Detection and Identification of Allergens from Canadian Mustard Varieties of Sinapis alba and Brassica juncea

Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Mustard Seed Flours
2.2. Optimization of Mustard Seed Protein Extraction
2.3. Protein Electrophoresis
2.4. Immunoblotting
2.5. Indirect ELISA
2.6. Identification of Protein Bands as Allergens by LC-MS/MS
2.7. Protein Sequence Comparisons with Known and Putative Allergens
3. Results and Discussion
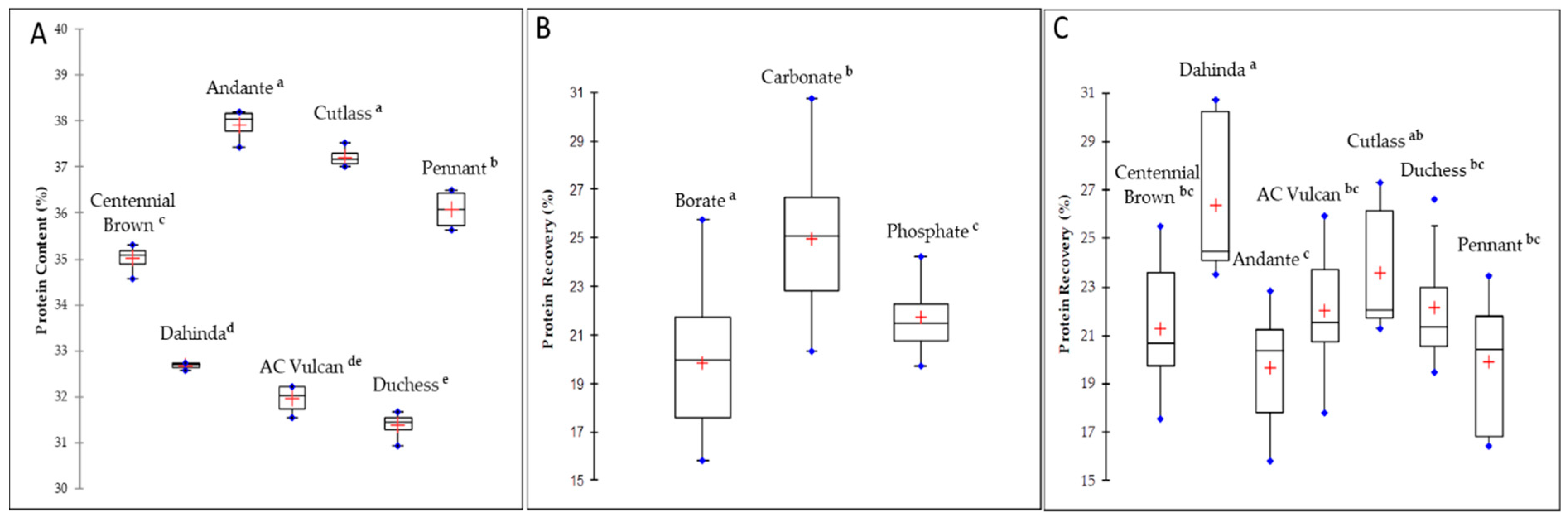
3.1. Protein Content and Extractability from Different Mustard Varieties
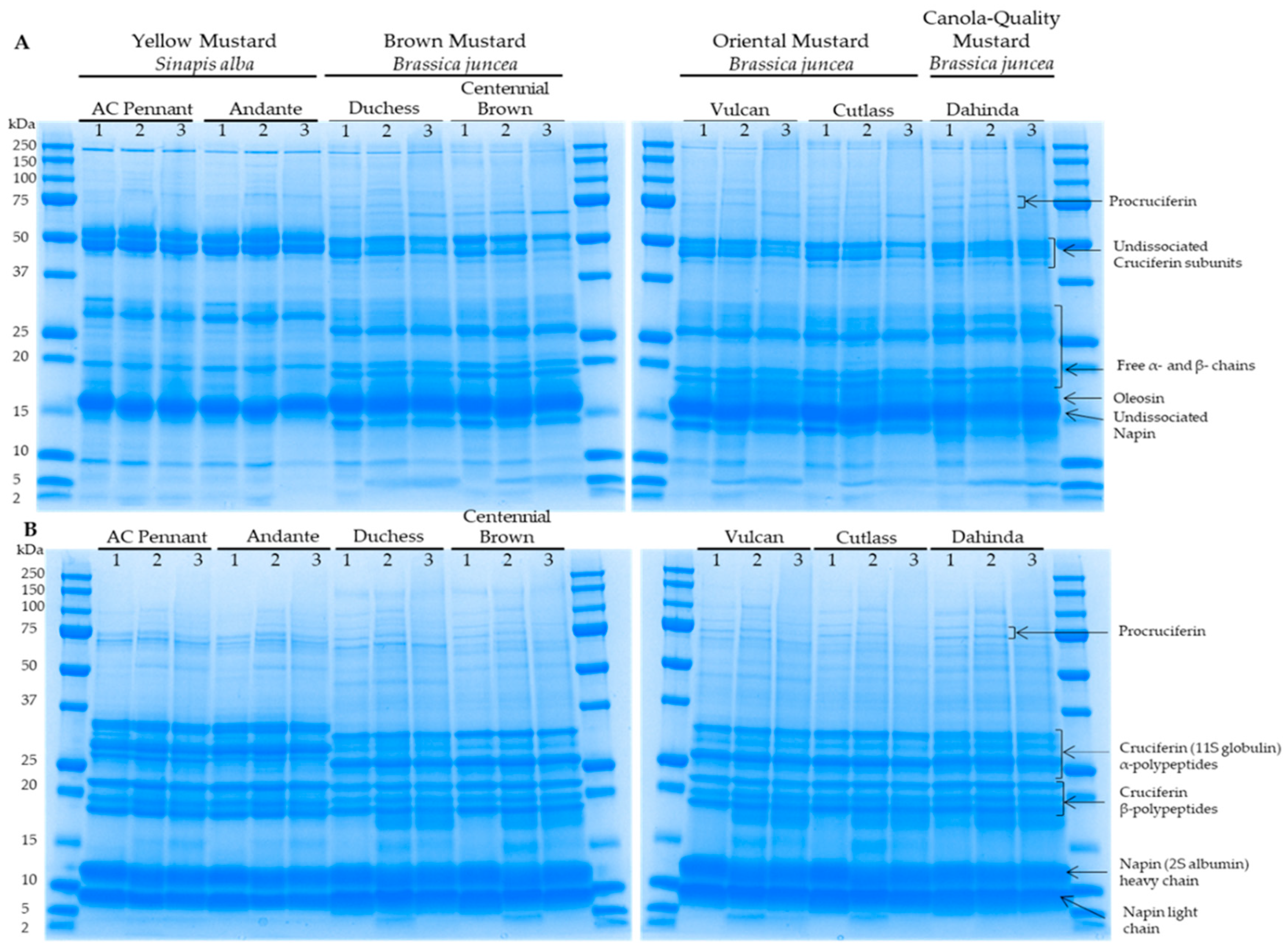
3.2. Protein Electrophoretic Profiles of Differents Mustard Varieties as a Function of Extraction Buffer
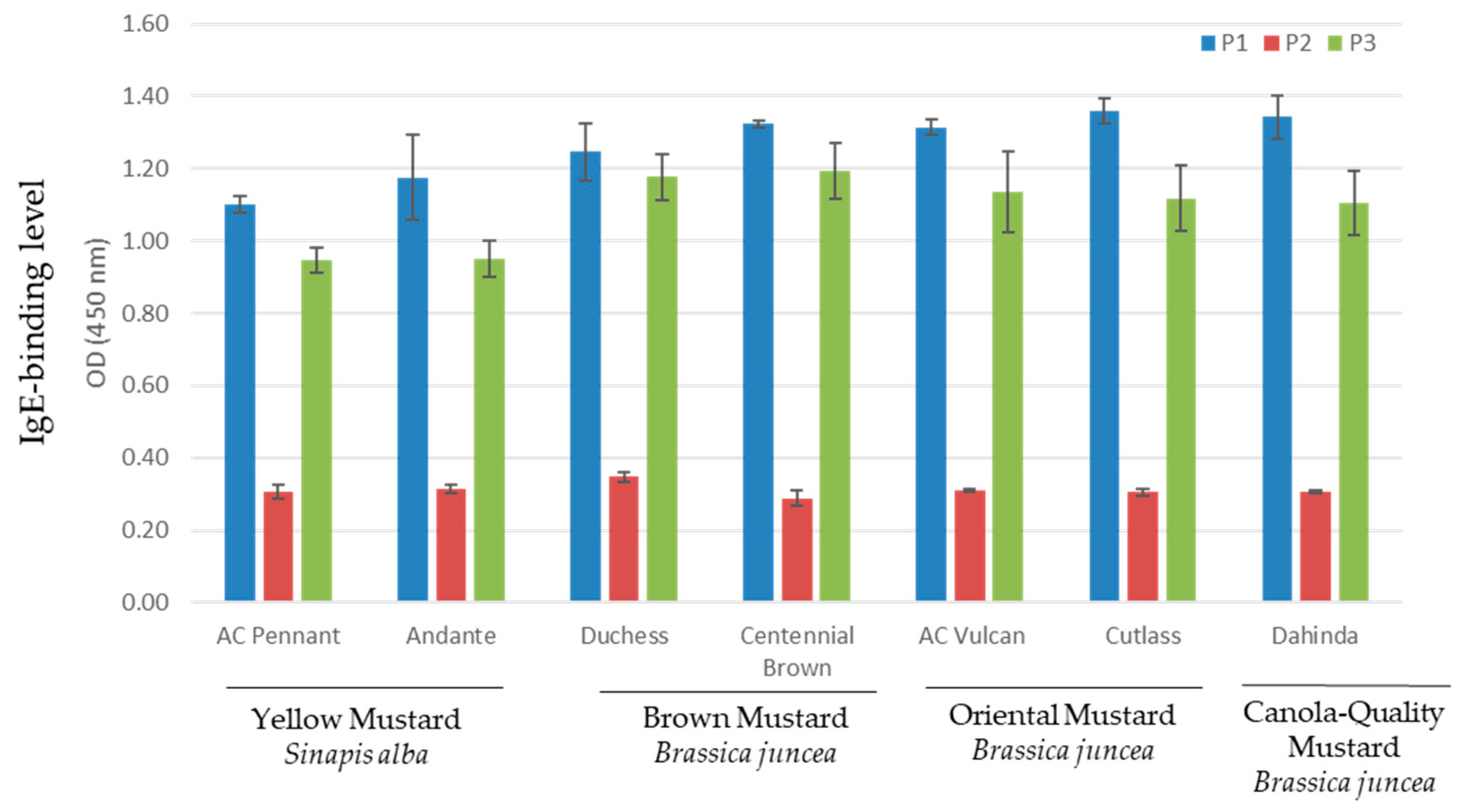
3.3. Indirect ELISA for Serum IgE Response to Mustard Varieties
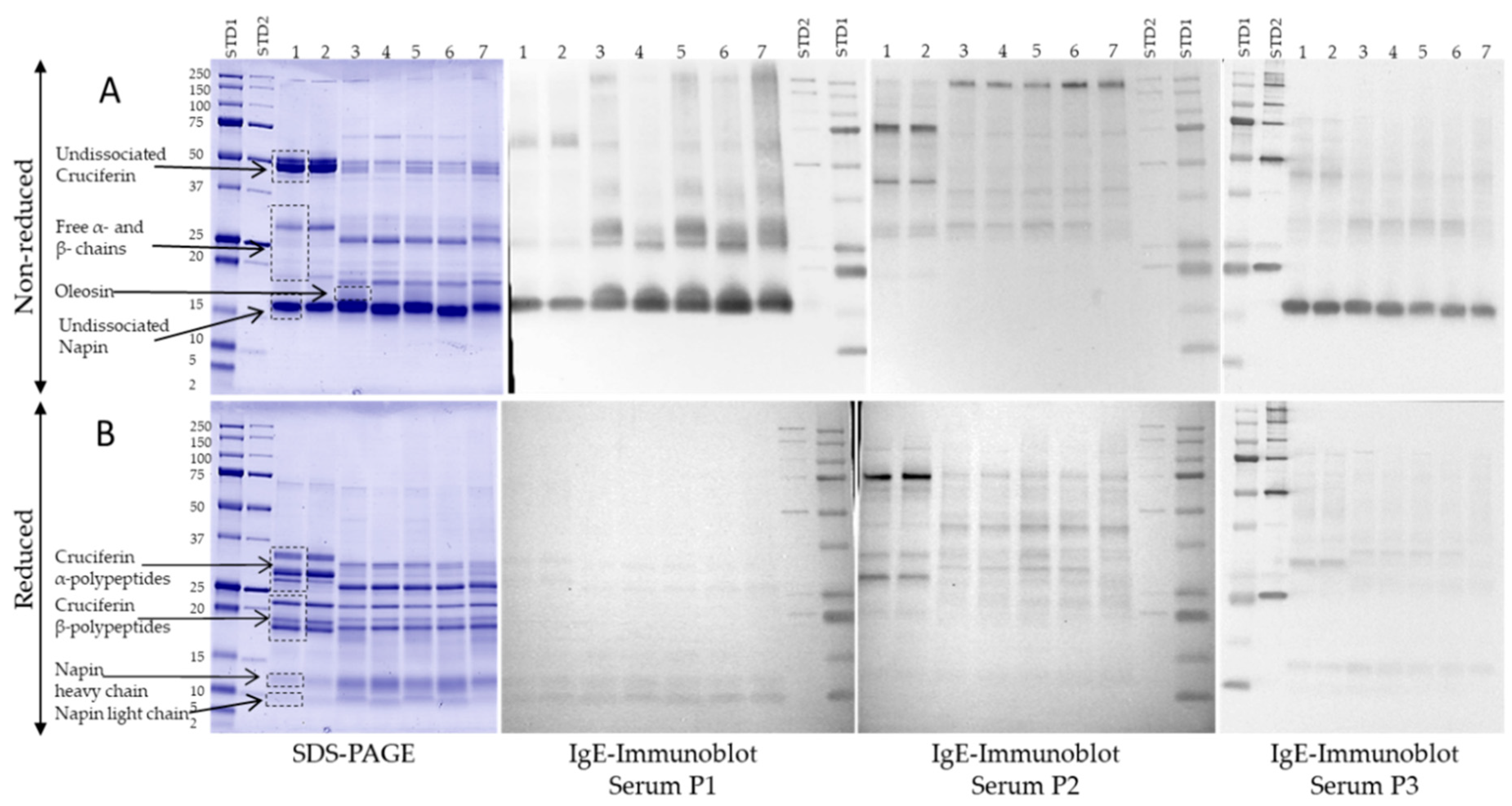
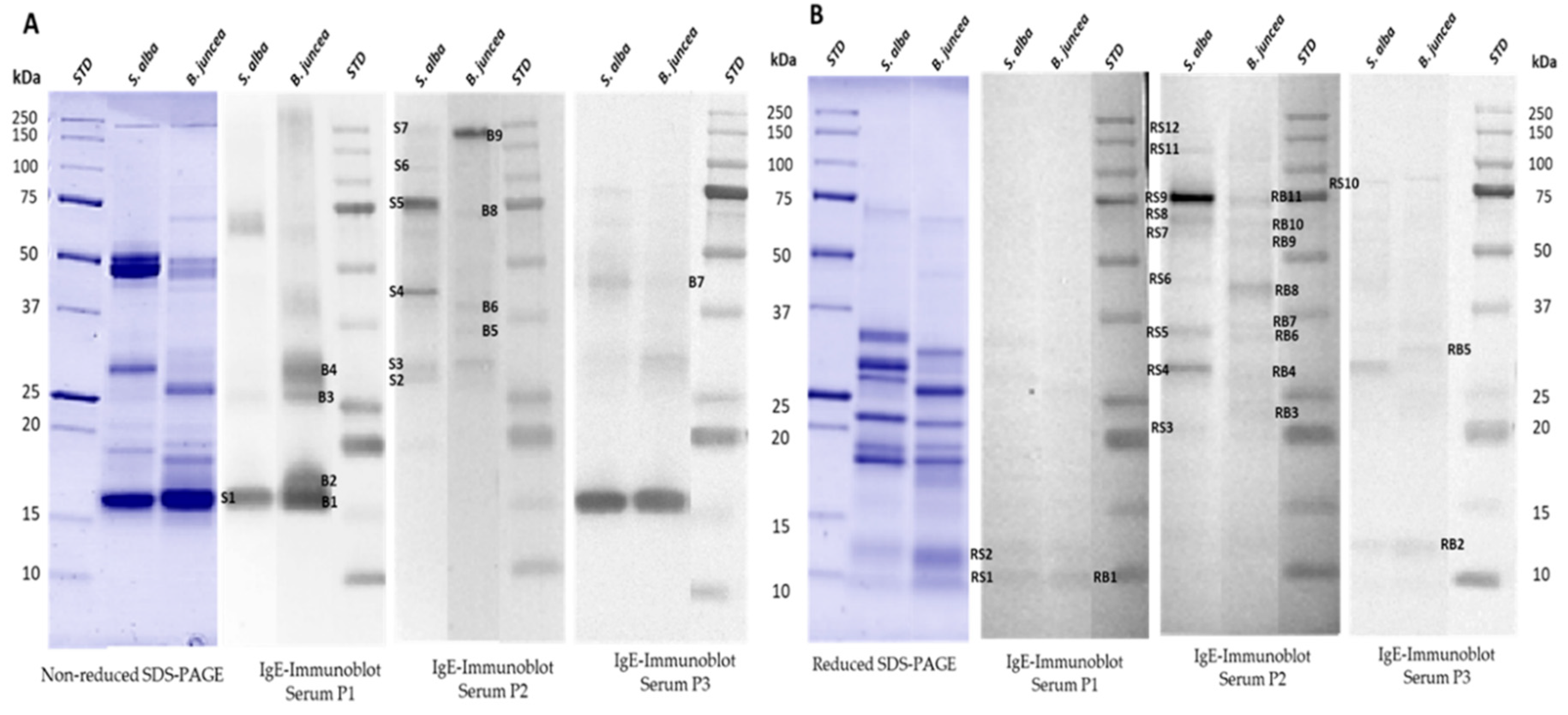
3.4. IgE-Binding Profiles of Select Canadian Mustard Varieties
3.5. Identification of Mustard IgE-Binding Proteins by Mass Spectroscopy
3.6. Bioinformatic Assessment of Potential Cross-Reactivity of Identified Mustard IgE-Binding Proteins with Known Allergens
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Caballero, T.; San-Martín, M.S.; Padial, M.A.; Contreras, J.; Cabañas, R.; Barranco, P.; López-Serrano, M.C. Clinical characteristics of patients with mustard hypersensitivity. Ann. Allergy Asthma Immunol. 2002, 89, 166–171. [Google Scholar] [CrossRef]
- Sharma, A.; Verma, A.K.; Gupta, R.K.; Neelabh; Dwivedi, P.D. A comprehensive review on mustard-induced allergy and implications for human health. Clin. Rev. Allergy Immunol. 2017, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Monneret-Vautrin, D.-A. Epidemiology of food allergies and relative prevalence of trophallergens. Cahiers Nutr. Diét. 2001, 36, 247–252. [Google Scholar]
- EuropeanFoodSafetyAuthority. Opinion of the scientific panel on dietetic products, nutrition and allergies [nda] on a request from the commission relating to the evaluation of allergenic foods for labelling purposes. EFSA J. 2004, 2, 32. [Google Scholar]
- Rancé, F.; Dutau, G.; Abbal M, M. Mustard allergy in children. Allergy 2000, 55, 496–500. [Google Scholar] [CrossRef] [PubMed]
- Rancé, F.; Abbal, M.; Dutau, G. Mustard allergy in children. Pediatr. Pulmonol. 2001, 32, 44–45. [Google Scholar] [CrossRef]
- Morisset, M.; Moneret-Vautrin, D.A.; Maadi, F.; Frémont, S.; Guénard, L.; Croizier, A.; Kanny, G. Prospective study of mustard allergy: First study with double-blind placebo-controlled food challenge trials (24 cases). Allergy 2003, 58, 295–299. [Google Scholar] [CrossRef]
- Figueroa, J.; Blanco, C.; Dumpiérrez, A.G.; Almeida, L.; Ortega, N.; Castillo, R.; Navarro, L.; Pérez, E.; Gallego, M.D.; Carrillo, T. Mustard allergy confirmed by double-blind placebo-controlled food challenges: Clinical features and cross-reactivity with mugwort pollen and plant-derived foods. Allergy Eur. J. Allergy Clin. Immunol. 2005, 60, 48–55. [Google Scholar] [CrossRef]
- World Atlas HomePage. Available online: https://www.worldatlas.com/articles/the-world-s-top-mustard-seed-producing-countries.html (accessed on 16 July 2019).
- Marambe, H.K.; McIntosh, T.C.; Cheng, B.; Wanasundara, J.P.D. Quantification of major 2s allergen protein of yellow mustard using anti-sin a 1 epitope antibody. Food Control 2014, 44, 233–241. [Google Scholar] [CrossRef]
- Canadian Grain Commission. Available online: https://www.grainscanada.gc.ca/en/grain-research/export-quality/oilseeds/mustard/2016/mustard-quality-report-16-1.html (accessed on 17 July 2019).
- Menéndez-Arias, L.; Monsalve, R.I.; Gavilanes, J.; Rodríguez, R. Molecular and spectroscopic characterisation of a low molecular weight seed storage protein from yellow mustard (sinapis alba l.). Int. J. Biochem. 1987, 19, 899–907. [Google Scholar] [CrossRef]
- Menéndez-Arias, L.; Moneo, I.; DomÍNguez, J.; RodrÍGuez, R. Primary structure of the major allergen of yellow mustard (sinapis alba l.) seed, sin a i. Eur. J. Biochem. 1988, 177, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Menéndez-Arias, L.; Domínguez, J.; Moneo, I.; Rodríguez, R. Epitope mapping of the major allergen from yellow mustard seeds, sin a i. Mol. Immunol. 1990, 27, 143–150. [Google Scholar] [CrossRef]
- Dominguez, J.; Cuevas, M.; Urena, V.; Munoz, T.; Moneo, I. Purification and characterization of an allergen of mustard seed. Ann. Allergy 1990, 64, 352–357. [Google Scholar] [PubMed]
