Associations of SNPs of the ADIPOQ Gene with Serum Adiponectin Levels, Unstable Angina, and Coronary Artery Disease

 , , , , ,
, , , , ,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Sample Collection and Measurements
2.3. SNP Selection
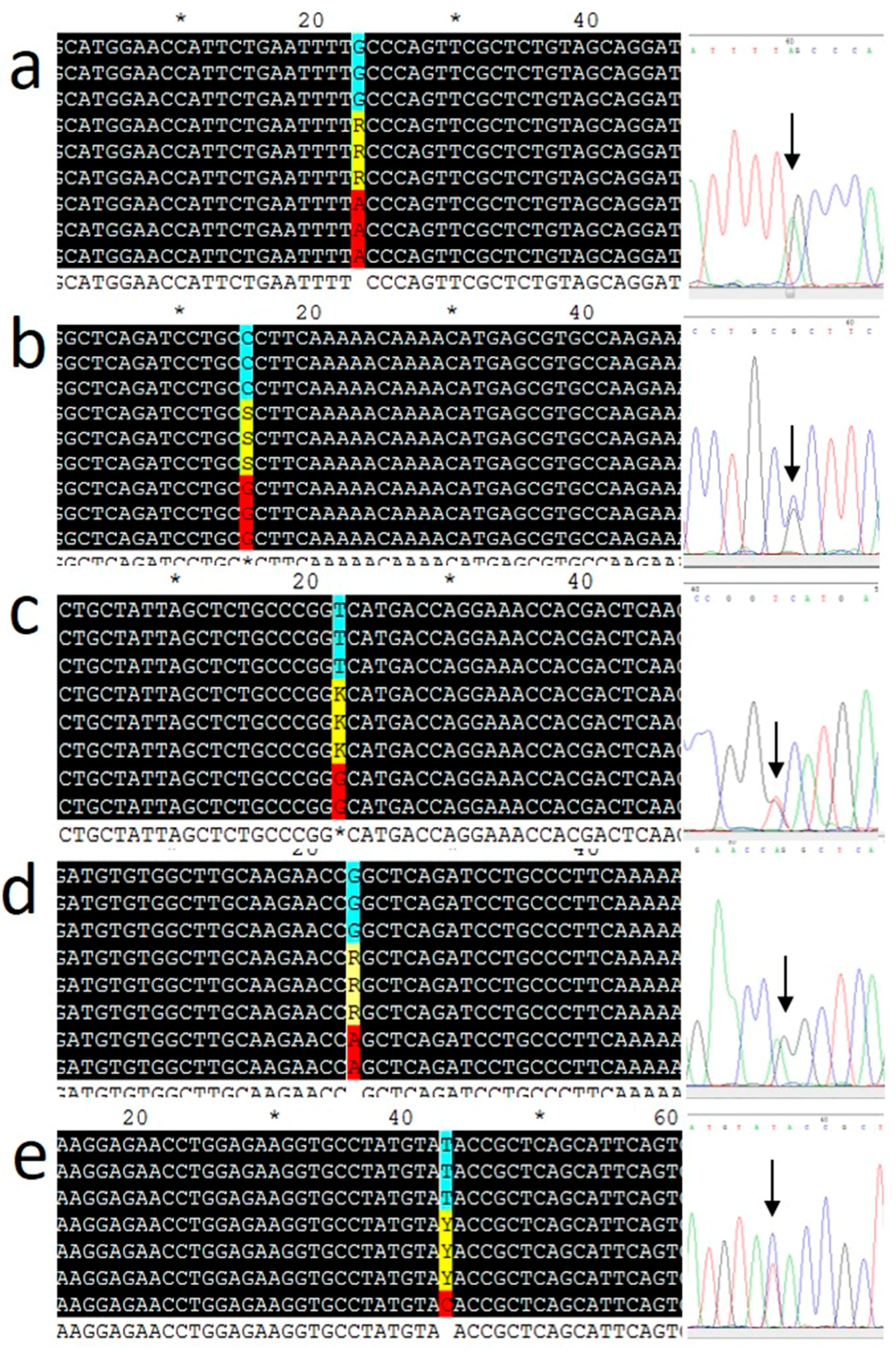
2.4. Genotyping
2.5. Statistical Analysis
3. Results
3.1. Subject Characteristics and Genotyping
3.2. Associations of the ADIPOQ SNPs with Serum Concentrations of Adiponectin
3.3. Associations of the rs266729 and rs182052 SNPs with CAD and T2D
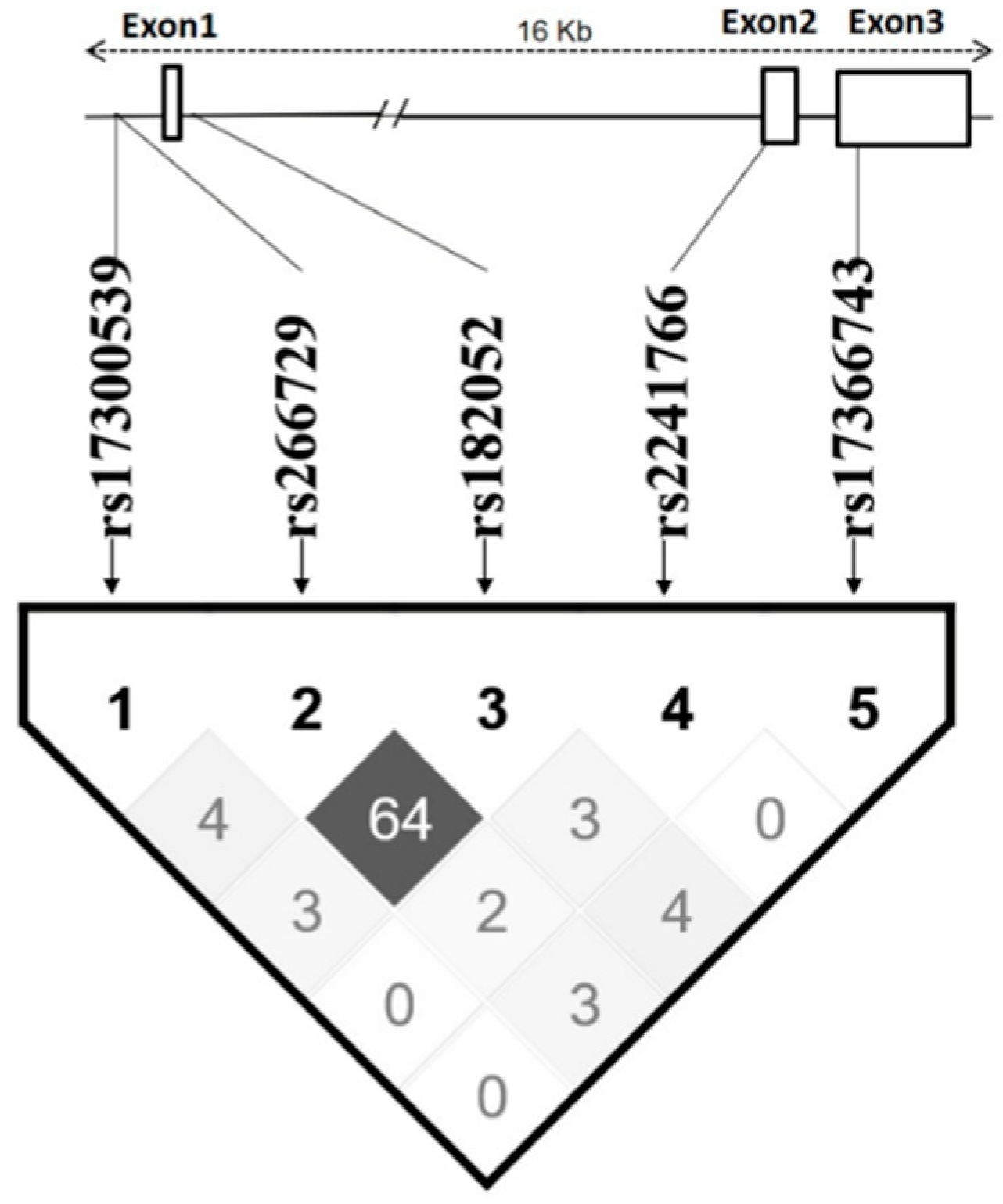
3.4. Haplotype Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Shimada, K.; Miyazaki, T.; Daida, H. Adiponectin and atherosclerotic disease. Clin. Chim. Acta 2004, 344, 1–12. [Google Scholar] [CrossRef]
- Ntaios, G.; Gatselis, N.K.; Makaritsis, K.; Dalekos, G.N. Adipokines as mediators of endothelial function and atherosclerosis. Atherosclerosis 2013, 227, 216–221. [Google Scholar] [CrossRef]
- Shehzad, A.; Iqbal, W.; Shehzad, O.; Lee, Y.S. Adiponectin: Regulation of its production and its role in human diseases. Hormones 2012, 11, 8–20. [Google Scholar] [CrossRef] [PubMed]
- Wassel, C.L.; Pankow, J.S.; Rasmussen-Torvik, L.J.; Li, N.; Taylor, K.D.; Guo, X.; Goodarzi, M.O.; Palmas, W.R.; Post, W.S. Associations of SNPs in ADIPOQ and subclinical cardiovascular disease in the multi-ethnic study of atherosclerosis (MESA). Obesity 2011, 19, 840–847. [Google Scholar] [CrossRef] [PubMed]
- Hivert, M.F.; Manning, A.K.; McAteer, J.B.; Florez, J.C.; Dupuis, J.; Fox, C.S.; O’Donnell, C.J.; Cupples, L.A.; Meigs, J.B. Common Variants in the Adiponectin Gene (ADIPOQ) Associated with Plasma Adiponectin Levels, Type 2 Diabetes, and Diabetes-Related Quantitative Traits. Diabetes 2008, 57, 3353–3359. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, J.F.; Phillips, C.M.; Tierney, A.C.; Pérez-Martínez, P.; Defoort, C.; Helal, O.; Lairon, D.; Planells, R.; Shaw, D.I.; Lovegrove, J.A.; et al. Gene-nutrient interactions in the metabolic syndrome: Single nucleotide polymorphisms in ADIPOQ and ADIPOR1interact with plasma saturated fatty acids to modulate insulin resistance. Am. J. Clin. Nutr. 2009, 91, 794–801. [Google Scholar] [CrossRef]
