
The Role of Tenascin-C on the Structural Plasticity of Perineuronal Nets and Synaptic Expression in the Hippocampus of Male Mice

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Model
2.2. Enriched Environment
2.3. Immunohistochemistry
2.4. Image Acquisition
2.5. Image Analysis
2.5.1. Quantification of WFA and PV Signals
2.5.2. Quantification of Density, Size and Intensity of Synaptic Puncta
2.5.3. Analysis of PNN Ultrastructure
2.6. Statistical Analysis
3. Results
3.1. TnC Shapes the Ulstrastructure of PNNs and Remodels the Excitatory and Inhibitory Puncta in DG
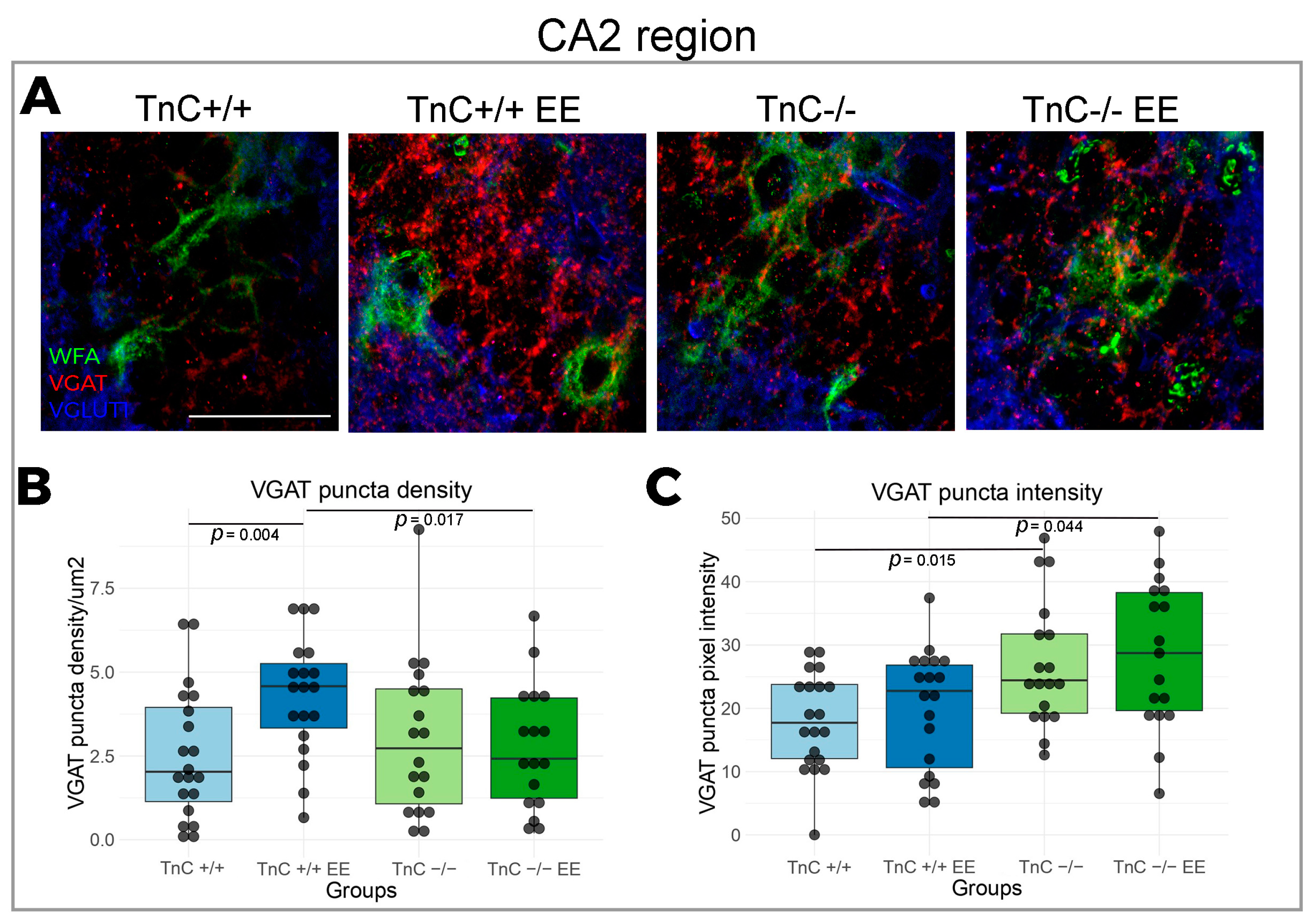
3.2. TnC Regulates the Number of PNNs and the Inhibitory Input in the CA2 Region
4. Discussion
4.1. Role of TnC in the Plasticity of the Dentate Gyrus
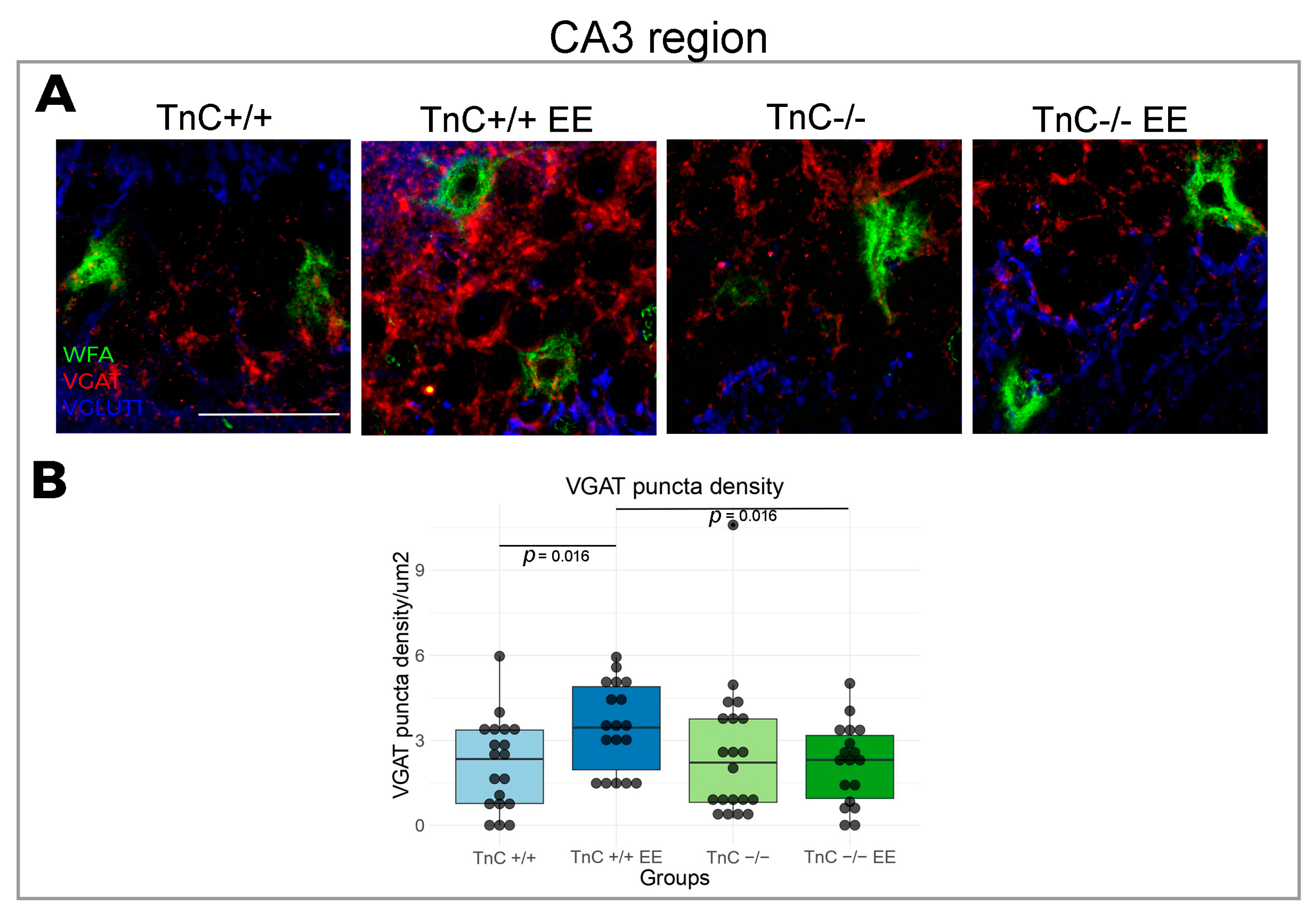
4.2. Role of TnC in the Plasticity of CA1-3 Regions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Celio, M.R.; Spreafico, R.; De Biasi, S.; Vitellaro-Zuccarello, L. Perineuronal Nets: Past and Present. Trends Neurosci. 1998, 21, 510–515. [Google Scholar] [CrossRef]
- Ruoslahti, E. Brain Extracellular Matrix. Glycobiology 1996, 6, 489–492. [Google Scholar] [CrossRef] [PubMed]
- Carstens, K.E.; Phillips, M.L.; Pozzo-Miller, L.; Weinberg, R.J.; Dudek, S.M. Perineuronal Nets Suppress Plasticity of Excitatory Synapses on CA2 Pyramidal Neurons. J. Neurosci. 2016, 36, 6312–6320. [Google Scholar] [CrossRef] [PubMed]
- Morikawa, S.; Ikegaya, Y.; Narita, M.; Tamura, H. Activation of Perineuronal Net-Expressing Excitatory Neurons during Associative Memory Encoding and Retrieval. Sci. Rep. 2017, 7, srep46024. [Google Scholar] [CrossRef] [PubMed]
- Bignami, A.; Hosley, M.; Dahl, D. Hyaluronic Acid and Hyaluronic Acid-Binding Proteins in Brain Extracellular Matrix. Anat. Embryol. 1993, 188, 419–433. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, Y. Lecticans: Organizers of the Brain Extracellular Matrix. Cell. Mol. Life Sci. 2000, 57, 276–289. [Google Scholar] [CrossRef] [PubMed]
- Celio, M.R.; Chiquet-Ehrismann, R. “Perineuronal Nets” around Cortical Interneurons Expressing Parvalbumin Are Rich in Tenascin. Neurosci. Lett. 1993, 162, 137–140. [Google Scholar] [CrossRef] [PubMed]
- Eill, G.J.; Sinha, A.; Morawski, M.; Viapiano, M.S.; Matthews, R.T. The Protein Tyrosine Phosphatase RPTPξ/Phosphacan Is Critical for Perineuronal Net Structure. J. Biol. Chem. 2020, 295, 955–968. [Google Scholar] [CrossRef] [PubMed]
- Stamenkovic, V.; Stamenkovic, S.; Jaworski, T.; Gawlak, M.; Jovanovic, M.; Jakovcevski, I.; Wilczynski, G.M.; Kaczmarek, L.; Schachner, M.; Radenovic, L.; et al. The Extracellular Matrix Glycoprotein Tenascin-C and Matrix Metalloproteinases Modify Cerebellar Structural Plasticity by Exposure to an Enriched Environment. Brain Struct. Funct. 2017, 222, 393–415. [Google Scholar] [CrossRef] [PubMed]
