
Combined Effects of Different Alleles of FLO2, Wx and SSIIa on the Cooking and Eating Quality of Rice
by

Yu Zhang
1,2,
Jiajia Zhao
1,
Yaqi Hu
1,
Yanni Zhang
1,
Yining Ying
1,
Feifei Xu
1 and
Jinsong Bao
1,2,* 1
Institute of Nuclear Agricultural Sciences, Key Laboratory for Nuclear Agricultural Sciences of Zhejiang Province and Ministry of Agriculture and Rural Affairs, Zhejiang University, Zijingang Campus, Hangzhou 310058, China
2
Hainan Institute of Zhejiang University, Yazhou Bay Science and Technology City, Yazhou District, Sanya 572025, China
*
Author to whom correspondence should be addressed.
Plants 2022, 11(17), 2249; https://doi.org/10.3390/plants11172249
Submission received: 8 August 2022
/
Revised: 19 August 2022
/
Accepted: 23 August 2022
/
Published: 30 August 2022
(This article belongs to the Special Issue Germplasm Enhancement and Breeding for Rice Quality Improvement)
Abstract
:The improvement of the cooking and eating quality (CEQ) of rice is one of the major objectives of current rice-breeding programs. A few major genes such as Waxy (Wx) and starch synthase IIa (SSIIa) have been successfully applied in molecular breeding. However, their interactive effects on CEQ have not been fully understood. In this study, a recombinant inbred line (RIL) population was constructed by crossing the white-core mutant GM645 with the transparent phenotype of the japonica rice variety Tainung 67 (TN67). GM645 and TN67 contain different alleles of FLOURY ENDOSPERM2 (FLO2), Wx, and SSIIa. The effects of different allele combinations of FLO2, Wx, and SSIIa on the CEQ of rice were investigated. The inbred lines with the mutation allele flo2 had a significantly lower apparent amylose content (AAC), viscosity characteristics except for setback (SB), and gel texture properties compared to those lines with the FLO2 allele. The allelic combination of FLO2 and Wx significantly affected the AAC, breakdown (BD), and gel textural properties, which could explain most of the variations in those rice quality traits that were correlated with AAC. The allelic combination of FLO2 and SSIIa significantly affected the hot paste viscosity (HPV) and pasting temperature (PT). The Wx × SSIIa interaction had a significant effect on the PT. The interaction of FLO2, Wx and SSIIa significantly affected the AAC, cold paste viscosity (CPV), PT, and consistency viscosity (CS). These results highlight the important roles of these quality-related genes in regulating the CEQ of rice and provide new clues for rice-quality improvement by marker-assisted selection.
1. Introduction
Rice (Oryza sativa L.), with a long history of cultivation and consumption, can provide more than 20% of the required energy for the entire global population [1,2]. Given the improvement in rice production techniques and people’s quality of life as well as the change in consumption styles and habits, people have placed higher demands on rice quality [3,4]. Cooking and eating quality (CEQ) and appearance quality are highly valued by consumers [5]. CEQ is usually indirectly reflected by the apparent amylose content (AAC), the viscosity characteristics measured by a rapid viscosity analyzer (RVA), and textural parameters such as hardness and cohesiveness measured by a texture profile analyzer [6]. Appearance quality includes indicators such as chalkiness (percentage of chalky grains and the degree of chalkiness), grain length, grain width, and length to width ratio [7].
Starch is the most important compound component in rice seeds [8,9], which accounts for 75–85% of their dry weight and is related to the appearance, stickiness, hardness, palatability, and digestibility of cooked rice [10]. Starch consists of linear polysaccharides containing α-1,4 glycosidic bonds (amylose) and highly branched polysaccharides containing both α-1,4 and α-1,6 glycosidic bonds (amylopectin) [11,12]. Starch is synthesized in the endosperm by the action of ADP-glucose pyrophosphorylase (AGPase), starch synthases (SSs), branching enzymes (BEs), and debranching enzymes (DBEs). Among them, the granulated starch synthase (GBSS) encoded by the Waxy (Wx) gene is responsible for amylose synthesis, while SSs extend the glucan chains of starch linked by α-1,4 glycosidic bonds, BEs introduce the branching point of the α-1,6 glycosidic linkage, and finally starch relies on DBEs to remove incorrect branch points. The latter three kinds of enzymes work together to complete the synthesis of amylopectin [13,14,15].
In the rice germplasm, various Wx alleles have been identified, including Wxa, Wxb, Wxin, Wxop, Wxhp, Wxmq, Wxmp, Wxlv, and wx [16,17,18,19,20]. Among the non-glutinous rice varieties, Wxa and Wxb are the two main functional genes of Wx [2]. Wxa, which leads to a high AAC, is mainly found in indica rice; Wxb, which leads to a low or moderate AAC, is mainly found in japonica rice. Compared with Wxa, the single nucleotide polymorphism (SNP) G at the first intron splice site becomes T in Wxb. This mutation reduces the splicing efficiency of pre-mRNA and reduces the accumulation of GBSS, which in turn results in a lower AAC [21]. Multiple other alleles have been identified from different rice varieties in the last decades [18,19,20]. In addition to the above allelic variants, the 5′-untranslated region of Wx exon 1 also contains a (CT)n microsatellite locus, and the polymorphism of this locus correlates with the AAC [22,23,24]. The SS family, which is involved in the branching extension of amylopectin, contains the SSI, SSII, SSIII, and SSIV isoforms. Among them, the gene-encoding SSIIa, also known as the main effector gene in regulating gelatinization temperature [24,25], can extend the A and B1 short chains of DP <10 to form the B1 long chain of amylopectin [26]. The SNP variation of GC/TT in exon 8 has been found to have the greatest effect on pasting temperature (PT) [27,28], and varieties containing GC usually show a high PT, while varieties containing TT usually show a low PT [27].
The floury endosperm is very similar to the chalky phenotype in terms of the grains’ transparency and the starch granule morphology and arrangement, so it can be regarded as an extreme chalky phenotype [29]. Many floury mutant genes, such as FLO2, FLO4–8, FLO10–16, and FLO18–19, have been cloned and characterized [30,31,32,33,34,35,36,37,38,39,40,41,42,43,44,45]. Among them, the FLO2 is located on the chromosome 4 and encodes a protein with a tetratricopeptide repeat motif (TPR) domain containing three motifs, which can mediate protein interactions and is involved in regulating the expression of a variety of starch synthesis-related enzyme genes and storage protein genes [30]. The FLO2 mutant can reduce AAC and storage proteins, and alter the structure of amylopectin [30,46,47,48].
At present, a large volume of research has been conducted on the Wx and/or SSIIa genes regarding their functions and their allelic effects on grain quality [10,49,50], but no studies have been conducted that analyze the specific effects of different allele combinations of the FLO2, Wx, and SSIIa genes on the CEQ of rice. To this end, a recombinant inbred line (RIL) population of GM645/Tainung67 (TN67), obtained in a previous study [51], is employed to investigate the effects of different combinations of Wx, SSIIa, and FLO2 on the CEQ of rice. Our results will facilitate an understanding of the effects of gene interactions on rice starch synthesis and CEQ formation and provide new avenues for rice-quality improvement by marker-assisted selection.1
2. Results
2.1. Variation Analysis of Quality Traits in Parents and RIL
The eleven starch characteristics of the parents were analyzed, and significant differences were found between the two parents for seven traits: AAC, PV, SB, PT, HD, ADH, and COH (Table 1). The coefficient of variation of SB (247.8%) was the largest among all the traits, which ranged from −121.1 to 135.4 RUV. The smallest coefficient of variation was found in PT (6.1%), which ranged from 66.9 to 83.9 °C (Table 1). All the rice quality traits showed a continuous variation in the RIL population (Figure 1). A significant transgressive segregation was found for all the traits, and the mean values of the traits except the HPV, PT, CS, and HD were between the two parents (Table 1). The skewness and kurtosis for the PV, HPV, CPV, BD, SB, and CS were <1.0, which displayed an approximately normal distribution, indicating that these traits were quantitative traits that are controlled by multiple genes (Table 1; Figure 1).
A principal component analysis was performed on the 11 rice quality traits (Figure 2). The PC1 and PC2 could explain 91.6% of the total variance. The AAC had the largest positive effect on PC1, and the HPV, CPV, CS, ADH, HD, and SB had a strong correlation with PC1. All the above seven traits were strongly correlated with each other. COH had a large negative effect on PC1, and negatively correlated with the seven traits above. The PV, BD, and PT had a large positive effect on PC2, and SB had the largest negative effect on PC2, so PV, BD, and PT were negatively correlated with SB.