- Hou, H.; Ge, S.; Zhao, L.; Wang, C.; Wang, W.; Zhao, X.; Sun, Z. An Updated Systematic Review and Meta-analysis of Association Between Adiponectin Gene Polymorphisms and Coronary Artery Disease. OMICS J. Integr. Boil. 2017, 21, 340–351. [Google Scholar] [CrossRef]
- Kanu, J.S.; Qiu, S.; Cheng, Y.; Li, R.; Kou, C.; Gu, Y.; Bai, Y.; Shi, J.; Li, Y.; Liu, Y.; et al. Associations between three common single nucleotide polymorphisms (rs266729, rs2241766, and rs1501299) of ADIPOQ and cardiovascular disease: A meta-analysis. Lipids Health Dis. 2018, 17, 126. [Google Scholar] [CrossRef]
- Anagnostis, P.; Athyros, V.G.; Kita, M.; Karagiannis, A. Is there any association between adiponectin gene polymorphisms and cardiovascular disease? Angiology 2013, 64, 253–256. [Google Scholar] [CrossRef] [PubMed]
- Potapov, V.A.; Chistiakov, D.A.; Dubinina, A.; Shamkhalova, M.S.; Shestakova, M.V.; Nosikov, V.V. Adiponectin and Adiponectin Receptor Gene Variants in Relation to Type 2 Diabetes and Insulin Resistance-Related Phenotypes. Rev. Diabet. Stud. 2008, 5, 28–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korytina, G.F.; Akhmadishina, L.Z.; Kochetova, O.V.; Aznabaeva, Y.G.; Zagidullin, S.Z.; Victorova, T.V. Inflammatory and Immune Response Genes Polymorphisms are Associated with Susceptibility to Chronic Obstructive Pulmonary Disease in Tatars Population from Russia. Biochem. Genet. 2016, 54, 388–412. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Fall, T.; van Dam, R.M.; Flyvbjerg, A.; Zethelius, B.; Ingelsson, E.; Hägg, S. Evidence of a causal relationship between adiponectin levels and insulin sensitivity: A Mendelian randomization study. Diabetes 2013, 62, 1338–1344. [Google Scholar] [CrossRef] [PubMed]
- Menzaghi, C.; Salvemini, L.; Concetta De Bonis, P.; Fini, G.; Doria, A.; Trischitta, V. Circulating HMW adiponectin isoform is heritable and shares a common genetic background with insulin resistance in non diabetic white Caucasians from Italy: Evidence from a family-based study. J. Intern. Med. 2010, 267, 287–294. [Google Scholar] [CrossRef] [PubMed]