- Delapena, M.A.G.; Villalba, M.; Garcialopez, J.L.; Rodriguez, R. Cloning and expression of the major allergen from yellow mustard seeds, sin a i. Biochem. Biophys. Res. Commun. 1993, 190, 648–653. [Google Scholar] [CrossRef]
- De La Peña, M.A.G.; Monsalve, R.I.; Batanero, E.; Villalba, M.; Rodríguez, R. Expression in escherichia coli of sin a 1, the major allergen from mustard. Eur. J. Biochem. 1996, 237, 827–832. [Google Scholar] [CrossRef] [PubMed]
- Shim, Y.-Y.; Wanasundara, J.P.D. Quantitative detection of allergenic protein sin a 1 from yellow mustard (sinapis alba l.) seeds using enzyme-linked immunosorbent assay. J. Agric. Food Chem. 2008, 56, 1184–1192. [Google Scholar] [CrossRef] [PubMed]
- Palomares. Isolation and identification of an 11s globulin as a new major allergen in mustard seeds. Ann. Allergy Asthma Immunol. 2005, 94, 586–592. [Google Scholar] [CrossRef]
- Palomares. Cloning, sequencing, and recombinant production of sin a 2, an allergenic 11s globulin from yellow mustard seeds. J. Allergy Clin. Immunol. 2007, 119, 1189–1196. [Google Scholar] [CrossRef]
- Sirvent, S.; Palomares, O.; Vereda, A.; Villalba, M.; Cuesta-Herranz, J.; Rodríguez, R. Nsltp and profilin are allergens in mustard seeds: Cloning, sequencing and recombinant production of sin a 3 and sin a 4. Clin. Exp. Allergy 2009, 39, 1929–1936. [Google Scholar] [CrossRef]
- Sirvent, S.; Palomares, O.; Cuesta-Herranz, J.; Villalba, M.; Rodríguez, R. Analysis of the structural and immunological stability of 2s albumin, nonspecific lipid transfer protein, and profilin allergens from mustard seeds. J. Agric. Food Chem. 2012, 60, 6011–6018. [Google Scholar] [CrossRef]
- Gonzalez de la Pena, M.; Menendez-Arias, L.; Monsalve, R.I.; Rodriguez, R. Isolation and characterization of a major allergen from oriental mustard seeds, braj i. Int. Arch. Allergy Appl. Immunol. 1991, 96, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Monsalve, R.I.; Gonzalez de la Pena, M.A.; Menendez-Arias, L.; Lopez-Otin, C.; Villalba, M.; Rodriguez, R. Characterization of a new oriental-mustard (brassica juncea) allergen, bra j ie: Detection of an allergenic epitope. Biochem. J. 1993, 293, 625–632. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage t4. Nature 1970, 227, 680. [Google Scholar] [CrossRef] [PubMed]
- Towbin, H.; Staehelin, T.; Gordon, J. Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: Procedure and some applications. Proc. Natl. Acad. Sci. USA 1979, 76, 4350–4354. [Google Scholar] [CrossRef] [PubMed]
- Rioux, G.; Majeau, N.; Leclerc, D. Mapping the surface-exposed regions of papaya mosaic virus nanoparticles. FEBS J. 2012, 279, 2004–2011. [Google Scholar] [CrossRef] [PubMed]
- Keller, A.; Nesvizhskii, A.I.; Kolker, E.; Aebersold, R. Empirical statistical model to estimate the accuracy of peptide identifications made by ms/ms and database search. Anal. Chem. 2002, 74, 5383–5392. [Google Scholar] [CrossRef] [PubMed]
- Nesvizhskii, A.I.; Keller, A.; Kolker, E.; Aebersold, R. A statistical model for identifying proteins by tandem mass spectrometry. Anal. Chem. 2003, 75, 4646–4658. [Google Scholar] [CrossRef] [PubMed]
- Goodman, R.E.; Ebisawa, M.; Ferreira, F.; Sampson, H.A.; van Ree, R.; Vieths, S.; Baumert, J.L.; Bohle, B.; Lalithambika, S.; Wise, J.; et al. Allergenonline: A peer-reviewed, curated allergen database to assess novel food proteins for potential cross-reactivity. Mol. Nutr. Food Res. 2016, 60, 1183–1198. [Google Scholar] [CrossRef] [PubMed]
- Alimentarius, C. Alinorm 03/34: Joint fao/who food standard programme, codex alimentarius commission. In Proceedings of the Twenty-Fifth Session, Rome, Italy, 30 June–5 July 2003; pp. 47–60, Appendix III, Guideline for the conduct of food safety assessment of foods derived from recombinant-DNA plants and Appendix IV, Annex on the assessment of possible allergenicity. [Google Scholar]
- Henikoff, S.; Henikoff, J.G. Amino acid substitution matrices from protein blocks. Proc. Natl. Acad. Sci. USA 1992, 89, 10915–10919. [Google Scholar] [CrossRef]
- Wanasundara, J.P.D.; Abeysekara, S.J.; McIntosh, T.C.; Falk, K.C. Solubility differences of major storage proteins of brassicaceae oilseeds. J. Am. Oil Chem. Soc. 2012, 89, 869–881. [Google Scholar] [CrossRef]
- L’Hocine, L.; Pitre, M. Quantitative and qualitative optimization of allergen extraction from peanut and selected tree nuts. Part 2. Optimization of buffer and ionic strength using a full factorial experimental design. Food Chem. 2016, 194, 820–827. [Google Scholar] [CrossRef] [PubMed]
- Wanasundara, J.P.D. Proteins of brassicaceae oilseeds and their potential as a plant protein source. Crit. Rev. Food Sci. Nutr. 2011, 51, 635–677. [Google Scholar] [CrossRef]
