RNA Sequencing Provides Insights into the Regulation of Solanesol Biosynthesis in Nicotiana tabacum Induced by Moderately High Temperature

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growing Conditions
2.2. Analysis of Total Solanesol Content
2.3. Measurement of Leaf Dry Weight and Net Photosynthetic Rate
2.4. RNA Extraction and Sequencing (RNA-Seq)
2.5. RNA-Seq Quality Control and Reads Mapping to the Reference Genome
2.6. Quantification of Gene Expression Levels and Differential Gene Expression Analysis
2.7. GO and KEGG Enrichment Analysis of Differentially Expressed Genes
2.8. Quantitative Real-Time PCR
2.9. Statistical Analysis
2.10. GenBank Accession Code
3. Results
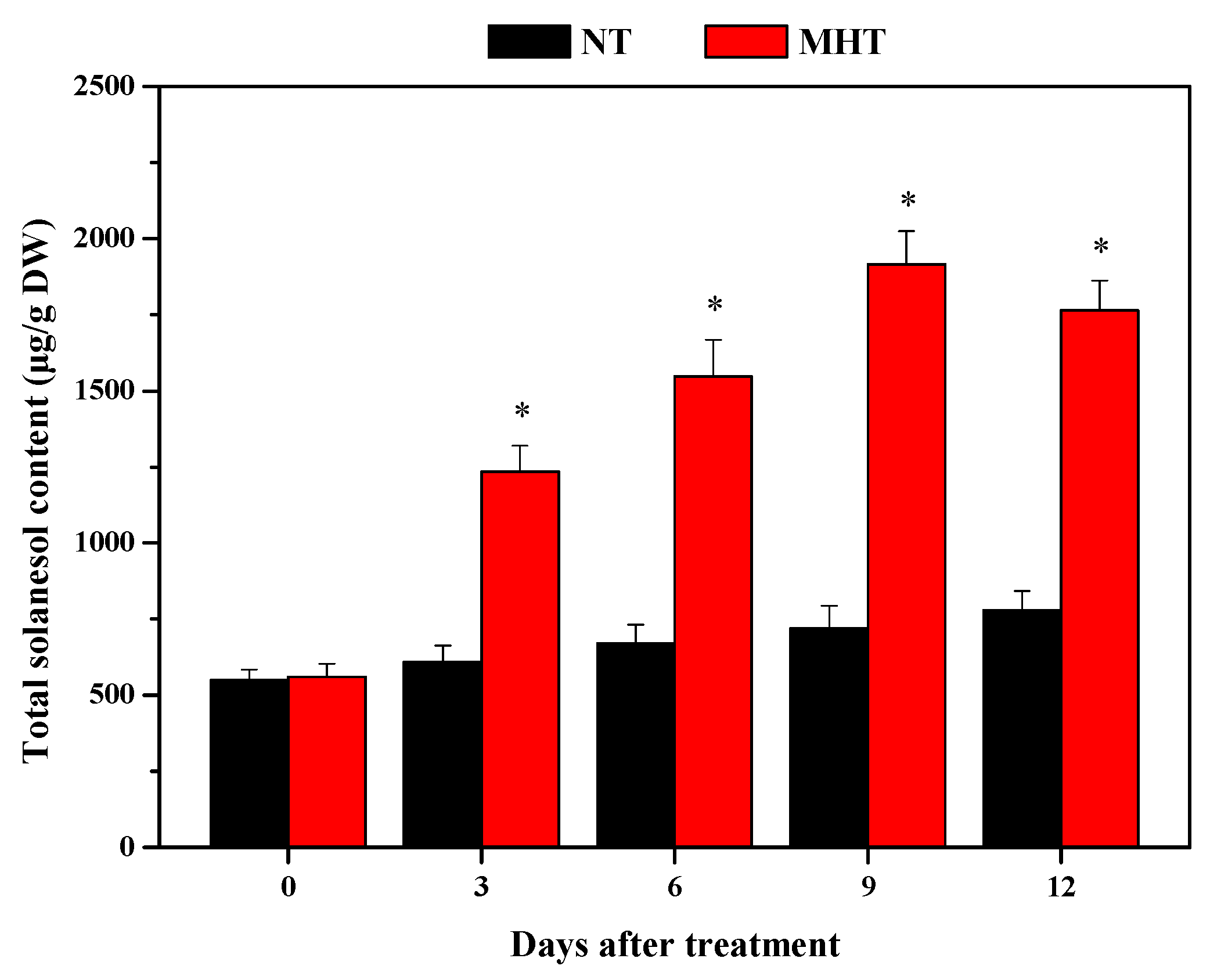
3.1. Total Solanesol Contents in Leaves of Tobacco Plants Grown at Normal Temperature and Moderately High Temperature
3.2. Leaf Dry Weight and Net Photosynthetic Rate in Leaves of Tobacco Plants Grown at Normal Temperature and Moderately High Temperature
3.3. Quality Assessment of Sequencing Data, Sequence Alignment, and Transcription Analysis of Differentially Expressed Genes
3.3.1. Quality Assessment of Sequencing Data
3.3.2. Sequence Alignment
3.3.3. Transcription Analysis of Differentially Expressed Genes
3.4. Gene Ontology Functional Enrichment Analysis of Differentially Expressed Genes
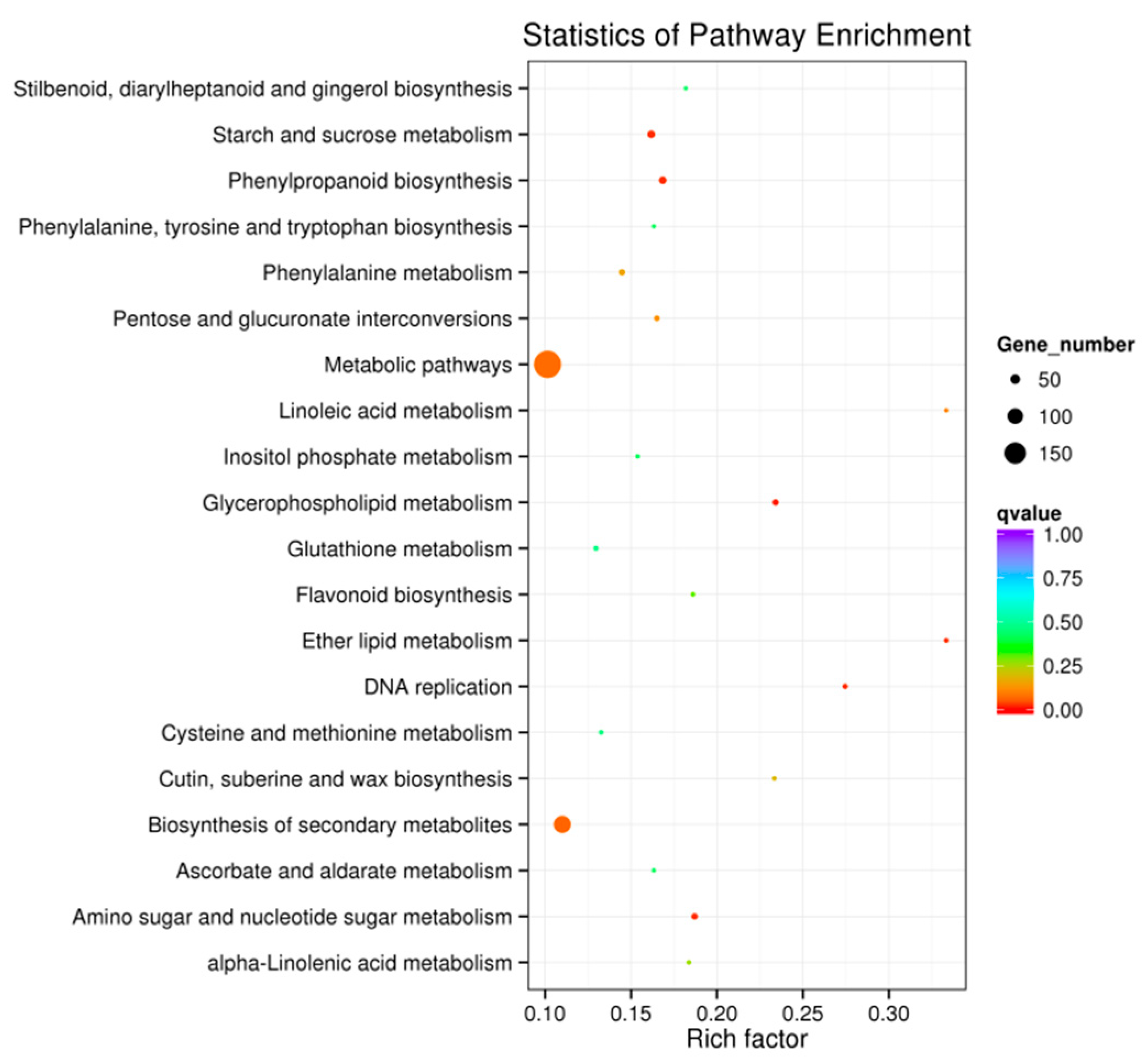
3.5. Kyoto Encyclopedia of Genes and Genomes Functional Enrichment Analysis of Differentially Expressed Genes
3.6. Transcription Factor Analysis of DEGs
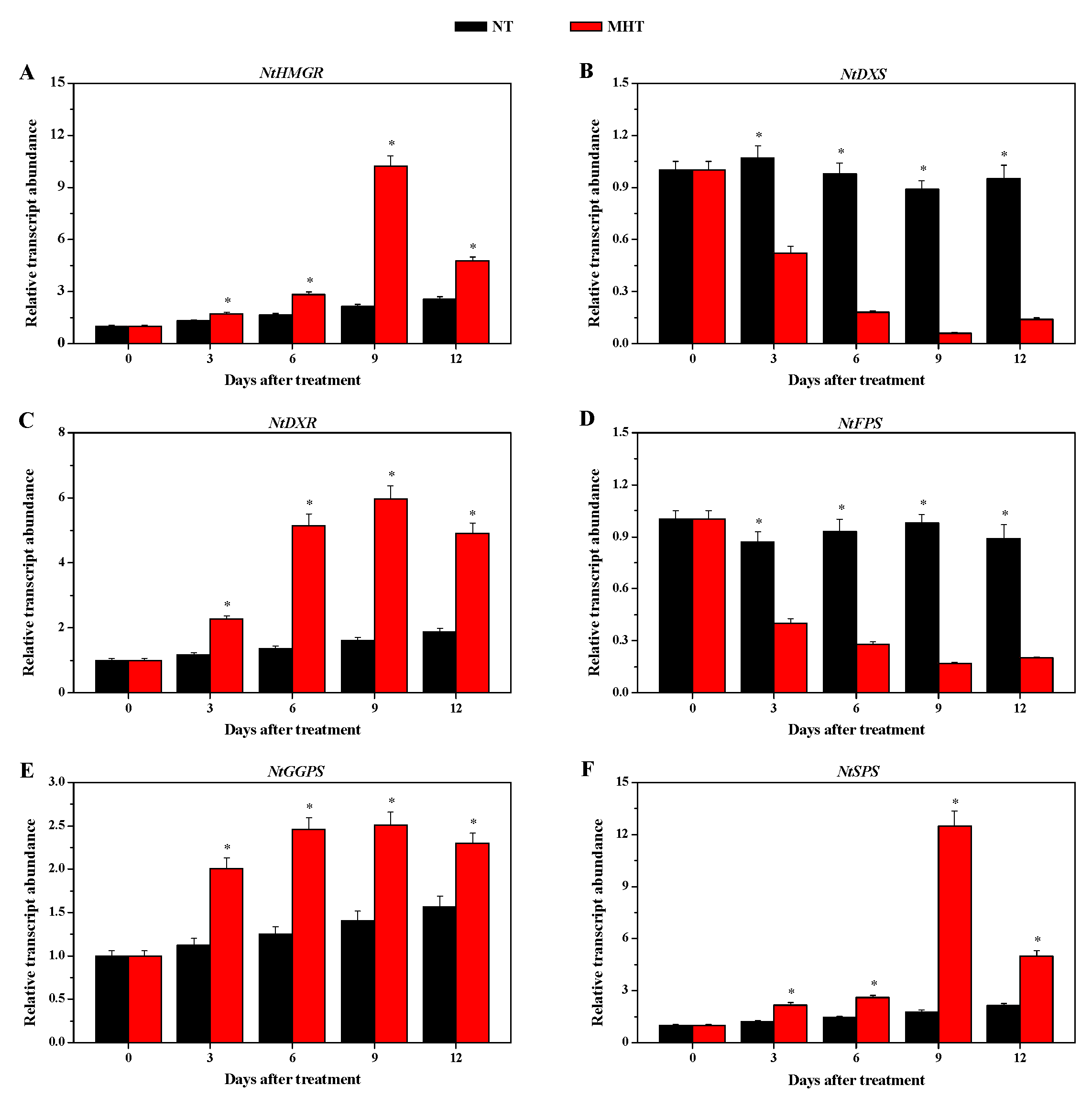
3.7. Transcription of Solanesol Biosynthesis Genes in Leaves of Tobacco Plants Grown at Normal Temperature and Moderately High Temperature
4. Discussion
4.1. MHT Enhances the Solanesol Accumulation in Tobacco Leaves
4.2. Transcriptome Analysis of Tobacco Leaves Treated with Moderately High Temperature
4.3. Transcription of Solanesol Biosynthesis Genes in Tobacco Leaves Treated with Moderately High Temperature
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Li, Y.; Li, Z.; Guo, P. Research progress on the bioactive components of Nicotiana tabacum L. Nat. Prod. Res. Develop. 2015, 27, 2157–2163. [Google Scholar]
