A Novel Kunitzin-Like Trypsin Inhibitor Isolated from Defensive Skin Secretion of Odorrana versabilis

 , ,
, , 
Abstract
:
1. Introduction
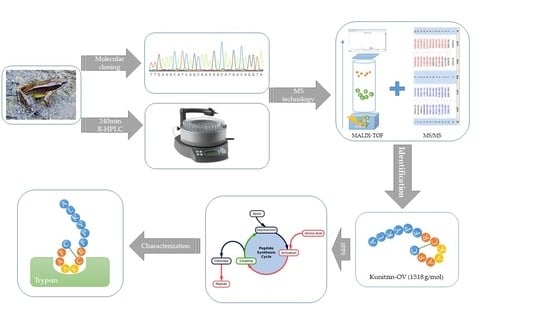
2. Materials and Methods
2.1. Specimen Biodata and Secretion Acquisition
2.2. Molecular Cloning of Kunitzin-OV Precursor-Encoding cDNA from the Skin Secretion-Derived cDNA Library of O. versabilis
2.3. Identification and Structural Analysis of Kunitzin-OV from Skin Secretion of O. veserbilis
2.4. Solid-Phase Peptide Synthesis of Kunitzin-OV and F9-Kunitzin-OV
2.5. Minimal Inhibitory Concentration Assay of Kunitzin-OV and Phe-Substituted Analogue
2.6. Haemolysis Assay
2.7. E. coli Cell Membrane Permeability Assay of Kunitzin-OV
2.8. Trypsin/Chymotrypsin Inhibition Assay
2.9. Trypsin Cleavage of Kunitzin-OV
3. Results
3.1. Molecular Cloning and Sequencing Analysis of Kunitzin-OV
3.2. Identification and Structural Characterisation of Kunitzin-OV
3.3. Synthesis of Kunitzin-OV and Phe-Substituted Variant
3.4. Antimicrobial/Haemolytic Assay of Kunitzin-OV and F9-Substituted Analogue
3.5. E. coli Cell Membrane Permeability of Kunitzin-OV
3.6. Trypsin/Chymotrypsin Inhibitory Activity of Kunizin-OV and Phe-Substituted Analogue
3.7. Trypsin Cleavage Assay of Kunitzin-OV
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Krowarsch, D.; Cierpicki, T.; Jelen, F.; Otlewski, J. Canonical Protein Inhibitors of Serine Proteases. Cell. Mol. Life Sci. 2003, 60, 2427–2444. [Google Scholar] [CrossRef] [PubMed]
- Habib, H.; Fazili, K.M. Plant Protease Inhibitors: A Defense Strategy in Plants. Biotechnol. Mol. Biol. Rev. 2007, 2, 68–85. [Google Scholar]
- Conlon, J.M.; Kim, J.B. A Protease Inhibitor of the Kunitz Family from Skin Secretions of the Tomato Frog, Dyscophus Guineti (Microhylidae). Biochem. Biophys. Res. Commun. 2000, 279, 961–964. [Google Scholar] [CrossRef] [PubMed]
- Ascenzi, P.; Bocedi, A.; Bolognesi, M.; Spallarossa, A.; Coletta, M.; Cristofaro, R.; Menegatti, E. The Bovine Basic Pancreatic Trypsin Inhibitor (Kunitz Inhibitor): A Milestone Protein. Curr. Protein Peptide Sci. 2003, 4, 231–251. [Google Scholar] [CrossRef]
- Hsieh, C.; Hernández-Ledesma, B.; Jeong, H.J.; Park, J.H.; Ben, O. Complementary Roles in Cancer Prevention: Protease Inhibitor Makes the Cancer Preventive Peptide Lunasin Bioavailable. PLoS ONE 2010, 5, e8890. [Google Scholar] [CrossRef] [PubMed]
- Cristina Oliveira de Lima, V.; Piuvezam, G.; Leal Lima Maciel, B.; Heloneida de Araújo Morais, A. Trypsin Inhibitors: Promising Candidate Satietogenic Proteins as Complementary Treatment for Obesity and Metabolic Disorders? J. Enzyme Inhib. Med. Chem. 2019, 34, 405–419. [Google Scholar] [CrossRef] [PubMed]
- Junker, Y.; Zeissig, S.; Kim, S.; Barisani, D.; Wieser, H.; Leffler, D.A.; Zevallos, V.; Libermann, T.A.; Dillon, S.; Freitag, T.L. Wheat Amylase Trypsin Inhibitors Drive Intestinal Inflammation Via Activation of Toll-Like Receptor 4. J. Exp. Med. 2012, 209, 2395–2408. [Google Scholar] [CrossRef] [PubMed]
- Całkosiński, I.; Zasadowski, A.; Bronowicka-Szydełko, A.; Dzierzba, K.; Seweryn, E.; Dobrzyński, M.; Gamian, A. Amphibian Skin Secretions as a New Source of Antibiotics and Biologically Active Substances. Postepy higieny i medycyny doswiadczalnej (Online) 2009, 63, 537–548. [Google Scholar] [PubMed]