- Lensjø, K.K.; Christensen, A.C.; Tennøe, S.; Fyhn, M.; Hafting, T. Differential Expression and Cell-Type Specificity of Perineuronal Nets in Hippocampus, Medial Entorhinal Cortex, and Visual Cortex Examined in the Rat and Mouse. Eneuro 2017, 4. [Google Scholar] [CrossRef]
- Guadagno, A.; Verlezza, S.; Long, H.; Wong, T.P.; Walker, C.-D. It Is All in the Right Amygdala: Increased Synaptic Plasticity and Perineuronal Nets in Male, but Not Female, Juvenile Rat Pups after Exposure to Early-Life Stress. J. Neurosci. 2020, 40, 8276–8291. [Google Scholar] [CrossRef] [PubMed]
- Carulli, D.; Rhodes, K.E.; Brown, D.J.; Bonnert, T.P.; Pollack, S.J.; Oliver, K.; Strata, P.; Fawcett, J.W. Composition of Perineuronal Nets in the Adult Rat Cerebellum and the Cellular Origin of Their Components. J. Comp. Neurol. 2006, 494, 559–577. [Google Scholar] [CrossRef] [PubMed]
- Lensjø, K.K.; Lepperød, M.E.; Dick, G.; Hafting, T.; Fyhn, M. Removal of Perineuronal Nets Unlocks Juvenile Plasticity through Network Mechanisms of Decreased Inhibition and Increased Gamma Activity. J. Neurosci. 2017, 37, 1269–1283. [Google Scholar] [CrossRef] [PubMed]
- Yamada, J.; Ohgomori, T.; Jinno, S. Perineuronal Nets Affect Parvalbumin Expression in GABAergic Neurons of the Mouse Hippocampus. Eur. J. Neurosci. 2015, 41, 368–378. [Google Scholar] [CrossRef] [PubMed]
- Tzilivaki, A.; Tukker, J.J.; Maier, N.; Poirazi, P.; Sammons, R.P.; Schmitz, D. Hippocampal GABAergic Interneurons and Memory. Neuron 2023, 111, 3154–3175. [Google Scholar] [CrossRef] [PubMed]
- Dityatev, A.; Brückner, G.; Dityateva, G.; Grosche, J.; Kleene, R.; Schachner, M.; Bru, G.; Dityateva, G.; Grosche, J.; Kleene, R.; et al. Activity-Dependent Formation and Functions of Chondroitin Sulfate-Rich Extracellular Matrix of Perineuronal Nets. Dev. Neurobiol. 2007, 67, 570–588. [Google Scholar] [CrossRef]
- Balmer, T.S.; Carels, V.M.; Frisch, J.L.; Nick, T.A. Modulation of Perineuronal Nets and Parvalbumin with Developmental Song Learning. J. Neurosci. 2009, 29, 12878–12885. [Google Scholar] [CrossRef] [PubMed]
- Brückner, G.; Brauer, K.; Härtig, W.; Wolff, J.R.; Rickmann, M.J.; Derouiche, A.; Delpech, B.; Girard, N.; Oertel, W.H.; Reichenbach, A.; et al. Perineuronal Nets Provide a Polyanionic, Glia-associated Form of Microenvironment around Certain Neurons in Many Parts of the Rat Brain. Glia 1993, 8, 183–200. [Google Scholar] [CrossRef] [PubMed]