- Dhillon, P.K.; Penney, K.L.; Schumacher, F.; Rider, J.R.; Sesso, H.D.; Pollak, M.; Mucci, L.A. Common polymorphisms in the adiponectin and its receptor genes, adiponectin levels and the risk of prostate cancer. Cancer Epidemiol. Prev. Biomark. 2011, 20, 2618–2627. [Google Scholar] [CrossRef]
- Han, L.Y.; Wu, Q.H.; Jiao, M.L.; Hao, Y.H.; Liang, L.B.; Gao, L.J.; Kang, Z. Associations between single-nucleotide polymorphisms (+ 45T> G,+ 276G> T,− 11377C> G,− 11391G> A) of adiponectin gene and type 2 diabetes mellitus: A systematic review and meta-analysis. Diabetologia 2011, 54, 2303–2314. [Google Scholar] [CrossRef] [PubMed]
- Guzman-Ornelas, M.O.; Chavarria-Avila, E.; Munoz-Valle, J.F.; Armas-Ramos, L.E.; Castro-Albarran, J.; Aldrete, M.E.; Oregon-Romero, E.; Vazquez-Del Mercado, M.; Navarro-Hernandez, R.E. Association of ADIPOQ+ 45T> G polymorphism with body fat mass and blood levels of soluble adiponectin and inflammation markers in a Mexican-Mestizo population. Diabetes Metab. Syndr. Obes. Targets Ther. 2012, 5, 369. [Google Scholar] [CrossRef]
- Ramya, K.; Ayyappa, K.A.; Ghosh, S.; Mohan, V.; Radha, V. Genetic association of ADIPOQ gene variants with type 2 diabetes, obesity and serum adiponectin levels in south Indian population. Gene 2013, 532, 253–262. [Google Scholar] [CrossRef] [PubMed]
- Gumanova, N.G.; Gavrilova, N.E.; Chernushevich, O.I.; Kots, A.Y.; Metelskaya, V.A. Ratios of leptin to insulin and adiponectin to endothelin are sex-dependently associated with extent of coronary atherosclerosis. Biomarkers 2017, 22, 239–245. [Google Scholar] [CrossRef] [PubMed]
- Gensini, G.G. A more meaningful scoring system for determining the severity of coronary heart disease. Am. J. Cardiol. 1983, 51, 606. [Google Scholar] [CrossRef]
- Hamm, C.W.; Bassand, J.P.; Agewall, S.; Boersma, E.; Bueno, H.; Caso, P.; Dudek, D.; Gielen, S.; Huber, K.; Ohman, M.; et al. ESC Guidelines for the management of acute coronary syndromes in patients presenting without persistent ST-segment elevation: The Task Force for the management of acute coronary syndromes (ACS) in patients presenting without persistent ST-segment elevation of the European Society of Cardiology (ESC). Eur. Heart J. 2011, 32, 2999–3054. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. Use of Glycated Haemoglobin (HbA1c) in the Diagnosis of Diabetes Mellitus: Abbreviated Report of a WHO Consultation; WHO Guidelines Approved by the Guidelines Review Committee; World Health Organization: Geneva, Switzerland, 2011. [Google Scholar]