- Proaño-Bolaños, C.; Li, R.; Zhou, M.; Wang, L.; Xi, X.; Tapia, E.E.; Coloma, L.A.; Chen, T.; Shaw, C. Novel Kazal-Type Proteinase Inhibitors from the Skin Secretion of the Splendid Leaf Frog, Cruziohyla Calcarifer. EuPA Open Proteom. 2017, 15, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Wang, H.; Jiang, Y.; Yu, Y.; Wang, L.; Zhou, M.; Zhang, Y.; Chen, T.; Shaw, C. A Novel Kazal-Type Trypsin Inhibitor from the Skin Secretion of the Central American Red-Eyed Leaf Frog, Agalychnis Callidryas. Biochimie 2012, 94, 1376–1381. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Wang, L.; Chen, T.; Walker, B.; Zhou, M.; Sui, D.; Conlon, J.M.; Shaw, C. Identification and Molecular Cloning of a Novel Amphibian Bowman Birk-Type Trypsin Inhibitor from the Skin of the Hejiang Odorous Frog; Odorrana Hejiangensis. Peptides 2012, 33, 245–250. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Wang, H.; Shen, Y.; Wang, L.; Zhou, M.; Chen, T.; Shaw, C. Kunitzins: Prototypes of a New Class of Protease Inhibitor from the Skin Secretions of European and Asian Frogs. Biochem. Biophys. Res. Commun. 2016, 477, 302–309. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.; Tikhonova, I.G.; Jewhurst, H.L.; Drysdale, O.C.; Dvořák, J.; Robinson, M.W.; Cwiklinski, K.; Dalton, J.P. Unexpected Activity of a Novel Kunitz-Type Inhibitor: Inhibition of Cysteine Proteases but Not Serine Proteases. J. Biol. Chem. 2016, 291, 19220–19234. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wu, J.; Wang, Y.; Xu, X.; Liu, T.; Lai, R.; Zhu, H. A Small Trypsin Inhibitor from the Frog of Odorrana Grahami. Biochimie 2008, 90, 1356–1361. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Long, Q.; Xu, Y.; Guo, S.; Chen, T.; Wang, L.; Zhou, M.; Zhang, Y.; Shaw, C.; Walker, B. A Structural and Functional Analogue of a Bowman Birk-Type Protease Inhibitor from Odorrana Schmackeri. Biosci. Rep. 2017. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Wu, D.; Xi, X.; Wu, Y.; Ma, C.; Zhou, M.; Wang, L.; Yang, M.; Chen, T.; Shaw, C. Identification and Characterisation of the Antimicrobial Peptide, Phylloseptin-PT, from the Skin Secretion of Phyllomedusa Tarsius, and Comparison of Activity with Designed, Cationicity-Enhanced Analogues and Diastereomers. Molecules 2016, 21, 1667. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Chen, D.; Wang, L.; Lin, C.; Ma, C.; Xi, X.; Chen, T.; Shaw, C.; Zhou, M. Dermaseptin-PH: A Novel Peptide with Antimicrobial and Anticancer Activities from the Skin Secretion of the South American Orange-Legged Leaf Frog, Pithecopus (Phyllomedusa) Hypochondrialis. Molecules 2017, 22, 1805. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Lee, W.; Zhang, Y. Extremely Abundant Antimicrobial Peptides Existed in the Skins of Nine Kinds of Chinese Odorous Frogs. J. Proteome Res. 2011, 11, 306–319. [Google Scholar] [CrossRef]
- Bahar, A.A.; Ren, D. Antimicrobial Peptides. Pharmaceuticals 2013, 6, 1543–1575. [Google Scholar] [CrossRef] [Green Version]
- Sonohara, R.; Muramatsu, N.; Ohshima, H.; Kondo, T. Difference in Surface Properties between Escherichia Coli and Staphylococcus Aureus as Revealed by Electrophoretic Mobility Measurements. Biophys. Chem. 1995, 55, 273–277. [Google Scholar] [CrossRef]
