
Arabidopsis thaliana Accessions from the Chernobyl Exclusion Zone Show Decreased Sensitivity to Additional Acute Irradiation
 , , , ,
, , , ,  , , and
, , and
Abstract
:1. Introduction
2. Results
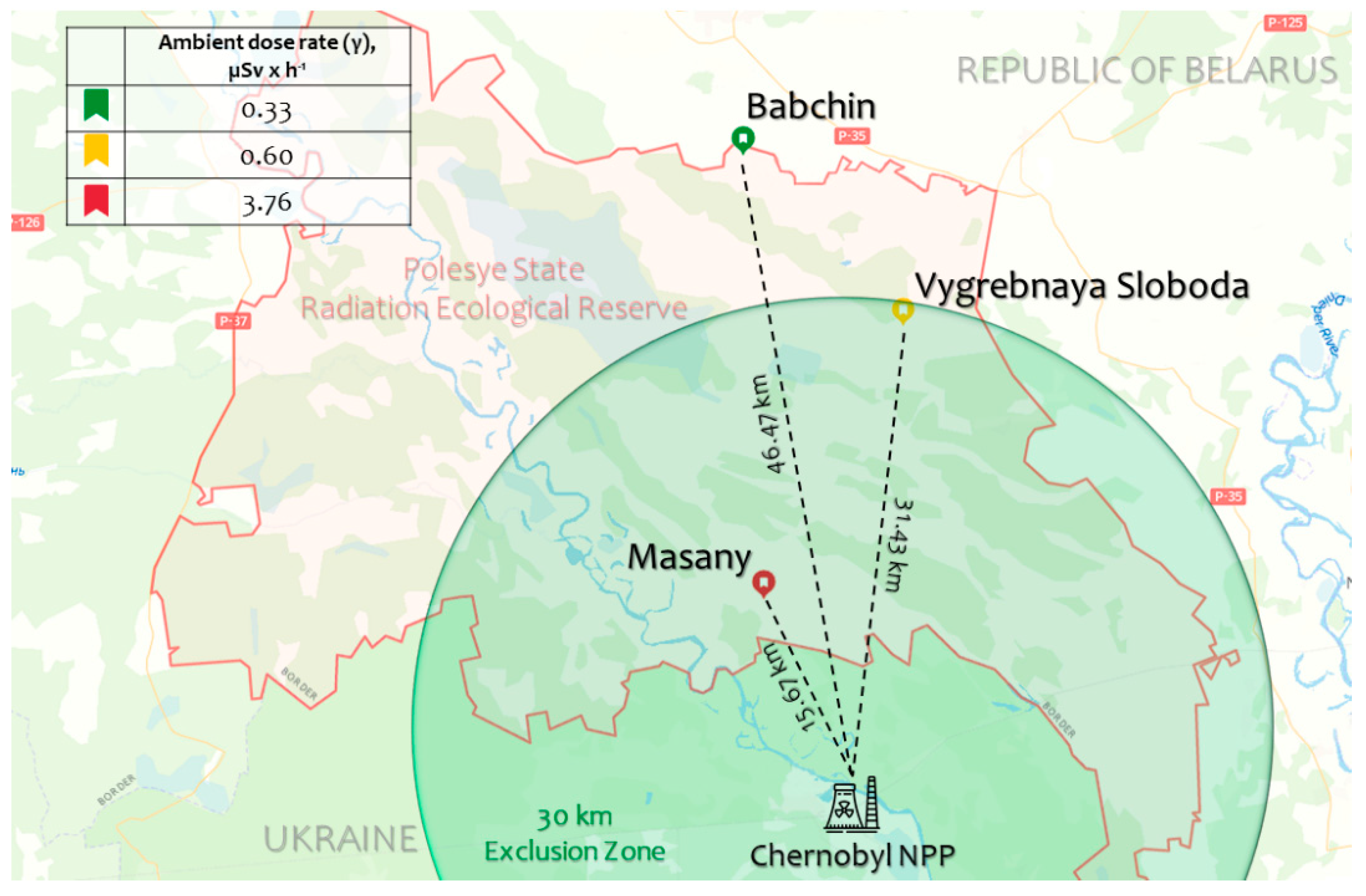
2.1. Arabidopsis Natural Accessions in the Chernobyl Exclusion Zone
2.2. Germination Dynamics Analysis
2.3. Leaf Area Analysis
2.4. Photosynthetic Parameters
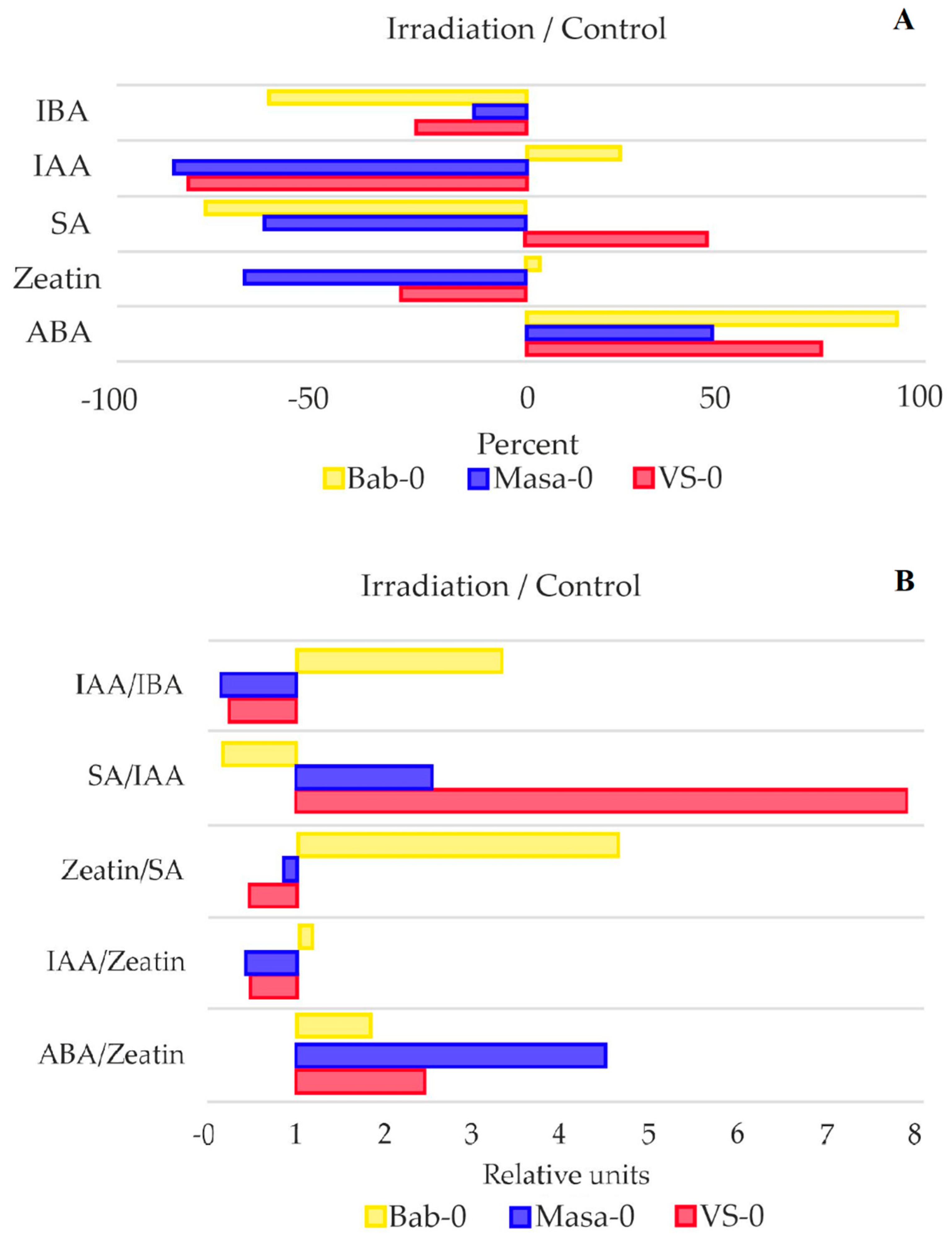
2.5. Phytohormone Measurements
2.6. Differential Gene Expression Analysis of Control-Grown Chernobyl Natural Accessions
2.7. Differential Gene Expression Analysis of Chernobyl A. thaliana Accessions Recovering from Acute High-Dose γ-Irradiation
3. Discussion
3.1. Growth and Physiological Responses of Chernobyl Accessions to Chronic and Acute Irradiation
3.2. Transcriptional Responses of Natural Accessions Recovering from Chronic Radiation Exposure
3.3. Transcriptional Responses of Natural Accessions to Acute Radiation Exposure
4. Conclusions
5. Materials and Methods
5.1. Sampling in the Chernobyl Exclusion Zone
5.2. Acute γ-Irradiation of Seeds
5.3. Growth Conditions and Germination Assay
5.4. Leaf Area and Fluorescence Measurements
5.5. Processing of Morphological and Fluorescence Data
5.6. Phytohormone Measurement
5.7. RNA Extraction
5.8. Illumina Sequencing
5.9. Data Processing and Functional Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Van Hoeck, A.; Horemans, N.; Nauts, R.; Van Hees, M.; Vandenhove, H.; Blust, R. Lemna minor plants chronically exposed to ionising radiation: RNA-seq analysis indicates a dose rate dependent shift from acclimation to survival strategies. Plant Sci. 2017, 257, 84–95. [Google Scholar] [CrossRef] [PubMed]
- Grinberg, M.A.; Gudkov, S.A.; Balalaeva, I.V.; Gromova, E.; Sinitsyna, Y.; Sukhov, V.; Vodeneev, V. Effect of chronic β-radiation on long-distance electrical signals in wheat and their role in adaptation to heat stress. Environ. Exp. Bot. 2021, 184, 104378. [Google Scholar] [CrossRef]
- Volkova, P.Y.; Duarte, G.T.; Kazakova, E.A.; Makarenko, E.S.; Bitarishvili, S.V.; Bondarenko, V.S.; Perevolotskii, A.N.; Geras’Kin, S.A.; Garbaruk, D.K.; Turchin, L.M. Radiosensitivity of herbaceous plants to chronic radiation exposure: Field study in the Chernobyl exclusion zone. Sci. Total Environ. 2021, 777, 146206. [Google Scholar] [CrossRef]
- Esnault, M.-E.; Legue, F.; Chenal, C. Ionizing radiation: Advances in plant response. Environ. Exp. Bot. 2010, 68, 231–237. [Google Scholar] [CrossRef]
- Gudkov, S.V.; Grinberg, M.A.; Sukhov, V.; Vodeneev, V. Effect of ionizing radiation on physiological and molecular processes in plants. J. Environ. Radioact. 2019, 202, 8–24. [Google Scholar] [CrossRef]
- Caplin, N.; Willey, N. Ionizing radiation, higher plants, and radioprotection: From acute high doses to chronic low doses. Front. Plant Sci. 2018, 9, 847. [Google Scholar] [CrossRef]
- Volkova, P.Y.; Geraskin, S.A. ‘Omic’ technologies as a helpful tool in radioecological research. J. Environ. Radioact. 2018, 189, 156–167. [Google Scholar] [CrossRef]
