1. Introduction
Vasoactive intestinal peptide (VIP), a neuropeptide present exclusively in hippocampal interneurons [
1], is a modulator of hippocampal synaptic transmission and excitability [
2]. Its endogenous actions are believed to be most relevant in conditions eliciting high-frequency firing such as occurring during hippocampal synaptic plasticity and seizures [
2,
3,
4]. Its actions are mediated by hippocampal VPAC
1 and VPAC
2 receptors that bind with similar affinity to the neuropeptide pituitary adenylate cyclase-activating polypeptide (PACAP). These integrate the VIP/PACAP family of G protein-coupled receptors [
2] that includes yet another receptor, PAC
1, that has a significantly higher affinity for PACAP than for VIP.
The hippocampal actions of VIP are largely dependent on GABAergic transmission [
2,
5,
6] and involve multiple cellular and molecular targets, as expected from the expression of VIP in different interneuron populations with different target selectivity [
2,
6] and from the opposing actions of VIP on exocytotic GABA release when activating VPAC
1 or VPAC
2 receptors [
7]. Nevertheless, VIP enhancement of synaptic transmission to CA1 pyramidal cell dendrites is mostly mediated by VPAC
1 receptors located in the
strata oriens and
radiatum [
2,
8,
9]. This, in turn, triggers disinhibition of pyramidal cell dendrites by inhibiting dendritic-targeting GABAergic interneurons [
6]. Notwithstanding, VIP also enhances pyramidal cell excitability in the absence of GABAergic transmission [
6,
10], an effect believed to occur through the activation of VPAC
2 receptors in pyramidal cell bodies [
11].
Long-term potentiation (LTP) as induced by theta-burst stimulation (TBS), a form of activity-dependent synaptic plasticity, is a key cellular mechanism for memory storage [
12,
13]. TBS is a sequence of electrical stimuli that mimics CA3 and CA1 pyramidal complex-spike cell discharges perceived during the hippocampal theta rhythm (3–7 Hz). This EEG pattern is linked to hippocampal spatial memory formation believed to work as a ‘tag’ for short-term memory processing [
13,
14]. TBS triggers an early LTP, acting through suppression of feedforward inhibition by a priming burst/single pulse. This allows for enough depolarization to trigger NMDA receptors [
15,
16], through an indirect mechanism that involves activation of GABA
B autoreceptors, thus suppressing GABA release from feedforward interneurons [
17] and synaptic GABA availability.
Endogenous VIP, acting on VPAC
1 receptors, is an important endogenous modulator of NMDA-dependent TBS-induced LTP as well as LTD and depotentiation in the CA1 area of the hippocampus [
3,
4]. This is a prospective indirect effect through modulation of disinhibition since it is fully dependent on GABAergic transmission. Yet this, together with previous reports that VIP directly enhances NMDA currents in CA1 pyramidal cells, an effect mimicked by VPAC
2 and to a lesser extent by VPAC
1 selective agonists [
18], highlights the diversity and importance of VIP mechanisms in the regulation of hippocampal synaptic plasticity phenomena. Accordingly, VIP is a crucial endogenous modulator of several hippocampal-dependent learning tasks [
19,
20,
21,
22,
23]. Furthermore, VIP-deficient mice fail to develop hippocampal-dependent learning skills like reversal learning [
24] while the VIP knockout is unviable. VIP interneuron dysfunction is also implicated in three prominent neurodevelopmental disorders, Rett syndrome, Dravet Syndrome, and Down’s syndrome [
25,
26,
27], and may contribute to enhanced seizure susceptibility and epileptogenesis associated with altered synaptic plasticity in these developmental disorders as in other acquired epilepsies [
2,
28,
29,
30].
The activity of hippocampal GABAergic interneurons influences not only the balance between excitation and inhibition but also a complex network of circuits regulating feedforward, feedback, and disinhibitory mechanisms [
31]. Synaptic inhibition at pyramidal cell dendrites is crucial for selective Ca
2+-dependent input selectivity and precision of LTP induction [
32] yet the long-lasting plasticity responses of interneurons to mild TBS remain largely unknown. Accumulated evidence suggests that interneurons serve as mediators of normal circuit development by regulating critical period plasticity and shaping the formation of sensory maps [
31]. Furthermore, although interneurons are present in the hippocampus from early postnatal development, a few of these, like parvalbumin-expressing interneurons only fully develop and set in human hippocampal circuits until adulthood [
33] This can dramatically influence the dynamics of hippocampal inhibition from weaning to adulthood, and its control of synaptic plasticity phenomena. We showed recently that TBS-induced LTP undergoes post-weaning developmental maturation until adulthood [
34], and preliminary data from our lab suggests this may be related to postweaning maturation of GABAergic circuits [
35].
In this paper, we investigated the postweaning developmental changes in VPAC1 receptor modulation of hippocampal LTP induced by mild TBS by endogenous VIP and their correlation with developmental changes in the expression of VIP and VIP receptors, GABAergic and glutamatergic pre and postsynaptic markers and overall synaptic density.