- Yan, N.; Liu, Y.; Gong, D.; Du, Y.; Zhang, H.; Zhang, Z. Solanesol: A review of its resources, derivatives, bioactivities, medicinal applications, and biosynthesis. Phytochem. Rev. 2015, 14, 403–417. [Google Scholar] [CrossRef]
- Yan, N.; Du, Y.; Liu, X.; Zhang, H.; Liu, Y.; Zhang, P.; Gong, D.; Zhang, Z. Chemical structures, biosynthesis, bioactivities, biocatalysis and semisynthesis of tobacco cembranoids: An overview. Ind. Crop. Prod. 2016, 83, 66–80. [Google Scholar] [CrossRef]
- Taylor, M.A.; Fraser, P.D. Solanesol: Added value from Solanaceous waste. Phytochemistry 2011, 72, 1323–1327. [Google Scholar] [CrossRef] [PubMed]
- Yan, N.; Zhang, H.; Zhang, Z.; Shi, J.; Timko, M.P.; Du, Y.; Liu, X.; Liu, Y. Organ-and growing stage-specific expression of solanesol biosynthesis genes in Nicotiana tabacum reveals their association with solanesol content. Molecules 2016, 21, 1536. [Google Scholar] [CrossRef]
- Yan, N.; Liu, Y.; Zhang, H.; Du, Y.; Liu, X.; Zhang, Z. Solanesol biosynthesis in plants. Molecules 2017, 22, 510. [Google Scholar] [CrossRef] [PubMed]
- Mehan, S.; Rajput, M.; Dudi, R.; Ghimire, K. Neuroprotective strategies of solanesol in mitochondrial impairment in experimentally induced Huntington disease. J. Pharm. Toxicol. 2018, 1, 3–7. [Google Scholar]
- Yao, X.; Bai, Q.; Yan, D.; Li, G.; Lü, C.; Xu, H. Solanesol protects human hepatic L02 cells from ethanol-induced oxidative injury via upregulation of HO-1 and Hsp70. Toxicol. In Vitro 2015, 29, 600–608. [Google Scholar] [CrossRef]
- Yao, X.; Lu, B.; Lü, C.; Bai, Q.; Yan, D.; Wu, Y.; Hong, Z.; Xu, H. Solanesol induces the expression of heme oxygenase-1 via p38 and Akt and suppresses the production of proinflammatory cytokines in RAW264.7 cells. Food Funct. 2017, 8, 132–141. [Google Scholar] [CrossRef]
- Qin, B.; Liu, L.; Pan, Y.; Zhu, Y.; Wu, X.; Song, S.; Han, G. PEGylated solanesol for oral delivery of coenzyme Q10. J. Agr. Food Chem. 2017, 65, 3360–3367. [Google Scholar] [CrossRef]