- Chang, S.; Lee, U.; Hong, M.J.; Jo, Y.D.; Kim, J.-B. High-throughput phenotyping (HTP) data reveal dosage effect at growth stages in Arabidopsis thaliana irradiated by gamma rays. Plants 2020, 9, 557. [Google Scholar] [CrossRef]
- Abramov, V.I.; Fedorenko, O.M.; Shevchenko, V.A. Genetic consequences of radioactive contamination for populations of Arabidopsis. Sci. Total Environ. 1992, 112, 19–28. [Google Scholar] [CrossRef]
- Kovalchuk, I.; Abramov, V.; Pogribny, I.; Kovalchuk, O. Molecular aspects of plant adaptation to life in the Chernobyl zone. Plant Physiol. 2004, 135, 357–363. [Google Scholar] [CrossRef]
- Horemans, N.; Nauts, R.; Battle, J.V.; Van Hees, M.; Jacobs, G.; Voorspoels, S.; Gaschak, S.; Nanba, K.; Saenen, E. Genome-wide DNA methylation changes in two Brassicaceae species sampled alongside a radiation gradient in Chernobyl and Fukushima. J. Environ. Radioact. 2018, 192, 405–416. [Google Scholar] [CrossRef] [PubMed]
- Morozova, V.; Kashparova, E.; Levchuk, S.; Bishchuk, Y.; Kashparov, V. The progeny of Chernobyl Arabidopsis thaliana plants does not exhibit changes in morphometric parameters and cellular antioxidant defence system of shoots. J. Environ. Radioact. 2020, 211, 106076. [Google Scholar] [CrossRef] [PubMed]
- Klimenko, O.; Pernis, M.; Danchenko, M.; Skultéty, L.; Klubicová, K.; Shevchenko, G. Natural ecotype of Arabidopsis thaliana (L.) Heynh (Chernobyl-07) respond to cadmium stress more intensively than the sensitive ecotypes Oasis and Columbia. Ecotoxicol. Environ. Saf. 2019, 30, 86–95. [Google Scholar] [CrossRef] [PubMed]
- Food and Agriculture Organization of the United Nations (FAO). Micronutrients. Assessment at the Country Level; Soil Bulletin No. 63; FAO: Roma, Italy, 1990. [Google Scholar]
- Gupta, D.K.; Tiwari, S.; Chatterjee, S.; Walther, C. Potassium and its role in cesium transport in plants. Biologia 2018, 73, 885–896. [Google Scholar] [CrossRef]
- Bilbro, J.D.; Wanjura, D.F. Soil Crusts and Cotton Emergence Relationships. Trans. ASAE 1982, 25, 1484–1487. [Google Scholar] [CrossRef]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence—A practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef]
- Kramer, D.M.; Johnson, G.; Kiirats, O.; Edwards, G.E. New fluorescence parameters for the determination of QA redox state and excitation energy fluxes. Photosynth. Res. 2004, 79, 209. [Google Scholar] [CrossRef]
- Buschmann, C. Photochemical and non-photochemical quenching coefficients of the chlorophyll fluorescence: Comparison of variation and limits. Photosynthetica 1999, 37, 217–224. [Google Scholar] [CrossRef]
- Fernández-Marín, B.; Gulías, J.; Figueroa, C.M.; Iñiguez, C.; Clemente-Moreno, M.J.; Nunes-Nesi, A.; Fernie, A.R.; Cavieres, L.A.; Bravo, L.; García-Plazaola, J.I.; et al. How do vascular plants perform photosynthesis in extreme environments? An integrative ecophysiological and biochemical story. Plant J. 2020, 101, 979–1000. [Google Scholar] [CrossRef]
- Wani, S.H.; Kumar, V.; Shriram, V.; Sah, S.K. Phytohormones and their metabolic engineering for abiotic stress tolerance in crop plants. Crop J. 2016, 4, 162–176. [Google Scholar] [CrossRef]