- Schwenke, K.D.; Raab, B.; Linow, K.J.; Pahtz, W.; Uhlig, J. Isolation of the 12 s globulin from rapeseed (brassica napus l.) and characterization as a “neutral” protein on seed proteins. Part 13. Food/Nahrung 1981, 25, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Potts, D.A.; Rakow, G.R.; Males, D.R. Canola-quality brassica juncea, a new oilseed crop for the canadian prairies. In Proceedings of the 10th International Rapeseed Congress, Canberra, Australia, 1999. Contribution No 70. [Google Scholar]
- Malabat, C.; Atterby, H.; Chaudhry, Q.; Renard, M.; Guéguen, J. Genetic variability of rapeseed protein composition. In 11th International Rapeseed Congress; Royal Veterinary and Agricultural University: Frederiksberg, Denmark, 2003. [Google Scholar]
- Perera, S.P.; McIntosh, T.C.; Wanasundara, J.P.D. Structural properties of cruciferin and napin of brassica napus (canola) show distinct responses to changes in ph and temperature. Plants 2016, 5, 36. [Google Scholar] [CrossRef]
- Aluko, R.; McIntosh, T. Electrophoretic and functional properties of mustard seed meals and protein concentrates. J. Am. Oil Chem. Soc. 2004, 81, 679–683. [Google Scholar] [CrossRef]
- Tzen, J.T.C.; Cao, Y.; Laurent, P.; Ratnayake, C.; Huang, A.H.C. Lipids, proteins, and structure of seed oil bodies from diverse species. Plant physiol. 1993, 101, 267–276. [Google Scholar] [CrossRef]
- Marambe, H.K.; McIntosh, T.C.; Cheng, B.; Wanasundara, J.P.D. Structural stability and sin a 1 anti-epitope antibody binding ability of yellow mustard (sinapis alba l.) napin during industrial-scale myrosinase inactivation process. Food Funct. 2015, 6, 2384–2395. [Google Scholar] [CrossRef]
- Vereda, A.; Sirvent, S.; Villalba, M.; Rodríguez, R.; Cuesta-Herranz, J.; Palomares, O. Improvement of mustard (sinapis alba) allergy diagnosis and management by linking clinical features and component-resolved approaches. J. Allergy Clin. Immunol. 2011, 127, 1304–1307. [Google Scholar] [CrossRef] [PubMed]
- Hummel, M.; Wigger, T.; Brockmeyer, J. Characterization of mustard 2s albumin allergens by bottom-up, middle-down, and top-down proteomics: A consensus set of isoforms of sin a 1. J. Proteome Res. 2015, 14, 1547–1556. [Google Scholar] [CrossRef] [PubMed]
- Abeysekara, S. Solubility, In Vitro Digestibility and Allergenicity of Brassica Juncea, Brassica Napus and Sinapis Alba Proteins. Master’s Thesis, University of Saskatchewan, Saskatoon, SK, Canada, 2011. [Google Scholar]
- Dalgalarrondo, M.; Robin, J.-M.; Azanza, J.-L. Subunit composition of the globulin fraction of rapeseed (brassica napus l.). Plant Sci. 1986, 43, 115–124. [Google Scholar] [CrossRef]
- Robin, J.M.; Inquello, V.; Mimouni, B.; Azanza, J.L. Relationship between immunological properties and structural model of 11s rapeseed globulin. Phytochemistry 1991, 30, 3511–3513. [Google Scholar] [CrossRef]
- Leduc, V.; Moneret-Vautrin, D.A.; Tzen, J.T.C.; Morisset, M.; Guerin, L.; Kanny, G. Identification of oleosins as major allergens in sesame seed allergic patients. Allergy 2006, 61, 349–356. [Google Scholar] [CrossRef] [PubMed]
- Pons, L.; Olszewski, A.; Guéant, J.-L. Characterization of the oligomeric behavior of a 16.5 kda peanut oleosin by chromatography and electrophoresis of the iodinated form. J. Chromatogr. 1998, 706, 131–140. [Google Scholar] [CrossRef]
- Pons, L.; Chéry, C.; Mrabet, N.; Schohn, H.; Lapicque, F.; Guéant, J.-L. Purification and cloning of two high molecular mass isoforms of peanut seed oleosin encoded by cdnas of equal sizes. Plant Physiol. Biochem. 2005, 43, 659–668. [Google Scholar] [CrossRef]
- Kobayashi, S.; Katsuyama, S.; Wagatsuma, T.; Okada, S.; Tanabe, S. Identification of a new ige-binding epitope of peanut oleosin that cross-reacts with buckwheat. Biosci. Biotechnol. Biochem. 2012, 76, 1182–1188. [Google Scholar] [CrossRef]
- Akkerdaas, J.H.; Schocker, F.; Vieths, S.; Versteeg, S.; Zuidmeer, L.; Hefle, S.L.; Aalberse, R.C.; Richter, K.; Ferreira, F.; van Ree, R. Cloning of oleosin, a putative new hazelnut allergen, using a hazelnut cdna library. Mol. Nutr. Food Res. 2006, 50, 18–23. [Google Scholar] [CrossRef]
- King, T.P.; Hoffman, D.; Lowenstein, H.; Marsh, D.G.; Platts-Mills, T.A.E.; Thomas, W. Allergen nomenclature. Int. Arch. Allergy Immunol. 1994, 105, 224–233. [Google Scholar] [CrossRef]
- Falk, A.; Rask, L. Expression of a zeatin-o-glucoside-degrading [beta]-glucosidase in Brassica napus. Plant Physiol. 1995, 108, 1369–1377. [Google Scholar] [CrossRef] [PubMed]