2. Material and Methods
The experiments were performed in
juvenile (3 weeks old),
young adult (6–7 weeks old), and
adult (12 weeks old) male outbred Wistar rats (Charles River, Barcelona, Spain) essentially as previously described [
3,
34] and all protocols and procedures were performed according to ARRIVE guidelines for experimental design, analysis, and their reporting. Animal housing and handling was performed in accordance with Portuguese law (DL 113/2013) and European Community guidelines (86/609/EEC and 63/2010/CE). The animals were anesthetized with fluothane, decapitated, and the right hippocampus dissected free in ice-cold artificial cerebrospinal fluid (aCSF) of the following composition in mM: NaCl 124, KCl 3, NaH
2PO
4 1.25, NaHCO
3 26, MgSO
4 1.5, CaCl
2 2, glucose 10, and gassed with a 95% O
2–5% CO
2 mixture.
2.1. LTP Experiments
Hippocampal slices (400 µm thick) were cut perpendicularly to the long axis of the hippocampus with a McIlwain tissue chopper (Campden Instruments, Loughborough, UK), then they were kept in a resting chamber in gassed aCSF at room temperature 22 °C–25 °C for at least 1 h to allow energetic and functional recovery. Each slice was transferred at a time to a submerged recording chamber of 1 mL capacity and continuously superfused at a rate of 3 mL/min with gassed aCSF at 30.5 °C. To obtain electrophysiological recordings slices were stimulated (rectangular pulses of 0.1 ms) through bipolar concentric wire electrodes placed in two separate sets (S1 and S2) of the Schaffer collateral/commissural fibres in the
stratum radiatum (
Figure S1). Responses were evoked alternately on the two pathways every 10 s and thus each pathway was stimulated every 20 s (0.05 Hz). Evoked field excitatory post-synaptic potentials (fEPSPs) were recorded extracellularly from CA1
stratum radiatum using aCSF-filled micropipettes. The initial stimulus intensity was chosen to elicit a field excitatory post-synaptic potential (fEPSP) of 600–900 μV amplitude (about 50% of the maximal response) while minimizing contamination by the population spike, and of similar magnitude in both pathways. The averages of six consecutive fEPSP responses from each pathway were obtained, measured, graphically plotted, and recorded for further analysis with a personal computer using the WinLTP software (version 2.96, WinLTP Ltd., Bristol, UK) [
36]. The slope of the initial phase of the potential was used to quantify fEPSPs intensity. At the end of the experiments, the independence of the two pathways was tested by studying paired-pulse facilitation (PPF) across both pathways, with less than 10% facilitation being usually observed. PPF was elicited by stimulating the two Schaffer pathways with a 50 ms interval between pulses. The ratio P2/P1 between the fEPSP slopes elicited by the second P2 and the first P1 stimuli was used to quantify synaptic facilitation.
When a stable fEPSP slope baseline was observed for at least 20 min, LTP was induced either by mild TBS (five trains of 100 Hz, 4 stimuli, separated by 200 ms) or a moderate TBS (fifteen trains of 100 Hz, 4 stimuli, separated by 200 ms). The intensity of the stimulus was not changed during these stimulation protocols. LTP was taken as the % change in the average slope of fEPSPs observed from 50 to 60 min after the induction protocol, in relation to one measured during the 10 min that preceded TBS. Control and test conditions were tested in independent pathways in the same slice. In all experiments, S1 always refers to the first pathway (left or right, randomly assigned) to which TBS was applied. Test drugs were added to the perfusion solution 20 min before TBS stimulation of the test pathway (S2) and were present until the end of the experiment.
2.2. Western Blot Analysis of Synaptic Proteins
The hippocampi of 3 week-old, 6–7-week-old, and 12–14 week-old rats were collected in sucrose solution (320 mM Sucrose, 1 mg/mL BSA, 10 mM HEPES e 1 mM EDTA, pH 7.4) containing protease (complete, mini, EDTA-free Protease Inhibitor Cocktail, Sigma, St. Louis, MO, USA) and phosphatase (1 mM PMSF, 2 mM Na
3VO
4, and 10 mM NaF) inhibitors, homogenized with a Potter–Elvejham apparatus (Labbox, Barcelona, Spain) and either total hippocampal membranes or hippocampal synaptosomes were isolated as described [
6,
34]. For isolation of total hippocampal membranes, the hippocampal homogenates were centrifuged at 1500×
g for 10 min. The supernatant was collected and further centrifuged at 14,000×
g for 12 min. The pellet was washed twice with modified aCSF (20 mM HEPES, 1 mM MgCl
2, 1.2 mM NaH
2PO
4, 120 mM NaCl; 2.7 mM KCl, 1.2 mM CaCl
2, 10 mM glucose, pH 7.4) also containing protease and phosphatase inhibitors and resuspended in 300 µL modified aCSF per hippocampus. For isolation of hippocampal synaptosomes, homogenates were centrifuged at 3000×
g for 10 min at 4 °C; the supernatant was then centrifuged at 14,000×
g for 12 min at 4 °C and the pellet resuspended in 3 mL of a Percoll 45% (
v/
v) in modified aCSF. The top layer (synaptosomal fraction) obtained after centrifugation at 14,000×
g for 2 min at 4 °C was washed twice with aCSF and resuspended in 200 µL modified aCSF per hippocampus. Aliquots of these suspensions of hippocampal membranes/synaptosomes were snap-frozen in liquid nitrogen and stored at −80 °C until use.