- Tewari, B.P.; Chaunsali, L.; Campbell, S.L.; Patel, D.C.; Goode, A.E.; Sontheimer, H. Perineuronal Nets Decrease Membrane Capacitance of Peritumoral Fast Spiking Interneurons in a Model of Epilepsy. Nat. Commun. 2018, 9, 4724. [Google Scholar] [CrossRef] [PubMed]
- Pizzorusso, T. Reactivation of Ocular Dominance Plasticity in the Adult Visual Cortex. Science 2002, 298, 1248–1251. [Google Scholar] [CrossRef] [PubMed]
- Suttkus, A.; Morawski, M.; Arendt, T. Protective Properties of Neural Extracellular Matrix. Mol. Neurobiol. 2016, 53, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Carceller, H.; Guirado, R.; Ripolles-Campos, E.; Teruel-Marti, V.; Nacher, J. Perineuronal Nets Regulate the Inhibitory Perisomatic Input onto Parvalbumin Interneurons and γ Activity in the Prefrontal Cortex. J. Neurosci. 2020, 40, 5008–5018. [Google Scholar] [CrossRef] [PubMed]
- Jakovljevic, A.; Agatonovic, G.; Aleksic, D.; Aksic, M.; Reiss, G.; Förster, E.; Stamatakis, A.; Jakovcevski, I.; Poleksic, J. The Impact of Early Life Maternal Deprivation on the Perineuronal Nets in the Prefrontal Cortex and Hippocampus of Young Adult Rats. Front. Cell Dev. Biol. 2022, 10, 982663. [Google Scholar] [CrossRef] [PubMed]
- Mauney, S.A.; Athanas, K.M.; Pantazopoulos, H.; Shaskan, N.; Passeri, E.; Berretta, S.; Woo, T.W. Developmental Pattern of Perineuronal Nets in the in Schizophrenia. Biol. Psychiatry 2013, 74, 427–435. [Google Scholar] [CrossRef] [PubMed]
- Crapser, J.D.; Spangenberg, E.E.; Barahona, R.A.; Arreola, M.A.; Hohsfield, L.A.; Green, K.N. Microglia Facilitate Loss of Perineuronal Nets in the Alzheimer’s Disease Brain. eBioMedicine 2020, 58, 102919. [Google Scholar] [CrossRef] [PubMed]
- Dzyubenko, E.; Manrique-Castano, D.; Kleinschnitz, C.; Faissner, A.; Hermann, D.M. Topological Remodeling of Cortical Perineuronal Nets in Focal Cerebral Ischemia and Mild Hypoperfusion. Matrix Biol. 2018, 74, 121–132. [Google Scholar] [CrossRef] [PubMed]
- Härtig, W.; Mages, B.; Aleithe, S.; Nitzsche, B.; Altmann, S.; Barthel, H.; Krueger, M.; Michalski, D. Damaged Neocortical Perineuronal Nets Due to Experimental Focal Cerebral Ischemia in Mice, Rats and Sheep. Front. Integr. Neurosci. 2017, 11, 15. [Google Scholar] [CrossRef] [PubMed]
- Sigal, Y.M.; Bae, H.; Bogart, L.J.; Hensch, T.K.; Zhuang, X. Structural Maturation of Cortical Perineuronal Nets and Their Perforating Synapses Revealed by Superresolution Imaging. Proc. Natl. Acad. Sci. USA 2019, 116, 7071–7076. [Google Scholar] [CrossRef] [PubMed]
- Fawcett, J.W.; Fyhn, M.; Jendelova, P.; Kwok, J.C.F.; Ruzicka, J.; Sorg, B.A. The Extracellular Matrix and Perineuronal Nets in Memory. Mol. Psychiatry 2022, 27, 3192–3203. [Google Scholar] [CrossRef] [PubMed]