2.2. Genotyping of RIL
The white-core phenotype of GM645 is caused by the presence of a 1 bp thymine deletion in exon 18 of the FLO2 gene, leading to an early appearance of the terminator codon [49]. The PCR product containing the thymine deletion can be specifically identified and cleaved by the HinfI (Figure 3). Hereafter, FLO2 indicates the wild-type and flo2 indicates the mutant allele, so GM645 possesses the flo2 allele and TN67 possesses the FLO2 allele. Among the 127 RILs, 41 lines had the flo2 allele and 86 lines had the FLO2 allele.
The white-core mutant GM645 had the Wxa-(CT)11 and SSIIa(GC) alleles, while the japonica Tainung67 (TN67) with the transparent endosperm had the Wxb-(CT)17 and SSIIa(TT) alleles. The polyacrylamide gel electrophoresis plots of the PCR products from the 127 RIL populations showed that there were 59 Wxa lines and 68 Wxb lines (Figure 3). The GC/TT polymorphism in exon 8 of SSIIa could be genotyped using four primers [28], and according to the results of agarose gel electrophoresis, there were 77 lines with the SSIIa(GC) and 50 lines containing the SSIIa(TT) in the RIL population (Figure 3).
2.3. Effects of FLO2, Wx, and SSIIa Single Gene on the AAC, Viscosity Properties, and Textural Properties in the RIL Population
FLO2 and its mutant allele flo2 had significant effects on nine traits (Table 2). Compared to FLO2, the RIL lines carrying the flo2 allele showed lower levels in all traits except COH (p < 0.05), indicating that the loss of function of FLO2 had a great impact on the starch-related physicochemical properties and the CEQ of rice.
Wx is the major gene regulating the AAC, viscosity properties, and textural properties in rice endosperm. Wxa maintained higher levels in most of the starch quality traits except for the CPV, PT, and COH. The different alleles of SSIIa only had a significant effect on HPV and PT (p < 0.05) but had no effects on the AAC and textural properties (Table 2).
2.4. Differences in the AAC, Viscosity and Textural Properties among Eight Genetic Combinations
Since FLO2, Wx, and SSIIa are genetically segregated in the RIL population, there may be specific interactive effects among these three genes. Theoretically, a total of eight genotype combinations can be generated. A total of 24 lines had the FLO2/Wxa/SSIIa(GC) combination (genotype), 18 had the FLO2/Wxa/SSIIa(TT) genotype, 24 had the FLO2/Wxb/SSIIa(GC) genotype, 20 the had FLO2/Wxb/SSIIa(TT) genotype, 13 had the flo2/Wxa/SSIIa(GC) genotype, 4 had the flo2/Wxa/SSIIa(TT) genotype, 16 had the flo2/Wxb/SSIIa(GC) genotype, and 8 had the flo2/Wxb/SSIIa(TT) genotype. The starch quality traits among the eight combinations are shown in Table 3.
Among the eight combinations, all eleven starch quality traits were different (p < 0.05), indicating that there were significant interactions between FLO2, Wx, and SSIIa. In terms of the AAC, the FLO2/Wxa/SSIIa(TT) genotype had the highest average value (26.23 ± 2.67%), and the lowest average value was in the flo2/Wxb/SSIIa(TT) genotype (10.87 ± 2.08%). The SSIIa(TT) allele was involved in both combinations, suggesting that SSIIa may have a weak effect on the AAC. In terms of viscosity characteristics, the BD (136.0 ± 35.1 RVU) and PT (81.7 ± 1.2 °C) were highest in the FLO2/Wxb/SSIIa(GC) genotype. The PVs of the FLO2/Wxb/SSIIa(TT) genotype (289.7 ± 35.4 RVU) and FLO2/Wxb/SSIIa(GC) genotype (283.3 ± 48.3 RVU) were the highest. The highest CPV (345.4 ± 25.1 RVU) and CS (152.1 ± 19.6 RVU) were found in the FLO2/Wxa/SSIIa(GC) genotype, but the FLO2/Wxa/SSIIa(TT) genotype had the same CPV (343.5 ± 40.6 RVU). The FLO2/Wxa/SSIIa(TT) combination showed the highest HPV (214.5 ± 38.7 RVU). The flo2/Wxa/SSIIa(TT) combination had the highest SB (97.4 ± 6.0 RVU). Most of the lowest average values appeared in the flo2/Wxb/SSIIa(TT) combination, except for the BD and SB. In terms of textural properties, the highest values of the two traits, HD and ADH, appeared in the FLO2/Wxa/SSIIa(GC) combination, while the lowest average values appeared in the flo2/Wxb/SSIIa(TT) combination. The highest average value of COH belongs to the FLO2/Wxb/SSIIa(TT) combination.
2.5. Effects of Interaction between FLO2, Wx, and SSIIa on AAC, Viscosity Properties, and Textural Properties of RIL
The analysis of the interactions between the genes for the variation in the starch properties was carried out by ANOVA, and the results are shown in Figure 4. The Wx gene accounted for more than 91% of the total variation of AAC, so the AAC was mainly controlled by the Wx gene. Similarly, Wx could explain more than 53% of the variation in the CPV, BD, SB, CS, HD, ADH, and COH, indicating that those traits were also controlled by the Wx and had a close correlation with the AAC. FLO2 accounted for more than 90% of the PV variation and was the only factor that had a significant effect on the PV (p < 0.001). FLO2 also explained 56% of the total variation in the HPV, 43% of the total variation in the CPV, and 16–27% of the total variation in the BD, PT, CS, HD and ADH, indicating that the viscosity and gel textural traits were also controlled by FLO2. SSIIa was responsible for PT, which explained 71% of the total variation.
Significant interactions between the genes were detected. Among the interactions, the FLO2 × Wx interaction accounted for 9–16% of the total variation in the gel textural properties. The FLO2 × Wx interaction was also significant for AAC and BD, but it only explained less than 2% of the total variation.
FLO2 × SSIIa interaction accounted for 6% of the total variation in the HPV. Although the interaction was significant for the PT, it only explained around 1% of the total variation. The Wx × SSIIa interaction was only detected in the PT, explaining less than 1% of the total variation. A significant FLO2 × Wx × SSIIa interaction was detected for most traits (Figure 4). This triple interaction could explain 6% of the total variation in the CS, and around 2% of the total variation in the PV, CPV, BD, HD, ADH, and COH, and less than 1% of the total variation of other traits.
2.6. Cluster Analysis Based on the Quality Traits of RIL Population
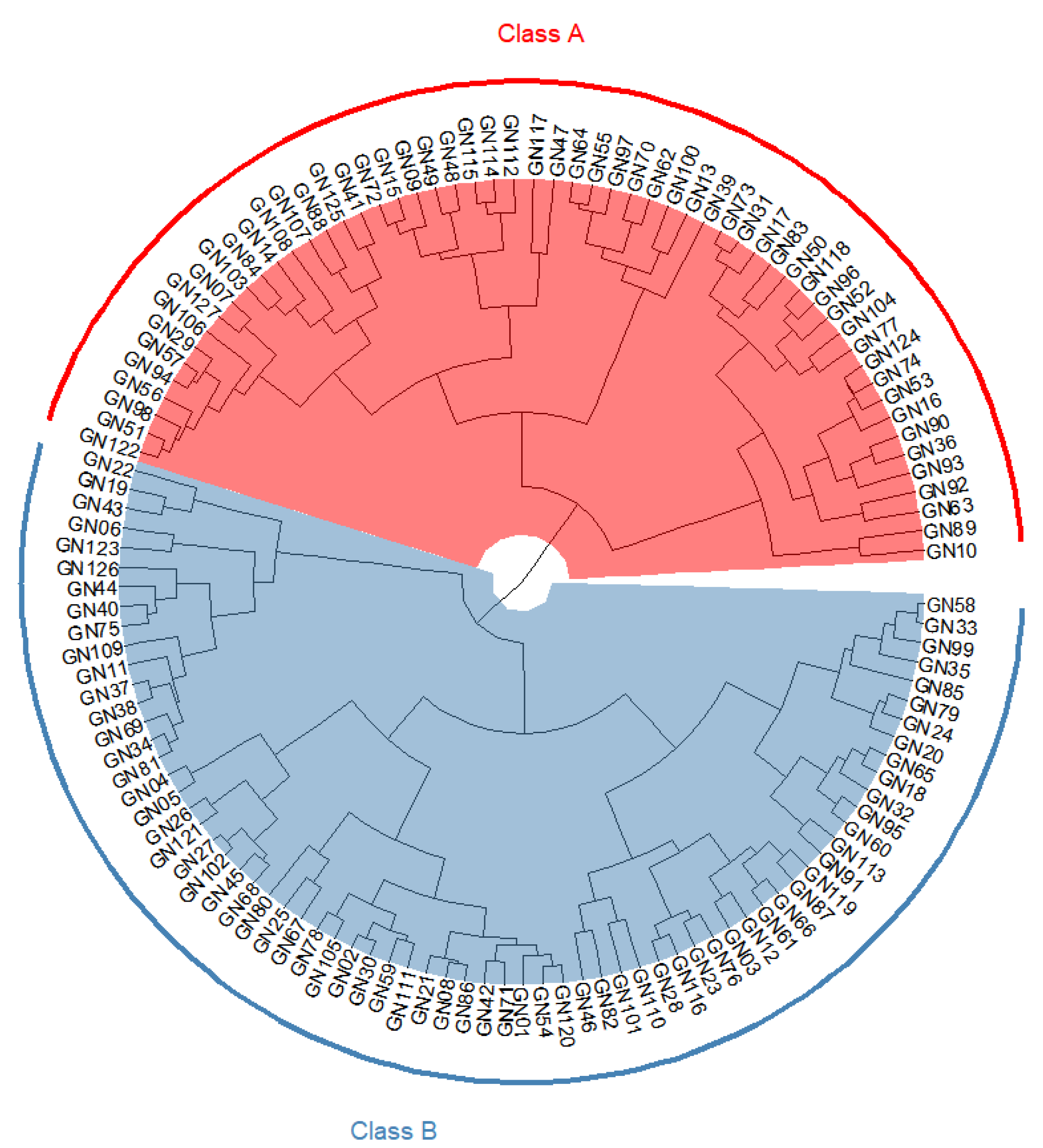
A cluster analysis was performed using hierarchical clustering with all 11 starch quality traits as variable indicators. The 127 populations of RIL were divided into two classes (Figure 5). When analyzed in combination with the genotype combinations contained in the 127 populations, Class A was found to contain FLO2/Wxa/SSIIa(GC), FLO2/Wxa/SSIIa(TT), flo2/Wxa/SSIIa(GC), and flo2/Wxa/SSIIa(TT); Class B contained FLO2/Wxb/SSIIa(GC), FLO2/Wxb/SSIIa(TT), flo2/Wxb/SSIIa(GC), and flo2/Wxb/SSIIa(TT). The clustering results showed that the classification of the RIL population basically depended on the genotype of Wx. Class A contained lines with a high AAC, while class B contained lines with a low to medium AAC. This also proved that Wx was the most important gene regulating the CEQ traits of rice.
