Molecular Cytogenetics of Eurasian Species of the Genus Hedysarum L. (Fabaceae)

 , and
, and
Abstract
:1. Introduction
2. Results
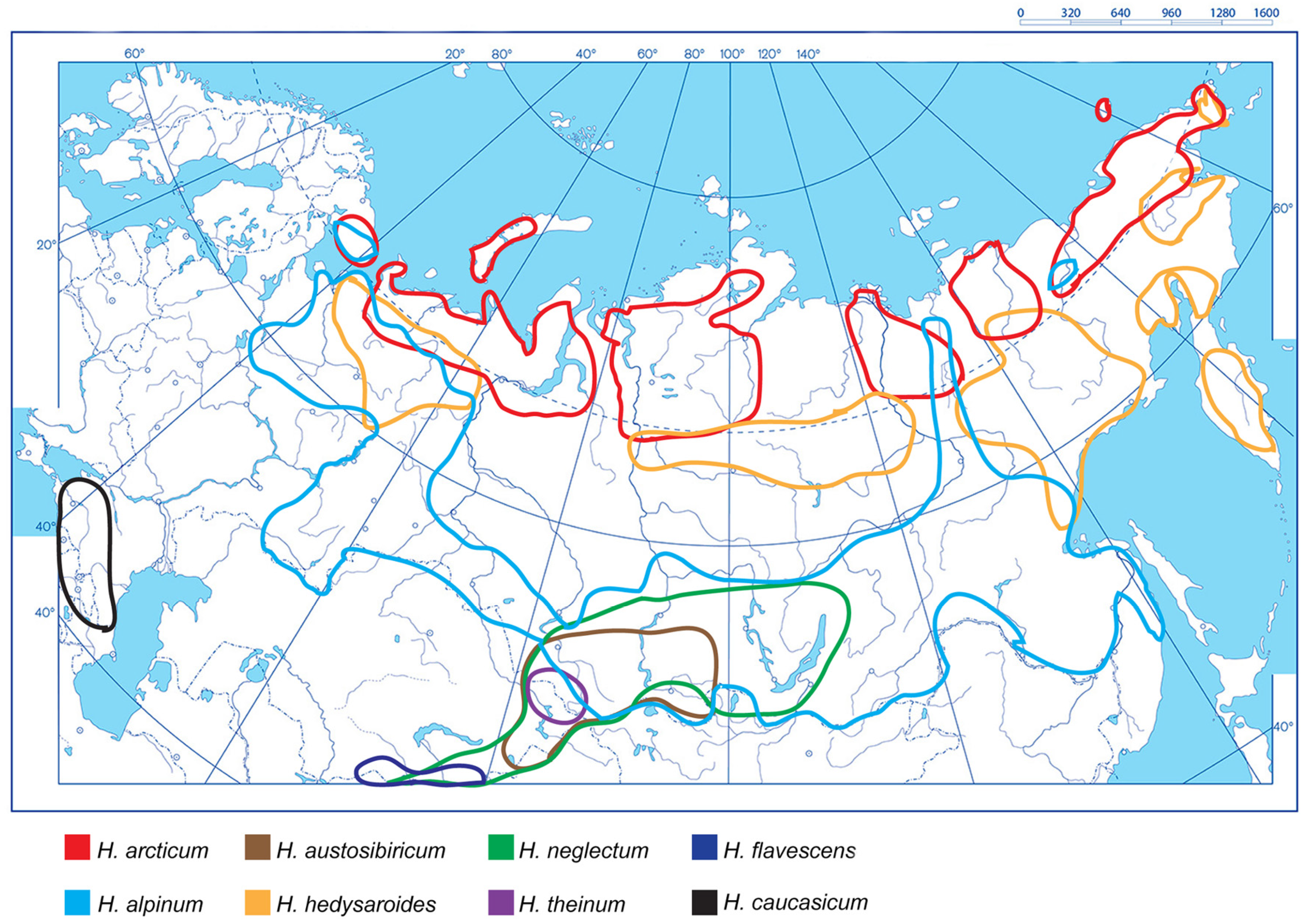
2.1. Distribution Areas of the Studied Hedysarum Species
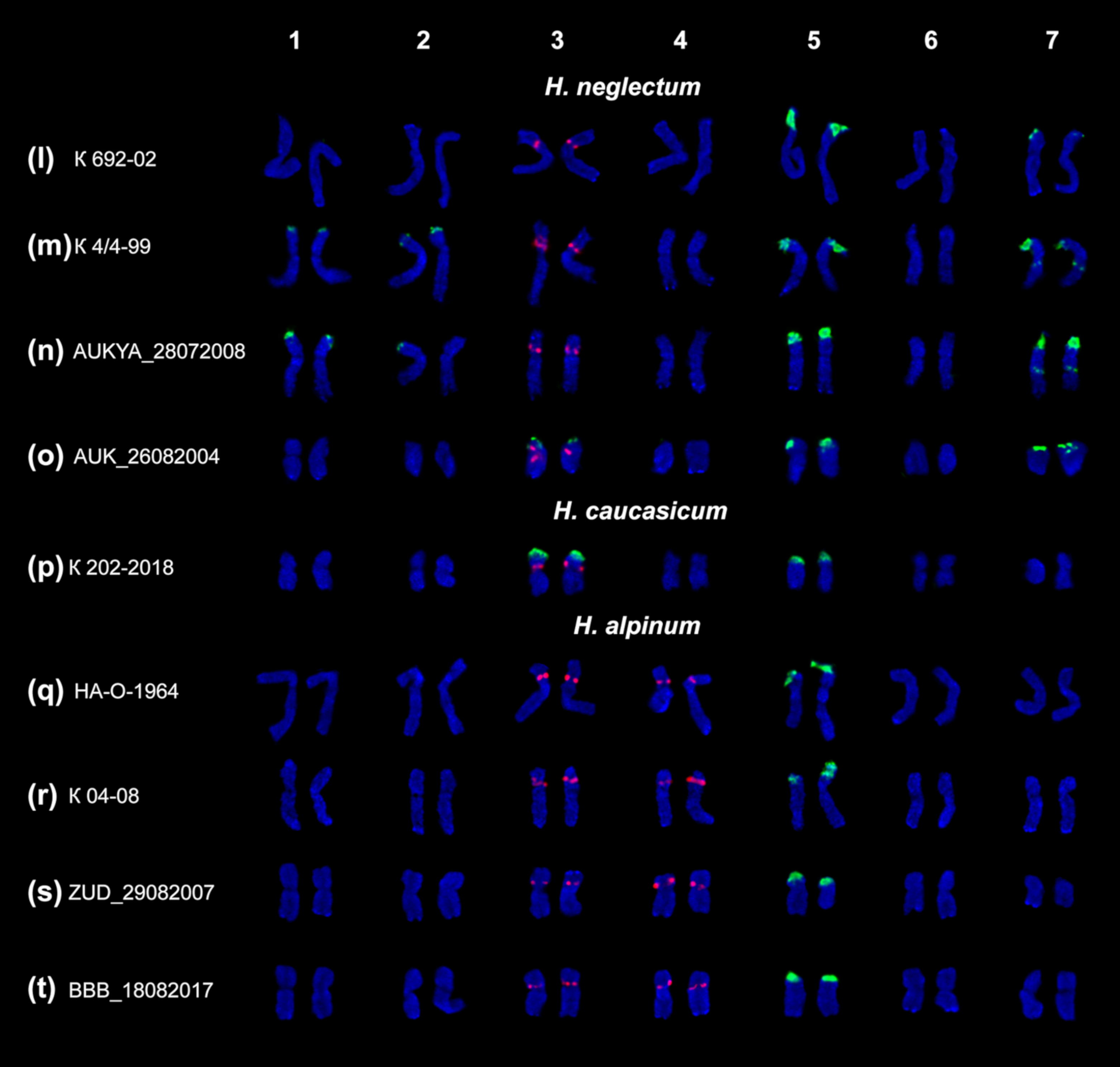
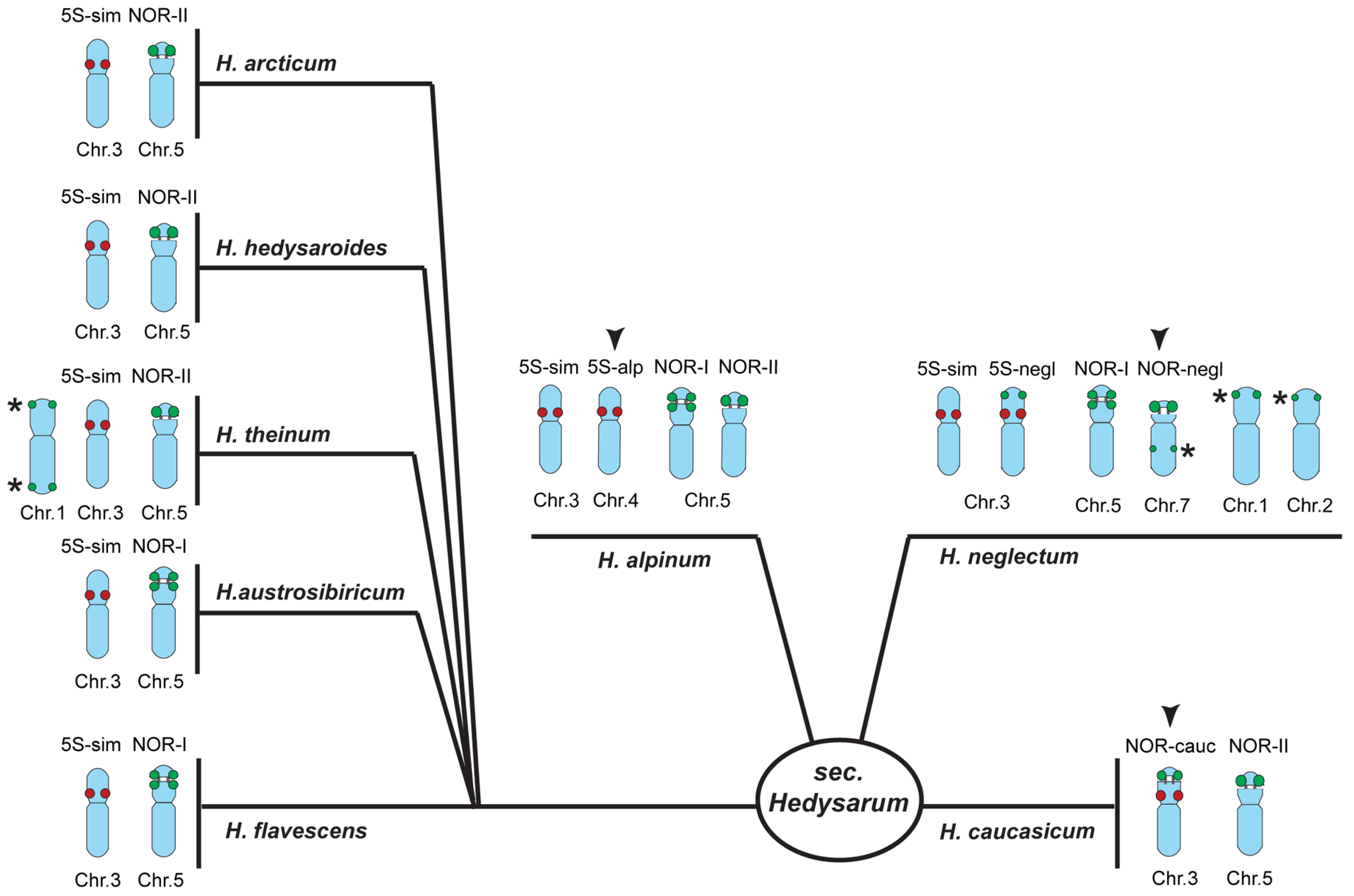
2.2. Chromosomal Structural Variations in the Studied Species
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Chromosome Spread Preparation
4.3. FISH Procedure
4.4. Chromosome Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fedtschenko, B.A. The genus Hedysarum. Trudy Imp. S. Petersburgsk Bot. Sada 1902, 19, 185–342. [Google Scholar]
- Roskov, Y.R. Hedysarea. In Legumes of Northern Eurasia: A Checklist; Yakovlev, G.P., Sytin, A.K., Roskov, Y.R., Eds.; Royal Botanic Gardens: London, UK, 1996; pp. 379–426. [Google Scholar]
- Choi, B.-H.; Ohashi, H. Proposal to conserve the name Hedysarum (Leguminosae: Papilionoideae) with a conserved type. Taxon 1998, 47, 877. [Google Scholar] [CrossRef]
- Choi, B.-H.; Ohashi, H.; Ohashi, B.-H.C. Generic criteria and an infrageneric system for Hedysarum and related genera (Papilionoideae-Leguminosae). Taxon 2003, 52, 567–576. [Google Scholar] [CrossRef]
- Basiner, T.F. Enumeratio monographica specierum generis Hedysari. Bull. Phys. Math. Acad. Sci. (Petersb.) 1845, 4, 305–315. [Google Scholar]
- Yurtsev, B.A. Arctic flora of the USSR [Arkticheskaya flora SSSR]; Nauka: Saint Petersburg, Russia, 1986; Volume 9. [Google Scholar]
- Mironov, Y.M. Pericarp anatomy of East European species of the genus Hedysarum L. (Papilionaceae): Sections Gamotion and Multicaulia. Bull. Mosc. Soc. Nat. Ser. Biol. 2000, 105, 50–53. [Google Scholar]
- Choi, B.H. Foliar Flavonoids of the genus Hedysarum and related genera (tribe Hedysareae—Leguminosae). Korean J. Plant Taxon. 1994, 24, 259–264. [Google Scholar] [CrossRef]