- Nithianantharajah, J.; Hannan, A.J. Enriched Environments, Experience-Dependent Plasticity and Disorders of the Nervous System. Nat. Rev. Neurosci. 2006, 7, 697–709. [Google Scholar] [CrossRef] [PubMed]
- Foscarin, S.; Ponchione, D.; Pajaj, E.; Leto, K.; Gawlak, M.; Wilczynski, G.M.; Rossi, F.; Carulli, D. Experience-Dependent Plasticity and Modulation of Growth Regulatory Molecules at Central Synapses. PLoS ONE 2011, 6, e16666. [Google Scholar] [CrossRef]
- Madinier, A.; Quattromani, M.J.; Sjölund, C.; Ruscher, K.; Wieloch, T. Enriched Housing Enhances Recovery of Limb Placement Ability and Reduces Aggrecan-Containing Perineuronal Nets in the Rat Somatosensory Cortex after Experimental Stroke. PLoS ONE 2014, 9, e93121. [Google Scholar] [CrossRef] [PubMed]
- Trévia, N.; Almeida, I.N.F.; Oliveira, P.S.; Warwick, L.V.; Marques, V.; dos Santos, D.C.; Paulo, D.C.; Viegas, M.L.; Picanço-Diniz, C.W. Enriched Environment Contributes to Recovery of Visual Acuity and Increases Perineuronal Nets in Monocular-Deprived Animals. Psychol. Neurosci. 2011, 4, 49–56. [Google Scholar] [CrossRef]
- Cattaud, V.; Bezzina, C.; Rey, C.C.; Lejards, C.; Dahan, L.; Verret, L. Early Disruption of Parvalbumin Expression and Perineuronal Nets in the Hippocampus of the Tg2576 Mouse Model of Alzheimer’s Disease Can Be Rescued by Enriched Environment. Neurobiol. Aging 2018, 72, 147–158. [Google Scholar] [CrossRef] [PubMed]
- Midwood, K.S.; Chiquet, M.; Tucker, R.P.; Orend, G. Tenascin-C at a Glance. J. Cell Sci. 2016, 129, 4321–4327. [Google Scholar] [CrossRef] [PubMed]
- Šekeljić, V.; Andjus, P.R. Tenascin-C and Its Functions in Neuronal Plasticity. Int. J. Biochem. Cell Biol. 2012, 44, 825–829. [Google Scholar] [CrossRef] [PubMed]
- Ferhat, L.; Louis, N.C.A.; Jorquera, I.; Niquet, J.; Khrestchatisky, M.; Ben-Ari, Y.; Represa, A. Transient Increase of Tenascin-C in Immature Hippocampus: Astroglial and Neuronal Expression. J. Neurocytol. 1996, 25, 53–66. [Google Scholar] [CrossRef] [PubMed]
- Irintchev, A.; Rollenhagen, A.; Troncoso, E.; Kiss, J.Z.; Schachner, M. Structural and Functional Aberrations in the Cerebral Cortex of Tenascin-C Deficient Mice. Cereb. Cortex 2005, 15, 950–962. [Google Scholar] [CrossRef] [PubMed]
- Tucić, M.; Stamenković, V.; Andjus, P. The Extracellular Matrix Glycoprotein Tenascin C and Adult Neurogenesis. Front. Cell Dev. Biol. 2021, 9, 674199. [Google Scholar] [CrossRef] [PubMed]
- Yuasa, S. Development of Astrocytes in the Mouse Hippocampus as Tracked by Tenascin-C Gene Expression. Arch. Histol. Cytol. 2001, 64, 149–158. [Google Scholar] [CrossRef] [PubMed]