3. Discussion
3.1. FLO2 Affects Rice Quality
The quality of rice, especially the CEQ, plays a crucial role in the market value of rice. It has been found that a series of genes directly or indirectly involved in starch synthesis can regulate the quality of rice [52,53]. Among them, FLO2 plays a key role in the regulation of rice grain size and starch quality by regulating the expression levels of starch synthesis-related genes and storage protein genes in the endosperm [30,46,47]. FLO2 contains a total of 23 exons and 22 introns, encoding a protein with two domains: one is a TPR domain containing three repeats of the TPR motif, and the other is an unproven protein clueless (CLU) domain containing 245 amino acid residues [30,54]. The TPR domain is a mediator domain that is completely composed of a helical structure and participates in the interaction between proteins. It plays an important role in the formation of protein complexes. Different motifs in the same protein have different effects and different characteristics [55,56]. The CLU domain is a large and highly conserved protein involved in normal mitochondrial function [57].
This study found that the AAC decreased in the RIL population lines containing the flo2 mutant allele, which is consistent with previous findings [30,48], indicating that FLO2 had an effect on amylose synthesis. Compared with the wild-type Guangluai4 (GLA4), the GM645 carrying the flo2 mutant allele only reduced the AAC by 6%, and the difference between the two accessions did not reach a significant level [48]. In this study, under the same Wx background, the flo2 allele can reduce the AAC by 17% in the Wxa background and 11% in the Wxb background (Table 3). However, in previous studies, other flo2 alleles showed a very significant effect on the AAC. She et al. [30] identified a flo2 allele with a mutated site in exon 14 and found the AAC decreased by about 40% compared to the wild-type. Wu, et al. [46] reported three flo2 alleles with mutations in exon 19, intron 9, and intron 11, and found that the AAC decreased by 24%−26% compared to the wild-type. Qiu [54] identified nine flo2 alleles with mutation sites in exons 6, 10, 11, and 21 and introns 13, 14, 16, and 17. The AAC was reduced by about 67%−73%. These evidences imply that different mutation sites in FLO2 will lead to different degrees of influence on the AAC. This may be due to the different effects of mutations on the TPR domain and the CLU protein domain, and different TPR motifs may have different roles in regulating amylose synthesis. The phenotypic differences caused by different allelic mutants have also been reported in flo4 and flo6 [58].
All the viscosity characteristics of the RIL population containing the flo2 allele were significantly lower than those of the line containing the FLO2 allele except SB, which was consistent with the results of previous studies [46,48]. The viscosity characteristics of rice are usually correlated with the AAC. Our principal component analysis also confirmed that the AAC was significantly correlated with most RVA viscosities except the BD and PT (Figure 2). Thus, the effect of FLO2 on the RVA viscosity was mainly due to its effect on the AAC. However, the fine structure of amylopectin also has an important impact on the RVA profile. Studies have shown that the short chain in amylopectin (DP 6–12) reduces the SB and CPV by reducing the rate of molecular polymerization [59]. The longer the branched chains are, the more easily they are intertwined with each other, which is conducive to maintaining the integrity of starch granules and reducing the capacity for water absorption and swelling, resulting in a lower BD, PV, and HPV [60,61]. Compared with the FLO2 from GLA4, the flo2 from GM645 increased the short chain (DP 6–9 and DP 22–35) and ultra-long chain DP ≥ 44 [48]. Combined with the change in the RVA profile, it is the flo2 mutation that caused the change in the amylopectin side chain distribution and then the change of RVA viscosity parameters. However, She et al. [30] showed that a DP ≤ 9 and DP 22–28 in the amylopectin short chain of the mutant increased, while the DP 9–21 and long chain (DP ≥ 38) decreased, suggesting that different FLO2 mutation sites may have different effects on the fine structure of amylopectin. In flo2 mutants, the expression of genes related to starch synthesis, such as AGPS1, GBSSI, SSIIa, BEI, BEIIa, BEIIb, PUL, and ISA1, were down regulated [30,62], which can explain why the AAC decreased and why the amylopectin chain length distribution changed. This study also found that FLO2 has the most significant effect on the PV (Figure 4), explaining more than 90% of the total variation (Figure 4). The PV indicates the swelling degree of starch particles and their ability to bind to water, which is related to the AAC, amylopectin fine structure, and non-carbohydrate components (lipids and proteins) in starches. Therefore, FLO2 plays a unique role in the PV.
The texture of rice gel is mainly affected by the content and morphology of starch, protein, non-starch polysaccharides, and other substances [63]. Most of the texture parameters in the FLO2 mutant were lower than those in the wild-type. In addition to the reduced AAC invoked in the flo2 mutant, other studies have confirmed that the total starch content and total protein content were decreased to some degrees, but that the soluble sugar content was increased [54]. The AAC is negatively correlated with COH, and positively correlated with HD and ADH (Figure 2). According to the results of this study, the effect of FLO2 on rice gel texture was due to its effect on the AAC. Whether FLO2 could regulate the texture characteristics by affecting the amylopectin fine structure or protein content needs to be further explored.
3.2. Interaction Effect of FLO2, Wx and SSIIa
The rice CEQ is regulated by a series of genes. The interactive effects between different genes/enzymes are diverse and complex, forming a complex regulatory network [15,64,65,66]. As a coding gene of GBSSI, Wx can regulate the content of amylose in rice, which is related to a variety of rice-quality indicators and plays an important role in the CEQ of rice. Among the 11 CEQ parameters, Wx had a significant effect on 10 traits except for PV (Figure 4). The cluster analysis revealed that the grouping of RIL lines was based on the different alleles of Wx (Figure 5). In addition to Wx, CEQ is also regulated by other starch synthesis-related genes such as SSIIa. However, this study found that SSIIa only had a significant effect on the AAC, BD, SB, and PT.
The FLO2 × Wx gene interaction has a significant impact on the AAC, BD, HD, ADH and COH. This gene combination can explain most of the changes in rice quality traits involving the AAC. The FLO2 × SSIIa gene interaction only has a significant effect on the HPV and PT. Wx is the main gene regulating the AC and gel consistency, and it is also the minor gene affecting the gelatinization temperature (GT) [49]. On the contrary, SSIIa mainly regulates the GT, but also has a certain effect on the AC and GC [49]. In previous studies, Wx × SSIIa was considered to have a great impact on RVA characteristics [10]. Our study found that the interaction between Wx and SSIIa only has a significant effect on the PT.