- Vysochina, G.I.; Kukushkina, T.A.; Karnaukhova, N.A.; Selyutina, I.Y. Flavonoids of wild and introduced plants of several species of the Hedysarum L Genus. Chem. Sustain. Dev. 2011, 19, 327–333. [Google Scholar]
- Dong, Y.-M.; Tang, D.; Zhang, N.; Li, Y.; Zhang, C.; Li, L.; Li, M.-H. Phytochemicals and biological studies of plants in genus Hedysarum. Chem. Cent. J. 2013, 7, 124. [Google Scholar] [CrossRef] [Green Version]
- Rybachenko, A.I.; Krivut, B.A.; Georgievskii, V.P. Fluorodensitometric determination of mangiferin and isomangiferin in Hedysarum flavescens and H. alpinum. Chem. Nat. Compd. 1976, 12, 395–396. [Google Scholar] [CrossRef]
- Komissarenko, A.N.; Nadezhina, T.P.; Komissarenko, N.F. Flavonoids and xanthones of Hedysarum connatum and H. alpinum. Chem. Nat. Compd. 1994, 30, 521–522. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, Q.-Y.; Zhao, Y.-Y.; Wang, B.; Hai, L.-Q.; Ying, Y.-P.; Chen, H.-B. Saponins from the roots of Hedysarum polybotrys. Biochem. Syst. Ecol. 2007, 35, 389–391. [Google Scholar] [CrossRef]
- Nechepurenko, I.V.; Polovinka, M.P.; Komarova, N.I.; Korchagina, D.V.; Salakhutdinov, N.F.; Nechepurenko, S.B. Low-molecular-weight phenolic compounds from Hedysarum theinum roots. Chem. Nat. Compd. 2008, 44, 31–34. [Google Scholar] [CrossRef]
- Zinner, N.S.; Vysochina, G.I.; Kukushkina, T.A.; Sviridova, T.P. Biologically active substances Hedysarum alpinum and H. theinum (Fabaceae) introduced to Tomsk region. Tomsk State Univ. J. Biol. 2010, 4, 116–122. [Google Scholar]
- Fedtschenko, B.A. Hedysarum. In Flora URSS; Komarov, V.L., Shishkin, B.K., Bobrov, E.G., Eds.; Akad. Scient. URSS: Moscow, Russia, 1948; Volume 13, pp. 259–379. [Google Scholar]
- Bajtenov, M.S. Genus Hedysarum L. In Flora of Kazahstan; Pavlov, N.V., Ed.; Izd-vo AN Kazah. SSR: Alma-Ata, Kazakhstan, 1961; Volume V, pp. 418–442. [Google Scholar]
- Bajtenov, M.S. Flora of Kazakhstan. Generic Complex of Flora; Gylym: Almaty, Kazakhstan, 2001; Volume 2. [Google Scholar]
