
Effects of Cannabidiol, ∆9-Tetrahydrocannabinol, and WIN 55-212-22 on the Viability of Canine and Human Non-Hodgkin Lymphoma Cell Lines

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Canine Peripheral Blood Mononuclear Cells
2.3. Lymphoma Cell Lines
2.4. Cell Culture
2.5. Treatment Design for Evaluating Lymphoma Cell Viability and Mechanisms of Cannabinoid-Induced Cytotoxicity
2.6. Evaluation of Cell Viability
2.7. IC50 Calculation
2.8. Total Protein Quantification
2.9. Biochemical Analysis
2.9.1. Hydrogen Peroxide (H2O2) Content
2.9.2. Lipid Peroxidation
2.9.3. Reactive Oxygen Species (ROS) Generation
2.9.4. Nitrite Content
2.9.5. Glutathione (GSH) Content
2.9.6. NADH Content
2.9.7. Mitochondrial Complex-I Activity
2.9.8. Interleukin Converting Enzyme-I (ICE-1) Activity
2.9.9. Cyclooxygenase (COX) Activity
2.9.10. Caspase-3, -8, and -9 Activities
2.10. Statistical Analysis
3. Results
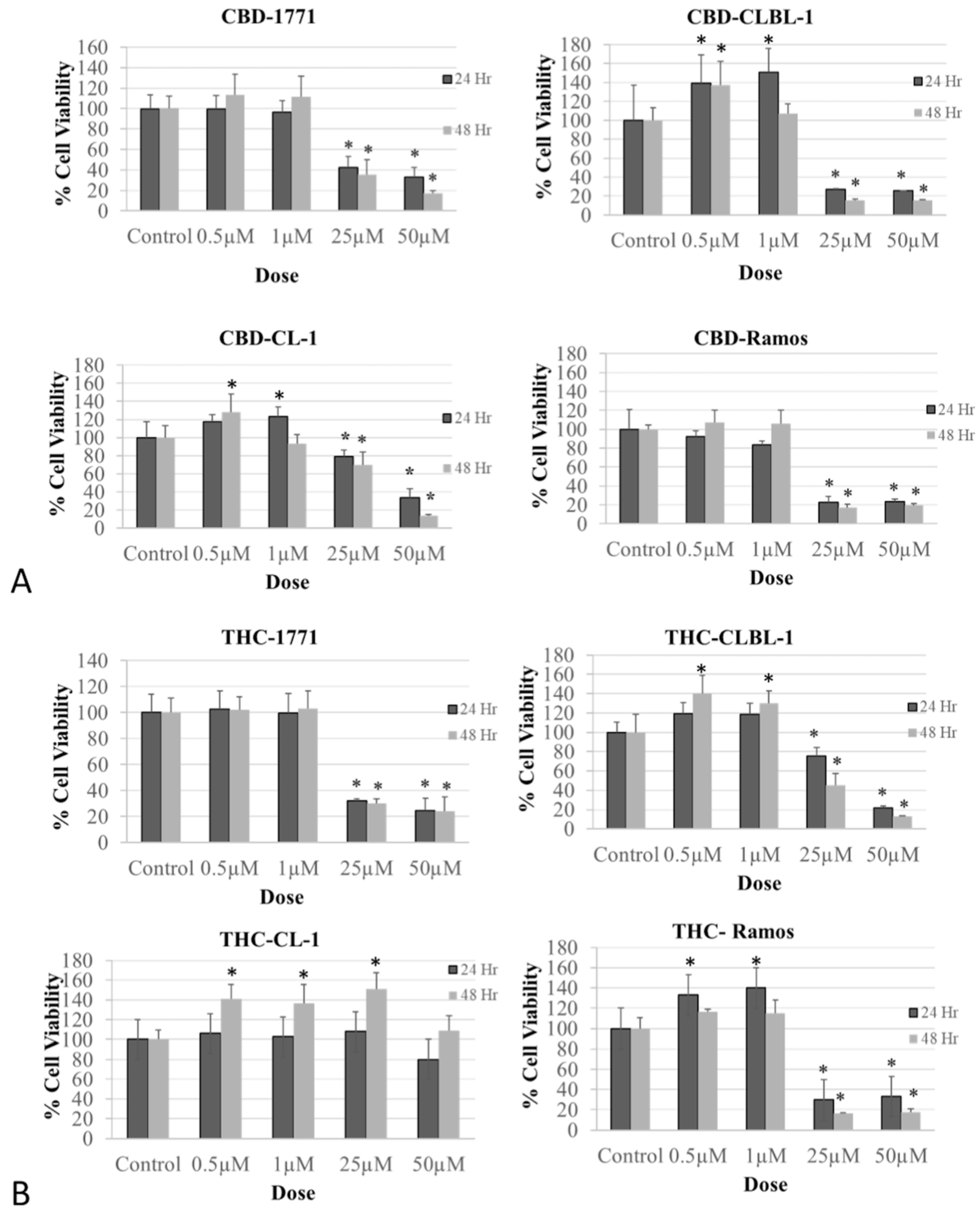
3.1. Biphasic Effect of CBD, THC, and WIN on Canine and Human Non-Hodgkin Lymphoma (NHL) Cell Viability
3.1.1. Treatment with CBD and THC Phytocannabinoids Altered Cell Viability in Human and Canine Lymphoma Cells
3.1.2. Treatment with WIN Altered NHL Cell Viability in Human and Canine Lymphoma Cells
3.1.3. Treatment with CBD Does Not Affect Healthy Lymphocytes’ Viability
3.2. IC50 Calculation of Phytocannabinoids and a Synthetic Cannabinoid Based on Canine and Human Non-Hodgkin Lymphoma (NHL) Cell Viability
3.3. CBD, THC, and WIN Treatments Induced Oxidative Stress in Canine 1771 B-Cell Lymphoma Cell Line
3.4. CBD, THC, and WIN Treatments Induced Inflammation in Canine 1771 B-Cell Lymphoma Cell Line
3.5. CBD, THC, and WIN Treatments Induced Apoptosis in Canine 1771 B-Cell Lymphoma Cell Line
3.6. Effects of CBD, THC, and WIN Cannabinoid Treatments on Mitochondrial Function
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ito, D.; Frantz, A.M.; Modiano, J.F. Canine lymphoma as a comparative model for human non-Hodgkin lymphoma: Recent progress and applications. Vet. Immunol. Immunopathol. 2014, 159, 192–201. [Google Scholar] [CrossRef] [PubMed]
- Zandvliet, M. Canine lymphoma: A review. Vet. Q. 2016, 36, 76–104. [Google Scholar] [CrossRef] [PubMed]
- Breen, M.; Modiano, J.F. Evolutionarily conserved cytogenetic changes in hematological malignancies of dogs and humans--man and his best friend share more than companionship. Chromosome Res. 2008, 16, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Castor, A.; Nilsson, L.; Astrand-Grundstrom, I.; Buitenhuis, M.; Ramirez, C.; Anderson, K.; Strombeck, B.; Garwicz, S.; Bekassy, A.N.; Schmiegelow, K.; et al. Distinct patterns of hematopoietic stem cell involvement in acute lymphoblastic leukemia. Nat. Med. 2005, 11, 630–637. [Google Scholar] [CrossRef] [PubMed]