FLO2 and flo2 have different effects on rice quality traits under different allele backgrounds of Wx and SSIIa. Under the condition of the same SSIIa allele, in the Wxa background, a total of 10 traits, including the AAC, PV, HPV, BD, CPV, PT, CS, HD, ADH, and COH, were affected differently by FLO2 and flo2 (Table 3). In the background of Wxb, the PV, HPV, BD, CPV, PT, and CS were significantly affected (Table 3). With the same Wx allele, when in the background of SSIIa(TT), the RILs containing FLO2 had a higher AAC, PV, HPV, BD, CPV, PT, CS, and ADH than those RILs containing flo2 (Table 3). In the background of SSIIa(GC), the PV, HPV, BD, CPV, PT, CS, HD, and ADH were significantly different between FLO2 and flo2 (Table 3). These results indicated that there are significant interactions between FLO2, Wx and SSIIa. As expected, the ANOVA revealed that the FLO2 × Wx × SSIIa effects reached significant levels for the AAC, CPV, PT, and CS.
High quality rice should generally have a good palatability with a soft and elastic texture. Cooked rice with a 14%–20% medium amylose content will have a fluffy and soft texture. This study found that among the eight allele combinations composed of three genes, the FLO2/Wxb/SSIIa(TT) has the lowest AAC, PT, and HD, implying that it has a soft texture and poor retrogradation. This allele combination is expected to have the best CEQ.
In this study, it was found that single gene FLO2, Wx, and SSIIa are the main factors affecting the variation in all the rice quality traits. The FLO2 and Wx genes have significant effects on the variation of most viscosity and texture properties, which is mainly due to the fact that amylose content is mainly regulated by both FLO2 and Wx. However, the interactive effects are also significant for many CEQ traits. The FLO2 × Wx allele combination showed much greater effects on quality traits than the FLO2 × SSIIa and Wx × SSIIa. It is important to explore the allelic combinations of these genes for the improvement of the CEQ of rice.
4. Materials and Methods
4.1. Plant Materials
The white-core endosperm mutant GM645 was obtained from induced mutation breeding of the indica rice (Oryza sativa L.) variety Guangluai4 (GLA4) [48]. The white-core endosperm phenotype is caused by the deletion of a T nucleotide in exon 18 of the FLO2 gene [51]. A recombinant inbred line (RIL) population with 127 lines was constructed from the cross between GM645 and japonica Tainung67 (TN67) with transparent endosperm. The F7 lines were harvested in Sanya, Hainan Province, in April 2021.
4.2. Preparation of Rice Flour
The paddy was dried in the sun until the moisture content was about 12%, and then stored at room temperature for three months. The rice grains were dehulled (Type THU, Satake Co., Tokyo, Japan), polished (Type TM05C, Satake Manufacturing, Suzhou, China), and then ground to flour (Cyclone Sample Mill, UDY Corporation, Fort Collins, CO, USA) to pass through a 100-mesh sieve.
4.3. DNA Extraction and Genotyping
Five rice seeds of each line and parents were germinated. Genomic DNA was extracted from the leaves of 10-day-old seedlings using the CTAB method. The deletion of 1 bp thymine on exon 18 of the FLO2 gene was detected using the method described in [51] with specific primers (5’TGAACCAGCGTAACGACATTGTG3’ and 5’TATGAAGAGAGTTACGGGGATTTATCTGACT3’). The PCR products were digested with restriction endonuclease HinfI for 1 h and separated on an 8% polyacrylamide gel for around 3h. The primers used to amplify the (CT)n microsatellite in the Wx gene were as follows: 5′CTTTGTCTCTCTCTCTCAGACAC3′ (484) and 5′TTGCAGATTTCTCTTCGATG3′ (485) [23,24]. The PCR products were also separated on the 8% polyacrylamide gels. For genotyping of the SSIIa allele, four of the allele-specific primers were used in a simple PCR (CGAGCGCACACACAG, GGCCGTGCAGATTAACCAT, CAAGAGAGGCTGGAGGGGC, and ACATGCGCACCTGGAAA) [28]. The PCR products were separated on a 2% agarose gel for around 2 h.
4.4. Apparent Amylose Content (AAC)
AAC was measured using a method described in Bao et al. [50]. The absorbance of the solution was measured at 620 nm using a Microplate spectrophotometer (Epoch, Biotek, Winooski, VT, USA). AAC was calculated using a standard curve made from five rice samples with known AAC.
4.5. Pasting Viscosity
The Pasting viscosity of rice flour was determined using Rapid visco analyser (RVA, Model 4500, Perten Instrument, Hägersten, Sweden). Three grams of rice flour (12% m.b.) were mixed with twenty-five grams of ddH2O in an aluminum RVA sample can. The heating and cooling cycle program was set as follows. The initial temperature was 50 °C. After 60 s, the sample was heated to 95 °C at a rate of 12 °C/60 s, maintained for 150 s, and then cooled to 50 °C with the cooling rate the same as the heating rate. The total program time was 12 min and 30 s. The starting speed of the RVA paddle was 960 rpm, which was reduced to 160 rpm after 15 s and maintained until the end of the program. The peak viscosity (PV), hot paste viscosity (HPV), cool paste viscosity (CPV), breakdown (BD, = PV – HPV), setback (SB, = CPV – PV), consistency (CS, = CPV – HPV) and pasting temperature (PT) were directly read out or calculated by TCW3 (Thermoline for Windows) software.
4.6. Gel textural Properties
The aluminum cans with rice flour gels were sealed by Parafilm after the RVA analysis and stored at 4 °C for 24 h. Texture characteristics were measured by a texture analyzer (TA.XTC-18, Shanghai Bosin Industrial Development Co., Shanghai, China) using a standard two cycle TPA program. A 5 mm diameter probe was used to compress the gel for 10 mm at 1 mm/s test speed. The hardness (HD, g), adhesiveness (ADH, g·s), and cohesiveness (COH) were derived from the software of the instrument.
4.7. Data Analysis
All the measurements were carried out at least in duplicate. Results were expressed as mean ± standard deviation (SD), and data were processed using analysis of variance (ANOVA) and multiple comparisons (Duncan’s multiple range test method) to determine significant differences by SAS (Version 9.3). The distribution of the data was determined, and principal component analysis was carried out using Origin 2017. The cluster analysis was carried out using the hclust function of the ggtree software package of R4.1.1 statistical software. The pie charts were created in Office 2019.
Author Contributions
Conceptualization, J.B.; investigation, Y.Z. (Yu Zhang), J.Z., Y.H., Y.Z. (Yanni Zhang), Y.Y., & F.X.; resources, J.B.; data curation, Y.Z. (Yu Zhang); writing—original draft preparation, Y.Z. (Yu Zhang) & J.B.; writing—review and editing, J.B.; funding acquisition, J.B. All authors have read and agreed to the published version of the manuscript.