- Qin, B.; Liu, L.; Wu, X.; Liang, F.; Hou, T.; Pan, Y.; Song, S. mPEGylated solanesol micelles as redox-responsive nanocarriers with synergistic anticancer effect. Acta Biomater. 2017, 64, 211–222. [Google Scholar] [CrossRef] [PubMed]
- Roe, S.J.; Oldfield, M.F.; Geach, N.; Baxter, A. A convergent stereocontrolled synthesis of [3-14C] solanesol. J. Labelled Compound. Rad. 2013, 56, 485–491. [Google Scholar] [CrossRef] [PubMed]
- Campbell, R.; Freitag, S.; Bryan, G.J.; Stewart, D.; Taylor, M.A. Environmental and genetic factors associated with solanesol accumulation in potato leaves. Front. Plant Sci. 2016, 7, 1263. [Google Scholar] [CrossRef] [PubMed]
- Xiang, D.; Yao, Z.; Liu, Y.; Gai, X.; Du, Y.; Zhang, Z.; Yan, N.; Wang, A.; Fu, Q. Analysis on solanesol content and genetic diversity of Chinese flue-cured tobacco (Nicotiana tabacum L.). Crop Sci. 2017, 57, 847–855. [Google Scholar] [CrossRef]
- Xiang, D.; Zhao, T.; Du, Y.; Zhang, Z.; Yan, N.; Huang, W.; Wang, A.; Fu, Q.; Gong, Y. Genetic analysis on solanesol content of tobacco. Chinese Tob. Sci. 2015, 36, 1–7. [Google Scholar]
- Bajda, A.; Konopka-Postupolska, D.; Krzymowska, M.; Hennig, J.; Skorupinska-Tudek, K.; Surmacz, L.; Wojcik, J.; Matysiak, Z.; Chojnacki, T.; Skorzynska-Polit, E.; et al. Role of polyisoprenoids in tobacco resistance against biotic stresses. Physiol. Plant. 2009, 135, 351–364. [Google Scholar] [CrossRef] [PubMed]
- Hancock, R.D.; Morris, W.L.; Ducreux, L.J.; Morris, J.A.; Usman, M.; Verrall, S.R.; Fuller, J.; Simpson, C.G.; Zhang, R.; Hedley, P.E.; et al. Physiological, biochemical and molecular responses of the potato (Solanum tuberosum L.) plant to moderately elevated temperature. Plant Cell Environ. 2014, 37, 439–450. [Google Scholar] [CrossRef]
- Mangelsen, E.; Kilian, J.; Harter, K.; Jansson, C.; Wanke, D.; Sundberg, E. Transcriptome analysis of high-temperature stress in developing barley caryopses: early stress responses and effects on storage compound biosynthesis. Mol. Plant 2011, 4, 97–115. [Google Scholar] [CrossRef]