For Western blot, samples incubated at 95 °C for 5 min with Laemmli buffer (125 mM Tris-BASE, 4% SDS, 50% glycerol, 0.02% Bromophenol Blue, 10% β-mercaptoethanol), were run on standard 12% sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to PVDF membranes (pore size 0.45 μm, GE Healthcare Life Sciences, Marlborough, MA, USA). These were then blocked for 1 h with either a 3% BSA solution or 5% milk solution in Tris-buffered saline (20 mM Tris, 150 mM NaCl) containing 0.1% Tween-20 (TBST), and incubated overnight at 4 °C with and either rabbit polyclonal anti-VPAC1 (1:600, Alomone Labs (Jerusalem, IL, USA) #AVR-001, RRID: AB_2341081), rabbit polyclonal anti-VPAC2 (1:500, Alomone Labs #AVR-002, RRID: RRID: AB_2341082), rabbit polyclonal anti VIP (1:300, Proteintech Europe (Manchester, UK), # 16233-1-AP, RRID: AB_2878233), mouse monoclonal anti-gephyrin (1:3000, #147011, Synaptic Systems (Göttingen, Germany), AB_2810214), rabbit polyclonal anti-PSD-95 (1:750, #CST-2507, Cell Signalling Tech. (Danvers, MA, USA), AB_561221), rabbit polyclonal anti-synaptophysin (1:7500, Synaptic Systems #101002, RRID:AB_887905), rabbit polyclonal anti-VGAT (1:2500, Synaptic Systems #131002, RRID: AB_887871), rabbit polyclonal anti-VGlut1 (1:3000, Synaptic Systems #135302, RRID: AB_887877), and either mouse monoclonal anti-β-actin (1:5000, Proteintech #60008-1-Ig, RRID: AB_2289225) or rabbit polyclonal anti-alpha-tubulin (1:5000, Proteintech #11224-1-AP; RRID: AB_2210206) primary antibodies. After washing the membranes were incubated for 1 h with anti-rabbit or anti-mouse IgG secondary antibody both conjugated with horseradish peroxidase (HRP) (Proteintech) at room temperature. HRP activity was visualized by enhanced chemiluminescence with Clarity ECL Western Blotting Detection System (Bio-Rad, Richmond, CA, USA). The intensity of the bands was evaluated with the Image J software (version 1.52a, public domain, NIH, USA). Alpha-tubulin band density was used as a loading control.
2.3. Materials
PG 97-269 and PG 99-465, (Phoenix peptides, Europe) were made up of 0.1 mM stock solution in CH3COOH 1% (v/v−1). The maximal CH3COOH concentration added to the slices, 0.001% (v/v) was devoid of effects on fEPSP slope (n = 4). Aliquots of the stock solutions were kept frozen at −20 °C until use. In each experiment, one aliquot was thawed and diluted in aCSF.
2.4. Statistics
LTP values are depicted as the mean ± S.E.M of n experiments. Each n represents a single experiment performed in slices obtained from one different animal for LTP experiments and experiments performed in duplicate for each single animal in Western blot studies. The significance of the differences between the means was calculated using the paired Student’s t-test when comparing control and test conditions for LTP expression in electrophysiological studies, or with repeated measures ANOVA with Tukey’s post-hoc test (when F was significant) when comparing the levels of synaptic proteins in different age groups. p values of 0.05 or less were considered to represent statistically significant differences.
3. Results
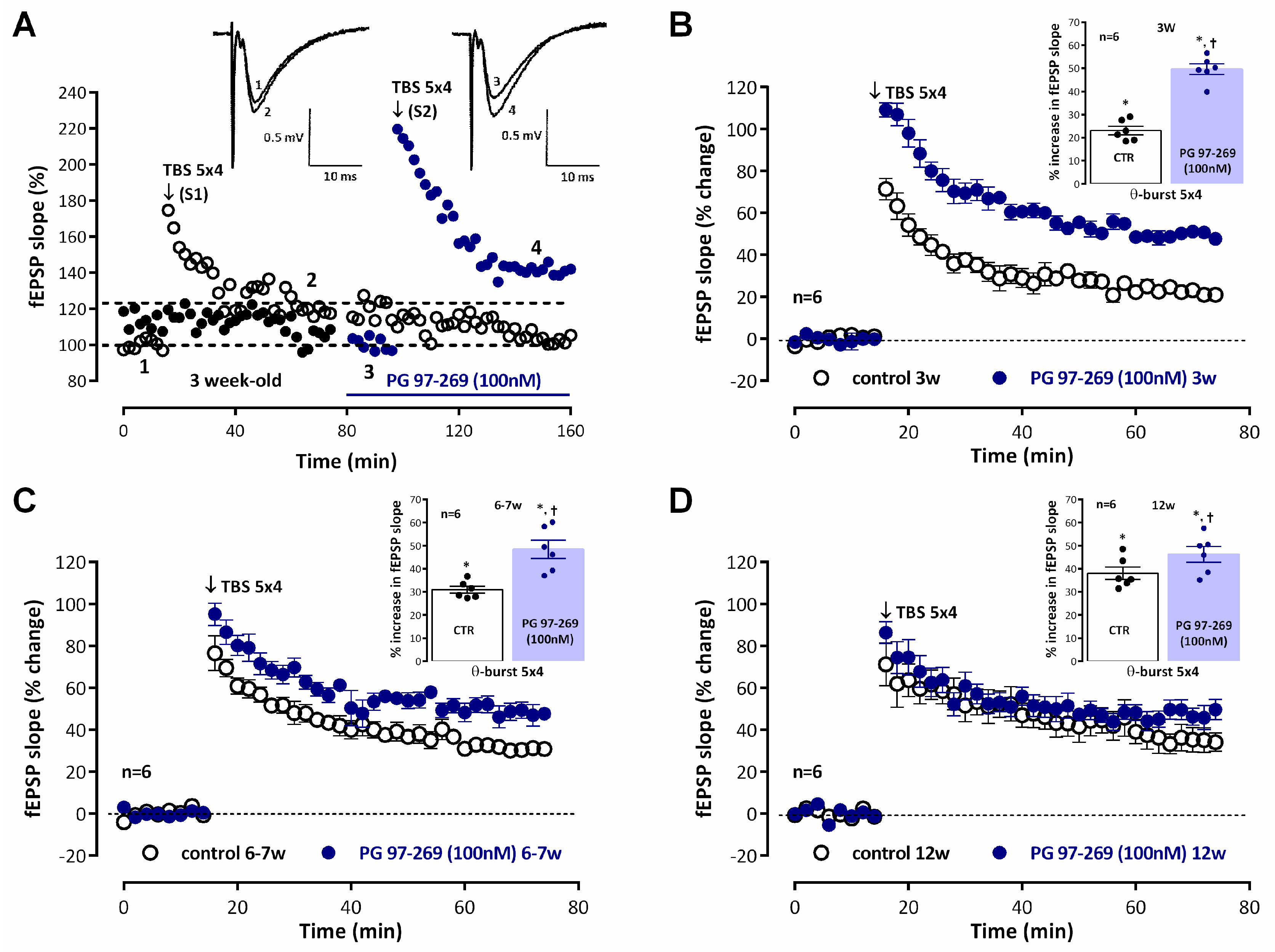
Recordings of fEPSP obtained under basal stimulation conditions (40–60% of the maximal response in each slice) in hippocampal slices from