- Šotkovský, P.; Sklenář, J.; Halada, P.; Cinová, J.; Šetinová, I.; Kainarová, A.; Goliáš, J.; Pavlásková, K.; Honzová, S.; Tučková, L. A new approach to the isolation and characterization of wheat flour allergens. Clin. Exp. Allergy 2011, 41, 1031–1043. [Google Scholar] [CrossRef] [PubMed]
- Zhang, E.; Brewer, J.M.; Minor, W.; Carreira, L.A.; Lebioda, L. Mechanism of enolase: The crystal structure of asymmetric dimer enolase−2-phospho-d-glycerate/enolase−phosphoenolpyruvate at 2.0 å resolution. Biochemistry 1997, 36, 12526–12534. [Google Scholar] [CrossRef] [PubMed]
- Simon-Nobbe, B.; Probst, G.; Kajava, A.V.; Oberkofler, H.; Susani, M.; Crameri, R.; Ferreira, F.; Ebner, C.; Breitenbach, M. Ige-binding epitopes of enolases, a class of highly conserved fungal allergens. J. Allergy Clin. Immunol. 2000, 106, 887–895. [Google Scholar] [CrossRef] [PubMed]
- Wagner, S.; Breiteneder, H.; Simon-Nobbe, B.; Susani, M.; Krebitz, M.; Niggemann, B.; Brehler, R.; Scheiner, O.; Hoffmann-Sommergruber, K. Hev b 9, an enolase and a new cross-reactive allergen from hevea latex and molds. Eur. J. Biochem. 2000, 267, 7006–7014. [Google Scholar] [CrossRef] [PubMed]
- Lai, H.Y.; Tam, M.F.; Tang, R.B.; Chou, H.; Chang, C.Y.; Tsai, J.J.; Shen, H.D. Cdna cloning and immunological characterization of a newly identified enolase allergen from penicillium citrinum and aspergillus fumigatus. Int. Arch. Allergy Immunol. 2002, 127, 181–190. [Google Scholar] [CrossRef]
- Satinover, S.M.; Reefer, A.J.; Pomes, A.; Chapman, M.D.; Platts-Mills, T.A.E.; Woodfolk, J.A. Specific ige and igg antibody-binding patterns to recombinant cockroach allergens. J. Allergy Clin. Immunol. 2005, 115, 803–809. [Google Scholar] [CrossRef]
- Shankar, J.; Gupta, P.D.; Sridhara, S.; Singh, B.P.; Gaur, S.N.; Arora, N. Immunobiochemical analysis of cross-reactive glutathione-s-transferase allergen from different fungal sources. Immunol. Investig. 2005, 34, 37–51. [Google Scholar] [CrossRef]
- Pahr, S.; Constantin, C.; Mari, A.; Scheiblhofer, S.; Thalhamer, J.; Ebner, C.; Vrtala, S.; Mittermann, I.; Valenta, R. Molecular characterization of wheat allergens specifically recognized by patients suffering from wheat-induced respiratory allergy. Clin. Exp. Allergy 2012, 42, 597–609. [Google Scholar] [CrossRef]
- Aalberse, R.C. Structural biology of allergens. J. Allergy Clin. Immunol. 2000, 106, 228–238. [Google Scholar] [CrossRef]
- Hileman, R.E.; Silvanovich, A.; Goodman, R.E.; Rice, E.A.; Holleschak, G.; Astwood, J.D.; Hefle, S.L. Bioinformatic methods for allergenicity assessment using a comprehensive allergen database. Int. Arch. Allergy Immunol. 2002, 128, 280–291. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Band Intensity (1 × 106) | ||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sinapis alba | Brassica juncea | |||||||||||||||||||||||
| MW a | AC Pennant | Andante | MW a | Duchess | Centennial Brown | Vulcan | Cutlass | Dahinda | ||||||||||||||||
| Band Name | kDa | 1 | 2 | 3 | 1 | 2 | 3 | Band Name | kDa | 1 | 2 | 3 | 1 | 2 | 3 | 1 | 2 | 3 | 1 | 2 | 3 | 1 | 2 | 3 |
| Sin a 1 LC b | 9.5 | 24.1 | 18.9 | 19.1 | 23.4 | 18.3 | 18.3 | Bra j 1 LC b | 9.5 | 23.7 | 15.0 | 19.6 | 23.7 | 13.0 | 15.7 | 22.4 | 18.1 | 19.9 | 27.2 | 16.7 | 21.2 | 26.4 | 23.1 | 22.5 |
| Sin a 1 HC c | 12 | 25.0 | 19.2 | 19.4 | 11.8 | 19.4 | 19.1 | Bra j 1 HC c | 12 | 19.2 | 11.1 | 14.1 | 17.7 | 10.0 | 18.0 | 13.9 | 12.9 | 14.7 | 19.6 | 10.8 | 15.0 | 16.1 | 13.9 | 14.6 |
| Unknown | 15.1 | 0.6 | 2.8 | 0.8 | 2.1 | Unknown | 14.6 | 2.4 | 1.2 | 3.3 | 1.1 | 2.8 | 1.1 | 3.7 | 0.8 | |||||||||
| Cruciferin | Cruciferin | 16.8 | 7.1 | 6.8 | 5.5 | 5.4 | 7.3 | 8.2 | 6.8 | 6.0 | 3.5 | 10.5 | 9.9 | |||||||||||
| β-polypeptides | 18 | 13.3 | 14.8 | 15.0 | 11.8 | 14.2 | 10.6 | β-polypeptides | 18 | 11.3 | 10.3 | 8.7 | 12.7 | 11.8 | 10.9 | 13.9 | 12.3 | 9.7 | 14.8 | 12.5 | 11.8 | 13.7 | 14.3 | 13.3 |
| 11 S globulin | 19.1 | 5.9 | 11.6 | 9.6 | 6.4 | 12.0 | 5.1 | 11 S globulin | 19.1 | 5.7 | 6.7 | 5.2 | 5.7 | 6.4 | 5.3 | 6.6 | 7.9 | 5.9 | 5.7 | 6.5 | 4.7 | 6.7 | 8.2 | 7.0 |
| Sin a 2 | 21.6 | 8.8 | 12.6 | 10.5 | 9.0 | 12.5 | 7.8 | Sin a 2 | 20.9 | 6.3 | 6.1 | 5.2 | 7.5 | 7.1 | 6.5 | 7.2 | 6.5 | 5.2 | 7.2 | 6.4 | 5.4 | 7.9 | 7.7 | 6.7 |
| Cruciferin | Cruciferin | 23.7 | 1.4 | 1.3 | 2.3 | 2.2 | 1.7 | 2.2 | 1.9 | 1.6 | 2.2 | 1.9 | 1.6 | 2.6 | 2.7 | 2.7 | ||||||||
| α-polypeptides | 26.8 | 6.2 | 7.0 | 6.5 | 4.4 | 4.2 | 4.7 | α-polypeptides | 25.3 | 10.5 | 10.4 | 10.4 | 13.0 | 12.6 | 12.1 | 13.7 | 12.5 | 10.7 | 14.1 | 12.9 | 11.9 | 11.1 | 10.4 | 10.0 |
| 11S globulin | 28.4 | 10.6 | 13.2 | 11.7 | 11.7 | 14.1 | 12.3 | 11S globulin | 28 | 2.6 | 2.3 | 1.6 | 2.8 | 3.1 | 2.5 | 2.7 | 3.0 | 2.1 | 2.8 | 2.8 | 2.1 | 3.7 | 3.5 | 3.1 |