- Wang, Z.D.; Yan, N.; Wang, Z.H.; Zhang, X.H.; Zhang, J.Z.; Xue, H.M.; Wang, L.X.; Zhan, Q.; Xu, Y.P.; Guo, D.P. RNA-seq analysis provides insight into reprogramming of culm development in Zizania latifolia induced by Ustilago esculenta. Plant Mol. Biol. 2017, 95, 533–547. [Google Scholar] [CrossRef]
- Wang, Z.; Gerstein, M.; Snyder, M. RNA-Seq: A revolutionary tool for transcriptomics. Nat. Rev. Genet. 2009, 10, 57–63. [Google Scholar] [CrossRef]
- Mortazavi, A.; Williams, B.A.; McCue, K.; Schaeffer, L.; Wold, B. Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat. Methods 2008, 5, 621–628. [Google Scholar] [CrossRef]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, X.; Du, Y.; Liu, X.; Zhang, H.; Liu, Y.; Yan, N.; Zhang, Z. Genome-wide analysis of long non-coding RNAs in potato and their potential role in tuber sprouting process. Int. J. Mol. Sci. 2018, 19, 101. [Google Scholar]
- Yan, N.; Du, Y.; Liu, X.; Zhang, H.; Liu, Y.; Shi, J.; Xue, S.J.; Zhang, Z. Analyses of effects of α-cembratrien-diol on cell morphology and transcriptome of Valsa mali var. mali. Food Chem. 2017, 214, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.O.; Perez-Fons, L.; Robertson, F.P.; Bramley, P.M.; Fraser, P.D. Functional characterization of long-chain prenyl diphosphate synthases from tomato. Biochem. J. 2013, 449, 729–740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Block, A.; Fristedt, R.; Rogers, S.; Kumar, J.; Barnes, B.; Barnes, J.; Elowsky, C.G.; Wamboldt, Y.; Mackenzie, S.A.; Redding, K.; et al. Functional modeling identifies paralogous solanesyl-diphosphate synthases that assemble the side chain of plastoquinone-9 in plastids. J. Biol. Chem. 2013, 288, 27594–27606. [Google Scholar] [CrossRef]
- Ksas, B.; Becuwe, N.; Chevalier, A.; Havaux, M. Plant tolerance to excess light energy and photooxidative damage relies on plastoquinone biosynthesis. Sci. Rep. 2015, 5, 10919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, N.; Wang, X.; Xu, X.; Guo, D.; Wang, Z.; Zhang, J.; Hyde, K.D.; Liu, H. Plant growth and photosynthetic performance of Zizania latifolia are altered by endophytic Ustilago esculenta infection. Physiol. Mol. Plant Pathol. 2013, 83, 75–83. [Google Scholar] [CrossRef]