- Sviridova, T.P.; Zinner, N.S. Prospects of cultivation Hedysarum alpinum L. and H. theinum Krasnob. in conditions of Tomsk region. Tomsk State Univ. J. Biol. 2008, 2, 5–11. [Google Scholar]
- Erst, A.A.; Zheleznichenko, T.V.; Novikova, T.I.; Dorogina, O.V.; Banaev, E.V. Ecological and geographic variability of Hedysarum theinum and features of its propagation in vitro. Contemp. Probl. Ecol. 2014, 7, 67–71. [Google Scholar] [CrossRef]
- Arslan, E.; Ertugrul, K.; Tugay, O.; Dural, H. Karyological studies of the genus Onobrychis Mill. and the related genera Hedysarum L. and Sartoria Boiss. and Heldr. (Fabaceae, Hedysareae) from Turkey. Caryologia 2012, 65, 11–17. [Google Scholar] [CrossRef]
- Zvyagina, N.S.; Dorogina, O.V.; Catalan, P. Genetic relatedness and taxonomy in closely related species of Hedysarum (Fabaceae). Biochem. Syst. Ecol. 2016, 69, 176–187. [Google Scholar] [CrossRef] [Green Version]
- Kumar, P.; Rana, P.K.; Singhal, V.K.; Singh, H.; Kholia, B.S. Chromosome count, meiotic abnormalities and pollen sterility in Lahaul sweetvetch (Hedysarum astragaloides Benth. ex Baker, Fabaceae), an endemic and threatened species from India. Acta Bot. Croat. 2018, 77, 203–208. [Google Scholar] [CrossRef]
- Issolah, R.; Benhizia, H.; Khalfallah, N. Karyotype variation within some natural populations of Sulla (Hedysarum coronarium L., Fabaceae) in Algeria. Genet. Resour. Crop Evol. 2006, 53, 1653. [Google Scholar] [CrossRef]
- Tropicos.org. Missouri Botanical Garden. IPCN Chromosome Reports. Available online: http://www.tropicos.org/Name/40022354 (accessed on 21 November 2020).
- Benhizia, H.; Benhizia, Y.; Ghernoub, L. Meiotic behaviour and karyotype features of endangered endemic fodder species Hedysarum perrauderianum (Fabaceae) in some populations from Algeria. Caryologia 2013, 66, 195–204. [Google Scholar] [CrossRef] [Green Version]