Funding
The work was financially supported by the Zhejiang Provincial Natural Science Foundation (Grant No. LZ21C130003).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are contained within the article.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| AAC | Apparent amylose content |
| ADH | Gel adhesiveness |
| BD | Breakdown |
| CPV | Cold paste viscosity |
| COH | Gel cohesiveness |
| CS | Consistency viscosity |
| CEQ | Cooking and eating quality |
| FLO | Floury |
| HD | Gel hardness |
| GT | Gelatinization temperature |
| HPV | Hot paste viscosity |
| PC | Principal component |
| PCA | Principal component analysis |
| PT | Pasting temperature |
| PV | Peak viscosity |
| RIL | Recombinant inbred line |
| RVA | Rapid Visco-Analyzer |
| RVU | Rapid Visco Unit |
| SB | Setback |
| SNP | Single nucleotide polymorphism |
| SS | Soluble starch synthase |
| Wx | Waxy |
References
- Wing, R.A.; Purugganan, M.D.; Zhang, Q. The rice genome revolution: From an ancient grain to Green Super Rice. Nat. Rev. Genet. 2018, 19, 505–517. [Google Scholar] [CrossRef] [PubMed]
- Tian, Z.; Qian, Q.; Liu, Q.; Yan, M.; Liu, X.; Yan, C.; Liu, G.; Gao, Z.; Tang, S.; Zeng, D.; et al. Allelic diversities in rice starch biosynthesis lead to a diverse array of rice eating and cooking qualities. Proc. Natl. Acad. Sci. USA 2009, 106, 21760–21765. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q. Strategies for developing Green Super Rice. Proc. Natl. Acad. Sci. USA 2007, 104, 16402–16409. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.-H.; Zhuang, J.-Y.; Fan, Y.-Y.; Du, J.-H.; Cao, L.-Y. Progress in research and development on hybrid rice: A super-domesticate in China. Ann. Bot. 2007, 100, 959–966. [Google Scholar] [CrossRef]
- Bao, J. Toward understanding the genetic and molecular bases of the eating and cooking qualities of rice. Cereal Foods World 2012, 57, 148–156. [Google Scholar] [CrossRef]
- Bao, J.; Ying, Y.; Zhou, X.; Xu, Y.; Wu, P.; Xu, F.; Pang, Y. Relationships among starch biosynthesizing protein content, fine structure and functionality in rice. Carbohydr. Polym. 2020, 237, 116118. [Google Scholar] [CrossRef] [PubMed]
- Ayaad, M.; Han, Z.; Zheng, K.; Hu, G.; Abo-Yousef, M.; Sobeih, S.E.S.; Xing, Y. Bin-based genome-wide association studies reveal superior alleles for improvement of appearance quality using a 4-way MAGIC population in rice. J. Adv. Res. 2021, 28, 183–194. [Google Scholar] [CrossRef]
- Zhang, Z.; Hu, Y.; Zhao, J.; Zhang, Y.; Ying, Y.; Xu, F.; Bao, J. The role of different Wx and BEIIb allele combinations on fine structures and functional properties of indica rice starches. Carbohydr. Polym. 2022, 278, 118972. [Google Scholar] [CrossRef]
- Xu, Y.; Ying, Y.; Ouyang, S.; Duan, X.; Sun, H.; Jiang, S.; Sun, S.; Bao, J. Factors affecting sensory quality of cooked japonica rice. Rice Sci. 2018, 25, 330–339. [Google Scholar] [CrossRef]
- Huang, L.; Gu, Z.; Chen, Z.; Yu, J.; Chu, R.; Tan, H.; Zhao, D.; Fan, X.; Zhang, C.; Li, Q.; et al. Improving rice eating and cooking quality by coordinated expression of the major starch synthesis-related genes, SSII and Wx, in endosperm. Plant Mol. Biol. 2021, 106, 419–432. [Google Scholar] [CrossRef]
- Buléon, A.; Colonna, P.; Planchot, V.; Ball, S. Starch granules: Structure and biosynthesis. Int. J. Biol. Macromol. 1998, 23, 85–112. [Google Scholar] [CrossRef]
- Pérez, S.; Bertoft, E. The molecular structures of starch components and their contribution to the architecture of starch granules: A comprehensive review. Starch-Stärke 2010, 62, 389–420. [Google Scholar] [CrossRef]
- Nishi, A.; Nakamura, Y.; Tanaka, N.; Satoh, H. Biochemical and genetic analysis of the effects of amylose-extender mutation in rice endosperm. Plant Physiol. 2001, 127, 459–472. [Google Scholar] [CrossRef]
- Nakamura, Y. Towards a Better Understanding of the Metabolic System for Amylopectin Biosynthesis in Plants: Rice Endosperm as a Model Tissue. Plant Cell Physiol. 2002, 43, 718–725. [Google Scholar] [CrossRef]
- Jeon, J.-S.; Ryoo, N.; Hahn, T.-R.; Walia, H.; Nakamura, Y. Starch biosynthesis in cereal endosperm. Plant Physiol. Biochem. 2010, 48, 383–392. [Google Scholar] [CrossRef]
- Wanchana, S.; Toojinda, T.; Tragoonrung, S.; Vanavichit, A. Duplicated coding sequence in the waxy allele of tropical glutinous rice (Oryza sativa L.). Plant Sci. 2003, 165, 1193–1199. [Google Scholar] [CrossRef]
- Zhang, C.; Zhu, J.; Chen, S.; Fan, X.; Li, Q.; Lu, Y.; Wang, M.; Yu, H.; Yi, C.; Tang, S.; et al. Wxlv, the ancestral allele of rice Waxy gene. Mol. Plant 2019, 12, 1157–1166. [Google Scholar] [CrossRef] [PubMed]
- Mikami, I.; Uwatoko, N.; Ikeda, Y.; Yamaguchi, J.; Hirano, H.Y.; Suzuki, Y.; Sano, Y. Allelic diversification at the wx locus in landraces of Asian rice. Theor. Appl. Genet. 2008, 116, 979–989. [Google Scholar] [CrossRef]
- Sato, H.; Suzuki, Y.; Sakai, M.; Imbe, T. Molecular Characterization of Wx-mq, a Novel Mutant Gene for Low-amylose Content in Endosperm of Rice (Oryza sativa L.). Breed. Sci. 2002, 52, 131–135. [Google Scholar] [CrossRef]
- Yang, J.; Wang, J.; Fan, F.-J.; Zhu, J.-Y.; Chen, T.; Wang, C.-L.; Zheng, T.-Q.; Zhang, J.; Zhong, W.-G.; Xu, J.-L. Development of AS-PCR marker based on a key mutation confirmed by resequencing of Wx-mp in Milky Princess and its application in japonica soft rice (Oryza sativa L.) breeding. Plant Breed. 2013, 132, 595–603. [Google Scholar] [CrossRef]
- Cai, X.L.; Wang, Z.Y.; Xing, Y.Y.; Zhang, J.L.; Hong, M.M. Aberrant splicing of intron 1 leads to the heterogeneous 5′ UTR and decreased expression of waxy gene in rice cultivars of intermediate amylose content. Plant J. 1998, 14, 459–465. [Google Scholar] [CrossRef] [PubMed]
- Bligh, H.F.J.; Till, R.I.; Jones, C.A. A microsatellite sequence closely linked to the Waxy gene of Oryza sativa. Euphytica 1995, 86, 83–85. [Google Scholar] [CrossRef]
- Ayres, N.M.; McClung, A.M.; Larkin, P.D.; Bligh, H.F.J.; Jones, C.A.; Park, W.D. Microsatellites and a single-nucleotide polymorphism differentiate apparent amylose classes in an extended pedigree of US rice germ plasm. Theor. Appl. Genet. 1997, 94, 773–781. [Google Scholar] [CrossRef]
- Bao, J.S.; Corke, H.; Sun, M. Microsatellites, single nucleotide polymorphisms and a sequence tagged site in starch-synthesizing genes in relation to starch physicochemical properties in nonwaxy rice (Oryza sativa L.). Theor. Appl. Genet. 2006, 113, 1185–1196. [Google Scholar] [CrossRef]
- Gao, Z.; Zeng, D.; Cui, X.; Zhou, Y.; Yan, M.; Huang, D.; Li, J.; Qian, Q. Map-based cloning of the ALK gene, which controls the gelatinization temperature of rice. Sci. China Ser. C Life Sci. 2003, 46, 661–668. [Google Scholar] [CrossRef]
- Nakamura, Y.; Francisco, P.B.; Hosaka, Y.; Sato, A.; Sawada, T.; Kubo, A.; Fujita, N. Essential amino acids of starch synthase IIa differentiate amylopectin structure and starch quality between japonica and indica rice varieties. Plant Mol. Biol. 2005, 58, 213–227. [Google Scholar] [CrossRef] [PubMed]
- Bao, J.S.; Corke, H.; Sun, M. Nucleotide diversity in starch synthase IIa and validation of single nucleotide polymorphisms in relation to starch gelatinization temperature and other physicochemical properties in rice (Oryza sativa L.). Theor. Appl. Genet. 2006, 113, 1171–1183. [Google Scholar] [CrossRef]
- Jin, L.; Lu, Y.; Shao, Y.; Zhang, G.; Xiao, P.; Shen, S.; Corke, H.; Bao, J. Molecular marker assisted selection for improvement of the eating, cooking and sensory quality of rice (Oryza sativa L.). J. Cereal Sci. 2010, 51, 159–164. [Google Scholar] [CrossRef]
- Mo, Y.; Jeung, J.-U. The use of floury endosperm mutants to develop rice cultivars suitable for dry milling. Plant Biotechnol. Rep. 2020, 14, 185–191. [Google Scholar] [CrossRef]
- She, K.-C.; Kusano, H.; Koizumi, K.; Yamakawa, H.; Hakata, M.; Imamura, T.; Fukuda, M.; Naito, N.; Tsurumaki, Y.; Yaeshima, M.; et al. A novel factor FLOURY ENDOSPERM2 Is involved in regulation of rice grain size and starch quality. Plant Cell 2010, 22, 3280–3294. [Google Scholar] [CrossRef] [Green Version]
- Kang, H.G.; Park, S.; Matsuoka, M.; An, G. White-core endosperm floury endosperm-4 in rice is generated by knockout mutations in the C-type pyruvate orthophosphate dikinase gene (OsPPDKB). Plant J. 2005, 42, 901–911. [Google Scholar] [CrossRef] [PubMed]
- Ryoo, N.; Yu, C.; Park, C.-S.; Baik, M.-Y.; Park, I.M.; Cho, M.-H.; Bhoo, S.H.; An, G.; Hahn, T.-R.; Jeon, J.-S. Knockout of a starch synthase gene OsSSIIIa/Flo5 causes white-core floury endosperm in rice (Oryza sativa L.). Plant Cell Rep. 2007, 26, 1083–1095. [Google Scholar] [CrossRef]