- Challa, A.; Eliopoulos, A.G.; Holder, M.J.; Burguete, A.S.; Pound, J.D.; Chamba, A.; Grafton, G.; Armitage, R.J.; Gregory, C.D.; Martinez-Valdez, H.; et al. Population depletion activates autonomous CD154-dependent survival in biopsylike Burkitt lymphoma cells. Blood 2002, 99, 3411–3418. [Google Scholar] [CrossRef] [PubMed]
- Guzman, M. Cannabinoids: Potential anticancer agents. Nat. Rev. Cancer 2003, 3, 745–755. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, C.; de Ceballos, M.L.; Gomez del Pulgar, T.; Rueda, D.; Corbacho, C.; Velasco, G.; Galve-Roperh, I.; Huffman, J.W.; Ramon y Cajal, S.; Guzman, M. Inhibition of glioma growth in vivo by selective activation of the CB(2) cannabinoid receptor. Cancer Res. 2001, 61, 5784–5789. [Google Scholar] [PubMed]
- Bifulco, M.; Laezza, C.; Portella, G.; Vitale, M.; Orlando, P.; De Petrocellis, L.; Di Marzo, V. Control by the endogenous cannabinoid system of ras oncogene-dependent tumor growth. FASEB J. 2001, 15, 2745–2747. [Google Scholar] [CrossRef]
- Velasco, G.; Galve-Roperh, I.; Sanchez, C.; Blazquez, C.; Guzman, M. Hypothesis: Cannabinoid therapy for the treatment of gliomas? Neuropharmacology 2004, 47, 315–323. [Google Scholar] [CrossRef]
- Sarker, K.P.; Obara, S.; Nakata, M.; Kitajima, I.; Maruyama, I. Anandamide induces apoptosis of PC-12 cells: Involvement of superoxide and caspase-3. FEBS Lett. 2000, 472, 39–44. [Google Scholar] [CrossRef]
- Maccarrone, M.; Lorenzon, T.; Bari, M.; Melino, G.; Finazzi-Agro, A. Anandamide induces apoptosis in human cells via vanilloid receptors. Evidence for a protective role of cannabinoid receptors. J. Biol. Chem. 2000, 275, 31938–31945. [Google Scholar] [CrossRef]
- Blasco-Benito, S.; Seijo-Vila, M.; Caro-Villalobos, M.; Tundidor, I.; Andradas, C.; Garcia-Taboada, E.; Wade, J.; Smith, S.; Guzman, M.; Perez-Gomez, E.; et al. Appraising the “entourage effect”: Antitumor action of a pure cannabinoid versus a botanical drug preparation in preclinical models of breast cancer. Biochem. Pharmacol. 2018, 157, 285–293. [Google Scholar] [CrossRef]
- Milian, L.; Mata, M.; Alcacer, J.; Oliver, M.; Sancho-Tello, M.; Martin de Llano, J.J.; Camps, C.; Galbis, J.; Carretero, J.; Carda, C. Cannabinoid receptor expression in non-small cell lung cancer. Effectiveness of tetrahydrocannabinol and cannabidiol inhibiting cell proliferation and epithelial-mesenchymal transition in vitro. PLoS ONE 2020, 15, e0228909. [Google Scholar] [CrossRef]
