
Susceptibility of Novel Promising Citrus Rootstocks to White Root Rot
 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
2.1. Identification of Fungal Isolate
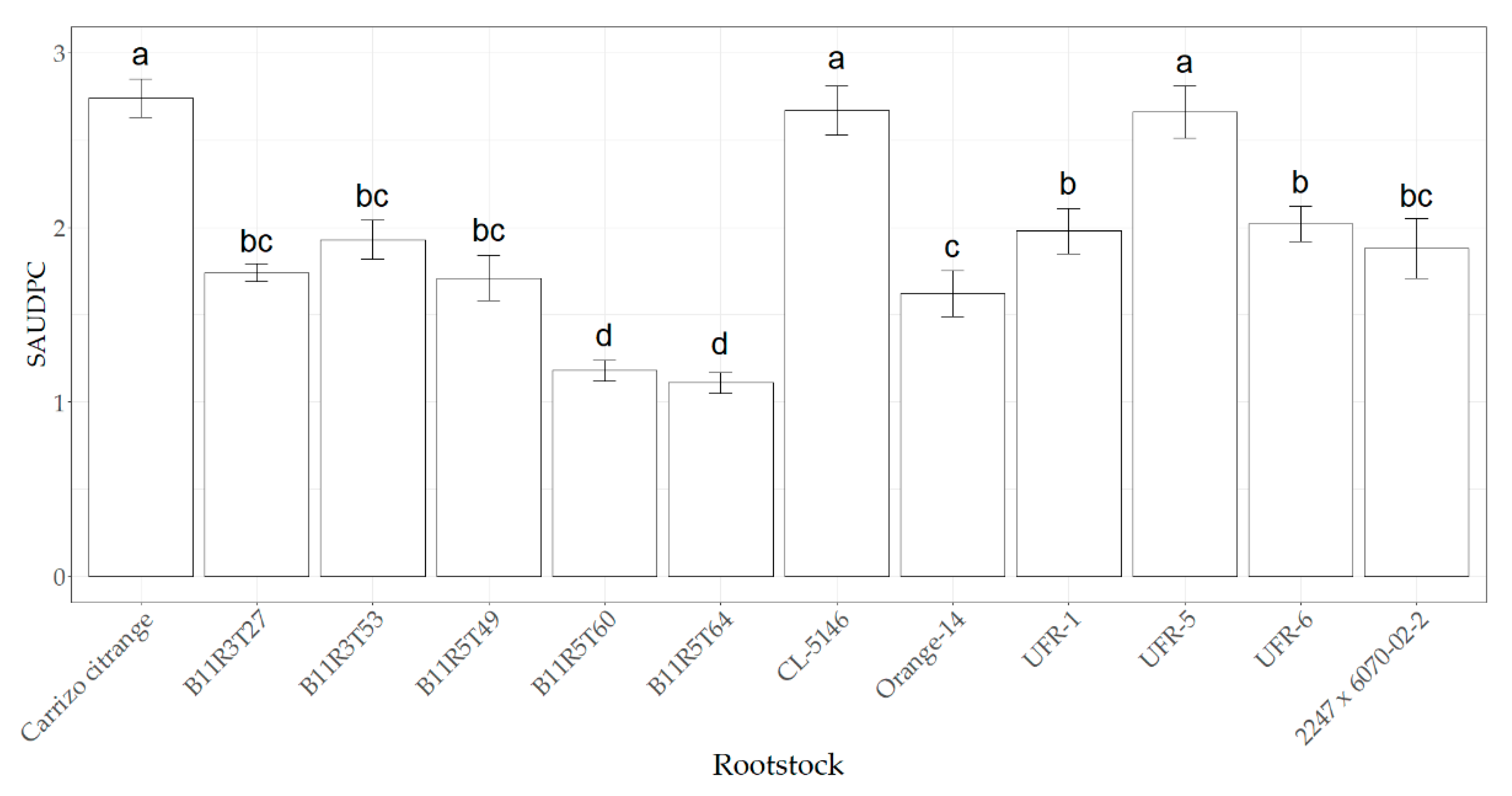
2.2. Plant Symptoms and Responses of Different Citrus Rootstocks
2.3. Content of Chlorophyll in the Citrus Rootstocks Assayed
2.4. Leaf Area Response in Different Citrus Rootstocks
2.5. Effect of R. necatrix on Biomass Production
2.5.1. Fresh Weight
2.5.2. Dry Weight
3. Discussion
4. Materials and Methods
4.1. Plant Material and Experimental Design

{kind=link}
{kind=link}
| Rootstock | Parentage | Origin | Ref. |
|---|---|---|---|
| Diploids | |||
| Carrizo citrange | Citrus sinensis ‘Washington’ × Poncirus trifoliata | USDA | [69] |
| B11R3T27 | P. trifoliata ‘Flying Dragon’ × C. paradisi ‘Duncan’ | CREC | [77] |
| B11R3T53 | (C. reticulata ‘Cleopatra’ × C. ichangensis) × (C. maxima × P. trifoliata) | CREC | [66] |
| B11R5T49 | P. trifoliata ‘Flying Dragon’ × C. sinensis ‘Ridge pineapple’ | CREC | [66] |
| B11R5T60 | P. trifoliata ‘Flying Dragon’ × C. sinensis ‘Ridge pineapple’ | CREC | [67] |
| B11R5T64 | P. trifoliata ‘Flying Dragon’ × C. sinensis ‘Ridge pineapple’ | CREC | [66] |
| CL-5146 | C. sunki × Citroncirus spp. ‘Swingle’ | CL | [66] |
| Tetraploids | |||
| Orange-14 * | C. reticulata ‘Nova’ + C. maxima HBP × C. reticulata ‘Cleopatra’ + P. trifoliata | CREC | [81] |
| UFR-1 (Orange-3) * | C. reticulata ‘Nova’ + C. maxima HBP × C. reticulata ‘Cleopatra’ + P. trifoliata | CREC | [75,82,83] |
