Activity of Phospholipase Cζ. J. Biol. Chem. 2005, 280, 21015–21021. [Google Scholar] [CrossRef]
- Nakanishi, T.; Ishibashi, N.; Kubota, H.; Inoue, K.; Ogonuki, N.; Ogura, A.; Kashiwabara, S.I.; Baba, T. Birth of Normal Offspring from Mouse Eggs Activated by a Phospholipase Cζ Protein Lacking Three EF-Hand Domains. J. Reprod. Dev. 2008, 54, 244–249. [Google Scholar] [CrossRef] [PubMed]
- Ellis, M.V.; Carne, A.; Katan, M. Structural Requirements of Phosphatidylinositol-specific Phospholipase C δ1 for Enzyme Activity. Eur. J. Biochem. 1993, 213, 339–347. [Google Scholar] [CrossRef] [PubMed]
- Katan, M. Families of Phosphoinositide-Specific Phospholipase C: Structure and Function. Biochim. Biophys. Acta—Mol. Cell Biol. Lipids 1998, 1436, 5–17. [Google Scholar] [CrossRef] [PubMed]
- Ellis, M.V.; James, S.R.; Perisic, O.; Downes, C.P.; Williams, R.L.; Katan, M. Catalytic Domain of Phosphoinositide-Specific Phospholipase C (PLC). J. Biol. Chem. 1998, 273, 11650–11659. [Google Scholar] [CrossRef] [PubMed]
- Kuroda, K.; Ito, M.; Shikano, T.; Awaji, T.; Yoda, A.; Takeuchi, H.; Kinoshita, K.; Miyazaki, S. The Role of X/Y Linker Region and N-Terminal EF-Hand Domain in Nuclear Translocation and Ca2+ oscillations-Inducing Activities of Phospholipase Cζ, a Mammalian Egg-Activating Factor. J. Biol. Chem. 2006, 281, 27794–27805. [Google Scholar] [CrossRef]
- Dingwall, C.; Laskey, R.A. Nuclear Targeting Sequences—A Consensus? Trends Biochem. Sci. 1991, 16, 478–481. [Google Scholar] [CrossRef]
- Okada, M.; Fujii, M.; Yamaga, M.; Sugimoto, H.; Sadano, H.; Osumi, T.; Kamata, H.; Hirata, H.; Yagisawa, H. Carboxyl-Terminal Basic Amino Acids in the X Domain Are Essential for the Nuclear Import of Phospholipase C δ1. Genes Cells 2002, 7, 985–996. [Google Scholar] [CrossRef]
- Yoda, A.; Oda, S.; Shikano, T.; Kouchi, Z.; Awaji, T.; Shirakawa, H.; Kinoshita, K.; Miyazaki, S. Ca2+ oscillations-Inducing Phospholipase C Zeta Expressed in Mouse Eggs Is Accumulated to the Pronucleus during Egg Activation. Dev. Biol. 2004, 268, 245–257. [Google Scholar] [CrossRef]
- Nomikos, M.; Elgmati, K.; Theodoridou, M.; Calver, B.L.; Nounesis, G.; Swann, K.; Lai, F.A. Phospholipase Cζ Binding to PtdIns(4,5)P2 Requires the XY-Linker Region. J. Cell Sci. 2011, 124, 2582–2590. [Google Scholar] [CrossRef]
- Kurokawa, M.; Yoon, S.Y.; Alfandari, D.; Fukami, K.; Sato, K.-i.; Fissore, R.A. Proteolytic Processing of Phospholipase Cζ and [Ca2+]i oscillations during Mammalian Fertilization. Dev. Biol. 2007, 312, 407–418. [Google Scholar] [CrossRef]
- Rizo, J.; Sudhof, T.C. C2-Domains, Structure and Function of a Universal Ca2+-Binding Domain. J. Biol. Chem. 1998, 273, 15879–15882. [Google Scholar] [CrossRef]