- Verma, V.; Ravindran, P.; Kumar, P.P. Plant hormone-mediated regulation of stress responses. BMC Plant Biol. 2016, 16, 86. [Google Scholar] [CrossRef] [PubMed]
- Volkova, P.Y.; Duarte, G.T.; Soubigou-Taconnat, L.; Kazakova, E.A.; Pateyron, S.; Bondarenko, V.S.; Bitarishvili, S.V.; Makarenko, E.S.; Churyukin, R.S.; Lychenkova, M.A.; et al. Early response of barley embryos to low- and high-dose gamma irradiation of seeds triggers changes in the transcriptional profile and an increase in hydrogen peroxide content in seedlings. J. Agron. Crop Sci. 2019, 206, 277–295. [Google Scholar] [CrossRef]
- Gruszka, D. Crosstalk of the brassinosteroid signalosome with phytohormonal and stress signaling components maintains a balance between the processes of growth and stress tolerance. Int. J. Mol. Sci. 2018, 19, 2675. [Google Scholar] [CrossRef]
- Podlešáková, K.; Ugena, L.; Spíchal, L.; Doležal, K.; De Diego, N. Phytohormones and polyamines regulate plant stress responses by altering GABA pathway. New Biotechnol. 2019, 48, 53–65. [Google Scholar] [CrossRef]
- Samsonova, Z.; Kiran, N.S.; Novak, O.; Spyroglou, I.; Skalák, J.; Hejátko, J.; Gloser, V. Steady-state levels of cytokinins and their derivatives may serve as a unique classifier of Arabidopsis ecotypes. Plants 2020, 9, 116. [Google Scholar] [CrossRef] [PubMed]
- Wan, J.S.H.; Pang, C.K.; Bonser, S.P. Does the cost of adaptation to extremely stressful environments diminish over time? A literature synthesis on how plants adapt to heavy metals and pesticides. Evol. Biol. 2017, 44, 411–426. [Google Scholar] [CrossRef]
- De Micco, V.; Arena, C.; Pignalosa, D.; Durante, M. Effects of sparsely and densely ionizing radiation on plants. Radiat. Environ. Biophys. 2011, 50, 1–19. [Google Scholar] [CrossRef]
- Naser, V.; Shani, E. Auxin response under osmotic stress. Plant Mol. Biol. 2016, 91, 661–672. [Google Scholar] [CrossRef]
- Wang, T.; Li, P.; Mu, T.; Dong, G.; Zheng, C.; Jin, S.; Chen, T.; Hou, B.; Li, Y. Overexpression of UGT74E2, an Arabidopsis IBA glycosyltransferase, enhances seed germination and modulates stress tolerance via ABA signaling in rice. Int. J. Mol. Sci. 2020, 21, 7239. [Google Scholar] [CrossRef]
- Tognetti, V.B.; Van Aken, O.; Morreel, K.; Vandenbroucke, K.; van de Cotte, B.; De Clercq, I.; Chiwocha, S.; Fenske, R.; Prinsen, E.; Boerjan, W.; et al. Perturbation of indole-3-butyric acid homeostasis by the UDP-glucosyltransferase UGT74E2 modulates Arabidopsis architecture and water stress tolerance. Plant Cell 2010, 22, 2660–2679. [Google Scholar] [CrossRef]
- Huang, X.; Hou, L.; Meng, J.; You, H.; Li, Z.; Gong, Z.; Yang, S.; Shi, Y. The antagonistic action of abscisic acid and cytokinin signaling mediates drought stress response in Arabidopsis. Mol. Plant 2018, 11, 970–982. [Google Scholar] [CrossRef] [PubMed]