- Benhizia, H.; Benhizia, Y.; Djeghar, R.; Siljak-Yakovlev, S.; Pustahija, F.; Khalfallah, N. Cytogenetic characterization, nuclear genome size, and pollen morphology of some Hedysarum L. taxa (Fabaceae) from Algeria, with emphasis on the origin of H. perrauderianum Coss. & Durieu. Genet. Resour. Crop Evol. 2020. [Google Scholar] [CrossRef]
- Marghali, S.; Panaud, O.; Lamy, F.; Ghariani, S.; Sarr, A.; Marrakchi, M.; Trifi-Farah, N. Exploration of intra- and inter-population genetic diversity in Hedysarum coronarium L. Genet. Resour. Crop Evol. 2005, 52, 277–284. [Google Scholar] [CrossRef]
- Bushman, B.S.; Larson, S.R.; Peel, M.; Pfrender, M.E. Population structure and genetic diversity in North American Hedysarum boreale Nutt. Crop Sci. 2007, 47, 1281–1288. [Google Scholar] [CrossRef] [Green Version]
- Zvyagina, N.S.; Dorogina, O.V. Genetic differentiation of Altai-Sayan endemic Hedysarum theinum Krasnob (Fabaceae) by inter-simple sequence repeat analysis. Russ. J. Genet. 2013, 49, 1030–1035. [Google Scholar] [CrossRef]
- Agafonova, M.; Agafonova, O. Polymorphism of seed polypeptides in closely related species Hedysarum theinum Krasnob. and H. neglectum Ledeb. (Fabaceae). Turczaninowia 2002, 5, 72–78. [Google Scholar]
- Dorogina, O.V.; Agafonova, M.A. Identification of closely related species Hedysarum theinum, H. neglectum and H. austrosibiricum (Fabaceae) by seed storage globulins. Bot. Zh. 2004, 89, 1637–1645. [Google Scholar]
- Duan, L.; Wen, J.; Yang, X.; Liu, P.L.; Arslan, E.; Ertuğrul, K.; Chang, Z.Y. Phylogeny of Hedysarum and tribe Hedysareae (Leguminosae: Papilionoideae) inferred from sequence data of ITS, matK, trnL-F and psbA-trnH. Taxon 2015, 64, 49–64. [Google Scholar] [CrossRef]
- Liu, P.L.; Wen, J.; Duan, L.; Arslan, E.; Ertuğrul, K.; Chang, Z.Y. Hedysarum L. (Fabaceae: Hedysareae) is not monophyletic—Evidence from phylogenetic analyses based on five nuclear and five plastid sequences. PLoS ONE 2017, 12, e0170596. [Google Scholar] [CrossRef] [PubMed]