| Sin a 2 | 30.9 | 1.3 | 0.8 | Sin a 2 | 30.5 | 9.6 | 9.1 | 7.3 | 12.0 | 10.6 | 9.8 | 10.2 | 8.7 | 7.2 | 10.2 | 8.7 | 7.2 | 6.2 | 5.6 | 5.0 | ||||
| 32.9 | 10.5 | 11.0 | 9.4 | 10.0 | 10.4 | 9.3 | ||||||||||||||||||
| 11S fragment | 55 | 1.7 | 1.5 | 1.9 | 1.6 | 11S fragment | 52 | 1.0 | 0.5 | 1.1 | 1.4 | 1.3 | 0.8 | 1.0 | 0.9 | 1.1 | 1.2 | 1.0 | 1.8 | 2.4 | 1.3 | |||
| Procruciferin | 63.5 | 1.0 | 1.0 | 1.1 | 0.9 | 1.6 | 3.0 | Procruciferin | 63.6 | 2.9 | 1.1 | 1.6 | 1.5 | 1.3 | 1.5 | 1.7 | 1.7 | 1.3 | 1.2 | 2.0 | 3.0 | 1.0 | ||
| 66.8 | 2.0 | 1.5 | 2.0 | 2.7 | 65.9 | 3.8 | 2.6 | 2.3 | 1.5 | 1.1 | 1.1 | 1.3 | 1.2 | |||||||||||
| 69.4 | 3.4 | 5.9 | 5.2 | 2.0 | 3.2 | 2.0 | 69.9 | 2.0 | 1.8 | 1.2 | 1.1 | 1.9 | 2.4 | 2.0 | 1.2 | |||||||||
| 73.9 | 2.7 | 2.5 | 1.7 | 2.3 | 3.3 | 72.5 | 2.7 | 1.0 | 1.3 | 2.8 | 2.3 | 2.4 | 1.4 | 3.4 | 1.2 | |||||||||
| Band Number a | MW on SDS-PAGE (KDa) | Protein Name b | Protein Accession Numbers | Protein MW (KDa) | Exclusive Unique Peptide Count | Percentage Sequence Coverage c |
|---|---|---|---|---|---|---|
| Sinapis alba (AC Pennant) | ||||||
| S1 | 16 | ALL1_SINAL Allergen Sin a 1 [Sinapis alba (White mustard)] | P15322 | 16.4 | 5 | 23.40% |
| S2 | 28 | SINAL 11S globulin [Sinapis alba (White mustard)] | Q2TLW0 | 56.5 | 16 | 39.40% |
| S3 | 30 | SINAL 11S globulin [Sinapis alba (White mustard)] | Q2TLW0 | 56.5 | 16 | 39.40% |
| CRU4_BRANA Cruciferin CRU4 [Brassica napus (Rape)] | P33522 | 51.4 | 13 | 30.10% | ||
| S4 | 44 | SINAL 11S globulin [Sinapis alba (White mustard)] | Q2TLW0 | 56.5 | 19 | 44.30% |
| CRU4_BRANA Cruciferin CRU4 [Brassica napus (Rape)] | P33522 | 51.4 | 14 | 29.70% | ||
| S5 | 75 | SINAL 11S globulin [Sinapis alba (White mustard)] | Q2TLW0 | 56.5 | 18 | 44.30% |
| CRU4_BRANA Cruciferin CRU4 [Brassica napus (Rape)] | P33522 | 51.4 | 7 | 23.40% | ||
| S6 | 107 | SINAL 11S globulin [Sinapis alba (White mustard)] | Q2TLW0 | 56.5 | 21 | 46.50% |
| S7 | 223 | SINAL 11S globulin [Sinapis alba (White mustard)] | Q2TLW0 | 56.5 | 20 | 39.40% |
| BRANA Beta-glucosidase [Brassica napus (Rape)] | Q42618 | 56.5 | 20 | 34.40% | ||
| RS1 | 9.5 | ALL1_SINAL Allergen Sin a 1 [Sinapis alba (White mustard)] | P15322 | 16.4 | 7 | 26.90% |
| RS2 | 12 | ALL1_SINAL Allergen Sin a 1 [Sinapis alba (White mustard)] | P15322 | 16.4 | 9 | 57.90% |
| RS3 | 20 | SINAL 11S globulin [Sinapis alba (White mustard)] | Q2TLW0 | 56.5 | 19 | 36.10% |
| BRANA Cruciferin (Fragment) [Brassica napus (Rape)] | Q7XB53 | 51.3 | 8 | 23.20% | ||
| RS4 | 28 | SINAL 11S globulin [Sinapis alba (White mustard)] | Q2TLW0 | 56.5 | 14 | 25.90% |
| CRU3_BRANA Cruciferin CRU1 [Brassica napus (Rape)] | P33525 | 56.5 | 7 | 27.50% | ||
| RS5 | 34 | SINAL 11S globulin [Sinapis alba (White mustard)] | Q2TLW0 | 56.5 | 14 | 25.90% |
| SINAL 11S globulin [Sinapis alba (White mustard)] | Q2TLV9 | 57.9 | 5 | 34.60% | ||
| RS6 | 44 | SINAL 11S globulin [Sinapis alba (White mustard)] | Q2TLW0 | 56.5 | 18 | 46.70% |
| RS7 | 55 | SINAL 11S globulin [Sinapis alba (White mustard)] | Q2TLW0 | 56.5 | 15 | 37.60% |
| CRU4_BRANA Cruciferin CRU4 [Brassica napus (Rape)] | P33522 | 51.4 | 10 | 25.20% | ||
| RS8 | 62 | SINAL 11S globulin [Sinapis alba (White mustard)] | Q2TLW0 | 56.5 | 20 | 45.10% |
| RS9 | 75 | Q2TLW0_SINAL 11S globulin [Sinapis alba (White mustard)] | Q2TLW0 | 56.5 | 16 | 40.20% |
| BRANA Beta-glucosidase [Brassica napus (Rape)] | Q42618 | 58.5 | 12 | 20.80% | ||
| RS10 | 82 | Q2TLW0_SINAL 11S globulin [Sinapis alba (White mustard)] | Q2TLW0 | 56.5 | 14 | 37.10% |
| RS11 | 118 | SINAL 11S globulin [Sinapis alba (White mustard)] | Q2TLW0 | 56.5 | 12 | 27.50% |
| RS12 | 140 | SINAL 11S globulin [Sinapis alba (White mustard)] | Q2TLW0 | 56.5 | 9 | 24.50% |
| Brassicae juncea (AC Vulcan) | ||||||
| B1 | 15.5 | Allergen Bra j 1-E [Brassica juncea (Indian mustard)] | P80207 | 14.6 | 5 | 51.90% |
| OLES2_BRANA Oleosin S2-2 [Brassica napus (Rape)] | C3S7F1 | 19.9 | 15 | 46.80% | ||
| B2 | 17 | BRANA Oleosin S3-1 [Brassica napus (Rape)] | C3S7F8 | 19.6 | 10 | 41.70% |
| OLES2_BRANA Oleosin S2-2 [Brassica napus (Rape)] | C3S7F1 | 19.9 | 16 | 46.80% | ||
| B3 | 26 | BRANA Caleosin CLO1-2 [Brassica napus (Rape)] | C3S7H5 | 28.1 | 7 | 35.90% |
| CRU4_BRANA Cruciferin CRU4 [Brassica napus (Rape)] | P33522 | 51.4 | 21 | 49.50% | ||
| B4 | 30 | CRU3_BRANA Cruciferin CRU1 [Brassica napus (Rape)] | P33525 | 56.5 | 12 | 47.20% |
| OLES2_BRANA Oleosin S2-2 [Brassica napus (Rape)] | C3S7F1 | 19.9 | 11 | 46.30% | ||
| B5 | 29 | BRANA Cruciferin (Fragment) [Brassica napus (Rape)] | Q7XB53 | 51.3 | 4 | 8.80% |