- Sarfaraz, S.; Afaq, F.; Adhami, V.M.; Mukhtar, H. Cannabinoid receptor as a novel target for the treatment of prostate cancer. Cancer Res. 2005, 65, 1635–1641. [Google Scholar] [CrossRef]
- Patsos, H.A.; Greenhough, A.; Hicks, D.J.; Al Kharusi, M.; Collard, T.J.; Lane, J.D.; Paraskeva, C.; Williams, A.C. The endogenous cannabinoid, anandamide, induces COX-2-dependent cell death in apoptosis-resistant colon cancer cells. Int. J. Oncol. 2010, 37, 187–193. [Google Scholar] [CrossRef]
- Caffarel, M.M.; Andradas, C.; Mira, E.; Perez-Gomez, E.; Cerutti, C.; Moreno-Bueno, G.; Flores, J.M.; Garcia-Real, I.; Palacios, J.; Manes, S.; et al. Cannabinoids reduce ErbB2-driven breast cancer progression through Akt inhibition. Mol. Cancer 2010, 9, 196. [Google Scholar] [CrossRef]
- Guindon, J.; Hohmann, A.G. The endocannabinoid system and cancer: Therapeutic implication. Br. J. Pharmacol. 2011, 163, 1447–1463. [Google Scholar] [CrossRef]
- Zygmunt, P.M.; Petersson, J.; Andersson, D.A.; Chuang, H.; Sorgard, M.; Di Marzo, V.; Julius, D.; Hogestatt, E.D. Vanilloid receptors on sensory nerves mediate the vasodilator action of anandamide. Nature 1999, 400, 452–457. [Google Scholar] [CrossRef]
- Ramer, R.; Schwarz, R.; Hinz, B. Modulation of the Endocannabinoid System as a Potential Anticancer Strategy. Front. Pharmacol. 2019, 10, 430. [Google Scholar] [CrossRef]
- McAllister, S.D.; Christian, R.T.; Horowitz, M.P.; Garcia, A.; Desprez, P.Y. Cannabidiol as a novel inhibitor of ld-1 gene expression in aggressive breast cancer cells. Mol. Cancer Ther. 2007, 6, 2921–2927. [Google Scholar] [CrossRef]
- Omer, S.; Pathak, S.; Nadar, R.; Bowen, D.; Sandey, M.; Dhanasekaran, M.; Pondugula, S.; Mansour, M.; Boothe, D. Validating the anti-lymphoma pharmacodynamic actions of the endocannabinoids on canine non-Hodgkin lymphoma. Life Sci. 2023, 327, 121862. [Google Scholar] [CrossRef]
- Pondugula, S.R.; Ferniany, G.; Ashraf, F.; Abbott, K.L.; Smith, B.F.; Coleman, E.S.; Mansour, M.; Bird, R.C.; Smith, A.N.; Karthikeyan, C.; et al. Stearidonic acid, a plant-based dietary fatty acid, enhances the chemosensitivity of canine lymphoid tumor cells. Biochem. Biophys. Res. Commun. 2015, 460, 1002–1007. [Google Scholar] [CrossRef]
- McKallip, R.J.; Lombard, C.; Fisher, M.; Martin, B.R.; Ryu, S.; Grant, S.; Nagarkatti, P.S.; Nagarkatti, M. Targeting CB2 cannabinoid receptors as a novel therapy to treat malignant lymphoblastic disease. Blood 2002, 100, 627–634. [Google Scholar] [CrossRef]
- Massi, P.; Vaccani, A.; Ceruti, S.; Colombo, A.; Abbracchio, M.P.; Parolaro, D. Antitumor effects of cannabidiol, a nonpsychoactive cannabinoid, on human glioma cell lines. J. Pharmacol. Exp. Ther. 2004, 308, 838–845. [Google Scholar] [CrossRef]