- Nishiyama, R.; Watanabe, Y.; Fujita, Y.; Le, D.T.; Kojima, M.; Werner, T.; Vankova, R.; Yamaguchi-Shinozaki, K.; Shinozaki, K.; Kakimoto, T.; et al. Analysis of cytokinin mutants and regulation of cytokinin metabolic genes reveals important regulatory roles of cytokinins in drought, salt and abscisic acid responses, and abscisic acid biosynthesis. Plant Cell 2011, 23, 2169–2183. [Google Scholar] [CrossRef] [PubMed]
- Duarte, G.T.; Volkova, P.Y.; Geraskin, S.A. The response profile to chronic radiation exposure based on the transcriptome analysis of Scots pine from Chernobyl affected zone. Environ. Pollut. 2019, 250, 618–626. [Google Scholar] [CrossRef]
- Rizhsky, L.; Liang, H.; Shuman, J.; Shulaev, V.; Davletova, S.; Mittler, R. When defense pathways collide. The response of Arabidopsis to a combination of drought and heat stress. Plant Physiol. 2004, 134, 1683–1696. [Google Scholar] [CrossRef]
- Huang, Y.; Hussain, M.A.; Luo, D.; Xu, H.; Zeng, C.; Havlickova, L.; Bancroft, I.; Tian, Z.; Zhang, X.; Cheng, Y.; et al. A Brassica napus reductase gene dissected by associative transcriptomics enhances plant adaption to freezing stress. Front. Plant Sci. 2020, 11, 971. [Google Scholar] [CrossRef]
- Bourbousse, C.; Vegesna, N.; Law, K.A. SOG1 activator and MYB3R repressors regulate a complex DNA damage network in Arabidopsis. Proc. Natl. Acad. Sci. USA 2018, 115, E12453–E12462. [Google Scholar] [CrossRef] [PubMed]
- Volkova, P.Y.; Bondarenko, E.V.; Kazakova, E.A. Radiation hormesis in plants. Curr. Opin. Toxicol. 2022, 30, 100334. [Google Scholar] [CrossRef]
- Lee, S.; Lee, S.; Yang, K.; Kim, Y.-M.; Park, S.-Y.; Kim, S.Y.; Soh, M.-S. Overexpression of PRE1 and its homologous genes activates gibberellin-dependent responses in Arabidopsis thaliana. Plant Cell Physiol. 2006, 47, 591–600. [Google Scholar] [CrossRef] [PubMed]
- Leivar, P.; Tepperman, J.M.; Cohn, M.M.; Monte, E.; Al-Sady, B.; Erickson, E.; Quail, P.H. Dynamic antagonism between phytochromes and PIF family basic helix-loop-helix factors induces selective reciprocal responses to light and shade in a rapidly responsive transcriptional network in Arabidopsis. Plant Cell 2012, 24, 1398–1419. [Google Scholar] [CrossRef]
- Hieno, A.; Naznin, H.; Inaba-Hasegawa, K.; Yokogawa, T.; Hayami, N.; Nomoto, M.; Tada, Y.; Yokogawa, T.; Higuchi-Takeuchi, M.; Hanada, K.; et al. Transcriptome analysis and identification of a transcriptional regulatory network in the tesponse to H2O2. Plant Physiol. 2019, 180, 1629–1646. [Google Scholar] [CrossRef]
- Verbruggen, N.; Hermans, C. Proline accumulation in plants: A review. Amino Acids 2008, 35, 753–759. [Google Scholar] [CrossRef] [PubMed]
- Funck, D.; Eckard, S.; Muller, G. Non-redundant functions of two proline dehydrogenase isoforms in Arabidopsis. BMC Plant Biol. 2010, 10, 70. [Google Scholar] [CrossRef]
- Hayat, S.; Hayat, Q.; Alyemeni, M.N.; Wani, A.S.; Pichtel, J.; Ahmad, A. Role of proline under changing environments. Plant Signal. Behav. 2012, 7, 1456–1466. [Google Scholar] [CrossRef] [PubMed]
- Macovei, A.; Pagano, A.; Sabatini, M.E.; Grandi, S.; Balestrazzi, A. The human tyrosyl-DNA phosphodiesterase 1 (hTdp1) inhibitor NSC120686 as an exploratory tool to investigate plant Tdp1 genes. Genes 2018, 9, 186. [Google Scholar] [CrossRef] [PubMed]