- Yan, N.; Xu, X.; Wang, Z.; Huang, J.; Guo, D. Interactive effects of temperature and light intensity on photosynthesis and antioxidant enzyme activity in Zizania latifolia Turcz. plants. Photosynthetica 2013, 51, 127–138. [Google Scholar] [CrossRef]
- Sierro, N.; Battey, J.N.; Ouadi, S.; Bakaher, N.; Bovet, L.; Willig, A.; Goepfert, S.; Peitsch, M.C.; Ivanov, N.V. The tobacco genome sequence and its comparison with those of tomato and potato. Nat. Commun. 2014, 5, 3833. [Google Scholar] [CrossRef] [PubMed]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.; Pertea, G.; Trapnell, C.; Pimentel, H.; Kelley, R.; Salzberg, S.L. TopHat2: Accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol. 2013, 14, R36. [Google Scholar] [CrossRef] [PubMed]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—a Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Genome Biol. 2010, 11, R106. [Google Scholar] [CrossRef] [PubMed]
- Young, M.D.; Wakefield, M.J.; Smyth, G.K.; Oshlack, A. Gene ontology analysis for RNA-seq: Accounting for selection bias. Genome Biol. 2010, 11, R14. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Goto, S.; Sato, Y.; Kawashima, M.; Furumichi, M.; Tanabe, M. Data, information, knowledge and principle: Back to metabolism in KEGG. Nucleic Acids Res. 2014, 42, D199–D205. [Google Scholar] [CrossRef]
- Mizoi, J.; Shinozaki, K.; Yamaguchi-Shinozaki, K. AP2/ERF family transcription factors in plant abiotic stress responses. BBA-Gene Regul. Mech. 2012, 1819, 86–96. [Google Scholar] [CrossRef] [PubMed]
- Darabi, M.; Izadi-Darbandi, A.; Masoudi-Nejad, A.; Naghavi, M.R.; Nemat-Zadeh, G. Bioinformatics study of the 3-hydroxy-3-methylglotaryl-coenzyme A reductase (HMGR) gene in Gramineae. Mol. Biol. Rep. 2012, 39, 8925–8935. [Google Scholar] [CrossRef]
- Wang, L.J.; Fang, X.; Yang, C.Q.; Li, J.X.; Chen, X.Y. Biosynthesis and regulation of secondary terpenoid metabolism in plants. Sci. Sin. Vitae 2013, 43, 1030–1046. [Google Scholar] [CrossRef]