juvenile (3-week-old) rats (
Figure 1A, raw data from a single experiment) showed an average slope of 0.579 ± 0.036 mV/ms (
n = 12). When
mild TBS (5 × 4 pulses with a 200 ms interval) was applied to the control pathway (S1) an LTP was induced, corresponding to a 25.6 ± 1.9% increase in fEPSP slope (
n = 12,
p < 0.05) observed 50–60 min after TBS. Application of a second
mild TBS train in the test pathway (S2) in the absence of drugs always resulted in an LTP of similar magnitude (% increase in fEPSP slope of 26.1 ± 3.4%,
n = 4) to the one observed in the control pathway (S1), i.e., LTP obtained under these experimental conditions was similar in S1 and S2. When
mild TBS of the test pathway (S2) was delivered in the presence of the VPAC
1 receptor antagonist PG 97-269 (100 nM) LTP was almost twice as large, since the enhancement of fEPSP slope observed 1 h later was 49.6 ± 2.3%, (
n = 6), suggesting a strong inhibition of LTP by endogenous VIP acting on VPAC
1 receptors. This effect was larger than the one previously observed in
young adult rats (6–7 weeks old) [
4]. In this study, fEPSPs in
young adult rats had an average slope of 0.553 ± 0.022 mV/ms (
n = 17). Induction of LTP with a
mild TBS caused a larger potentiation of the fEPSP slope (% increase in fEPSP slope 30.0 ± 1.2%,
n= 12,
p < 0.05,
Figure 1C) than the one observed in
juvenile rats as previously described [
34]. When
mild TBS was delivered to S2 in the presence of the VPAC
1 receptor antagonist PG 97-269 (100 nM), this resulted in an enhancement of LTP (% increase in fEPSP slope of 48.4 ± 3.9%,
n = 6,
Figure 1C) that was less pronounced than the one observed in
juvenile rats (
Figure 1B). In
adult rats (12 weeks old) the average fEPSP slope was 0.582 ± 0.017 mV/ms (
n = 16). The same
mild TBS paradigm caused an even larger LTP since
mild TBS now induced a potentiation of the fEPSP slope of 38.1 ± 2.7%, n = 6,
p < 0.05,
Figure 1D). When LTP was induced in the presence of the VPAC
1 receptor antagonist PG 97-269 (100 nM) the enhancement of LTP was even more reduced, since
mild TBS induced an enhancement of 46.3 ± 3.4% (
n = 6,
Figure 1D) in fEPSP slope. PG 97-269 (100 nM) did not significantly change basal synaptic transmission when added to the hippocampal slices.
Our previous studies did not show any influence of endogenous VIP on LTP induced by
mild TBS through activation of VPAC
2 receptors in
young adult rats [
4]. However, there could be post-weaning developmental changes in this respect. As shown in
Figure 2, when LTP was induced by
mild TBS in the presence of the selective VPAC
2 receptor antagonist PG 99-465 (100 nM) there was a mild but significant enhancement of LTP in
juvenile rats for which an enhancement of 34.2 ± 3.4% (
n = 5,
Figure 2A) in fEPSP slope was observed upon
mild TBS stimulation. An even milder tendency to an increase in LTP by VPAC
2 receptor blockage was observed in
young adult and
adult rats, but it was not consistently observed in all animals (
Figure 2B,C), and this effect did not reach statistical significance. When added to the hippocampal slices PG 99-465 (100 nM) did not significantly change basal synaptic transmission.