- Hurny, A.; Cuesta, C.; Cavallari, N.; Ötvös, K.; Duclercq, J.; Dokládal, L.; Montesinos, J.C.; Gallemí, M.; Semerádová, H.; Rauter, T.; et al. SYNERGISTIC ON AUXIN AND CYTOKININ 1 positively regulates growth and attenuates soil pathogen resistance. Nat. Commun. 2020, 11, 2170. [Google Scholar] [CrossRef] [PubMed]
- Vercruysse, J.; Baekelandt, A.; Gonzalez, N.; Inze, D. Molecular networks regulating cell division during Arabidopsis leaf growth. J. Exp. Bot. 2020, 71, 2365–2378. [Google Scholar] [CrossRef] [PubMed]
- Stortenbeker, N.; Bemer, M. The SAUR gene family: The plant’s toolbox for adaptation of growth and development. J. Exp. Bot. 2019, 70, 17–27. [Google Scholar] [CrossRef]
- Babina, D.; Podobed, M.; Bondarenko, E.; Kazakova, E.; Bitarishvili, S.; Podlutskii, M.; Mitsenyk, A.; Prazyan, A.; Gorbatova, I.; Shesterikova, E.; et al. Seed gamma irradiation of Arabidopsis thaliana ABA-mutant lines alters germination and does not inhibit the photosynthetic efficiency of juvenile plants. Dose Response 2020, 18, 1559325820979249. [Google Scholar] [CrossRef]
- Ralph, P.; Gademann, R. Rapid Light Curves: A powerful tool to assess photosynthetic activity. Aquat. Bot. 2005, 82, 222–237. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Buschmann, C.; Knapp, M. How to correctly determine the different chlorophyll fluorescence parameters and the chlorophyll fluorescence decrease ratio RFd of leaves with the PAM fluorometer. Photosynthetica 2005, 43, 379–393. [Google Scholar] [CrossRef]
- Easlon, H.M.; Bloom, A.J. Easy Leaf Area: Automated digital image analysis for rapid and accurate measurement of leaf area. Plant Sci. 2014, 2, 1400033. [Google Scholar] [CrossRef] [PubMed]
- Bitarishvili, S.V.; Volkova, P.Y.; Geras’kin, S.A. γ-Irradiation of barley seeds and its effect on the phytohormonal status of seedlings. Russ. J. Plant Physiol. 2018, 65, 446–454. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Langmead, B.; Salzberg, S. HISAT: A fast-spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. EdgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2009, 26, 139–140. [Google Scholar] [CrossRef] [PubMed]
- Afgan, E.; Baker, D.; van den Beek, M.; Blankenberg, D.; Bouvier, D.; Čech, M.; Chilton, J.; Clements, D.; Coraor, N.; Eberhard, C.; et al. The Galaxy platform for accessible, reproducible and collaborative biomedical analyses: 2016 update. Nucleic Acids Res. 2016, 44, W3–W10. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experimental Plot (Natural Accession) | pH | HA | TEB | Humus | Av. P2O5 | Av. K2O | Av. Ca | Av. Mg | Av. Na |
|---|---|---|---|---|---|---|---|---|---|
| mg-eqv per 100 g | % | mg/kg | mg-eqv per 100 g | ||||||
| Babchin (Bab-0) | 6.61 | 6.4 | 6.1 ± 0.7 | 1.17 ± 0.01 | 210.7 ± 5.2 | 91.0 ± 1.0 | 3.41 ± 0.20 | 0.85 ± 0.02 | 0.61 ± 0.04 |
| Vygrebnaya Sloboda (VS-0) | 6.33 ± 0.02 | 4.3 | 1.1 ± 0.1 | 0.60 ± 0.02 | 47.2 ± 0.6 | 19.0 ± 0.1 | 1.06 ± 0.08 | 0.23 ± 0.01 | 0.71 ± 0.01 |