- Wang, K.; Jia, F.; Dang, W.; Zhao, Y.; Zhu, R.; Sun, M.; Qiu, S.; An, X.; Ma, Z.; Zhu, Y.; et al. Antifungal Effect and Action Mechanism of Antimicrobial Peptide polybia-cp. J. Peptide Sci. 2016, 22, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.; Sun, T.; Hung, W.; Huang, H.W. Process of Inducing Pores in Membranes by Melittin. Proc. Natl. Acad. Sci. USA 2013, 110, 14243–14248. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, L.; Dufton, M.J. Evolutionary Trace Analysis of the Kunitz/BPTI Family of Proteins: Functional Divergence may have been Based on Conformational Adjustment. J. Mol. Biol. 1999, 285, 1589–1607. [Google Scholar] [CrossRef] [PubMed]
- Otlewski, J.; Jaskólski, M.; Buczek, O.; Cierpicki, T.; Czapińska, H.; Krowarsch, D.; Smalas, A.O.; Stachowiak, D.; Szpineta, A.; Dadlez, M. Structure-Function Relationship of Serine Protease-Protein Inhibitor Interaction. Acta Biochim. Pol. 2001, 48, 419–428. [Google Scholar] [PubMed]
- Perona, J.J.; Craik, C.S. Evolutionary Divergence of Substrate Specificity within the Chymotrypsin-Like Serine Protease Fold. J. Biol. Chem. 1997, 272, 29987–29990. [Google Scholar] [CrossRef] [PubMed]
- Derache, C.; Epinette, C.; Roussel, A.; Gabant, G.; Cadene, M.; Korkmaz, B.; Gauthier, F.; Kellenberger, C. Crystal Structure of Greglin, a Novel Non-Classical Kazal Inhibitor, in Complex with Subtilisin. FEBS J. 2012, 279, 4466–4478. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sequence | Net Charge | GRAVY 1 | |
|---|---|---|---|
| Kunitzin-OV | ALKYPFRCKAAFC | +3 | 0.354 |
| F9-Kunitzin-OV | ALKYPFRCFAAFC | +2 | 0.869 |
| Peptides | Sequence | Minimum Inhibitory Concentration (µM) | ||
|---|---|---|---|---|
| S. aureus | E. coli | C. albicans | ||
| Kunitzin-OV | ALKYPFRCKAAFC | 512 | 512 | 512 |
| F9-Kunitzin-OV | ALKYPFRCFAAFC | 512 | 512 | 512 |
| Name | Sequence Length | Reactive Site | Source |
|---|---|---|---|
| Kunitzin-OV | 13 | -F R C K A A F- | This study |
| VKTO1_HAPHA | 55 | -G R C K A S F- | UniProtKB-D2Y2Q6 |
| VKTCT_OPHHA | 58 | -G F C K A Y I- | UniProtKB-B6RLX2 |
| BPT1_BOVIN | 58 | -G P C K A R I- | UniProtKB-P00974 |
| CSTI_BOMMO | 55 | -G P C K G S F- | UniProtKB-P81902 |
| SPIT2_HUMAN domain 2 | 51 | -G P C R A S F- | UniProtKB-O43291 |
| AMBP_HUMAN domain 2 | 51 | -G P C R A F I- | UniProtKB-P02760 |
| VKT_OXYSC | 51 | -G P C R A A I- | UniProtKB-B7S4N9 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, Y.; Shi, D.; Ying, Y.; Xi, X.; Chen, X.; Wang, L.; Zhou, M.; Wu, Q.; Ma, C.; Chen, T. A Novel Kunitzin-Like Trypsin Inhibitor Isolated from Defensive Skin Secretion of Odorrana versabilis. Biomolecules 2019, 9, 254. https://doi.org/10.3390/biom9070254
Dong Y, Shi D, Ying Y, Xi X, Chen X, Wang L, Zhou M, Wu Q, Ma C, Chen T. A Novel Kunitzin-Like Trypsin Inhibitor Isolated from Defensive Skin Secretion of Odorrana versabilis. Biomolecules. 2019; 9(7):254. https://doi.org/10.3390/biom9070254
Chicago/Turabian StyleDong, Yanjing, Daning Shi, Yuan Ying, Xinping Xi, Xiaoling Chen, Lei Wang, Mei Zhou, Qinan Wu, Chengbang Ma, and Tianbao Chen. 2019. "A Novel Kunitzin-Like Trypsin Inhibitor Isolated from Defensive Skin Secretion of Odorrana versabilis" Biomolecules 9, no. 7: 254. https://doi.org/10.3390/biom9070254
APA StyleDong, Y., Shi, D., Ying, Y., Xi, X., Chen, X., Wang, L., Zhou, M., Wu, Q., Ma, C., & Chen, T. (2019). A Novel Kunitzin-Like Trypsin Inhibitor Isolated from Defensive Skin Secretion of Odorrana versabilis. Biomolecules, 9(7), 254. https://doi.org/10.3390/biom9070254