Increasing the number of bursts to 15 (
moderate TBS) enhanced the resulting potentiation evaluated 50–60 min after TBS, leading to a 46.2 ± 1.8% (
n = 5,
p < 0.05) enhancement in fEPSP slope (
Figure 3A) in
young adult rats and to a 63.6 ± 3.3% (
n = 5,
p < 0.05) enhancement in fEPSP slope (
Figure 3B) in
adult rats, as previously described [
34]. Using this stronger LTP induction paradigm the influence of endogenous VIP, acting on VPAC
1 receptors, on LTP expression was altered since the potentiation induced in fEPSP slope by
moderate TBS in the presence of the selective VPAC
1 receptor antagonist PG 97-269 (100 nM) was of 59.9 ± 3.1% (
n = 5,
p < 0.05,
Figure 3A) in
young adult rats and of a 56.5 ± 4.4% (
n = 5,
p < 0.05,
Figure 3B) in
adult rats, thus displaying a much lower potentiation of LTP by the VPAC
1 receptor antagonist in
young adult rats and a mild but non-significant reversal of the effect in
adult rats, suggesting that a developmental reshaping of GABAergic synapses, VIPergic interneurons or its activation pathways may be occurring or that VIP receptor expression and its role in hippocampal GABAergic synapses may be altered.
To elucidate the changes in VIP, VIP receptors, GABAergic, and glutamatergic synapses during postweaning development in the hippocampus we studied the levels of VIP VPAC
1 and VPAC
2 receptors and synaptic glutamatergic and GABAergic markers by Western blot in total hippocampal membranes. The synaptic changes in glutamatergic and GABAergic transmission were inferred from the evolution of GABAergic and glutamatergic pre and postsynaptic markers and its relation to broad-spectrum presynaptic marker synaptophysin, which was taken as an estimator of the overall synaptic density in total hippocampal membranes. The levels of the synaptic vesicle integral protein synaptophysin enhanced from weaning to adulthood by 63.9 ± 14.9% (
n = 5,
p < 0.05,
Figure 4I). The levels of both pre- and post-synaptic glutamatergic synaptic proteins VGlutT-1 and PSD-95 were enhanced by 29.9 ± 6.6% (
n = 5,
p < 0.05,
Figure 4A,C) and 44.5 ± 11.0% (
n = 5,
p < 0.05,
Figure 4A,B), respectively, in relation to the observed protein levels at 3 weeks of age. The increase in pre- and post-synaptic GABAergic synaptic proteins VGAT and gephyrin was more pronounced since VGAT and gephyrin levels enhanced by 71.1 ± 16.0% (
n = 5,
p < 0.05,
Figure 4E,G) and 55.4 ± 10.9% (
n = 5,
p < 0.05,
Figure 4E,F), respectively, in relation to the protein levels observed at 3 weeks, the age of weaning. By plotting the VGlut1/PSD-95 and VGAT/gephyrin ratios (
Figure 4D,H) we could observe that the ratios do not significantly change along postweaning development, suggesting that the increase in these synaptic markers is indeed representative of an increase in the number of synapses. To estimate the rate of change of glutamatergic and GABAergic synapses vs global synaptic changes in hippocampal tissue we calculated the VGlut1/synaptophysin and the VGAT/synaptophysin immunoreactivity ratios (
Figure 4K,L). The decrease in the hippocampal VGlut1/synaptophysin ratio from 3 to 12 weeks suggests that glutamatergic synapses do not accompany the global increase in new synapses while GABAergic synapses seem to contribute to this increase in hippocampal synaptic content, as estimated by a stable VGAT/synaptophysin ratio from 3 to 12 weeks.