- Guide for Biologists. Available online: https://www.sciencegateway.org/protocols/ (accessed on 2 March 2021).
- Sobotta, M.C.; Barata, A.G.; Schmidt, U.; Mueller, S.; Millonig, G.; Dick, T.P. Exposing cells to H2O2: A quantitative comparison between continuous low-dose and one-time high-dose treatments. Free Radic. Biol. Med. 2013, 60, 325–335. [Google Scholar] [CrossRef]
- Zheng, M.; Ahuja, M.; Bhattacharya, D.; Clement, T.P.; Hayworth, J.S.; Dhanasekaran, M. Evaluation of differential cytotoxic effects of the oil spill dispersant Corexit 9500. Life Sci 2014, 95, 108–117. [Google Scholar] [CrossRef]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef]
- Majrashi, M.; Almaghrabi, M.; Fadan, M.; Fujihashi, A.; Lee, W.; Deruiter, J.; Randall Clark, C.; Dhanasekaran, M. Dopaminergic neurotoxic effects of 3-TFMPP derivatives. Life Sci. 2018, 209, 357–369. [Google Scholar] [CrossRef]
- Hong, S.; Kim, C.Y.; Lee, J.E.; Seong, G.J. Agmatine protects cultured retinal ganglion cells from tumor necrosis factor-alpha-induced apoptosis. Life Sci. 2009, 84, 28–32. [Google Scholar] [CrossRef]
- Dhanasekaran, M.; Tharakan, B.; Manyam, B.V. Antiparkinson drug--Mucuna pruriens shows antioxidant and metal chelating activity. Phytother. Res. 2008, 22, 6–11. [Google Scholar] [CrossRef]
- Giustarini, D.; Rossi, R.; Milzani, A.; Dalle-Donne, I. Nitrite and nitrate measurement by Griess reagent in human plasma: Evaluation of interferences and standardization. Methods Enzymol. 2008, 440, 361–380. [Google Scholar] [CrossRef]
- Almaghrabi, M. Investigate the Dopaminergic Neurotoxicity Profile of Designer Drugs (Piperazine Derivatives). Ph.D. Thesis, Auburn University, Auburn, AL, USA, 2018. [Google Scholar]
- Ramsay, R.R.; Dadgar, J.; Trevor, A.; Singer, T.P. Energy-driven uptake of N-methyl-4-phenylpyridine by brain mitochondria mediates the neurotoxicity of MPTP. Life Sci. 1986, 39, 581–588. [Google Scholar] [CrossRef]
- Ahuja, M.; Buabeid, M.; Abdel-Rahman, E.; Majrashi, M.; Parameshwaran, K.; Amin, R.; Ramesh, S.; Thiruchelvan, K.; Pondugula, S.; Suppiramaniam, V.; et al. Immunological alteration and toxic molecular inductions leading to cognitive impairment and neurotoxicity in transgenic mouse model of Alzheimer’s disease. Life Sci. 2017, 177, 49–59. [Google Scholar] [CrossRef]
- Bhattacharya, D.; Majrashi, M.; Ramesh, S.; Govindarajulu, M.; Bloemer, J.; Fujihashi, A.; Crump, B.R.; Hightower, H.; Bhattacharya, S.; Moore, T.; et al. Assessment of the cerebellar neurotoxic effects of nicotine in prenatal alcohol exposure in rats. Life Sci. 2018, 194, 177–184. [Google Scholar] [CrossRef]