- Nafisi, H.; Kazempour-Osaloo, S.; Mozaffarian, V.; Schneeweiss, G.M. Molecular phylogeny and divergence times of the genus Hedysarum (Fabaceae) with special reference to section Multicaulia in Southwest Asia. Plant Syst. Evol. 2019, 305, 1001–1017. [Google Scholar] [CrossRef]
- Kurbatsky, V.I. Genus Hedysarum L. In Flora Sibiri (Flora of Siberia), Fabaceae (Leguminosae); Polozhii, A., Ed.; Nauka: Novosibirsk, Russia, 1994; Volume 9, pp. 153–166. [Google Scholar]
- Malyshev, L.I. Manual of High Mountain Plants of the Southern Siberia; Nauka: Saint Petersburg, Russia, 1968. [Google Scholar]
- Kharkevich, S.S. Vascular Plants of the Soviet Far East. V.4; Nauka: Saint Petersburg, Russia, 1989. [Google Scholar]
- Litvinskaya, S.A.; Murtazaliyev, R.A. Flora of the North Caucasus: Atlas Continuant; Fiton XXI: Moscow, Russia, 2013. [Google Scholar]
- Imachuyeva, D.R.; Serebryanaya, F.K. The current state of study of plants of Hedysarum L. florae of the Caucasus. Pharm. Pharmacol. 2016, 4, 4–32. [Google Scholar] [CrossRef] [Green Version]
- Kurbatsky, V.I.; Malahova, L.A. Chromosome numbers for some species of Hedysarum L. from the territory of Siberia. Tomsk State Univ. 1992, 89, 3–5. [Google Scholar]
- Kulikov, P.V. Floristic findings in the Southern Ural (Chelyabinsk region). Bot. Zh. 1998, 83, 137–145. [Google Scholar]
- Knyasev, M.S. A new hybrid species of the genus Hedysarum (Fabaceae) from the East Europe. Bot. Zh. 2011, 96, 1122–1126. [Google Scholar]
- Krasnoborov, I.M.; Azovtsev, G.R.; Orlov, V.P. A new species of the genus Hedysarum L. (Fabaceae L.) from southern Siberia. Bot. Zh. 1985, 70, 968–973. [Google Scholar]
- Tutin, T.G.; Heywood, V.H.; Burges, N.A.; Moore, D.M.; Valentine, D.H.; Walters, S.M.; Webb, D.A. Flora Europaea, Leguminosae; Cambridge University Press: Cambridge, UK, 1968; Volume 2. [Google Scholar]
- Kobayashi, T. A new role of the rDNA and nucleolus in the nucleus—rDNA instability maintains genome integrity. Bioessays 2008, 30, 267–272. [Google Scholar] [CrossRef] [PubMed]