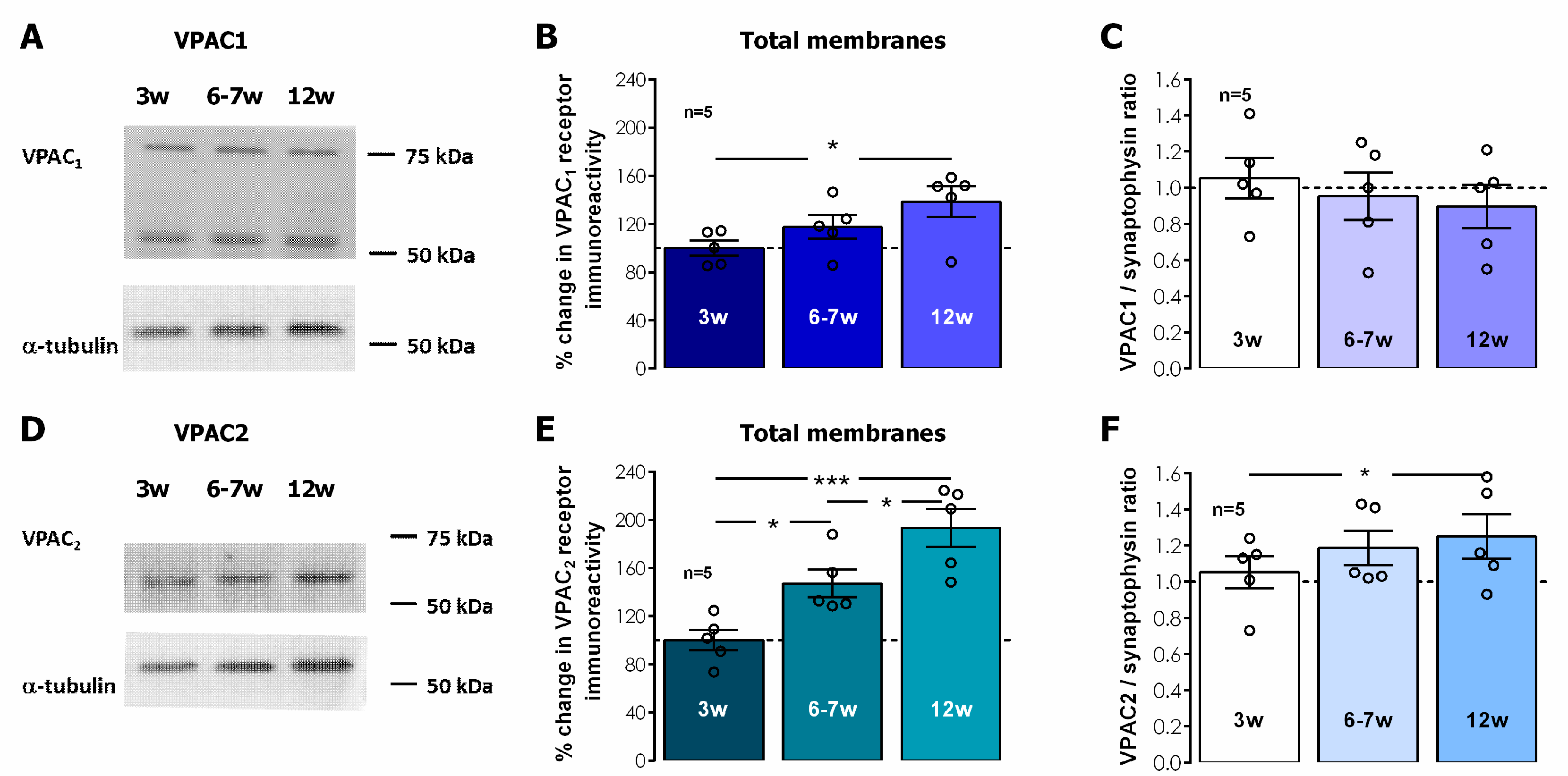
In total hippocampal membranes, VPAC
1 receptor immunostaining was detected by two independent bands located at 55 and 75 KDa, respectively, likely reflecting two different N-glycosylation levels, a feature that is also relevant for VPAC
1 membrane targeting, as previously described [
37]. The levels of both VPAC
1 and VPAC
2 receptors in total hippocampal membranes displayed a gradual postweaning developmental enhancement, that reached 42.5 ± 8.9% enhancement in 12-week-old rats for VPAC
1 receptors (
Figure 5A,B,
n = 5,
p < 0.05) and was even more pronounced for VPAC
2 receptors, for which the receptor levels nearly doubled (% increase 93.5 ± 15.7,
Figure 5D,E,
n = 5,
p < 0.05) in 12-week-old rats when compared to 3-week-old animals. By plotting the VPAC
1/ and VPAC
2/synaptophysin ratios (
Figure 5C,F) we were able to infer that the increase in VPAC
1 receptors accompanied the overall gain in synaptophysin yet VPAC
2 receptors were increasing even further, suggesting this increase was occurring also in non-synaptic locations.
To further clarify the cellular location of these receptors and their relevance for synaptic transmission and synaptic plasticity we performed also Western blot experiments to probe for VIP, VPAC
1, and VPAC
2 receptors in hippocampal synaptosomes, a preparation that is composed mainly of presynaptic nerve terminals, that often maintain attached part of the postsynaptic density [
38]. In hippocampal synaptosomes, both VIP and to a lesser extent VPAC
1 receptors increased along post-weaning development by 58.3 ± 15.9% and 43.7 ± 10.1% (
n = 5,
p < 0.05,
Figure 6A,B,E,F) at 12 weeks of age, respectively, suggesting that VIP and VPAC
1 containing nerve terminals enhance substantially along postweaning development. Conversely, VPAC
2 receptors did not significantly change hippocampal synaptosomes from 3- to 12-week-old rats (% change at 12 weeks vs. 3 weeks 20.3 ± 11.7%,
n = 5,
p > 0.05,
Figure 6C,D).
4. Discussion
The main findings of the present work are that: (1) VPAC1 receptor-mediated modulation of LTP induced by mild TBS in the hippocampal CA1 area by endogenous VIP is progressively weaker from weaning (3 week-old) through adulthood (12-week-old) in Wistar rats; (2) A stronger LTP-inducing train (moderate TBS) elicited an increased magnitude and stability of LTP in young-adult (6–7-week-old) and adult (12-week-old) Wistar rats, that was even less influenced by endogenous VIP acting on VPAC1 receptors; (3) The LTP elicited by mild TBS was only weakly dependent on endogenous VPAC2 receptor activation in juvenile (3-week-old) rats; (4) These observations were concurrent with an increase in both GABAergic and glutamatergic pre- and post-synaptic markers in total hippocampal membranes from weaning to adulthood, the enhancement in GABAergic being more prominent; (5) There was an increase in both VPAC1 and VPAC2 receptor levels in total hippocampal membranes during this developmental period while (6) VIP levels enhanced markedly, while only VPAC1, but not VPAC2, receptor levels were mildly enhanced in adult (12-week-old) vs juvenile (3-week-old) rats.