- Al Saqr, A.; Majrashi, M.; Alrbyawi, H.; Govindarajulu, M.; Fujihashi, A.; Gottumukkala, S.; Poudel, I.; Arnold, R.D.; Babu, R.J.; Dhanasekaran, M. Elucidating the anti-melanoma effect and mechanisms of Hispolon. Life Sci. 2020, 256, 117702. [Google Scholar] [CrossRef]
- Liu, K.; Kadannagari, S.; Deruiter, J.; Pathak, S.; Abbott, K.L.; Salamat, J.M.; Pondugula, S.R.; Akingbemi, B.T.; Dhanasekaran, M. Effects of developmental exposures to Bisphenol-A and Bisphenol-S on hepatocellular function in male Long-Evans rats. Life Sci. 2023, 326, 121752. [Google Scholar] [CrossRef]
- Copeland, R.A.; Williams, J.M.; Giannaras, J.; Nurnberg, S.; Covington, M.; Pinto, D.; Pick, S.; Trzaskos, J.M. Mechanism of selective inhibition of the inducible isoform of prostaglandin G/H synthase. Proc. Natl. Acad. Sci. USA 1994, 91, 11202–11206. [Google Scholar] [CrossRef]
- Usha, R.; Muralikrishnan, D.; Thomas, B.; Ghosh, S.; Mandal, C.; Mohanakumar, K.P. Region-specific attenuation of a trypsin-like protease in substantia nigra following dopaminergic neurotoxicity by 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine. Brain Res. 2000, 882, 191–195. [Google Scholar] [CrossRef]
- Ellert-Miklaszewska, A.; Ciechomska, I.A.; Kaminska, B. Cannabinoid Signaling in Glioma Cells. Adv. Exp. Med. Biol. 2020, 1202, 223–241. [Google Scholar] [CrossRef]
- Kovalchuk, O.; Kovalchuk, I. Cannabinoids as anticancer therapeutic agents. Cell Cycle 2020, 19, 961–989. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M. In Vitro Anticancer Activity of Plant-Derived Cannabidiol on Prostate Cancer Cell Lines. Pharmacol. Pharm. 2014, 5, 806–820. [Google Scholar] [CrossRef]
- McAllister, S.D.; Soroceanu, L.; Desprez, P.Y. The Antitumor Activity of Plant-Derived Non-Psychoactive Cannabinoids. J. Neuroimmune Pharmacol. 2015, 10, 255–267. [Google Scholar] [CrossRef] [PubMed]
- Jia, W.; Hegde, V.L.; Singh, N.P.; Sisco, D.; Grant, S.; Nagarkatti, M.; Nagarkatti, P.S. Δ9-Tetrahydrocannabinol-Induced Apoptosis in Jurkat Leukemia T Cells Is Regulated by Translocation of Bad to Mitochondria. Mol. Cancer Res. 2006, 4, 549–562. [Google Scholar] [CrossRef] [PubMed]
- McKallip, R.J.; Jia, W.; Schlomer, J.; Warren, J.W.; Nagarkatti, P.S.; Nagarkatti, M. Cannabidiol-induced apoptosis in human leukemia cells: A novel role of cannabidiol in the regulation of p22phox and Nox4 expression. Mol. Pharmacol. 2006, 70, 897–908. [Google Scholar] [CrossRef] [PubMed]