| CRU4_BRANA Cruciferin CRU4 [Brassica napus (Rape)] | P33522 | 51.4 | 8 | 24.30% | ||
| B6 | 34 | SINAL 11S globulin [Sinapis alba (White mustard)] | Q2TLW0 | 56.5 | 7 | 20.80% |
| CRU3_BRANA Cruciferin CRU1 [Brassica napus (Rape)] | P33525 | 56.5 | 3 | 21.80% | ||
| B7 | 42.5 | CRU3_BRANA Cruciferin CRU1 [Brassica napus (Rape)] | P33525 | 56.5 | 12 | 45.20% |
| CRU4_BRANA Cruciferin CRU4 [Brassica napus (Rape)] | P33522 | 51.4 | 20 | 52.30% | ||
| B8 | 68 | CRU3_BRANA Cruciferin CRU1 [Brassica napus] | P33525 | 56.5 | 14 | 52.80% |
| CRU4_BRANA Cruciferin CRU4 [Brassica napus] | P33522 | 51.4 | 13 | 40.90% | ||
| B9 | 200 | Malate synthase. glyoxysomal [Brassica napus (Rape)] | P13244 | 63.7 | 14 | 23.00% |
| BRANA Beta-glucosidase [Brassica napus (Rape)] | Q42618 | 58.5 | 23 | 44.20% | ||
| RB1 | 9.5 | Allergen Bra j 1-E [Brassica juncea (Indian mustard)] | P80207 | 14.6 | 2 | 38.00% |
| RB2 | 12 | Allergen Bra j 1-E [Brassica juncea (Indian mustard)] | P80207 | 14.6 | 5 | 68.20% |
| ALL1_SINAL Allergen Sin a 1 [Sinapis alba (White mustard)] | P15322 | 16.5 | 4 | 35.20% | ||
| RB3 | 22 | CRU3_BRANA Cruciferin CRU1 [Brassica napus (Rape)] | P33525 | 56.5 | 11 | 39.50% |
| CRU4_BRANA Cruciferin CRU4 [Brassica napus (Rape)] | P33522 | 51.4 | 11 | 35.30% | ||
| RB4 | 27 | BRANA Caleosin CLO1-2 [Brassica napus (Rape)] | C3S7H5 | 28.1 | 8 | 37.60% |
| OLES2_BRANA Oleosin S2-2 [Brassica napus (Rape)] | C3S7F1 | 20.0 | 6 | 40.40% | ||
| BRAJU Glutathione S-transferase 3 [Brassica juncea] | Q7XZT2 | 24.1 | 7 | 46.00% | ||
| RB5 | 30 | CRU4_BRANA Cruciferin CRU4 [Brassica napus (Rape)] | P33522 | 51.4 | 17 | 42.60% |
| RB6 | 30 | CRU3_BRANA Cruciferin CRU1 [Brassica napus (Rape)] | P33525 | 56.5 | 12 | 45.20% |
| CRU4_BRANA Cruciferin CRU4 [Brassica napus (Rape)] | P33522 | 51.4 | 15 | 38.70% | ||
| RB7 | 34 | CRU3_BRANA Cruciferin CRU1 [Brassica napus (Rape)] | P33525 | 56.5 | 8 | 28.90% |
| BRAJU Cysteine synthase [Brassica juncea] | O23733 | 33.9 | 8 | 33.20% | ||
| RB8 | 47 | CRU4_BRANA Cruciferin CRU4 [Brassica napus (Rape)] | P33522 | 51.4 | 25 | 54.00% |
| BRANA Cruciferin (Fragment) [Brassica napus (Rape)] | Q7XB53 | 51.3 | 14 | 39.70% | ||
| RB9 | 52 | CRU4_BRANA Cruciferin CRU4 [Brassica napus (Rape)] | P33522 | 51.4 | 16 | 45.40% |
| BRACM Enolase [Brassica campestris (Field mustard)] | Q6W7E8 | 47.4 | 21 | 61.00% | ||
| RB10 | 62 | BRANA Beta-glucosidase [Brassica napus (Rape)] | Q42618 | 58.5 | 13 | 30.00% |
| RB11 | 76 | CRU4_BRANA Cruciferin CRU4 [Brassica napus (Rape)] | P33522 | 51.4 | 8 | 30.10% |
| SINAL 11S globulin [Sinapis alba (White mustard)] | Q2TLW0 | 56.5 | 5 | 15.30% | ||
| Protein Identification a | Species | Best %ID b | # Hits >35% | Full Alignment | NCBI Links f | ||
|---|---|---|---|---|---|---|---|
| E-value c | %ID d | Length e | |||||
| P15322: Allergen Sin a 1 [Sinapis alba (White mustard)] | |||||||
| gid|192|Allergen Allergen Sin a 1 precursor | Sinapis alba | 100.00% | 66 of 66 | 2.4e-34 | 100.0% | 145 | gi|51338758 |
| gid|1172|Putative 2S storage protein | Brassica rapa | 93.80% | 66 of 66 | 2.5e-29 | 88.30% | 145 | gi|17697 |
| gid|1142|Putative Napin-3 | Brassica napus | 91.20% | 66 of 66 | 2.6e-17 | 82.60% | 144 | gi|75107016 |
| gid|1170|Putative Allergen Bra j 1-E | Brassica juncea | 87.50% | 66 of 66 | 1.8e-17 | 79.90% | 144 | gi|32363444 |
| P80207: Allergen Bra j 1-E [Brassica juncea (Indian mustard)] | |||||||
| gid|1170|Putative Allergen Bra j 1-E (Bra j 1) | Brassica juncea | 100.00% | 50 of 50 | 7.1e-23 | 100.0% | 129 | gi|32363444 |
| gid|1142|Putative Napin-3 (Napin BnIII) | Brassica napus | 92.50% | 50 of 50 | 8.9e-20 | 89.10% | 129 | gi|75107016 |
| gid|192|Allergen allergen sin a 1.0104 | Sinapis alba | 88.78% | 50 of 50 | 2.5e-13 | 80.60% | 144 | gi|1009434 |
| gid|1172|Putative 2S storage protein | Brassica rapa | 85.00% | 50 of 50 | 1e-11 | 77.10% | 144 | gi|17697 |
| gid|386|Putative recombinant Ib pronapin | Brassica napus | 55.00% | 50 of 50 | 3.8e-6 | 50.00% | 114 | gi|26985163 |
| Q2TLWO: 11S globulin [Sinapis alba (White mustard)] | |||||||
| gid|837|Allergen 11S globulin precursor | Sinapis alba | 100.00% | 431 of 431 | 0 | 100.0% | 510 | gi|62240390 |
| gid|2066|Putative 11S legumin protein | Carya illinoinensis | 62.51% | 351 of 431 | 2.6e-61 | 43.20% | 521 | gi|158998782 |
| gid|2597|Putative legumin | Juglans nigra | 62.51% | 362 of 431 | 2.7e-58 | 42.00% | 528 | gi|1126299828 |
| gid|76|Allergen allergen Ana 0 2 | Anacardium occidentale | 60.04% | 380 of 431 | 2.3e-70 | 44.80% | 498 | gi|25991543 |