- Theodoridou, M.; Nomikos, M.; Parthimos, D.; Gonzalez-Garcia, J.R.; Elgmati, K.; Calver, B.L.; Sideratou, Z.; Nounesis, G.; Swann, K.; Lai, F.A. Chimeras of Sperm PLCζ Reveal Disparate Protein Domain Functions in the Generation of Intracellular Ca2+ oscillations in Mammalian Eggs at Fertilization. Mol. Hum. Reprod. 2013, 19, 852–864. [Google Scholar] [CrossRef]
- Yoshida, N.; Amanai, M.; Fukui, T.; Kajikawa, E.; Brahmajosyula, M.; Iwahori, A.; Nakano, Y.; Shoji, S.; Diebold, J.; Hessel, H.; et al. Broad, Ectopic Expression of the Sperm Protein PLCZ1 Induces Parthenogenesis and Ovarian Tumours in Mice. Development 2007, 134, 3941–3952. [Google Scholar] [CrossRef]
- Bianchi, E.; Doe, B.; Goulding, D.; Wright, G.J. Juno Is the Egg Izumo Receptor and Is Essential for Mammalian Fertilization. Nature 2014, 508, 483–487. [Google Scholar] [CrossRef]
- Meng, X.; Melo, P.; Jones, C.; Ross, C.; Mounce, G.; Turner, K.; Child, T.; Coward, K. Use of Phospholipase C Zeta Analysis to Identify Candidates for Artificial Oocyte Activation: A Case Series of Clinical Pregnancies and a Proposed Algorithm for Patient Management. Fertil. Steril. 2020, 114, 163–174. [Google Scholar] [CrossRef] [PubMed]
- Allahveisi, A.; Yousefian, E. Assessment of Expression Levels and Localization Patterns of Phospholipase C Zeta in Different Grades of HOST in Human Sperm. Int. J. Fertil. Steril. 2023, 18, 26. [Google Scholar] [CrossRef]
- Jones, C.; Meng, X.; Coward, K. SPERM FACTORS AND EGG ACTIVATION Phospholipase C Zeta (PLCZ1) and the Clinical Diagnosis of Oocyte Activation Deficiency. Reproduction 2022, 164, F53–F66. [Google Scholar] [CrossRef]
- Heytens, E.; Parrington, J.; Coward, K.; Young, C.; Lambrecht, S.; Yoon, S.Y.; Fissore, R.A.; Hamer, R.; Deane, C.M.; Ruas, M.; et al. Reduced Amounts and Abnormal Forms of Phospholipase C Zeta (PLCζ) in Spermatozoa from Infertile Men. Hum. Reprod. 2009, 24, 2417–2428. [Google Scholar] [CrossRef]
- Kashir, J.; Konstantinidis, M.; Jones, C.; Lemmon, B.; Chang Lee, H.; Hamer, R.; Heindryckx, B.; Deane, C.M.; De Sutter, P.; Fissore, R.A.; et al. Maternally Inherited Autosomal Point Mutation in Human Phospholipase C Zeta (PLCζ) Leads to Male Infertility. Hum. Reprod. 2012, 27, 222–231. [Google Scholar] [CrossRef]
- Kashir, J.; Jones, C.; Lee, H.C.; Rietdorf, K.; Nikiforaki, D.; Durrans, C.; Ruas, M.; Tee, S.T.; Heindryckx, B.; Galione, A.; et al. Loss of Activity Mutations in Phospholipase C Zeta (PLCζ) Abolishes Calcium Oscillatory Ability of Human Recombinant Protein in Mouse Oocytes. Hum. Reprod. 2011, 26, 3372–3387. [Google Scholar] [CrossRef]
- Escoffier, J.; Lee, H.C.; Yassine, S.; Zouari, R.; Martinez, G.; Karaouzène, T.; Coutton, C.; Kherraf, Z.E.; Halouani, L.; Triki, C.; et al. Homozygous Mutation of PLCZ1 Leads to Defective Human Oocyte Activation and Infertility That Is Not Rescued by the WW-Binding Protein PAWP. Hum. Mol. Genet. 2016, 25, 878–891. [Google Scholar] [CrossRef]