- Emery, S.M.; Alotaibi, M.R.; Tao, Q.; Selley, D.E.; Lichtman, A.H.; Gewirtz, D.A. Combined antiproliferative effects of the aminoalkylindole WIN55,212-2 and radiation in breast cancer cells. J. Pharmacol. Exp. Ther. 2014, 348, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Shah, S.A.; Gupta, A.S.; Kumar, P. Emerging role of cannabinoids and synthetic cannabinoid receptor 1/cannabinoid receptor 2 receptor agonists in cancer treatment and chemotherapy-associated cancer management. J. Cancer Res. Ther. 2021, 17, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Ellert-Miklaszewska, A.; Ciechomska, I.A.; Kaminska, B. Synthetic Cannabinoids Induce Autophagy and Mitochondrial Apoptotic Pathways in Human Glioblastoma Cells Independently of Deficiency in TP53 or PTEN Tumor Suppressors. Cancers 2021, 13, 419. [Google Scholar] [CrossRef] [PubMed]
- Flygare, J.; Gustafsson, K.; Kimby, E.; Christensson, B.; Sander, B. Cannabinoid receptor ligands mediate growth inhibition and cell death in mantle cell lymphoma. FEBS Lett. 2005, 579, 6885–6889. [Google Scholar] [CrossRef] [PubMed]
- Hodges, E.L.; Marshall, J.P.; Ashpole, N.M. Age-dependent hormesis-like effects of the synthetic cannabinoid CP55940 in C57BL/6 mice. npj Aging Mech. Dis. 2020, 6, 7. [Google Scholar] [CrossRef]
- Murakami, A. Hormesis-Mediated Mechanisms Underlying Bioactivities of Phytochemicals. Curr. Pharmacol. Rep. 2020, 6, 325–334. [Google Scholar] [CrossRef]
- Jodynis-Liebert, J.; Kujawska, M. Biphasic Dose-Response Induced by Phytochemicals: Experimental Evidence. J. Clin. Med. 2020, 9, 718. [Google Scholar] [CrossRef] [PubMed]
- Son, T.G.; Camandola, S.; Mattson, M.P. Hormetic dietary phytochemicals. Neuromol. Med. 2008, 10, 236–246. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, E.J.; Mattson, M.P.; Calabrese, V. Resveratrol commonly displays hormesis: Occurrence and biomedical significance. Hum. Exp. Toxicol. 2010, 29, 980–1015. [Google Scholar] [CrossRef] [PubMed]
- Anand, U.; Jones, B.; Korchev, Y.; Bloom, S.R.; Pacchetti, B.; Anand, P.; Sodergren, M.H. CBD Effects on TRPV1 Signaling Pathways in Cultured DRG Neurons. J. Pain. Res. 2020, 13, 2269–2278. [Google Scholar] [CrossRef] [PubMed]
- Gustafsson, K.; Wang, X.; Severa, D.; Eriksson, M.; Kimby, E.; Merup, M.; Christensson, B.; Flygare, J.; Sander, B. Expression of cannabinoid receptors type 1 and type 2 in non-Hodgkin lymphoma: Growth inhibition by receptor activation. Int. J. Cancer 2008, 123, 1025–1033. [Google Scholar] [CrossRef] [PubMed]
- Derocq, J.M.; Segui, M.; Marchand, J.; Le Fur, G.; Casellas, P. Cannabinoids enhance human B-cell growth at low nanomolar concentrations. FEBS Lett. 1995, 369, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Carayon, P.; Marchand, J.; Dussossoy, D.; Derocq, J.M.; Jbilo, O.; Bord, A.; Bouaboula, M.; Galiegue, S.; Mondiere, P.; Penarier, G.; et al. Modulation and functional involvement of CB2 peripheral cannabinoid receptors during B-cell differentiation. Blood 1998, 92, 3605–3615. [Google Scholar] [CrossRef] [PubMed]