- Friedewald, W.T.; Levy, R.I.; Fredrickson, D.S. Estimation of the concentration of low-density lipoprotein cholesterol in plasma, without use of the preparative ultracentrifuge. Clin. Chem. 1972, 18, 499–502. [Google Scholar] [PubMed]
- Shen, D.; Xing, S.; Chen, C. Adiponectin gene polymorphisms contributes to ischemic stroke risk: A meta-analysis. J. Renin Angiotensin Aldosterone Syst. 2015, 16, 178–184. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, S.; Gaunt, T.R.; Day, I.N.M. Hardy-Weinberg equilibrium testing of biological ascertainment for Mendelian randomization studies. Am. J. Epidemiol. 2009, 169, 505–514. [Google Scholar] [CrossRef] [PubMed]
- Groeneveld, R.A.; Meeden, G. Measuring Skewness and Kurtosis. J. R. Stat. Soc. Ser. D Stat. 1984, 33, 391–399. [Google Scholar] [CrossRef]
- Zhang, Z. Missing data imputation: Focusing on single imputation. Ann. Transl. Med. 2016, 4, 9. [Google Scholar] [CrossRef] [PubMed]
- Hart, A. Mann–Whitney test is not just a test of medians: Differences in spread can be important. BMJ 2001, 323, 391–393. [Google Scholar] [CrossRef] [PubMed]
- Westfall, P.H.; Young, S.S. Resampling-Based Multiple Testing: Examples and Methods for p-Value Adjustment; John Wiley & Sons: Hoboken, NJ, USA, 1993; Volume 279. [Google Scholar]
- Pollard, K.S.; Dudoit, S.; Van Der Laan, M.J. Multiple Testing Procedures: The multtest Package and Applications to Genomics. In Nonclinical Statistics for Pharmaceutical and Biotechnology Industries; Springer Science and Business Media LLC: New York, NY, USA, 2005; pp. 249–271. [Google Scholar]
- Sheather, S. A Modern Approach to Regression with R; Springer Science and Business Media LLC: New York, NY, USA, 2009. [Google Scholar]
- Barrett, J.; Fry, B.; Maller, J.; Daly, M.J. Haploview: Analysis and visualization of LD and haplotype maps. Bioinformatics 2004, 21, 263–265. [Google Scholar] [CrossRef]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.; Bender, D.; Sham, P.C. PLINK: A tool set for whole-genome association and population-based linkage analyses. Am. J. Hum. genet. 2007, 81, 559–575. [Google Scholar] [CrossRef] [PubMed]