- Schubert, I.; Wobus, U. In situ hybridization confirms jumping nucleolus organizing regions in Allium. Chromosoma 1985, 92, 143–148. [Google Scholar] [CrossRef]
- Raskina, O.; Belyayev, A.; Nevo, E. Quantum speciation in Aegilops: Molecular cytogenetic evidence from rDNA cluster variability in natural populations. Proc. Natl. Acad. Sci. USA 2004, 101, 14818–14823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roa, F.; Guerra, M. Distribution of 45S rDNA sites in chromosomes of plants: Structural and evolutionary implications. BMC Evol. Biol. 2012, 12, 225. [Google Scholar] [CrossRef] [Green Version]
- Rosato, M.; Álvarez, I.; Feliner, G.N.; Rosselló, J.A. High and uneven levels of 45S rDNA site-number variation across wild populations of a diploid plant genus (Anacyclus, Asteraceae). PLoS ONE 2017, 12, e0187131. [Google Scholar] [CrossRef] [Green Version]
- Pedrosa-Harand, A.; de Almeida, C.C.; Mosiolek, M.; Blair, M.W.; Schweizer, D.; Guerra, M. Extensive ribosomal DNA amplification during Andean common bean (Phaseolus vulgaris L.) evolution. Theor. Appl. Genet. 2006, 112, 924–933. [Google Scholar] [CrossRef]
- Scaldaferro, M.A.; da Cruz, M.V.; Cecchini, N.M.; Moscone, E.A. FISH and AgNor mapping of the 45S and 5S rRNA genes in wild and cultivated species of Capsicum (Solananceae). Genome 2016, 59, 95–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amosova, A.V.; Samatadze, T.E.; Mozgova, G.V.; Kipen, V.N.; Dubovskaya, A.G.; Artemyeva, Y.O.Y.; Zoshchuk, S.A.; Lemesh, V.A.; Muravenko, O.V. Genomic markers associated with cold-hardiness in Brassica rapa L. Mol. Biol. 2020, 54, 541–552. [Google Scholar] [CrossRef]
- Gerlach, W.L.; Bedbrook, J.R. Cloning and characterization of ribosomal RNA genes from wheat and barley. Nucleic Acids Res. 1979, 7, 1869–1885. [Google Scholar] [CrossRef] [PubMed]