- Yuan, P.; Zheng, L.; Liang, H.; Lin, Q.; Ou, S.; Zhu, Y.; Lai, L.; Zhang, Q.; He, Z.; Wang, W. Novel Mutations in the PLCZ1 Gene Associated with Human Low or Failed Fertilization. Mol. Genet. Genomic Med. 2020, 8, e1470. [Google Scholar] [CrossRef]
- Ruan, J.L.; Liang, S.S.; Pan, J.P.; Chen, Z.Q.; Teng, X.M. Artificial Oocyte Activation with Ca2+ Ionophore Improves Reproductive Outcomes in Patients with Fertilization Failure and Poor Embryo Development in Previous ICSI Cycles. Front. Endocrinol. 2023, 14, 1244507. [Google Scholar] [CrossRef]
- Fawzy, M.; Emad, M.; Mahran, A.; Sabry, M.; Fetih, A.N.; Abdelghafar, H.; Rasheed, S. Artificial Oocyte Activation with SrCl2 or Calcimycin after ICSI Improves Clinical and Embryological Outcomes Compared with ICSI Alone: Results of a Randomized Clinical Trial. Hum. Reprod. 2018, 33, 1636–1644. [Google Scholar] [CrossRef]
- Torra-Massana, M.; Cornet-Bartolomé, D.; Barragán, M.; Durban, M.; Ferrer-Vaquer, A.; Zambelli, F.; Rodriguez, A.; Oliva, R.; Vassena, R. Novel Phospholipase C Zeta 1 Mutations Associated with Fertilization Failures after ICSI. Hum. Reprod. 2019, 34, 1494–1504. [Google Scholar] [CrossRef]
- Campos, G.; Sciorio, R.; Esteves, S.C. Total Fertilization Failure after ICSI: Insights into Pathophysiology, Diagnosis, and Management through Artificial Oocyte Activation. Hum. Reprod. Update 2023, 29, 369–394. [Google Scholar] [CrossRef]
- Wei, Y.; Wang, J.; Qu, R.; Zhang, W.; Tan, Y.; Sha, Y.; Li, L.; Yin, T. Genetic mechanisms of fertilization failure and early embryonic arrest: A comprehensive review. Hum. Reprod. Update 2024, 30, 48–80. [Google Scholar] [CrossRef]
- Dai, J.; Dai, C.; Guo, J.; Zheng, W.; Zhang, T.; Li, Y.; Lu, C.; Gong, F.; Lu, G.; Lin, G. Novel Homozygous Variations in PLCZ1. Lead to Poor or Failed Fertilization Characterized by Abnormal Localization Patterns of PLCζ in Sperm. Clin. Genet. 2020, 97, 347–351. [Google Scholar] [CrossRef]
- Mu, J.; Zhang, Z.; Wu, L.; Fu, J.; Chen, B.; Yan, Z.; Li, B.; Zhou, Z.; Wang, W.; Zhao, L.; et al. The Identification of Novel Mutations in PLCZ1 Responsible for Human Fertilization Failure and a Therapeutic Intervention by Artificial Oocyte Activation. Mol. Hum. Reprod. 2020, 26, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Yuan, P.; Yang, C.; Ren, Y.; Yan, J.; Nie, Y.; Yan, L.; Qiao, J. A Novel Homozygous Mutation of Phospholipase C Zeta Leading. to Defective Human Oocyte Activation and Fertilization Failure. Hum. Reprod. 2021, 35, 977–985. [Google Scholar] [CrossRef]
- Wu, A.T.H.; Sutovsky, P.; Manandhar, G.; Xu, W.; Katayama, M.; Day, B.N.; Park, K.W.; Yi, Y.J.; Yan, W.X.; Prather, R.S.; et al. PAWP, a Sperm-Specific WW Domain-Binding Protein, Promotes Meiotic Resumption and Pronuclear Development during Fertilization. J. Biol. Chem. 2007, 282, 12164–12175. [Google Scholar] [CrossRef]