- Ramakrishna, A.; Ravishankar, G.A. Influence of abiotic stress signals on secondary metabolites in plants. Plant Signal. Behav. 2011, 6, 1720–1731. [Google Scholar] [Green Version]
- Zobayed, S.M.; Afreen, F.; Kozai, T. Temperature stress can alter the photosynthetic efficiency and secondary metabolite concentrations in St. John’s wort. Plant Physiol. Biochem. 2005, 43, 977–984. [Google Scholar] [CrossRef] [PubMed]
- Kubien, D.S.; Sage, R.F. The temperature response of photosynthesis in tobacco with reduced amounts of Rubisco. Plant Cell Environ. 2008, 31, 407–418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rushton, P.J.; Bokowiec, M.T.; Han, S.; Zhang, H.; Brannock, J.F.; Chen, X.; Laudeman, T.W.; Timko, M.P. Tobacco transcription factors: novel insights into transcriptional regulation in the Solanaceae. Plant Physiol. 2008, 147, 280–295. [Google Scholar] [CrossRef]
- Rushton, P.J.; Bokowiec, M.T.; Laudeman, T.W.; Brannock, J.F.; Chen, X.; Timko, M.P. TOBFAC: the database of tobacco transcription factors. BMC Bioinformatics 2008, 9, 53. [Google Scholar] [CrossRef] [PubMed]
- Hemmerlin, A.; Hoeffler, J.F.; Meyer, O.; Tritsch, D.; Kagan, I.A.; Grosdemange-Billiard, C.; Rohmer, M.; Bach, T.J. Cross-talk between the cytosolic mevalonate and the plastidial methylerythritol phosphate pathways in tobacco bright yellow-2 cells. J. Biol. Chem. 2003, 278, 26666–26676. [Google Scholar] [CrossRef] [PubMed]
- Mendoza-Poudereux, I.; Kutzner, E.; Huber, C.; Segura, J.; Eisenreich, W.; Arrillaga, I. Metabolic cross-talk between pathways of terpenoid backbone biosynthesis in spike lavender. Plant Physiol. Biochem. 2015, 95, 113–120. [Google Scholar] [CrossRef]
- Schaller, H.; Grausem, B.; Benveniste, P.; Chye, M.L.; Tan, C.T.; Song, Y.H.; Chua, N.H. Expression of the Hevea brasiliensis (HBK) Mull. Arg. 3-hydroxy-3-methylglutaryl-coenzyme A reductase 1 in tobacco results in sterol overproduction. Plant Physiol. 1995, 109, 761–770. [Google Scholar] [CrossRef]
- Szkopińska, A.; Płochocka, D. Farnesyl diphosphate synthase: regulation of product specificity. Acta Biochim Pol. 2005, 52, 45–55. [Google Scholar]