| gid|160|Allergen HMW glutenin | Triticum aestivum | 60.00% | 84 of 431 | 3.5e-18 | 37.60% | 213 | gi|288860106 |
| gid|392|Allergen 11S globulin | Corylus avellana | 58.00% | 387 of 431 | 3.6e-61 | 45.70% | 534 | gi|18479082 |
| gid|1572|Putative prunin 2 precursor | Prunus dulcis | 57.80% | 418 of 431 | 3e-61 | 45.60% | 522 | gi|307159114 |
| gid|2650|Allergen 11S globulin- | Actinidia chinensis | 57.50% | 352 of 431 | 2.3e-68 | 41.40% | 503 | gi|82469930 |
| gid|2274|Putative 11S globulin | Sesamum indicum | 55.00% | 342 of 431 | 3.7e-60 | 42.10% | 470 | gi|13183173 |
| gid|1093|Putative 11S globulin | Pistacia vera | 55.00% | 356 of 431 | 6.5e-53 | 41.80% | 505 | gi|156001070 |
| gid|531|Putative allergenic protein | Fagopyrum tataricum | 55.00% | 256 of 431 | 1.5e-43 | 38.10% | 486 | gi|113200131 |
| gid|733|Putative glycinin subunit G3 | Glycine max | 53.10% | 272 of 431 | 2e-38 | 36.20% | 514 | gi|18639 |
| gid|347|Putative 11S globulin | Bertholletia excelsa | 52.50% | 318 of 431 | 5.8e-59 | 39.20% | 510 | gi|30313867 |
| Q7XB53: Cruciferin (Fragment) [Brassica napus (Rape)] | |||||||
| gid|837|Allergen 11S globulin precursor | Sinapis alba | 76.50% | 387 of 387 | 3.5e-29 | 61.00% | 472 | gi|62240390 |
| gid|392|Allergen Cor a 9 allergen | Corylus avellana | 63.70% | 353 of 387 | 9.4e-22 | 44.00% | 509 | gi|557792009 |
| gid|1572|Putative Pru du 6 allergen | Prunus dulcis | 62.51% | 160 of 387 | 3.2e-10 | 48.20% | 193 | gi|523916668 |
| gid|76|Allergen allergen Ana 0 2 | Anacardium occidentale | 61.30% | 333 of 387 | 1.2e-22 | 44.70% | 474 | gi|25991543 |
| gid|2597|Putative legumin | Juglans nigra | 58.79% | 298 of 387 | 3.8e-21 | 42.70% | 503 | gi|1126299828 |
| gid|2066|Putative 11S legumin protein | Carya illinoinensis | 58.79% | 326 of 387 | 3.1e-21 | 43.50% | 506 | gi|158998782 |
| gid|817|Putative seed storage protein | Juglans regia | 57.52% | 325 of 387 | 4.4e-21 | 42.90% | 513 | gi|56788031 |
| gid|1093|Putative Pis v 2.0201 allergen 11S | Pistacia vera | 56.30% | 347 of 387 | 1.7e-33 | 41.60% | 459 | gi|110349085 |
| gid|347|Putative 11S globulin | Bertholletia excelsa | 53.80% | 349 of 387 | 1.2e-24 | 43.30% | 466 | gi|30313867 |
| gid|733|Putative glycinin subunit G3 | Glycine max | 53.80% | 274 of 387 | 4.8e-22 | 38.00% | 479 | gi|18639 |
| gid|2274|Putative 11S globulin | Sesamum indicum | 52.46% | 318 of 387 | 1.6e-24 | 40.80% | 476 | gi|13183173 |
| gid|291|Allergen trypsin inhibitor | Arachis hypogaea | 49.40% | 59 of 387 | 1.9e-5 | 35.80% | 204 | gi|22135348 |
| P33525: Cruciferin CRU4 [Brassica napus (Rape) | |||||||
| gid|837|Allergen 11S globulin precursor | Sinapis alba | 98.80% | 430 of 430 | 2.4e-210 | 91.60% | 510 | gi|62240390 |
| gid|2066|Putative 11S legumin protein | Carya illinoinensis | 65.00% | 358 of 430 | 1.9e-61 | 44.70% | 535 | gi|158998782 |
| gid|2597|Putative legumin | Juglans nigra | 63.79% | 373 of 430 | 4.1e-61 | 43.30% | 533 | gi|1126299828 |
| gid|817|Putative seed storage protein | Juglans regia | 63.79% | 368 of 430 | 1e-63 | 44.50% | 533 | gi|56788031 |
| gid|1572|Putative Pru du 6 allergen | Prunus dulcis | 60.04% | 168 of 430 | 4.6e-20 | 49.70% | 195 | gi|523916668 |
| gid|76|Allergen allergen Ana 0 2 | Anacardium occidentale | 60.04% | 379 of 430 | 7.9e-73 | 46.20% | 487 | gi|25991543 |
| gid|392|Allergen Cor a 9 allergen | Corylus avellana | 58.79% | 391 of 430 | 4.6e-62 | 45.50% | 528 | gi|557792009 |
| gid|160|Allergen glutenin | Triticum aestivum | 56.26% | 70 of 430 | 2.5e-11 | 33.30% | 207 | gi|736319 |
| gid|1093|Putative Pis v 2.0101 11S globulin | Pistacia vera | 55.00% | 345 of 430 | 6.7e-51 | 41.10% | 518 | gi|110349083 |
| gid|2274|Putative 11S globulin | Sesamum indicum | 53.80% | 343 of 430 | 4.8e-63 | 42.10% | 478 | gi|13183173 |
| gid|531|Putative allergenic protein | Fagopyrum tataricum | 53.80% | 255 of 430 | 4.4e-44 | 35.60% | 523 | gi|113200131 |
| gid|347|Putative 11S globulin | Bertholletia excelsa | 52.50% | 309 of 430 | 2.3e-59 | 39.70% | 506 | gi|30313867 |
| gid|733|Putative glycinin subunit G3 | Glycine max | 51.20% | 267 of 430 | 3e-39 | 36.00% | 516 | gi|18639 |
| gid|291|Allergen allergen Arah3/Arah4 | Arachis hypogaea | 47.50% | 219 of 430 | 2.1e-25 | 33.20% | 548 | gi|21314465 |
| gid|574|Putative glycinin precursor | Glycine max | 47.50% | 116of430 | 4.9e-28 | 39.80% | 191 | gi|169971 |
| Q2TLV9: 11S globulin [Sinapis alba (White mustard)] | |||||||
| gid|837|Allergen 11S globulin precursor | Sinapis alba | 100.00% | 444 of 444 | 0 | 100.00% | 523 | gi|62240392 |
| gid|160|Allergen HMW glutenin | Triticum aestivum | 68.80% | 99 of 444 | 1.3e-12 | 38.60% | 228 | gi|288860106 |