- Gerlach, W.L.; Dyer, T.A. Sequence organization of the repeating units in the nucleus of wheat which contain5S rRNA genes. Nucleic Acids Res. 1980, 8, 4851–4855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yurkevich, O.Y.; Samatadze, T.E.; Levinskikh, M.A.; Zoshchuk, S.A.; Signalova, O.B.; Surzhikov, S.A.; Sychev, V.N.; Amosova, A.V.; Muravenko, O.V. Molecular cytogenetics of Pisum sativum L. grown under spaceflight-related stress. BioMed Res. Int. 2018, 2018, 1–10. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Accession Number/Voucher | Origin/Seed Source |
|---|---|---|
| H. alpinum L. | HA-O-1964 | Omsk region, RF/germplasm collection of CSBG, 2009 |
| H. alpinum L. | K 04-08 | Unknown/germplasm collection of AIMAP, 2017 |
| H. alpinum L. | ZUD_29082007 | 51°7′ N; 112°11′ E; Zabaikalye region, RF/collected by Dr. N.A. Karnaukhova, 2007 |
| H. alpinum L. | BBB_18082017 | 53°58′ N; 113°35′ E; Bauntovskii region, Buryatia, RF/collected by Dr. I.Yu. Selyutina, 2017 |
| H. arcticum B. Fedtsch. | K 02252/818_24072016 | 67°25′ N; 65°11′ E; lake Shchuch’e, Komi Republic, RF/collected by Dr. N.A. Suprun, 2016 |
| H. arcticum B. Fedtsch. | K 499-17 | VRBG/germplasm collection of AIMAP, 2017 |
| H. arcticum B. Fedtsch. | K 292-19 | VRBG/germplasm collection of AIMAP, 2017 |
| H. austrosibiricum B. Fedtsch. | KZ_RIP_25071998 | 50°19′ N; 83°50′ E; Ivanovskiy Ridge, 2000 m above sea level, Kazakhstan/collected by Dr. I.Yu. Selyutina, 1998 |
| H. caucasicum M. Bieb. | K 202-2018 | Gothenburg Botanical Garden, Sweden/germplasm collection of AIMAP, 2018 |
| H. flavescens Rgl. et Schmalh. | CSBG_Fl_U30072007 | University of Uppsala Botanical Garden, Sweden, Uppsala/germplasm collection of CSBG, 2008 |
| H. hedysaroides (L.) Schinz et Thell. | K 518-17 | Botanical Garden of the University of Vienna, Austria/germplasm collection of AIMAP, 2017 |
| H. hedysaroides (L.) Schinz et Thell. | K 445-17 | University of Tartu Botanical Gardens, Estonia/germplasm collection of AIMAP, 2017 |