- Aarabi, M.; Qin, Z.; Xu, W.; Mewburn, J.; Oko, R. Sperm-Borne Protein, PAWP, Initiates Zygotic Development in Xenopus laevis by Eliciting Intracellular Calcium Release. Mol. Reprod. Dev. 2010, 77, 249–256. [Google Scholar] [CrossRef]
- Aarabi, M.; Balakier, H.; Bashar, S.; Moskovtsev, S.I.; Sutovsky, P.; Librach, C.L.; Oko, R. Sperm-Derived WW Domain-Binding Protein, PAWP, Elicits Calcium oscillations and Oocyte Activation in Humans and Mice. FASEB J. 2014, 28, 4434–4440. [Google Scholar] [CrossRef]
- Harada, Y.; Matsumoto, T.; Hirahara, S.; Nakashima, A.; Ueno, S.; Oda, S.; Miyazaki, S.; Iwao, Y. Characterization of a Sperm Factor for Egg Activation at Fertilization of the Newt Cynops Pyrrhogaster. Dev. Biol. 2007, 306, 797–808. [Google Scholar] [CrossRef]
- Kang, W.; Harada, Y.; Yamatoya, K.; Kawano, N.; Kanai, S.; Miyamoto, Y.; Nakamura, A.; Miyado, M.; Hayashi, Y.; Kuroki, Y.; et al. Extra-Mitochondrial Citrate Synthase Initiates Calcium oscillations and Suppresses Age-Dependent Sperm Dysfunction. Lab. Investig. 2020, 100, 583–595. [Google Scholar] [CrossRef]
- Satouh, Y.; Nozawa, K.; Ikawa, M. Sperm Postacrosomal WW Domain-Binding Protein Is Not Required for Mouse Egg Activation. Biol. Reprod. 2015, 93, 94. [Google Scholar] [CrossRef]
- Sanders, J.R.; Ashley, B.; Moon, A.; Woolley, T.E.; Swann, K. PLCζ Induced Ca2+ Oscillations in Mouse Eggs Involve a Positive Feedback Cycle of Ca2+ Induced InsP3 Formation from Cytoplasmic PIP2. Front. Cell Dev. Biol. 2018, 6, 36. [Google Scholar] [CrossRef]
- Phillips, S.V.; Yu, Y.; Rossbach, A.; Nomikos, M.; Vassilakopoulou, V.; Livaniou, E.; Cumbes, B.; Lai, F.A.; George, C.H.; Swann, K. Divergent Effect of Mammalian PLCζ in Generating Ca2+ Oscillations in Somatic Cells Compared with Eggs. Biochem. J. 2011, 438, 545–553. [Google Scholar] [CrossRef]
- Tadano, M.; Edamatsu, H.; Minamisawa, S.; Yokoyama, U.; Ishikawa, Y.; Suzuki, N.; Saito, H.; Wu, D.; Masago-Toda, M.; Yamawaki-Kataoka, Y.; et al. Congenital Semilunar Valvulogenesis Defect in Mice Deficient in Phospholipase Cε. Mol. Cell Biol. 2005, 25, 2191–2199. [Google Scholar] [CrossRef]
- Bai, Y.; Edamatsu, H.; Maeda, S.; Saito, H.; Suzuki, N.; Satoh, T.; Kataoka, T. Crucial Role of Phospholipase Cε in Chemical Carcinogen-Induced Skin Tumor Development. Cancer Res. 2004, 64, 8808–8810. [Google Scholar] [CrossRef]
- Fukami, K.; Nakao, K.; Inoue, T.; Kataoka, Y.; Kurokawa, M.; Fissore, R.A.; Nakamura, K.; Katsuki, M.; Mikoshiba, K.; Yoshida, N.; et al. Requirement of Phospholipase Cδ4 for the Zona Pellucida-Induced Acrosome Reaction. Science 2001, 292, 920–923. [Google Scholar] [CrossRef]