- Faissner, A.; Roll, L.; Theocharidis, U. Tenascin-C in the Matrisome of Neural Stem and Progenitor Cells. Mol. Cell. Neurosci. 2017, 81, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Rigato, F.; Garwood, J.; Calco, V.; Heck, N.; Faivre-Sarrailh, C.; Faissner, A. Tenascin-C Promotes Neurite Outgrowth of Embryonic Hippocampal Neurons through the Alternatively Spliced Fibronectin Type III BD Domains via Activation of the Cell Adhesion Molecule F3/Contactin. J. Neurosci. 2002, 22, 6596–6609. [Google Scholar] [CrossRef] [PubMed]
- Bijelić, D.; Adžić, M.; Perić, M.; Jakovčevski, I.; Förster, E.; Schachner, M.; Andjus, P.R. Different Functions of Recombinantly Expressed Domains of Tenascin-C in Glial Scar Formation. Front. Immunol. 2021, 11, 624612. [Google Scholar] [CrossRef] [PubMed]
- Bijelić, D.; Adžić, M.; Perić, M.; Reiss, G.; Milošević, M.; Andjus, P.R.; Jakovčevski, I. Tenascin-C Fibronectin D Domain Is Involved in the Fine-Tuning of Glial Response to CNS Injury in Vitro. Front. Cell Dev. Biol. 2022, 10, 952208. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Lee, H.J.; Jakovcevski, I.; Shah, R.; Bhagat, N.; Loers, G.; Liu, H.-Y.; Meiners, S.; Taschenberger, G.; Kügler, S.; et al. The Extracellular Matrix Glycoprotein Tenascin-C Is Beneficial for Spinal Cord Regeneration. Mol. Ther. 2010, 18, 1769–1777. [Google Scholar] [CrossRef] [PubMed]
- Dzyubenko, E.; Manrique-Castano, D.; Pillath-Eilers, M.; Vasileiadou, P.; Reinhard, J.; Faissner, A.; Hermann, D.M. Tenascin-C Restricts Reactive Astrogliosis in the Ischemic Brain. Matrix Biol. 2022, 110, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Manrique-Castano, D.; Dzyubenko, E.; Borbor, M.; Vasileiadou, P.; Kleinschnitz, C.; Roll, L.; Faissner, A.; Hermann, D.M. Tenascin-C Preserves Microglia Surveillance and Restricts Leukocyte and, More Specifically, T Cell Infiltration of the Ischemic Brain. Brain. Behav. Immun. 2021, 91, 639–648. [Google Scholar] [CrossRef] [PubMed]
- Momčilović, M.; Stamenković, V.; Jovanović, M.; Andjus, P.R.; Jakovčevski, I.; Schachner, M.; Miljković, Đ. Tenascin-C Deficiency Protects Mice from Experimental Autoimmune Encephalomyelitis. J. Neuroimmunol. 2017, 302, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Metzger, M.; Bartsch, S.; Bartsch, U.; Bock, J.; Schachner, M.; Braun, K. Regional and Cellular Distribution of the Extracellular Matrix Protein Tenascin-C in the Chick Forebrain and Its Role in Neonatal Learning. Neuroscience 2006, 141, 1709–1719. [Google Scholar] [CrossRef] [PubMed]
- Bartsch, S.; Bartsch, U.; Dijrries, U.; Ekblom, P. Expression Cortex of Tenascin in the Developing and Adult Cerebellar. J. Neurosci. 1992, 12(3), 736–749. [Google Scholar] [CrossRef] [PubMed]