| H. hedysaroides (L.) Schinz et Thell. | K 96-19 | Botanical Garden of the University of Vienna, Austria/germplasm collection of AIMAP, 2017 |
| H. neglectum Ledeb. | K 4/4-99 | Kaira Yalbak Mountains, Altai region, RF/germplasm collection of AIMAP, 2017 |
| H. neglectum Ledeb. | K 692-02 | Altai region/germplasm collection of AIMAP, 2017 |
| H. neglectum Ledeb. | AUK_26082004 | 50°39′ N; 87°43′ E; Kubadru River, Altai region, RF/collected by Dr. I.Yu. Selyutina, 2004 |
| H. neglectum Ledeb. | AUKYA_28072008 | 50°52′ N; 85°15′ E; Yabogansky pass, Altai region, RF/collected by Dr. N.A. Karnaukhova, 2008 |
| H. theinum Krasnob. | AUKK_15082008 | 50°59′ N; 84°13′ E; Kumir river, Altai region, RF/collected by Dr. I.Yu. Selyutina, 2008 |
| H. theinum Krasnob. | KZ_RP_25072008 | 50°18′ N; 83°38′ E; Ridderskii region, Kazakhstan/collected by Dr. I.Yu. Selyutina, 2008 |
| H. theinum Krasnob. | AUSK_05092008 | 50°4′ N; 85°13′ E; Ust-Koksinsky region; Altai Mountains, RF/collected by Dr. N.A. Karnaukhova, 2008 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yurkevich, O.Y.; Samatadze, T.E.; Selyutina, I.Y.; Romashkina, S.I.; Zoshchuk, S.A.; Amosova, A.V.; Muravenko, O.V. Molecular Cytogenetics of Eurasian Species of the Genus Hedysarum L. (Fabaceae). Plants 2021, 10, 89. https://doi.org/10.3390/plants10010089
Yurkevich OY, Samatadze TE, Selyutina IY, Romashkina SI, Zoshchuk SA, Amosova AV, Muravenko OV. Molecular Cytogenetics of Eurasian Species of the Genus Hedysarum L. (Fabaceae). Plants. 2021; 10(1):89. https://doi.org/10.3390/plants10010089
Chicago/Turabian StyleYurkevich, Olga Yu., Tatiana E. Samatadze, Inessa Yu. Selyutina, Svetlana I. Romashkina, Svyatoslav A. Zoshchuk, Alexandra V. Amosova, and Olga V. Muravenko. 2021. "Molecular Cytogenetics of Eurasian Species of the Genus Hedysarum L. (Fabaceae)" Plants 10, no. 1: 89. https://doi.org/10.3390/plants10010089