- Klimushina, M.V.; Gumanova, N.G.; Kutsenko, V.A.; Divashuk, M.G.; Smetnev, S.A.; Kiseleva, A.V.; Metelskaya, V.A.; Meshkov, A.N. Association of common polymorphisms in IL-6 and IL6ST genes with levels of inflammatory markers and coronary stenosis. Meta Gene 2019, 21, 100593. [Google Scholar] [CrossRef]
- Guerre-Millo, M. Adiponectin: An update. Diabetes Metab. 2008, 34, 12–18. [Google Scholar] [CrossRef]
- Zhang, H.; Mo, X.; Hao, Y.; Gu, D. Association between polymorphisms in the adiponectin gene and cardiovascular disease: A meta-analysis. BMC Med. Genet. 2012, 13, 40. [Google Scholar] [CrossRef] [PubMed]
- Tong, G.; Wang, N.; Leng, J.; Tong, X.; Shen, Y.; Yang, J.; Ye, X.; Zhou, L.; Zhou, Y. Common variants in adiponectin gene are associated with coronary artery disease and angiographical severity of coronary atherosclerosis in type 2 diabetes. Cardiovasc. Diabetol. 2013, 12, 67. [Google Scholar] [CrossRef] [PubMed]
- Qiao, L.; MacLean, P.S.; Schaack, J.; Orlicky, D.J.; Darimont, C.; Pagliassotti, M.; Friedman, J.E.; Shao, J. C/EBP Regulates Human Adiponectin Gene Transcription Through an Intronic Enhancer. Diabetes 2005, 54, 1744–1754. [Google Scholar] [CrossRef] [PubMed]
- Grabe, N. AliBaba2: Context specific identification of transcription factor binding sites. Silico Biol. 2002, 2, S1–S15. [Google Scholar]
- Laumen, H.; Saningong, A.D.; Heid, I.M.; Hess, J.; Herder, C.; Claussnitzer, M.; Klopp, N. Functional characterization of promoter variants of the adiponectin gene complemented by epidemiological data. Diabetes 2009, 58, 984–991. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Ma, J.; Brismar, K.; Efendić, S.; Gu, H.F. A single nucleotide polymorphism alters the sequence of SP1 binding site in the adiponectin promoter region and is associated with diabetic nephropathy among type 1 diabetic patients in the Genetics of Kidneys in Diabetes Study. J. Diabetes Complicat. 2009, 23, 265–272. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| SNP | Oligonucleotide Primers | Probes | Amplicon Size (bp) |