| Masany (Masa-0) | 5.50 ± 0.01 | 3.8 | 1.0 ± 0.4 | 1.59 ± 0.02 | 54.5 ± 0.4 | 16.9 ± 0.2 | 1.16 ± 0.07 | 0.35 ± 0.01 | 0.61 |
| Experimental Plot (Natural Accession) | Ambient Dose Rate (γ) | α-Particles Flux Density | β-Particles Flux Density | Activity of 137Cs in Soil | Activity of 90Sr in Soil |
|---|---|---|---|---|---|
| μSv × h−1 | min−1 × cm−2 | Bq × kg−1 | |||
| Babchin (Bab-0) | 0.33 | 1.2 | 1.2 | 126 ± 4.5 | 9.9 ± 1.6 |
| Vygrebnaya Sloboda (VS-0) | 0.60 | 17.0 | 5.2 | 1074 ± 31 | 16.3 ± 2.7 |
| Masany (Masa-0) | 3.76 | 25.3 | 21.3 | 11,510 ± 294 | 233.8 ± 35.4 |
| Chronically Irradiated Accession VS-0 | ||
| GO Terms | Up/Downregulated Genes | p-Value |
| Molecular Function | ||
| protein serine/threonine kinase activity (GO:0004674) | Down | 3.9 × 10−6 |
| ADP binding (GO:0043531) | Down | 3.9 × 10−6 |
| ATP binding (GO:0005524) | Down | 3.8 × 10−4 |
| calcium ion binding (GO:0005509) | Down | 3.9 × 10−2 |
| Biological Process | ||
| protein phosphorylation (GO:0006468) | Down | 1.2 × 10−6 |
| defence response to bacterium (GO:0042742) | Down | 5.6 × 10−3 |
| systemic acquired resistance (GO:0009627) | Down | 3.0 × 10−3 |
| signal transduction (GO:0007165) | Down | 2.6 × 10−3 |
| response to ozone (GO:0010193) | Down | 2.6 × 10−3 |
| response to chitin (GO:0010200) | Down | 1.0 × 10−2 |
| positive regulation of innate immune response (GO:0045089) | Down | 1.0 × 10−2 |
| response to UV-B (GO:0010224) | Down | 1.7 × 10−2 |
| Chronically Irradiated Accession Masa-0 | ||
| GO Terms | Up/Downregulated Genes | p-Value |
| Molecular Function | ||
| ADP binding (GO:0043531) | Up | 2.3 × 10−4 |
| protein serine/threonine kinase activity (GO:0004674) | Down | 4.8 × 10−2 |
| carbohydrate binding (GO:0030246) | Down | 4.8 × 10−2 |
| Acutely Irradiated Plants of the Accession Bab-0 | ||
| GO Terms | Up/Downregulated Genes | p-Value |
| Molecular Function | ||
| glutathione transferase activity (GO:000258) | Up | 2.6 × 10−4 |
| kinase activity (GO:0016301) | Up | 3.5 × 10−4 |
| xyloglucan: xyloglucosyl transferase activity (GO:0016762) | Up | 2.3 × 10−3 |
| oxidoreductase activity, act. on a sulphur group of donors (GO:0016667) | Up | 4.3 × 10−2 |
| hydrolase activity, hydrolysing O-glycosyl compounds (GO:0046527) | Up/Down | 2.4 × 10−2/2.9 × 10−2 |
| peroxidase activity (GO:0004601) | Down | 9.9 × 10−5 |
| heme biding (GO:0020037) | Down | 2.1 × 10−4 |
| copper ion binding (GO:0005507) | Down | 2.8 × 10−3 |
| oxidoreductase activity, oxidizing metal ions (GO:0016722) | Down | 2.4 × 10−3 |
| Biological Process | ||
| response to toxic substance (GO:0009636) | Up | 9.5 × 10−6 |
| response to salicylic acid (GO:0009751) | Up | 8.3 × 10−5 |
| toxic catabolic process (GO:0009407) | Up | 4.4 × 10−5 |
| defence response to bacterium (GO:0042742) | Up | 4.0 × 10−5 |
| response to wounding (GO:0009611) | Up | 2.5 × 10−5 |
| response to jasmonic acid (GO:0009753) | Up | 1.5 × 10−4 |
| xyloglucan metabolic process (GO:0010411) | Up | 8.8 × 10−3 |
| response to virus (GO:0009615) | Up | 6.3 × 10−3 |
| response to abscisic acid (GO:0009737) | Up | 6.0 × 10−3 |