- Marcu, J.P.; Christian, R.T.; Lau, D.; Zielinski, A.J.; Horowitz, M.P.; Lee, J.; Pakdel, A.; Allison, J.; Limbad, C.; Moore, D.H.; et al. Cannabidiol enhances the inhibitory effects of delta9-tetrahydrocannabinol on human glioblastoma cell proliferation and survival. Mol. Cancer Ther. 2010, 9, 180–189. [Google Scholar] [CrossRef] [PubMed]
- Massi, P.; Vaccani, A.; Bianchessi, S.; Costa, B.; Macchi, P.; Parolaro, D. The non-psychoactive cannabidiol triggers caspase activation and oxidative stress in human glioma cells. Cell. Mol. Life Sci. 2006, 63, 2057–2066. [Google Scholar] [CrossRef]
- Wójcik, P.; Biernacki, M.; Domian, N.; Žarković, N.; Skrzydlewska, E. Influence of Inhibition of COX-2-Dependent Lipid Metabolism on Regulation of UVB-Induced Keratinocytes Apoptosis by Cannabinoids. Biomolecules 2022, 12, 842. [Google Scholar] [CrossRef]
- Meijerink, J.; Poland, M.; Balvers, M.G.J.; Plastina, P.; Lute, C.; Dwarkasing, J.; van Norren, K.; Witkamp, R.F. Inhibition of COX-2-mediated eicosanoid production plays a major role in the anti-inflammatory effects of the endocannabinoid N-docosahexaenoylethanolamine (DHEA) in macrophages. Br. J. Pharmacol. 2015, 172, 24–37. [Google Scholar] [CrossRef] [PubMed]
- Ruhaak, L.R.; Felth, J.; Karlsson, P.C.; Rafter, J.J.; Verpoorte, R.; Bohlin, L. Evaluation of the cyclooxygenase inhibiting effects of six major cannabinoids isolated from Cannabis sativa. Biol. Pharm. Bull. 2011, 34, 774–778. [Google Scholar] [CrossRef] [PubMed]
- Rimmerman, N.; Ben-Hail, D.; Porat, Z.; Juknat, A.; Kozela, E.; Daniels, M.P.; Connelly, P.S.; Leishman, E.; Bradshaw, H.B.; Shoshan-Barmatz, V.; et al. Direct modulation of the outer mitochondrial membrane channel, voltage-dependent anion channel 1 (VDAC1) by cannabidiol: A novel mechanism for cannabinoid-induced cell death. Cell Death Dis. 2013, 4, e949. [Google Scholar] [CrossRef] [PubMed]
- Singer, E.; Judkins, J.; Salomonis, N.; Matlaf, L.; Soteropoulos, P.; McAllister, S.; Soroceanu, L. Reactive oxygen species-mediated therapeutic response and resistance in glioblastoma. Cell Death Dis. 2015, 6, e1601. [Google Scholar] [CrossRef] [PubMed]
- Velasco, G.; Sanchez, C.; Guzman, M. Towards the use of cannabinoids as antitumour agents. Nat. Rev. Cancer 2012, 12, 436–444. [Google Scholar] [CrossRef] [PubMed]
- Gustafsson, K.; Christensson, B.; Sander, B.; Flygare, J. Cannabinoid receptor-mediated apoptosis induced by R(+)-methanandamide and Win55,212-2 is associated with ceramide accumulation and p38 activation in mantle cell lymphoma. Mol. Pharmacol. 2006, 70, 1612–1620. [Google Scholar] [CrossRef]