- Theodosis, D.T.; Pierre, K.; Cadoret, M.A.; Allard, M.; Faissner, A.; Poulain, D.A. Expression of High Levels of the Extracellular Matrix Glycoprotein, Tenascin-C, in the Normal Adult Hypothalamoneurohypophysial System. J. Comp. Neurol. 1997, 379, 386–398. [Google Scholar] [CrossRef]
- Evers, M.R.; Salmen, B.; Bukalo, O.; Rollenhagen, A.; Bösl, M.R.; Morellini, F.; Bartsch, U.; Dityatev, A.; Schachner, M. Impairment of L-Type Ca2+ Channel-Dependent Forms of Hippocampal Synaptic Plasticity in Mice Deficient in the Extracellular Matrix Glycoprotein Tenascin-C. J. Neurosci. 2002, 22, 7177–7194. [Google Scholar] [CrossRef] [PubMed]
- Stamenkovic, V.; Milenkovic, I.; Galjak, N.; Todorovic, V.; Andjus, P. Enriched Environment Alters the Behavioral Profile of Tenascin-C Deficient Mice. Behav. Brain Res. 2017, 331, 241–253. [Google Scholar] [CrossRef] [PubMed]
- Morellini, F.; Schachner, M. Enhanced Novelty-Induced Activity, Reduced Anxiety, Delayed Resynchronization to Daylight Reversal and Weaker Muscle Strength in Tenascin-C-Deficient Mice. Eur. J. Neurosci. 2006, 23, 1255–1268. [Google Scholar] [CrossRef] [PubMed]
- Morellini, F.; Malyshev, A.; Volgushev, M.; Chistiakova, M.; Papashvili, G.; Fellini, L.; Kleene, R.; Schachner, M.; Dityatev, A. Impaired Fear Extinction Due to a Deficit in Ca2+ Influx Through L-Type Voltage-Gated Ca2+ Channels in Mice Deficient for Tenascin-C. Front. Integr. Neurosci. 2017, 11, 16. [Google Scholar] [CrossRef] [PubMed]
- Andjus, P.R.; Bajić, A.; Zhu, L.; Schachner, M.; Strata, P. Short-Term Facilitation and Depression in the Cerebellum: Some Observations on Wild-Type and Mutant Rodents Deficient in the Extracellular Matrix Molecule Tenascin C. Ann. N. Y. Acad. Sci. 2005, 1048, 185–197. [Google Scholar] [CrossRef] [PubMed]
- Gottschling, C.; Wegrzyn, D.; Denecke, B.; Faissner, A. Elimination of the Four Extracellular Matrix Molecules Tenascin-C, Tenascin-R, Brevican and Neurocan Alters the Ratio of Excitatory and Inhibitory Synapses. Sci. Rep. 2019, 9, 13939. [Google Scholar] [CrossRef]
- Gurevicius, K.; Kuang, F.; Stoenica, L.; Irintchev, A.; Gureviciene, I.; Dityatev, A.; Schachner, M.; Tanila, H. Genetic Ablation of Tenascin-C Expression Leads to Abnormal Hippocampal CA1 Structure and Electrical Activity in Vivo. Hippocampus 2009, 19, 1232–1246. [Google Scholar] [CrossRef] [PubMed]
- Jansen, S.; Gottschling, C.; Faissner, A.; Manahan-Vaughan, D. Intrinsic Cellular and Molecular Properties of In Vivo Hippocampal Synaptic Plasticity Are Altered in the Absence of Key Synaptic Matrix Molecules. Hippocampus 2017, 27, 920–933. [Google Scholar] [CrossRef] [PubMed]
- Mitrovic, N.; Schachner, M. Detection of Tenascin-C in the Nervous System of the Tenascin-C Mutant Mouse. J. Neurosci. Res. 1995, 42, 710–717. [Google Scholar] [CrossRef] [PubMed]