In the present study
mild TBS (5 bursts, 4 pulses delivered at 100 Hz), an LTP-inducing stimulation pattern, elicited a long-lasting potentiation of fEPSP slope that was increasingly larger in 3-, 6–7-, and 12-week-old rats, as previously described by our group [
34]. This is consistent with previous studies showing that during postnatal development there is a reinforcement of the cellular mechanisms leading to LTP expression and stability [
13,
39,
40]. Stronger TBS, achieved by increasing the number of bursts to 15 (
moderate TBS), enhanced proportionally the resulting potentiation for all age groups studied [
34].
Mild TBS elicits an LTP that is fully dependent on NMDA and partially dependent on GABA
B receptor activation for its induction in our model [
34] and is of ideal magnitude for pharmacological studies aiming to improve LTP outcome, while
moderate TBS elicits a near-maximal LTP for this stimulation pattern [
13]. Suppression of feed-forward phasic inhibition, mediated by GABA
B autoreceptor inhibition of GABA release, underlies induction of LTP by TBS in excitatory synapses of the hippocampus [
13,
41,
42]. This allows for temporal summation of excitation and sustained depolarization ultimately activating NMDA receptors, a crucial mechanism for TBS LTP induction.
We previously described that LTP induced by
mild TBS-induced LTP is inhibited by endogenous VIP acting on VPAC
1 receptors in young adult rats [
4]. This work further describes that VPAC
1 receptor blockade with PG 97-269 elicited an enhancement of LTP and that effect was progressively smaller from weaning, at 3 weeks of age, to adulthood, at 12 weeks of age. In addition, the LTP obtained in the presence of PG 97-269 was very similar for all age groups (46.3–49.6%) while LTP induced by
moderate TBS, an LTP that is near maximal for this stimulation pattern (56.5–59.9%) [
13] could not be further enhanced by VPAC
1 receptor blockade. Last, the levels of VPAC
1 receptors in both total hippocampal membranes and hippocampal synaptosomes increased from weaning to adulthood. Altogether, this suggests that the decrease in endogenous VIP control of hippocampal CA1 LTP is not due to changes in VPAC
1 receptor levels and that enhanced LTP afforded by VPAC
1 receptor blockade reaches a near maximum value for
mild TBS-induced LTP. This is also consistent with previous observations of a developmental reinforcement of the cellular mechanisms leading to LTP expression and stability [
13,
39,
40].
Conversely, as previously described in young-adult rats [
4], VPAC
2 receptor blockade with PG 99-465 did not change LTP of synaptic transmission induced by
mild TBS in the CA1 area of the hippocampus except for a mild increase in
juvenile rats, suggesting that endogenous VIP controlling CA1 LTP does not significantly activate VPAC
2 receptors. This does not preclude an important role for VPAC
2 receptors in the control of other hippocampal plasticity of neuronal excitability mechanisms, such as modulation of pyramidal cell excitability, as inferred from the main localization of these receptors at the
stratum pyramidale in the CA1 [
8,
11,
18]. Nevertheless, the enhancement in VPAC
2 receptor levels observed from weaning to adulthood in total hippocampal membranes was not reproduced in hippocampal synaptosomes, suggesting that it may be occurring either in non-neuronal cells or away from synaptic sites. Astrocytes are important players in neuronal communication and express functional VPAC
2 receptors regulating glutamate uptake [
43,
44], suggesting a major influence in the control of hippocampal synaptic activity and plasticity by endogenous VIP. This should be further investigated but is beyond the scope of the current paper.
Feed-forward phasic inhibition to CA1 pyramidal cells is mediated by physiologically and morphologically distinct GABAergic interneurons [
45,
46] and its TBS-induced suppression, mediated by GABA
B autoreceptors, is important, but not fundamental, for TBS-induced CA1 LTP [
17,
34], as
mild TBS also activates postsynaptic mechanisms leading to a sustained potentiation of fast GABA
A synaptic transmission to pyramidal cells [
47]. The net result of all these mechanisms is potentiation of synaptic transmission by
mild TBS, and blocking GABA
A receptors using bicuculline elicits an enhancement of LTP in the CA1 area [
34,
48]. Altogether, this suggests that further excitatory and inhibitory pathways, like transient changes in tonic inhibition during LTP induction, may determine the outcome of LTP induced by TBS [
13,
49]. This is corroborated by observations that, both synaptic and non-synaptic inhibition can hinder spike backpropagation and Ca
2+ spikes in CA1 pyramidal cell dendrites [
32,
49], an essential mechanism in TBS-induced LTP.