- Bernhardt, M.L.; Kong, B.Y.; Kim, A.M.; O’Halloran, T.V.; Woodruff, T.K. A Zinc-Dependent Mechanism Regulates Meiotic Progression in Mammalian Oocytes. Biol. Reprod. 2012, 86, 114. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Yoshida, N.; Suzuki, E.; Okuda, E.; Perry, A.C.F. Full-Term Mouse Development by Abolishing Zn2+-Dependent Metaphase II Arrest without Ca2+ Release. Development 2010, 137, 2659–2669. [Google Scholar] [CrossRef]
- Lee, K.; Davis, A.; Zhang, L.; Ryu, J.; Spate, L.D.; Park, K.W.; Samuel, M.S.; Walters, E.M.; Murphy, C.N.; Machaty, Z.; et al. Pig Oocyte Activation Using a Zn2+ Chelator, TPEN. Theriogenology 2015, 84, 1024–1032. [Google Scholar] [CrossRef] [PubMed]
- Kim, A.M.; Bernhardt, M.L.; Kong, B.Y.; Ahn, R.W.; Vogt, S.; Woodruff, T.K.; O’Halloran, T.V. Zinc Sparks Are Triggered by Fertilization and Facilitate Cell Cycle Resumption in Mammalian Eggs. ACS Chem. Biol. 2011, 6, 716–723. [Google Scholar] [CrossRef] [PubMed]
- Que, E.L.; Bleher, R.; Duncan, F.E.; Kong, B.Y.; Gleber, S.C.; Vogt, S.; Chen, S.; Garwin, S.A.; Bayer, A.R.; Dravid, V.P.; et al. Quantitative Mapping of Zinc Fluxes in the Mammalian Egg Reveals the Origin of Fertilization-Induced Zinc Sparks. Nat. Chem. 2015, 7, 130–139. [Google Scholar] [CrossRef]
- Duncan, F.E.; Que, E.L.; Zhang, N.; Feinberg, E.C.; O’Halloran, T.V.; Woodruff, T.K. The Zinc Spark Is an Inorganic Signature of Human Egg Activation. Sci. Rep. 2016, 6, 24737. [Google Scholar] [CrossRef]
- Lee, H.C.; Edmonds, M.E.; Duncan, F.E.; O’Halloran, T.V.; Woodruff, T.K. Zinc Exocytosis Is Sensitive to Myosin Light Chain Kinase Inhibition in Mouse and Human Eggs. Mol. Hum. Reprod. 2020, 26, 228–239. [Google Scholar] [CrossRef]
- Que, E.L.; Duncan, F.E.; Lee, H.C.; Hornick, J.E.; Vogt, S.; Fissore, R.A.; O’Halloran, T.V.; Woodruff, T.K. Bovine Eggs Release Zinc in Response to Parthenogenetic and Sperm-Induced Egg Activation. Theriogenology 2019, 127, 41–48. [Google Scholar] [CrossRef]
- Seeler, J.F.; Sharma, A.; Zaluzec, N.J.; Bleher, R.; Lai, B.; Schultz, E.G.; Hoffman, B.M.; LaBonne, C.; Woodruff, T.K.; O’Halloran, T.V. Metal Ion Fluxes Controlling Amphibian Fertilization. Nat. Chem. 2021, 13, 683–691. [Google Scholar] [CrossRef]
- Hansen, D.V.; Tung, J.J.; Jackson, P.K. CaMKII and Polo-like Kinase 1 Sequentially Phosphorylate the Cytostatic Factor Emi2/XErp1 to Trigger Its Destruction and Meiotic Exit. Proc. Natl. Acad. Sci. USA 2006, 103, 608–613. [Google Scholar] [CrossRef] [PubMed]
- Tokuhiro, K.; Dean, J. Glycan-Independent Gamete Recognition Triggers Egg Zinc Sparks and ZP2 Cleavage to Prevent Polyspermy. Dev. Cell 2018, 46, 627–640.e5. [Google Scholar] [CrossRef] [PubMed]
- Kim, A.M.; Vogt, S.; O’Halloran, T.V.; Woodruff, T.K. Zinc Availability Regulates Exit from Meiosis in Maturing Mammalian Oocytes. Nat. Chem. Biol. 2010, 6, 674–681. [Google Scholar] [CrossRef] [PubMed]