- Wu, S.; Jiang, Z.; Kempinski, C.; Nybo, S.E.; Husodo, S.; Williams, R.; Chappell, J. Engineering triterpene metabolism in tobacco. Planta 2012, 236, 867–877. [Google Scholar] [CrossRef]
- Zhang, H.; Niu, D.; Wang, J.; Zhang, S.; Yang, Y.; Jia, H.; Cui, H. Engineering a platform for photosynthetic pigment, hormone and cembrane-related diterpenoid production in Nicotiana tabacum. Plant Cell Physiol. 2015, 56, 2125–2138. [Google Scholar] [CrossRef]
- Hasunuma, T.; Takeno, S.; Hayashi, S.; Okumoto, H.; Katto, H.; Kajiyami, S.; Kobayashi, A. Overexpression of 1-deoxy-Dxylulose-5-phosphate reductoisomerase gene in chloroplast contributes to increment of isoprenoid production. J. Biosci. Bioeng. 2008, 105, 518–526. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Forward Primer (5′–3′) | Reverse Primer (5′–3′) |
|---|---|---|
| Actin | CCACACAGGTGTGATGGTTG | GTGGCTAACACCATCACCAG |
| NtHMGR | GTCAGGTGGCGTGAGAAG | GTCCACGGCGGCTATCTT |
| NtDXS | ACCACCAACACCTCTTTT | TGATGACCAACATCCCAT |
| NtDXR | TGGTAAGAGGGTTCAGTGTT | CAGCCAGAGCATCTTTGAG |
| NtFPS | TGAGTTCCAGACTGCCTCT | GCCAATCTTACCCAGCAC |
| NtGGPS | CCCAATAAAACCTTCACTG | CACAGGTGGGTCTTTTACTA |
| NtSPS | GTTCCAGGTTGTTGATGAC | CTCGGAAAGGACTAGAAGG |
| GO ID | GO Terms | Type a | Corrected p-Value | Test b | Ref. c |
|---|---|---|---|---|---|
| GO:0004553 | Hydrolase activity, hydrolyzing O-glycosyl compounds | P | 1.43 × 10−9 | 77 | 1039 |
| GO:0016798 | Hydrolase activity, acting on glycosyl bonds | P | 1.43 × 10−9 | 80 | 1107 |
| GO:0005975 | Carbohydrate metabolic process | F | 1.13 × 10−7 | 117 | 2085 |
| GO:0006629 | Lipid metabolic process | F | 6.13 × 10−7 | 104 | 1831 |
| GO:0005507 | Copper ion binding | P | 6.13 × 10−7 | 28 | 243 |
| GO:0003824 | Catalytic activity | P | 6.13 × 10−7 | 810 | 23,582 |
| GO:0055114 | Oxidation-reduction process | F | 2.26 × 10−6 | 199 | 4547 |
| GO:0044710 | Single-organism metabolic process | F | 4.53 × 10−6 | 380 | 9926 |
| Gene Family | Number of TFs | Percentage (%) | TF Type |
|---|---|---|---|
| MYB | 16 | 13.11 | Transcription factor |