|---|---|---|---|
| rs17300539 | F: 5-TTGAAGTTGGTGCTGGCATC-3 | (FAM)-CAGGATCTGAGCCGGTTC-(RTQ1) | 193 |
| R: 5-GGAAGCTGCCACCCACTTA-3 | (R6G)-CAAGAACCAGCTCAGATCC-(BHQ2) | ||
| rs266729 | F: 5-GTTGGTGCTGGCATC-3 | (FAM)-CAGATCCTGCCCTTCAAA-(RTQ1) | 127 |
| R: 5-CCTTGGACTTTCTTGGCACG-3 | (R6G)-TGCGCTTCAAAAACAAAACAT-(BHQ2) | ||
| rs182052 | F: 5-CCTCCGTTCTCCCAC-3 | (FAM)-CCATTCTGAATTTTGCCCAGT-(RTQ1) | 145 |
| R: 5-ACCCTTCCACCTTACTGACC-3 | (R6G)-CCATTCTGAATTTTACCCAGTTCG-(BHQ2) | ||
| rs2241766 | F: 5-GGATTCCAGGGCTCAGGATG-3 | (FAM)-TCTGCCCGGTCATGA-(RTQ1) | 139 |
| R: 5-GCCATCCAACCTGTGCAG-3 | (R6G)-TCCTGGTCATGCCCGG-(BHQ2) | ||
| rs17366743 | F:5-GGCAGGAAAGGAGAACC-3 | (FAM)-AGCGGTATACATAGGCACC-(RTQ1) | 180 |
| R: 5-GTACAGCCCAGGAATGTTGC-3 | (R6G)-CTATGTACACCGCTCAGC-(BHQ2) |
| Parameter | Total Group (N = 447) |
|---|---|
| general characteristics | |
| Age, years | 61 ± 9 |
| Weight, kg | 85.2 ± 15.0 |
| Body mass index, kg/m2 | 28.73 (26.22–32.7) |
| Systolic blood pressure, mmHg | 131 ± 16 |
| Diastolic blood pressure, mmHg | 80 ± 9 |
| Heart rate, bpm | 68 (64–74] |
| serum biochemical parameters | |
| Total cholesterol, mmol/L | 4.98 ± 1.27 |
| LDL cholesterol, mmol/L | 3.15 ± 1.15 |
| HDL cholesterol, mmol/L | 1.00 ± 0.26 |
| Triglycerides, mmol/L | 1.56 (1.14–2.12) |
| Adiponectin, µg/mL | 7.87 (5.64–11.81) |
| Fasting glucose, mmol/L | 5.6 (5.2–6.2) |
| Insulin, µIU/mL | 10.5 (7.4–14.5) |
| HOMA-IR index | 2.66 (1.83–3.99) |
| C-reactive protein, mg/L | 2.7 (1.2–5.9) |
| diseases | |
| Type 2 diabetes mellitus, % | 17.2 (13.8–21.1) |
| Hypertension, % | 81.4 (77.5–84.9) |
| Unstable angina, % | 4.9 (3.1–7.4) |
| Coronary artery disease, % | 64.0 (59.3–68.4) |
| medications | |
| Statins, % | 92.8 (90.0–95.1) |
| Warfarin (anticoagulants), % | 7.4 (5.1–10.2) |
| Clopidogrel (antiplatelet drugs), % | 59.7 (55.0–64.3) |
| Aspirin (antiplatelet drugs), % | 89.9 (86.8–92.6) |
| Angiotensin converting enzyme inhibitors, % | 69.6 (65.1–73.8) |
| Beta-adrenoblockers, % | 87.0 (83.6–90.0) |
| Calcium antagonists, % | 27.7 (23.6–32.1) |
| Diuretics, % | 25.3 (21.3–29.6) |