- Grotenhermen, F. Pharmacokinetics and pharmacodynamics of cannabinoids. Clin. Pharmacokinet. 2003, 42, 327–360. [Google Scholar] [CrossRef]
- Britch, S.C.; Babalonis, S.; Walsh, S.L. Cannabidiol: Pharmacology and therapeutic targets. Psychopharmacology 2021, 238, 9–28. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Line | Cannabinoid | IC50 | SE * | R2 † |
|---|---|---|---|---|
| Canine B-cell lymphoma 1771 | CBD | 14 µM | 2 | 0.9 |
| THC | 41 µM | 9.6 | 0.9 | |
| WIN | NA | NA | NA | |
| Canine B-cell lymphoma CLBL-1 | CBD | 18 µM | 6.1 | 0.6 |
| THC | 34 µM | 2.2 | 0.9 | |
| WIN | 5.6 µM | 0.6 | 0.9 | |
| Canine T-cell lymphoma CL-1 | CBD | 39 µM | 3.2 | 0.8 |
| THC | NA | NA | NA | |
| WIN | NA | NA | NA | |
| Human B-cell lymphoma Ramos | CBD | 8.6 µM | 1.3 | 0.9 |
| THC | 21 µM | 2.6 | 0.8 | |
| WIN | 14 µM | 1.7 | 0.9 |
| Cellular Response to Cytotoxicity | Lymphoma Cells’ Response to Cannabinoid-Induced Cytotoxicity | |||
|---|---|---|---|---|
| Overall Effect | CBD | THC | WIN | |
| Markers of Oxidative Stress | ||||
| ROS |  | | | |
| GSH |  | | | |
| NADH | | |  | |
| H2O2 | | ND | ND | |
| Nitrite | | | | |
| Lipid peroxidation (LP) | | | | ND |
| Markers of Inflammation | ||||
| ICE-1 | | | | |
| COX | | | | |
| Markers of Apoptosis | ||||
| Caspase-3 | | | | |
| Caspase-8 | | | | |
| Caspase-9 | | | | |
| Markers of Mitochondrial function | ||||
| Complex-1 | | | | |
= Increased, = Decreased, = No significant effect, ND = Not determined].Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Omer, S.; Pathak, S.; Mansour, M.; Nadar, R.; Bowen, D.; Dhanasekaran, M.; Pondugula, S.R.; Boothe, D. Effects of Cannabidiol, ∆9-Tetrahydrocannabinol, and WIN 55-212-22 on the Viability of Canine and Human Non-Hodgkin Lymphoma Cell Lines. Biomolecules 2024, 14, 495. https://doi.org/10.3390/biom14040495
Omer S, Pathak S, Mansour M, Nadar R, Bowen D, Dhanasekaran M, Pondugula SR, Boothe D. Effects of Cannabidiol, ∆9-Tetrahydrocannabinol, and WIN 55-212-22 on the Viability of Canine and Human Non-Hodgkin Lymphoma Cell Lines. Biomolecules. 2024; 14(4):495. https://doi.org/10.3390/biom14040495
Chicago/Turabian StyleOmer, Saba, Suhrud Pathak, Mahmoud Mansour, Rishi Nadar, Dylan Bowen, Muralikrishnan Dhanasekaran, Satyanarayana R. Pondugula, and Dawn Boothe. 2024. "Effects of Cannabidiol, ∆9-Tetrahydrocannabinol, and WIN 55-212-22 on the Viability of Canine and Human Non-Hodgkin Lymphoma Cell Lines" Biomolecules 14, no. 4: 495. https://doi.org/10.3390/biom14040495