| gid|2066|Putative 11S legumin protein | Carya illinoinensis | 63.79% | 360 of 444 | 6.3e-41 | 41.90% | 532 | gi|158998782 |
| gid|817|Putative seed storage protein | Juglans regia | 62.51% | 367 of 444 | 1.1e-41 | 41.70% | 545 | gi|56788031 |
| gid|2597|Putative legumin | Juglans nigra | 62.51% | 364 of 444 | 2.5e-41 | 40.80% | 539 | gi|1126299828 |
| gid|76|Allergen allergen Ana 0 2 | Anacardium occidentale | 61.30% | 380 of 444 | 9.9e-70 | 42.70% | 510 | gi|25991543 |
| gid|1572|Putative Chain A | Prunus dulcis | 59.30% | 417 of 444 | 6.5e-53 | 45.20% | 524 | gi|258588247 |
| gid|392|Allergen Cor a 9 allergen | Corylus avellana | 59.30% | 390 of 444 | 3.4e-41 | 43.70% | 545 | gi|557792009 |
| gid|1572|Putative prunin 1 precursor | Prunus dulcis | 59.30% | 436 of 444 | 2.6e-53 | 44.90% | 543 | gi|307159112 |
| gid|160|Allergen HMW glutenin | Triticum aestivum | 57.52% | 76 of 444 | 4.2e-9 | 31.30% | 307 | gi|21751 |
| gid|1572|Putative Pru du 6 allergen | Prunus dulcis | 57.50% | 168 of 444 | 4.8e-20 | 46.20% | 208 | gi|523916668 |
| gid|2650|Allergen 11S globulin | Actinidia chinensis | 56.30% | 344 of 444 | 4.1e-67 | 39.70% | 514 | gi|82469930 |
| gid|1093|Putative Pis v 2.0201 allergen 11S | Pistacia vera | 56.30% | 332 of 444 | 9.2e-53 | 39.50% | 516 | gi|110349085 |
| gid|2274|Putative 11S globulin | Sesamum indicum | 53.80% | 345 of 444 | 2e-60 | 41.10% | 482 | gi|13183173 |
| gid|574|Putative glycinin A3B4 subunit | Glycine max | 53.80% | 241 of 444 | 6.4e-27 | 32.40% | 558 | gi|10566449 |
| gid|291|Allergen allergen Arah3/Arah4 | Arachis hypogaea | 47.50% | 208 of 444 | 1.2e-20 | 32.40% | 561 | gi|21314465 |
| C3S7F1: Oleosin S2-2 [Brassica napus (Rape)] | |||||||
| gid|2298|Allergen oleosin | Corylus avellana | 70.01% | 109 of 109 | 2.2e-42 | 52.50% | 158 | gi|49617323 |
| gid|2283|Putative oleosin 1 | Arachis hypogaea | 64.98% | 104 of 109 | 6.5e-39 | 46.20% | 171 | gi|113200509 |
| gid|1893|Putative oleosin | Sesamum indicum | 62.50% | 103 of 109 | 2.2e-31 | 45.90% | 157 | gi|10834827 |
| gid|389|Putative oleosin | Corylus avellana | 50.00% | 68 of 109 | 1.1e-21 | 41.00% | 117 | gi|29170509 |
| gid|2285|Allergen oleosin 3 | Arachis hypogaea | 45.10% | 77 of 109 | 9.1e-21 | 38.20% | 144 | gi|52001241 |
| C3S7F8: Oleosin S3-1 [Brassica napus (Rape] | |||||||
| gid|1238|Putative 15 kDa oleosin | Sesamum indicum | 76.20% | 101 of 101 | 6e-31 | 63.20% | 125 | gi|5381321 |
| gid|389|Putative oleosin | Corylus avellana | 73.80% | 98 of 101 | 1.1e-32 | 66.90% | 121 | gi|29170509 |
| gid|2284|Putative oleosin 1 | Arachis hypogaea | 70.00% | 94 of 101 | 1.2e-28 | 60.80% | 125 | gi|71040655 |
| Q6W7E8: Enolase [Brassica campestris (Field mustard)] | |||||||
| gid|586|Putative Enolase | Hevea brasiliensis | 98.80% | 365 of 365 | 1.1e-170 | 89.60% | 442 | gi|14423687 |
| gid|1955|Allergen Alpha-enolase | Thunnus albacares | 88.70% | 365 of 365 | 2.9e-129 | 69.50% | 442 | gi|576011129 |
| gid|1959|Allergen enolase | Salmo salar | 88.70% | 365 of 365 | 5.1e-130 | 69.90% | 442 | gi|385145180 |
| gid|2710|Allergen beta-enolase | Gallus gallus | 83.80% | 365 of 365 | 3.6e-119 | 65.10% | 441 | gi|46048765 |
| gid|396|Allergen Enolase | Candida albicans | 82.50% | 365 of 365 | 1.5e-111 | 63.10% | 444 | gi|232054 |
| gid|103|Putative enolase | Rhodotorula mucilaginosa | 80.00% | 365 of 365 | 9.5e-107 | 61.00% | 441 | gi|30314940 |
| Q7XZT2-BRAJU Glutathione S-transferase 3 [Brassica juncea] | |||||||
| gid|856|Allergen glutathione S transferase c | Periplaneta americana | 40.00% | 14 of 134 | 1.2e-6 | 29.40% | 163 | gi|359326557 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
L’Hocine, L.; Pitre, M.; Achouri, A. Detection and Identification of Allergens from Canadian Mustard Varieties of Sinapis alba and Brassica juncea. Biomolecules 2019, 9, 489. https://doi.org/10.3390/biom9090489
L’Hocine L, Pitre M, Achouri A. Detection and Identification of Allergens from Canadian Mustard Varieties of Sinapis alba and Brassica juncea. Biomolecules. 2019; 9(9):489. https://doi.org/10.3390/biom9090489
Chicago/Turabian StyleL’Hocine, Lamia, Mélanie Pitre, and Allaoua Achouri. 2019. "Detection and Identification of Allergens from Canadian Mustard Varieties of Sinapis alba and Brassica juncea" Biomolecules 9, no. 9: 489. https://doi.org/10.3390/biom9090489
APA StyleL’Hocine, L., Pitre, M., & Achouri, A. (2019). Detection and Identification of Allergens from Canadian Mustard Varieties of Sinapis alba and Brassica juncea. Biomolecules, 9(9), 489. https://doi.org/10.3390/biom9090489