| SNP | Location on Chromosome 3 a | Relation | Alleles, M/m | Genotypes, N for MM/Mm/mm | Observed MAF, % | MAF | |
|---|---|---|---|---|---|---|---|
| Mean MAF, % | Maximal MAF EUR, % | ||||||
| rs17300539 | 186841671 | Promoter | G/A | 385/60/2 | 0.07 | 0.05 | 0.07 |
| rs266729 | 186841685 | Promoter | C/G | 240/154/53 | 0.29 | 0.23 | 0.32 |
| rs182052 | 186842993 | Intron 1 | G/A | 177/196/74 | 0.38 | 0.39 | 0.56 |
| rs2241766 | 186853103 | Exon 2 coding synonymous | T/G | 391/54/2 | 0.06 | 0.05 | 0.12 |
| rs17366743 | 186854300 | Exon 3 coding nonsynonymous | T/C | 432/14/1 | 0.02 | 0.02 | 0.04 |
| SNP (MAF, %) | Univariate Wilcoxon–Mann–Whitney Test | Multivariate Linear Regression Parameters | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Genetic Model | Δ Log (Adiponectin Concentration) | CI 95% | p | Genetic Model | β | 95% CI | p | Adjusted p | |
| rs17300539 (7%) | D | 0.11 | −0.03, 0.25 | 0.125 | D | 0.14 | −0.03, 0.31 | 0.103 | 0.199 |
| rs266729 (29%) | R | −0.16 | −0.32, –0.01 | 0.040 a | A | −0.08 | −0.16, 0.01 | 0.079 | 0.193 |
| rs182052 (38%) | D | −0.11 | −0.22, –0.01 | 0.034 a | A | −0.11 | −0.19, -0.03 | 0.006 a | 0.016 a |
| rs2241766 (6%) | D | 0.11 | −0.06, 0.27 | 0.210 | A | 0.07 | −0.1, 0.23 | 0.436 | 0.645 |
| rs17366743 (2%) | R | 0.89 | −0.92, 3.65 | 0.151 | D | 0.05 | −0.25, 0.34 | 0.763 | 0.728 |
| Diseases | Univariate Fisher Exact Test | Multivariate Logistic Regression Parameters | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Genetic Model (N for OR) | % in Group with MM/ % in Group with mm | OR | CI 95% | p | Genetic Model | OR | 95% CI | p | Adjusted p | |
| rs266729 SNP | ||||||||||
| Unstable angina | R (394/53) | 4.1%/11.3% | 3.02 | 1.04–7.74 | 0.035 a | R | 3.59 | 1.17–10.01 | 0.018 a | 0.045 a |
| Type 2 diabetes | D (240/207) | 14%/21% | 1.59 | 0.97–2.62 | 0.078 | D | 1.59 | 0.93–2.72 | 0.091 | 0.091 |
| Coronary artery disease | R (394/53) | 67%/79% | 2.07 | 1.03–4.48 | 0.033 a | A | 1.48 | 1.09–2.03 | 0.015 a | 0.045 a |
| rs182052 SNP | ||||||||||
| Unstable angina | R (373/74) | 3.8%/10.8% | 3.11 | 1.20–7.56 | 0.017 a | A | 2.55 | 1.4–4.82 | 0.003 a | 0.018 a |
| Type 2 diabetes | D (177/270) | 11%/21% | 2.1 | 1.23–3.71 | 0.007 a | D | 2.29 | 1.29–4.21 | 0.006 a | 0.024 a |