- Nakic, M.; Manahan-Vaughan, D.; Reymann, K.G.; Schachner, M. Long-Term Potentiation In Vivo Increases Rat Hippocampal Tenascin-C Expression. J. Neurobiol. 1998, 37, 393–404. [Google Scholar] [CrossRef]
- Strekalova, T.; Sun, M.; Sibbe, M.; Evers, M.; Dityatev, A.; Gass, P.; Schachner, M. Fibronectin Domains of Extracellular Matrix Molecule Tenascin-C Modulate Hippocampal Learning and Synaptic Plasticity. Mol. Cell. Neurosci. 2002, 21, 173–187. [Google Scholar] [CrossRef] [PubMed]
- Day, J.M.; Olin, A.I.; Murdoch, A.D.; Canfield, A.; Sasaki, T.; Timpl, R.; Hardingham, T.E.; Aspberg, A. Alternative Splicing in the Aggrecan G3 Domain Influences Binding Interactions with Tenascin-C and Other Extracellular Matrix Proteins. J. Biol. Chem. 2004, 279, 12511–12518. [Google Scholar] [CrossRef] [PubMed]
- Grumet, M.; Milev, P.; Sakurai, T.; Karthikeyan, L.; Bourdon, M.; Margolis, R.K.; Margolis, R.U. Interactions with Tenascin and Differential Effects on Cell Adhesion of Neurocan and Phosphacan, Two Major Chondroitin Sulfate Proteoglycans of Nervous Tissue. J. Biol. Chem. 1994, 269, 12142–12146. [Google Scholar] [CrossRef] [PubMed]
- Milev, P.; Fischer, D.; Häring, M.; Schulthess, T.; Margolis, R.K.; Chiquet-Ehrismann, R.; Margolis, R.U. The Fibrinogen-like Globe of Tenascin-C Mediates Its Interactions with Neurocan and Phosphacan/Protein-Tyrosine Phosphatase-ζ/β. J. Biol. Chem. 1997, 272, 15501–15509. [Google Scholar] [CrossRef] [PubMed]
- Grigoryan, G.A. Molecular-Cellular Mechanisms of Plastic Restructuring Produced by an Enriched Environment. Eff. Learn. Mem. 2021, 15, 226–239. [Google Scholar] [CrossRef]
- Ippolito, D.M.; Eroglu, C. Quantifying Synapses: An Immunocytochemistry-Based Assay to Quantify Synapse Number. J. Vis. Exp. 2010, 45, e2270. [Google Scholar] [CrossRef]
- Paxinos, G.; Franklin, K.B.J. The Mouse Brain in Stereotaxic Coordinates, 2nd ed.; Academic Press: Cambridge, MA, USA, 2001; 360p. [Google Scholar]
- Murthy, S.; Kane, G.A.; Katchur, N.J.; Lara Mejia, P.S.; Obiofuma, G.; Buschman, T.J.; McEwen, B.S.; Gould, E. Perineuronal Nets, Inhibitory Interneurons, and Anxiety-Related Ventral Hippocampal Neuronal Oscillations Are Altered by Early Life Adversity. Biol. Psychiatry 2019, 85, 1011–1020. [Google Scholar] [CrossRef]
- Reichelt, A.C.; Hare, D.J.; Bussey, T.J.; Saksida, L.M. Perineuronal Nets: Plasticity, Protection, and Therapeutic Potential. Trends Neurosci. 2019, 42, 458–470. [Google Scholar] [CrossRef] [PubMed]