| AP2-EREBP | 12 | 9.84 | Transcription factor |
| NAC | 12 | 9.84 | Transcription factor |
| bHLH | 10 | 8.20 | Transcription factor |
| HB | 9 | 7.38 | Transcription factor |
| SBP | 7 | 5.74 | Transcription factor |
| C2C2-Dof | 5 | 4.10 | Transcription factor |
| LOB | 5 | 4.10 | Transcription factor |
| C3H | 4 | 3.28 | Transcription factor |
| CCAAT | 4 | 3.28 | Transcription factor |
| MADS | 4 | 3.28 | Transcription factor |
| Orphans | 4 | 3.28 | Transcriptional regulator |
| WRKY | 4 | 3.28 | Transcription factor |
| C2C2-GATA | 3 | 2.46 | Transcription factor |
| GRAS | 3 | 2.46 | Transcription factor |
| PLATZ | 3 | 2.46 | Transcription factor |
| bZIP | 2 | 1.64 | Transcription factor |
| C2H2 | 2 | 1.64 | Transcription factor |
| LIM | 2 | 1.64 | Transcription factor |
| SNF2 | 2 | 1.64 | Transcriptional regulator |
| TRAF | 2 | 1.64 | Transcriptional regulator |
| HSF | 1 | 0.82 | Transcription factor |
| LUG | 1 | 0.82 | Transcriptional regulator |
| S1Fa-like | 1 | 0.82 | Transcription factor |
| SET | 1 | 0.82 | Transcriptional regulator |
| TCP | 1 | 0.82 | Transcription factor |
| Trihelix | 1 | 0.82 | Transcription factor |
| zf-HD | 1 | 0.82 | Transcription factor |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yan, N.; Du, Y.; Zhang, H.; Zhang, Z.; Liu, X.; Shi, J.; Liu, Y. RNA Sequencing Provides Insights into the Regulation of Solanesol Biosynthesis in Nicotiana tabacum Induced by Moderately High Temperature. Biomolecules 2018, 8, 165. https://doi.org/10.3390/biom8040165
Yan N, Du Y, Zhang H, Zhang Z, Liu X, Shi J, Liu Y. RNA Sequencing Provides Insights into the Regulation of Solanesol Biosynthesis in Nicotiana tabacum Induced by Moderately High Temperature. Biomolecules. 2018; 8(4):165. https://doi.org/10.3390/biom8040165
Chicago/Turabian StyleYan, Ning, Yongmei Du, Hongbo Zhang, Zhongfeng Zhang, Xinmin Liu, John Shi, and Yanhua Liu. 2018. "RNA Sequencing Provides Insights into the Regulation of Solanesol Biosynthesis in Nicotiana tabacum Induced by Moderately High Temperature" Biomolecules 8, no. 4: 165. https://doi.org/10.3390/biom8040165