- Lisle, R.S.; Anthony, K.; Randall, M.A.; Diaz, F.J. Oocyte-Cumulus Cell Interactions Regulate Free Intracellular Zinc in Mouse Oocytes. Reproduction 2013, 145, 381–390. [Google Scholar] [CrossRef]
- Kong, B.Y.; Duncan, F.E.; Que, E.L.; Kim, A.M.; O’Halloran, T.V.; Woodruff, T.K. Maternally-Derived Zinc Transporters ZIP6 and ZIP10 Drive the Mammalian Oocyte-to-Egg Transition. Mol. Hum. Reprod. 2014, 20, 1077–1089. [Google Scholar] [CrossRef]
- Zhang, N.; Duncan, F.E.; Que, E.L.; O’Halloran, T.V.; Woodruff, T.K. The Fertilization-Induced Zinc Spark Is a Novel Biomarker of Mouse Embryo Quality and Early Development. Sci. Rep. 2016, 6, 22772. [Google Scholar] [CrossRef]
Figure 1.
Mechanism of rise(s) in intracellular Ca2+ level via PLCζ during mammalian fertilization. After the sperm–oocyte fusion, PLCζ is released from the sperm into the oocyte. PLCζ hydrolyzes phosphatidylinositol 4,5-bisphosphate (PIP2) to inositol trisphosphate (IP3) and diacylglycerol (DG). DG induces the exocytosis of cortical granules, results in the zona reaction, and IP3 binds to the IP3 receptor (IP3R), which leads to the release of Ca2+ from the endoplasmic reticulum (ER).
Figure 1.
Mechanism of rise(s) in intracellular Ca2+ level via PLCζ during mammalian fertilization. After the sperm–oocyte fusion, PLCζ is released from the sperm into the oocyte. PLCζ hydrolyzes phosphatidylinositol 4,5-bisphosphate (PIP2) to inositol trisphosphate (IP3) and diacylglycerol (DG). DG induces the exocytosis of cortical granules, results in the zona reaction, and IP3 binds to the IP3 receptor (IP3R), which leads to the release of Ca2+ from the endoplasmic reticulum (ER).

Figure 2.
Representative Ca2+ release at fertilization in oocytes of several species.

Figure 3.
Hypotheses regarding the sperm-inducing Ca2+ release during mammalian fertilization.

Figure 4.
Species-dependent localization of PLCζ in mammalian spermatozoa. Orange indicates the localization of PLCζ protein.
Figure 4.
Species-dependent localization of PLCζ in mammalian spermatozoa. Orange indicates the localization of PLCζ protein.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Sugita, H.; Takarabe, S.; Kageyama, A.; Kawata, Y.; Ito, J. Molecular Mechanism of Oocyte Activation in Mammals: Past, Present, and Future Directions. Biomolecules 2024, 14, 359. https://doi.org/10.3390/biom14030359
AMA Style
Sugita H, Takarabe S, Kageyama A, Kawata Y, Ito J. Molecular Mechanism of Oocyte Activation in Mammals: Past, Present, and Future Directions. Biomolecules. 2024; 14(3):359. https://doi.org/10.3390/biom14030359
Chicago/Turabian StyleSugita, Hibiki, Shunsuke Takarabe, Atsuko Kageyama, Yui Kawata, and Junya Ito. 2024. "Molecular Mechanism of Oocyte Activation in Mammals: Past, Present, and Future Directions" Biomolecules 14, no. 3: 359. https://doi.org/10.3390/biom14030359
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.