- van Praag, H.; Kempermann, G.; Gage, F.H. Neural Consequences of Enviromental Enrichment. Nat. Rev. Neurosci. 2000, 1, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Yamada, J.; Nadanaka, S.; Kitagawa, H.; Takeuchi, K.; Jinno, S. Increased Synthesis of Chondroitin Sulfate Proteoglycan Promotes Adult Hippocampal Neurogenesis in Response to Enriched Environment. J. Neurosci. 2018, 38, 8496–8513. [Google Scholar] [CrossRef] [PubMed]
- Booker, S.A.; Vida, I. Morphological Diversity and Connectivity of Hippocampal Interneurons. Cell Tissue Res. 2018, 373, 619–641. [Google Scholar] [CrossRef] [PubMed]
- Domínguez, S.; Rey, C.C.; Therreau, L.; Fanton, A.; Massotte, D.; Verret, L.; Piskorowski, R.A.; Chevaleyre, V. Maturation of PNN and ErbB4 Signaling in Area CA2 during Adolescence Underlies the Emergence of PV Interneuron Plasticity and Social Memory. Cell Rep. 2019, 29, 1099–1112.e4. [Google Scholar] [CrossRef] [PubMed]
- Cope, E.C.; Zych, A.D.; Katchur, N.J.; Waters, R.C.; Laham, B.J.; Diethorn, E.J.; Park, C.Y.; Meara, W.R.; Gould, E. Atypical Perineuronal Nets in the CA2 Region Interfere with Social Memory in a Mouse Model of Social Dysfunction. Mol. Psychiatry 2022, 27, 3520–3531. [Google Scholar] [CrossRef] [PubMed]
- Geissler, M.; Gottschling, C.; Aguado, A.; Rauch, U.; Wetzel, C.H.; Hatt, H.; Faissner, A. Primary Hippocampal Neurons, Which Lack Four Crucial Extracellular Matrix Molecules, Display Abnormalities of Synaptic Structure and Function and Severe Deficits in Perineuronal Net Formation. J. Neurosci. 2013, 33, 7742–7755. [Google Scholar] [CrossRef] [PubMed]
- Travaglia, A.; Steinmetz, A.B.; Miranda, J.M.; Alberini, C.M. Mechanisms of Critical Period in the Hippocampus Underlie Object Location Learning and Memory in Infant Rats. Learn. Mem. 2018, 25, 176–182. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jakovljević, A.; Stamenković, V.; Poleksić, J.; Hamad, M.I.K.; Reiss, G.; Jakovcevski, I.; Andjus, P.R. The Role of Tenascin-C on the Structural Plasticity of Perineuronal Nets and Synaptic Expression in the Hippocampus of Male Mice. Biomolecules 2024, 14, 508. https://doi.org/10.3390/biom14040508
Jakovljević A, Stamenković V, Poleksić J, Hamad MIK, Reiss G, Jakovcevski I, Andjus PR. The Role of Tenascin-C on the Structural Plasticity of Perineuronal Nets and Synaptic Expression in the Hippocampus of Male Mice. Biomolecules. 2024; 14(4):508. https://doi.org/10.3390/biom14040508
Chicago/Turabian StyleJakovljević, Ana, Vera Stamenković, Joko Poleksić, Mohammad I. K. Hamad, Gebhard Reiss, Igor Jakovcevski, and Pavle R. Andjus. 2024. "The Role of Tenascin-C on the Structural Plasticity of Perineuronal Nets and Synaptic Expression in the Hippocampus of Male Mice" Biomolecules 14, no. 4: 508. https://doi.org/10.3390/biom14040508