In this study, we observed a developmental decrease in VPAC
1 receptor influence on hippocampal LTP, even though there was a small but significant synaptic enhancement in both VIP and VPAC
1 receptors, which could be considered a paradox. Furthermore, VPAC
1 receptors mediate modulation of hippocampal synaptic transmission and plasticity through mechanisms that are fully dependent on GABAergic transmission [
2,
3,
4], VPAC
1 receptors are important nerve terminal modulators of GABA release [
7] and VIP is present in the hippocampus exclusively in three distinct interneuron populations with very distinct target selectivity [
1,
50]. In view of all this, and of the fact that developmental changes in tonic GABAergic inhibition at pyramidal cell dendrites were associated with developmental changes in LTP expression [
51], with relevance for LTP induced by
mild TBS at developmental stages addressed in this study, the distinct capacity of the VPAC
1 receptor antagonist (and thus endogenous VIP) to influence the outcome of LTP induced by mild TBS may be conditioned by postweaning maturation of GABAergic circuits. In fact, we previously showed that VPAC
1 receptor-mediated inhibition of CA1 LTP by endogenous VIP is majorly occurring by curtailing disinhibition impinging on phasic feed-forward inhibition [
4]. As such, a developmental enhancement in the influence of tonic inhibition on LTP expression would reduce the influence of endogenous VIP acting on VPAC
1 receptors to modulate the same LTP.
Given that VPAC
1 receptors can also be activated by endogenous PACAP, postweaning alteration of PACAP levels could conceptually be the cause for altered endogenous modulation of LTP through VPAC
1 receptors [
2]. However, it is worth noticing that PACAP is mostly present in glutamatergic nerve terminals, and likely co-released with glutamate, while this study and our previous study in young adults show this effect to be mostly dependent on GABAergic transmission [
4]. Our previous work also excluded the endogenous action of PACAP on VPAC
1 receptors in what concerns the modulation of TBS-induced LTP [
4]. It is thus not likely that PACAP is involved in the postweaning changes in VPAC
1 receptor modulation of LTP here reported.
To further explore the possible influence of GABAergic system maturation during postweaning development on modulation of hippocampal LTP in our rat model, we also investigated the alterations in GABAergic and glutamatergic synapses. Changes in synaptic GABAergic and glutamatergic synaptic markers were studied in total hippocampal membranes, which allowed us to perceive the increase in the total number of synapses in the whole tissue, while analysis of the ratios between pre- and post-synaptic GABAergic or glutamatergic markers allowed us to ascribe these changes to synaptic rather than single-molecule alterations. In addition, synaptophysin levels were taken as a measure of the global increase in the number of synapses. With this strategy, we were able to demonstrate that the increase in the number of hippocampal GABAergic synapses is larger than the increase in the number of glutamatergic synapses during this post-weaning developmental period. Studies on the maturation of hippocampal GABAergic transmission and its relations to GABAergic synapse establishment and synaptic plasticity are more often focused on early preweaning brain development [
52], but accumulated evidence suggests that the maturation of hippocampal circuits continues to progress into childhood and adolescence [
53]. This involves not only the maturation of resting properties and the setting up of new or altered synaptic connections but also the migration and insertion of new neurons, and involved most prominently interneurons and their synaptic spines [
33,
53,
54]. The observations in the present work are consistent with these studies.
The findings in this paper are of particular interest for the understanding of alterations of hippocampal synaptic plasticity in several developmental brain pathologies that have concomitantly been associated with alterations of VIP levels or VIP immunoreactive interneurons [
31,
55,
56]. VIP and VPAC
1 receptors are upregulated in mouse models of Down syndrome [
56], a disease associated with excessive GABAergic inhibition, which also displays impaired synaptic plasticity specifically in TBS but not high-frequency stimulation-induced LTP
in vitro [
57].
In vivo, the deficits in LTP and learning and memory deficits in this model can be rescued by selective GABA antagonists and an α5-selective GABA
A inverse agonist [
58,
59], a subunit associated with both tonic and synaptic inhibition by GABA
A receptors in the hippocampus [
60]. Enhanced VIP and VPAC
1 receptor levels may also be involved in restraining disinhibition contributing to impaired LTP in this model. Furthermore, VIP interneuron impairment promotes
in vivo circuit dysfunction in the cerebral cortex and autism-related behaviors in Dravet syndrome, a disease caused by mutation of the gene coding for voltage-gated sodium channel subunit Nav1.1, and in Rett syndrome, associated with mutations in the gene coding for methyl-CpG binding protein 2 (MECP2) located on the X chromosome, and both associated with recurrent seizures that ultimately lead to epilepsy [
31]. Although the firing patterns of irregular spiking VIP expressing interneurons were altered in a mouse model of Dravet syndrome (Scn 1a
+/−) [
25], suggesting this could contribute to Dravet syndrome cognitive pathogenesis, the alterations in the hippocampus are likely secondary to the development of seizures but should be further investigated, especially in what regards the development of epilepsy, since it has been recently demonstrated that inhibition of VIP-expressing interneurons of the ventral
subiculum was sufficient to reduce seizures in the intrahippocampal kainic acid model of epilepsy, thus suggesting a prominent role of these cells in seizure propagation and epileptogenesis [
30].
In conclusion, this paper reveals the postweaning alterations in VPAC1 receptor modulation of TBS-induced LTP by endogenous VIP in the rat hippocampus. Besides providing us with important insights into VPAC1 receptors as putative drug targets aiming to improve synaptic plasticity and ultimately memory, this study also sheds light into the ongoing physiological adaptations of GABAergic transmission and in the control of synaptic plasticity during postweaning development, a developmental period that is often overlooked in brain development. Understanding these physiological adaptations may prove useful to also understand the pathophysiology of neurodevelopmental disorders affecting the hippocampus thus providing a better guide to the discovery of new therapeutic targets.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