| Coronary artery disease | R (373/74) | 66%/78% | 1.93 | 1.07–3.64 | 0.024 a | A | 1.55 | 1.15–2.09 | 0.004 a | 0.021 a |
| No | Haplotype | Frequency, % | β, Log Adiponectin | OR, Unstable Angina | OR, Type 2 Diabetes | OR, Coronary Artery Disease |
|---|---|---|---|---|---|---|
| 1 | GCGTT a | 48.7 | - | - | - | |
| 2 | GGATT | 27.4 | −0.094 (p = 0.047) b | 2.054 (p = 0.041) b | 1.268 (p = 0.266) | 1.546 (p = 0.014) b |
| 3 | GCATT | 9.8 | −0.15 (p = 0.042) b | 3.597 (p = 0.007) b | 1.479 (p = 0.247) | 1.337 (p = 0.287) |
| 4 | ACGTT | 6.5 | 0.09 (p = 0.301) | 0.486 (p = 0.496) | 1.164 (p = 0.711) | 1.237 (p = 0.508) |
| 5 | GCGGT | 5.7 | 0.035 (p = 0.706) | 0.54 (p = 0.565) | 1.017 (p = 0.968) | 0.76 (p = 0.314) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Smetnev, S.; Klimushina, M.; Kutsenko, V.; Kiseleva, A.; Gumanova, N.; Kots, A.; Skirko, O.; Ershova, A.; Yarovaya, E.; Metelskaya, V.; et al. Associations of SNPs of the ADIPOQ Gene with Serum Adiponectin Levels, Unstable Angina, and Coronary Artery Disease. Biomolecules 2019, 9, 537. https://doi.org/10.3390/biom9100537
Smetnev S, Klimushina M, Kutsenko V, Kiseleva A, Gumanova N, Kots A, Skirko O, Ershova A, Yarovaya E, Metelskaya V, et al. Associations of SNPs of the ADIPOQ Gene with Serum Adiponectin Levels, Unstable Angina, and Coronary Artery Disease. Biomolecules. 2019; 9(10):537. https://doi.org/10.3390/biom9100537
Chicago/Turabian StyleSmetnev, Stepan, Marina Klimushina, Vladimir Kutsenko, Anna Kiseleva, Nadezhda Gumanova, Alexander Kots, Olga Skirko, Alexandra Ershova, Elena Yarovaya, Victoria Metelskaya, and et al. 2019. "Associations of SNPs of the ADIPOQ Gene with Serum Adiponectin Levels, Unstable Angina, and Coronary Artery Disease" Biomolecules 9, no. 10: 537. https://doi.org/10.3390/biom9100537
APA StyleSmetnev, S., Klimushina, M., Kutsenko, V., Kiseleva, A., Gumanova, N., Kots, A., Skirko, O., Ershova, A., Yarovaya, E., Metelskaya, V., Meshkov, A., & Drapkina, O. (2019). Associations of SNPs of the ADIPOQ Gene with Serum Adiponectin Levels, Unstable Angina, and Coronary Artery Disease. Biomolecules, 9(10), 537. https://doi.org/10.3390/biom9100537