| defence response to fungus (GO:0050832) | Up | 1.6 × 10−3 |
| oxidation-reduction process (GO:0055114) | Up/Down | 2.7 × 10−2/1.9 × 10−6 |
| leaf senescence (GO:0010150) | Up | 2.4 × 10−2 |
| response to salt stress (GO:0009651) | Up | 1.4 × 10−2 |
| response to water deprivation (GO:0009414) | Up | 1.7 × 10−2 |
| suberin biosynthetic process (GO:0010345) | Down | 1.4 × 10−6 |
| hydrogen peroxide catabolic process (GO:0042744) | Down | 4.5 × 10−5 |
| response to oxidative stress (GO:0006979) | Down | 5.7 × 10−4 |
| root morphogenesis (GO:0010015) | Down | 2.7 × 10−4 |
| lignin metabolic process (GO:0009808) | Down | 5.2 × 10−3 |
| plant-type cell wall organization (GO:0009664) | Down | 4.1 × 10−3 |
| Cellular Component | ||
| plasma membrane (GO:0005886) | Up | 8.0 × 10−4 |
| apoplast (GO:0048046) | Up | 2.7 × 10−3 |
| integral component of membrane (GO:0016021) | Up | 1.3 × 10−3 |
| cell wall (GO:0005618) | Up | 1.1 × 10−2 |
| extracellular region (GO:0005576) | Down | 5.2 × 10−5 |
| Acutely Irradiated Plants of the Accession VS-0 | ||
| GO Terms | Up/Downregulated Genes | p-Value |
| Molecular Function | ||
| xyloglucan: xyloglucosyl transferase activity (GO:0016762) | Up | 2.2 × 10−3 |
| peroxidase activity (GO:0004601) | Down | 3.6 × 10−8 |
| heme binding (GO:0020037) | Down | 3.8 × 10−6 |
| quercetin 3-O-glucosyltransferase activity (GO:0080043) | Down | 3.8 × 10−2 |
| quercetin 7-O-glucosyltransferase activity (GO:0080043) | Down | 3.8 × 10−2 |
| symporter activity (GO:0015293) | Down | 2.7 × 10−2 |
| Biological Process | ||
| response to jasmonic acid (GO:0009753) | Up | 3.9 × 10−6 |
| defence response to fungus (GO:0050832) | Up | 3.3 × 10−6 |
| response to salicylic acid (GO:0009751) | Up | 1.7 × 10−6 |
| defence response to bacterium (GO:0042742) | Up | 1.4 × 10−5 |
| response to reactive oxygen species (GO:0000302) | Up | 6.3 × 10−4 |
| response to salt stress (GO:0009651) | Up | 5.2 × 10−3 |
| xyloglucan metabolic process (GO:0010411) | Up/Down | 4.8 × 10−3/3.0 × 10−2 |
| cellular calcium ion homeostasis (GO:0006874) | Up | 3.0 × 10−3 |
| lignin metabolic process (GO:0009808) | Up/Down | 3.8 × 10−2/1.1 × 10−2 |
| response to abscisic acid (GO:0009737) | Up | 1.3 × 10−2 |
| toxin catabolic process (GO:0009407) | Up | 1.2 × 10−2 |
| hydrogen peroxide metabolic process (GO:0042744) | Down | 7.6 × 10−7 |
| oxidation-reduction process (GO:0055114) | Down | 5.8 × 10−5 |
| response to hypoxia (GO:0001666) | Down | 1.9 × 10−4 |
| response to oxidative stress (GO:000513) | Down | 5.1 × 10−3 |
| plant type cell wall organization (GO:0009664) | Down | 2.2 × 10−3 |
| root morphogenesis (GO:0010015) | Down | 3.0 × 10−2 |
| phenylpropanoid biosynthetic process (GO:0009699) | Down | 3.0 × 10−2 |
| cellular response to starvation (GO:0009267) | Down | 2.6 × 10−2 |
| Cellular Component | ||
| cell wall (GO:0005618) | Up/Down | 3.5 × 10−6/7.6 × 10−6 |
| plasma membrane (GO:0005886) | Up | 4.5 × 10−3 |
| extracellular space (GO:0005615) | Down | 3.3 × 10−2 |