- Fujita, N.; Yoshida, M.; Kondo, T.; Saito, K.; Utsumi, Y.; Tokunaga, T.; Nishi, A.; Satoh, H.; Park, J.-H.; Jane, J.-L.; et al. Characterization of SSIIIa-deficient mutants of rice: The function of SSIIIa and pleiotropic effects by SSIIIa deficiency in the rice endosperm. Plant Physiol. 2007, 144, 2009–2023. [Google Scholar] [CrossRef] [PubMed]
- Peng, C.; Wang, Y.; Liu, F.; Ren, Y.; Zhou, K.; Lv, J.; Zheng, M.; Zhao, S.; Zhang, L.; Wang, C.; et al. FLOURY ENDOSPERM6 encodes a CBM48 domain-containing protein involved in compound granule formation and starch synthesis in rice endosperm. Plant J. 2014, 77, 917–930. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Ren, Y.; Lu, B.; Yang, C.; Feng, Z.; Liu, Z.; Chen, J.; Ma, W.; Wang, Y.; Yu, X.; et al. FLOURY ENDOSPERM7 encodes a regulator of starch synthesis and amyloplast development essential for peripheral endosperm development in rice. J. Exp. Bot. 2016, 67, 633–647. [Google Scholar] [CrossRef]
- Long, W.; Dong, B.; Wang, Y.; Pan, P.; Wang, Y.; Liu, L.; Chen, X.; Liu, X.; Liu, S.; Tian, Y.; et al. FLOURY ENDOSPERM8, encoding the UDP-glucose pyrophosphorylase 1, affects the synthesis and structure of starch in rice endosperm. J. Plant Biol. 2017, 60, 513–522. [Google Scholar] [CrossRef]
- Wu, M.; Ren, Y.; Cai, M.; Wang, Y.; Zhu, S.; Zhu, J.; Hao, Y.; Teng, X.; Zhu, X.; Jing, R.; et al. Rice FLOURY ENDOSPERM10 encodes a pentatricopeptide repeat protein that is essential for the trans-splicing of mitochondrial nad1 intron 1 and endosperm development. New Phytol. 2019, 223, 736–750. [Google Scholar] [CrossRef]
- Zhu, X.; Teng, X.; Wang, Y.; Hao, Y.; Jing, R.; Wang, Y.; Liu, Y.; Zhu, J.; Wu, M.; Zhong, M.; et al. FLOURY ENDOSPERM11 encoding a plastid heat shock protein 70 is essential for amyloplast development in rice. Plant Sci. 2018, 277, 89–99. [Google Scholar] [CrossRef]
- Zhong, M.; Liu, X.; Liu, F.; Ren, Y.; Wang, Y.; Zhu, J.; Teng, X.; Duan, E.; Wang, F.; Zhang, H.; et al. FLOURY ENDOSPERM12 encoding alanine aminotransferase 1 regulates carbon and nitrogen metabolism in rice. J. Plant Biol. 2019, 62, 61–73. [Google Scholar] [CrossRef]
- Hu, T.; Tian, Y.; Zhu, J.; Wang, Y.; Jing, R.; Lei, J.; Sun, Y.; Yu, Y.; Li, J.; Chen, X.; et al. OsNDUFA9 encoding a mitochondrial complex I subunit is essential for embryo development and starch synthesis in rice. Plant Cell Rep. 2018, 37, 1667–1679. [Google Scholar] [CrossRef]
- Xue, M.; Liu, L.; Yu, Y.; Zhu, J.; Gao, H.; Wang, Y.; Wan, J. Lose-of-function of a rice nucleolus-localized pentatricopeptide repeat protein is responsible for the floury endosperm14 mutant phenotypes. Rice 2019, 12, 100. [Google Scholar] [CrossRef]
- You, X.; Zhang, W.; Hu, J.; Jing, R.; Cai, Y.; Feng, Z.; Kong, F.; Zhang, J.; Yan, H.; Chen, W.; et al. FLOURY ENDOSPERM15 encodes a glyoxalase I involved in compound granule formation and starch synthesis in rice endosperm. Plant Cell Rep. 2019, 38, 345–359. [Google Scholar] [CrossRef] [PubMed]
- Teng, X.; Zhong, M.; Zhu, X.; Wang, C.; Ren, Y.; Wang, Y.; Zhang, H.; Jiang, L.; Wang, D.; Hao, Y.; et al. FLOURY ENDOSPERM16 encoding a NAD-dependent cytosolic malate dehydrogenase plays an important role in starch synthesis and seed development in rice. Plant Biotechnol. J. 2019, 17, 1914–1927. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Wu, M.; Ren, Y.; Wang, Y.; Li, J.; Lei, C.; Sun, Y.; Bao, X.; Wu, H.; Yang, H.; et al. Rice FLOURY ENDOSPERM 18 encodes a pentatricopeptide repeat protein required for 5′ processing of mitochondrial nad5 messenger RNA and endosperm development. J. Integr. Plant Biol. 2021, 63, 834–847. [Google Scholar] [CrossRef] [PubMed]
- Lou, G.; Chen, P.; Zhou, H.; Li, P.; Xiong, J.; Wan, S.; Zheng, Y.; Alam, M.; Liu, R.; Zhou, Y.; et al. FLOURY ENDOSPERM19 encoding a class I glutamine amidotransferase affects grain quality in rice. Mol. Breed. 2021, 41, 36. [Google Scholar] [CrossRef]
- Wu, Y.; Pu, C.; Lin, H.; Huang, H.; Huang, Y.; Hong, C.; Chang, M.; Lin, Y. Three novel alleles of FLOURY ENDOSPERM2 (FLO2) confer dull grains with low amylose content in rice. Plant Sci. 2015, 233, 44–52. [Google Scholar] [CrossRef]
- Qiao, Y.; Lee, S.I.; Piao, R.; Jiang, W.; Ham, T.H.; Chin, J.H.; Piao, Z.; Han, L.; Kang, S.Y.; Koh, H.J. Fine mapping and candidate gene analysis of the floury endosperm gene, FLO(a), in rice. Mol. Cells 2010, 29, 167–174. [Google Scholar] [CrossRef]
- Kong, X.; Sun, X.; Xu, F.; Umemoto, T.; Chen, H.; Bao, J. Morphological and physicochemical properties of two starch mutants induced from a high amylose indica rice by gamma irradiation. Starch-Stärke 2014, 66, 157–165. [Google Scholar] [CrossRef]
- Xiang, X.; Kang, C.; Xu, S.; Yang, B. Combined effects of Wx and SSIIa haplotypes on rice starch physicochemical properties. J. Sci. Food Agric. 2017, 97, 1229–1234. [Google Scholar] [CrossRef]
- Bao, J.S.; Jin, L.; Xiao, P.; Shen, S.; Sun, M.; Corke, H. Starch physicochemical properties and their associations with microsatellite alleles of starch-synthesizing genes in a rice RIL population. J. Agric. Food Chem. 2008, 56, 1589–1594. [Google Scholar] [CrossRef]
- Bao, J.S.; Zhang, Y.; Zhao, J.J.; Chen, Y.L.; Wu, W.X.; Cao, L.Y.; Xu, F.F. Identification of a new allele of FLOURY ENDOSPERM2 in a white-core endosperm mutant of rice. Rice Sci. 2022, 29, 407–411. [Google Scholar] [CrossRef]
- Zhang, H.; Xu, H.; Jiang, Y.; Zhang, H.; Wang, S.; Wang, F.; Zhu, Y. Genetic control and high temperature effects on starch biosynthesis and grain quality in rice. Front. Plant Sci. 2021, 12, 2971. [Google Scholar] [CrossRef]
- Huang, L.; Tan, H.; Zhang, C.; Li, Q.; Liu, Q. Starch biosynthesis in cereal endosperms: An updated review over the last decade. Plant Commun. 2021, 2, 100237. [Google Scholar] [CrossRef] [PubMed]
- Qiu, J. Effects of Flo2 with Different Mutation Sites on Synthesis of Storage Substances in Rice Endosperm. Master’s Thesis, Yangzhou University, Yangzhou, China, 2021. [Google Scholar]
- Das, A.K.; Cohen, P.T.W.; Barford, D. The structure of the tetratricopeptide repeats of protein phosphatase 5: Implications for TPR-mediated protein–protein interactions. EMBO J. 1998, 17, 1192–1199. [Google Scholar] [CrossRef] [PubMed]
- Blatch, G.L.; Lässle, M. The tetratricopeptide repeat: A structural motif mediating protein-protein interactions. BioEssays 1999, 21, 932–939. [Google Scholar] [CrossRef]
- Sen, A.; Kalvakuri, S.; Bodmer, R.; Cox, R.T. Clueless, a protein required for mitochondrial function, interacts with the PINK1-Parkin complex in Drosophila. Dis. Models Mech. 2015, 8, 577–589. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhao, L.; Lin, L.; Zhao, L.; Liu, Q.; Wei, C. A novel mutation of OsPPDKB, encoding pyruvate orthophosphate dikinase, affects metabolism and structure of starch in the rice endosperm. Int. J. Mol. Sci. 2018, 19, 2268. [Google Scholar] [CrossRef]
- Kowittaya, C.; Lumdubwong, N. Molecular weight, chain profile of rice amylopectin and starch pasting properties. Carbohydr. Polym. 2014, 108, 216–223. [Google Scholar] [CrossRef]
- Tao, K.; Li, C.; Yu, W.; Gilbert, R.G.; Li, E. How amylose molecular fine structure of rice starch affects functional properties. Carbohydr. Polym. 2019, 204, 24–31. [Google Scholar] [CrossRef]
- Wang, K.; Hasjim, J.; Wu, A.C.; Henry, R.J.; Gilbert, R.G. Variation in amylose fine structure of starches from different botanical sources. J. Agric. Food Chem. 2014, 62, 4443–4453. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Pang, Y.; Bao, J. Expression profiles and protein complexes of starch biosynthetic enzymes from white-core and waxy mutants induced from high amylose indica rice. Rice Sci. 2020, 27, 152–161. [Google Scholar] [CrossRef]
- Huang, J.; Tang, X.; Pu, H. Research progress on the micro-structure, texture property and stability of starch gel. J. Food Sci. Biotechnol. 2017, 36, 673–679. [Google Scholar]
- Pfister, B.; Zeeman, S.C. Formation of starch in plant cells. Cell. Mol. Life Sci. 2016, 73, 2781–2807. [Google Scholar] [CrossRef] [PubMed]
- Ball, S.G.; Morell, M.K. From bacterial glycogen to starch: Understanding the biogenesis of the plant starch granule. Annu. Rev. Plant Biol. 2003, 54, 207–233. [Google Scholar] [CrossRef]
- Zeeman, S.C.; Kossmann, J.; Smith, A.M. Starch: Its metabolism, evolution, and biotechnological modification in plants. Annu. Rev. Plant Biol. 2010, 61, 209–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Distribution of apparent amylose content and RVA spectrum eigenvalues in RIL.