| Acutely Irradiated Plants of the Accession Masa-0 | ||
| GO Terms | Up/Downregulated Genes | p-Value |
| Biological Process | ||
| innate immune response (GO:0045087) | Up | 2.7 × 10−3 |
| regulation of growth (GO:0040008) | Up | 1.7 × 10−3 |
| hormone-mediated signaling pathway (GO:0009755) | Up | 1.2 × 10−2 |
| defence response to another organism (GO:0098542) | Up | 1.4 × 10−2 |
| Cellular Component | ||
| cell wall | Up | 3.5 × 10−2 |
| Gene | Description | Acute Irradiation 1 | Chronic Irradiation 2 |
|---|---|---|---|
| log2FC | |||
| AT4G05235 | Long non-coding RNA | 10.77 | −8.45 |
| AT1G15640 | Transmembrane protein | 7.93 | −8.59 |
| AT5G14160 | F-box family protein | 7.28 | −7.58 |
| AT3G21330 | Basic helix–loop–helix (bHLH) DNA-binding superfamily protein | 4.00 | |
| AT5G18010 | SAUR-like auxin-responsive protein family | 3.10 | |
| AT5G39860 | Basic helix–loop–helix (bHLH) DNA-binding family protein | 2.74 | |
| AT5G18050 | SAUR-like auxin-responsive protein family | 2.63 | |
| AT1G52400 | Beta glucosidase 18 | 2.47 | |
| AT5G18080 | SAUR-like auxin-responsive protein family | 2.39 | |
| AT5G18030 | SAUR-like auxin-responsive protein family | 2.39 | |
| AT3G09960 | Calcineurin-like metallo-phosphoesterase superfamily protein | 2.31 | −2.80 |
| AT3G57520 | Seed imbibition 2 | −2.14 | |
| AT3G30775 | Proline oxidase | −2.17 | |
| AT5G41080 | PLC-like phosphodiesterases superfamily protein | −2.45 | |
| AT1G02820 | Late embryogenesis abundant 3 (LEA3) family protein | −2.61 | |
| AT4G04223 | Other RNA | −2.90 | 4.90 |
| AT3G60950 | C2 calcium/lipid-binding endonuclease/exonuclease/phosphatase | −4.80 | −2.67 |
| AT3G62460 | Putative endonuclease or glycosyl hydrolase | −4.82 | 4.68 |
| AT3G61030 | Calcium-dependent lipid-binding (CaLB domain) family protein | −4.93 | −2.88 |
| AT2G29000 | Leucine-rich repeat protein kinase family protein | −8.74 | 8.74 |
| AT2G27402 | Hypothetical protein | −8.97 | 12.56 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Podlutskii, M.; Babina, D.; Podobed, M.; Bondarenko, E.; Bitarishvili, S.; Blinova, Y.; Shesterikova, E.; Prazyan, A.; Turchin, L.; Garbaruk, D.; et al. Arabidopsis thaliana Accessions from the Chernobyl Exclusion Zone Show Decreased Sensitivity to Additional Acute Irradiation. Plants 2022, 11, 3142. https://doi.org/10.3390/plants11223142
Podlutskii M, Babina D, Podobed M, Bondarenko E, Bitarishvili S, Blinova Y, Shesterikova E, Prazyan A, Turchin L, Garbaruk D, et al. Arabidopsis thaliana Accessions from the Chernobyl Exclusion Zone Show Decreased Sensitivity to Additional Acute Irradiation. Plants. 2022; 11(22):3142. https://doi.org/10.3390/plants11223142
Chicago/Turabian StylePodlutskii, Mikhail, Darya Babina, Marina Podobed, Ekaterina Bondarenko, Sofia Bitarishvili, Yana Blinova, Ekaterina Shesterikova, Alexander Prazyan, Larisa Turchin, Dmitrii Garbaruk, and et al. 2022. "Arabidopsis thaliana Accessions from the Chernobyl Exclusion Zone Show Decreased Sensitivity to Additional Acute Irradiation" Plants 11, no. 22: 3142. https://doi.org/10.3390/plants11223142