Figure 2.
Principal component analysis of rice quality properties.
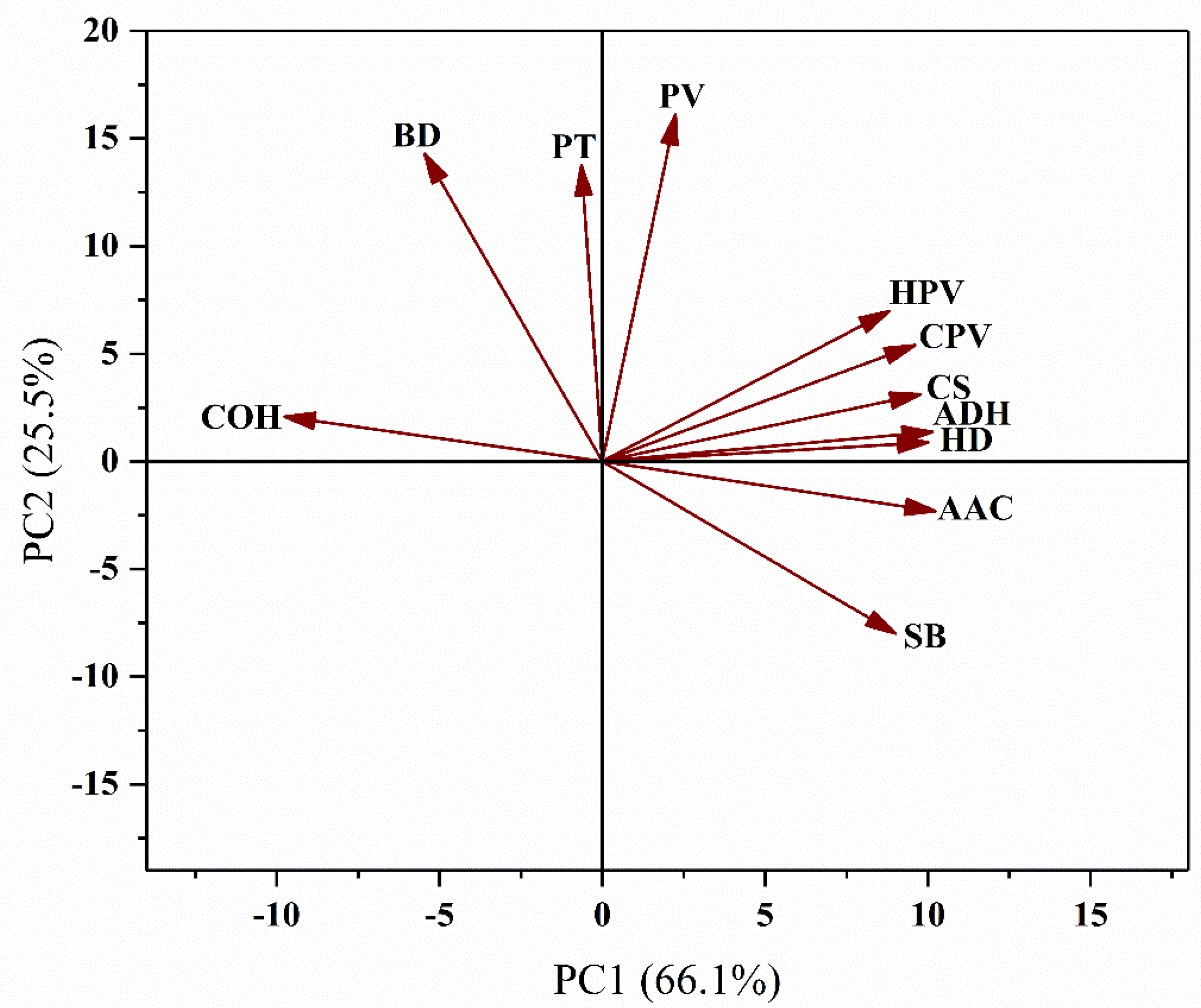
Figure 3.
Genotyping of FLO2, Wx, and SSIIa alleles in RIL. M—DNA marker; bp—base pair.

Figure 4.
Percentages of variance of starch properties in RIL population explained by the FLO2, Wx and SSIIa interactions.
Figure 4.
Percentages of variance of starch properties in RIL population explained by the FLO2, Wx and SSIIa interactions.

Figure 5.
Cluster analysis of 11 quality traits in RIL population. The red part represents Class A, including 59 RIL lines involving four allele combinations of FLO2/Wxa/SSIIa(GC), FLO2/Wxa/SSIIa(TT), flo2/Wxa/SSIIa(GC), and flo2/Wxa/SSIIa(TT). The blue part represents Class B, including 68 RIL lines involving four allele combinations of FLO2/Wxb/SSIIa(GC), FLO2/Wxb/SSIIa(TT), FLO2/Wxb/SSIIa(GC), and FLO2/Wxb/SSIIa(TT).
Figure 5.
Cluster analysis of 11 quality traits in RIL population. The red part represents Class A, including 59 RIL lines involving four allele combinations of FLO2/Wxa/SSIIa(GC), FLO2/Wxa/SSIIa(TT), flo2/Wxa/SSIIa(GC), and flo2/Wxa/SSIIa(TT). The blue part represents Class B, including 68 RIL lines involving four allele combinations of FLO2/Wxb/SSIIa(GC), FLO2/Wxb/SSIIa(TT), FLO2/Wxb/SSIIa(GC), and FLO2/Wxb/SSIIa(TT).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Rice-quality properties of parents and RIL.
| Parameter | Parents | RIL | |||||
|---|---|---|---|---|---|---|---|
| GM645 | TN67 | Mean ± SD | CV (%) | Range | Skewness | Kurtosis | |
| AAC (%) | 22.6 A | 16.3 B | 18 ± 6.8 | 37.8 | 6.7–29.1 | 0.2 | −1.5 |
| PV (RUV) | 215.3 B | 269.9 A | 246.5 ± 56.4 | 22.9 | 96.5–351.3 | −0.4 | −0.7 |
| HPV (RUV) | 176.7 A | 171.1 A | 163.7 ± 43.1 | 26.3 | 62.3–281.7 | 0.1 | −0.3 |
| CPV (RUV) | 279.1 A | 262.9 A | 271.7 ± 69.8 | 25.7 | 115.4–412.7 | −0.3 | −0.6 |
| BD (RUV) | 38.6 A | 98.8 A | 82.8 ± 44.0 | 53.1 | 11.3–198.2 | −0.3 | −0.5 |
| SB (RUV) | 63.8 A | −7.0 B | 25.2 ± 62.5 | 247.8 | −121.1 to 135.4 | −0.3 | −0.9 |
| PT (°C) | 72.8 B | 73.6 A | 75.5 ± 4.6 | 6.1 | 66.9–83.9 | 0.0 | −1.2 |
| CS (RUV) | 102.4 A | 91.8 A | 108.0 ± 34.9 | 32.4 | 34.7–182.6 | 0.1 | −0.6 |
| HD (g) | 11.5 A | 7.5 B | 11.6 ± 10.4 | 89.7 | 1.37–52.72 | 1.5 | 1.9 |
| ADH (g·s) | 24.8 A | 10.7 B | 23.2 ± 20.6 | 88.8 | 0.00–103.4 | 1.5 | 1.9 |
| COH | 0.58 B | 0.76 A | 0.61 ± 0.08 | 13.1 | 0.45–0.96 | 0.5 | 1.7 |
Different letters after the values between parents represent significant differences (p < 0.05). SD—standard deviation; CV—coefficient of variation.
Table 2.
Effects of different alleles of FLO2, Wx and SSIIa on rice quality properties.
| Parameter | FLO2 | Wx | SSIIa | |||
|---|---|---|---|---|---|---|
| FLO2 (86) | flo2 (41) | Wxa (59) | Wxb (68) | TT (50) | GC (77) | |
| AAC (%) | 19.2 ± 7.1 A | 15.6 ± 5.2 B | 24.6 ± 3.1 A | 12.3 ± 2.6B | 18.7 ± 6.7 A | 17.6 ± 6.7 A |
| PV (RUV) | 274.2 ± 40.4 A | 188.4 ± 37.8 B | 239.0 ± 46.7 A | 253.8 ± 62.9 A | 248.8 ± 60.9 A | 245.1 ± 53.2 A |
| HPV (RUV) | 180.6 ± 37.7 A | 128.4 ± 30.4 B | 188.1 ± 37.5 A | 142.6 ± 35.9 B | 175.4 ± 50.5 A | 156.2 ± 35.6 B |
| CPV (RUV) | 93.6 ± 43.8 A | 60.0 ± 34.7 B | 50.9 ± 22.9 B | 110.5 ± 38.9 A | 73.4 ± 40.7 A | 88.9 ± 45.0 A |
| BD (RUV) | 300.2 ± 57.3 A | 212.0 ± 54.2 B | 321.8 ± 46.8 A | 228.3 ± 56.1 B | 284.0 ± 71.2 A | 263.8 ± 67.8 A |
| SB (RUV) | 26.0 ± 64.8 A | 23.6 ± 57.3 A | 82.8 ± 21.1 A | −24.8 ± 39.1 B | 35.2 ± 54.1 A | 18.7 ± 66.6 A |
| PT (°C) | 76.6 ± 4.7 A | 73.3 ± 3.7 B | 74.6 ± 4.1 B | 76.3 ± 4.9 A | 71.0 ± 2.3 B | 78.4 ± 3.2 A |
| CS (RUV) | 119.6 ± 31.2 A | 83.7 ± 29.4 B | 133.7 ± 27.6 A | 85.7 ± 23.5 B | 108.6 ± 30.9 A | 107.6 ± 37.3 A |
| HD (g) | 13.9 ± 11.5 A | 6.8 ± 4.2 B | 19.7 ± 10.3 A | 4.5 ± 1.0 B | 11.1 ± 9.3 A | 11.9 ± 10.9 A |
| ADH (g·s) | 27.9 ± 22.5 A | 13.3 ± 10.3 B | 38.8 ± 20.9 A | 9.7 ± 4.2 B | 22.7 ± 18.5 A | 23.5 ± 21.8 A |
| COH | 0.60 ± 0.08 A | 0.63 ± 0.08 A | 0.55 ± 0.06 B | 0.65 ± 0.06 A | 0.61 ± 0.07 A | 0.60 ± 0.08 A |
The number after the genotype indicates the number of lines with this allele in the RIL population. Different letters after the values between two alleles of the same gene represent significant differences (p < 0.05).
Table 3.
Differences in rice quality traits in eight combinations of FLO2, Wx, and SSIIa.
| Genetic Combination | AAC (%) | PV (RUV) | HPV (RUV) | BD (RUV) | CPV (RUV) | SB (RUV) |
|---|---|---|---|---|---|---|
| FLO2/Wxa/SSIIa(GC) | 25.70 ± 2.07 A | 264.5 ± 28.6 A | 193.3 ± 23.5 AB | 71.2 ± 17.8 D | 345.4 ± 25.1 A | 80.9 ± 22.7 A |
| FLO2/Wxa/SSIIa(TT) | 26.23 ± 2.67 A | 257.9 ± 38.0 A | 214.5 ± 38.7 A | 43.4 ± 13.1 EF | 343.5 ± 40.6 A | 85.6 ± 24.4 A |
| FLO2/Wxb/SSIIa(GC) | 11.30 ± 1.94 D | 283.3 ± 48.3 A | 147.3 ± 21.7 D | 136.0 ± 35.1 A | 239.0 ± 33.8 C | −44.4 ± 39.7 D |
| FLO2/Wxb/SSIIa(TT) | 14.41 ± 2.45 C | 289.7 ± 35.4 A | 174.8 ± 30.7 BC | 114.9 ± 27.6 B | 280.5 ± 38.7 B | −9.2 ± 31.9 BC |
| flo2/Wxa/SSIIa(GC) | 20.89 ± 1.62 B | 184.8 ± 22.2 BC | 154.2 ± 24.0 CD | 30.6 ± 10.5 F | 262.8 ± 24.1 BC | 78.1 ± 11.1 A |
| flo2/Wxa/SSIIa(TT) | 22.20 ± 1.59 B | 176.8 ± 13.0 BC | 148.1 ± 10.4 D | 28.6 ± 11.4 F | 274.1 ± 14.2 B | 97.4 ± 6.0 A |
| flo2/Wxb/SSIIa(GC) | 12.01 ± 2.39 D | 207.5 ± 37.0 B | 115.5 ± 15.9 E | 92.0 ± 27.4 C | 179.4 ± 30.3 D | −28.1 ± 36.0 BC |
| flo2/Wxb/SSIIa(TT) | 10.87 ± 2.08 D | 161.9 ± 46.8 C | 102.2 ± 30.8 E | 59.7 ± 21.8 DE | 163.7 ± 40.1 D | 1.7 ± 25.3 B |
| Genetic Combination | PT (°C) | CS (RUV) | HD (g) | ADH (g·s) | COH | |
| FLO2/Wxa/SSIIa(GC) | 79.0 ± 1.4 B | 152.1 ± 19.6 A | 25.14 ± 10.38 A | 48.68 ± 21.91 A | 0.52 ± 0.04 E | |
| FLO2/Wxa/SSIIa(TT) | 70.7 ± 1.8 E | 129.0 ± 29.0 B | 21.21 ± 7.78 A | 41.95 ± 16.23 A | 0.55 ± 0.05 DE | |
| FLO2/Wxb/SSIIa(GC) | 81.7 ± 1.2 A | 91.7 ± 16.0 D | 4.70 ± 0.91 CD | 10.25 ± 2.58 CD | 0.66 ± 0.04 AB | |
| FLO2/Wxb/SSIIa(TT) | 72.8 ± 1.6 D | 105.7 ± 13.6 CD | 4.72 ± 0.76 CD | 11.59 ± 3.60 CD | 0.67 ± 0.04 A | |
| flo2/Wxa/SSIIa(GC) | 73.8 ± 1.2 D | 108.7 ± 14.3 C | 9.64 ± 3.45 BC | 19.96 ± 9.77 BC | 0.61 ± 0.07 BC | |
| flo2/Wxa/SSIIa(TT) | 68.7 ± 0.9 F | 126.0 ± 15.5 B | 12.70 ± 5.46 B | 25.87 ± 10.99 B | 0.58 ± 0.05CD | |
| flo2/Wxb/SSIIa(GC) | 76.5 ± 2.0 C | 63.9 ± 18.0 E | 4.48 ± 1.34 CD | 8.49 ± 5.66 D | 0.64 ± 0.10AB | |
| flo2/Wxb/SSIIa(TT) | 68.2 ± 1.0 F | 61.4 ± 12.3 E | 3.71 ± 0.52 D | 5.65 ± 2.36 D | 0.65 ± 0.06AB |
Different letters after the values in each column indicated significant differences (p < 0.05).
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zhang, Y.; Zhao, J.; Hu, Y.; Zhang, Y.; Ying, Y.; Xu, F.; Bao, J. Combined Effects of Different Alleles of FLO2, Wx and SSIIa on the Cooking and Eating Quality of Rice. Plants 2022, 11, 2249. https://doi.org/10.3390/plants11172249
AMA Style
Zhang Y, Zhao J, Hu Y, Zhang Y, Ying Y, Xu F, Bao J. Combined Effects of Different Alleles of FLO2, Wx and SSIIa on the Cooking and Eating Quality of Rice. Plants. 2022; 11(17):2249. https://doi.org/10.3390/plants11172249
Chicago/Turabian StyleZhang, Yu, Jiajia Zhao, Yaqi Hu, Yanni Zhang, Yining Ying, Feifei Xu, and Jinsong Bao. 2022. "Combined Effects of Different Alleles of FLO2, Wx and SSIIa on the Cooking and Eating Quality of Rice" Plants 11, no. 17: 2249. https://doi.org/10.3390/plants11172249
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.