| UFR-5 (White 4) * | C. reticulata ‘Nova’ + C. maxima HBP × C. sinensis ‘Succari’ + P. trifoliata | CREC | [75,84] |
| UFR-6 ** | C. reticulata ‘Changsha’ + P. trifoliata ‘50-7’ | CREC | [75,85] |
| 2247 × 6070-02-2 * | C. reticulata ‘Nova’ + C. maxima HBP × C. aurantium+Poncirus trifoliata ‘Flying Dragon’ | CREC | [67] |
4.2. Fungal Isolate
4.3. Plant Inoculation
4.4. Plant Symptoms Evaluation
4.5. Evaluation of Leaf Chlorophyll Content
4.6. Leaf Area Assessment
4.7. Biomass
4.8. Fungal Isolation from Inoculated Plants
4.9. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- AOSTAT. Food and Agriculture Organization (FAO) of the United Nations. 2022. Available online: http://www.fao.org/faostat/es/#home (accessed on 6 August 2022).
- ten Hoopen, G.M.; Krauss, U. Biology and control of Rosellinia bunodes, Rosellinia necatrix and Rosellinia pepo:A review. Crop Prot. 2006, 25, 89–107. [Google Scholar] [CrossRef]
- Fungal Databases, U.S. National Fungus Collection, United States Department of Agriculture. 2022. Available online: https://nt.ars-grin.gov/fungaldatabases/ (accessed on 25 August 2022).
- Sivanesan, A.; Holliday, P. Rosellinia Necatrix. CMI Descriptions of Pathogenic Fungi and Bacteria; Commonwealth Mycological Institute: Kew, UK, 1972; ISBN 0009-9716. [Google Scholar]
- Sztejnberg, A.; Madar, Z. Host range of Dematophora necatrix, the cause of white root rot disease in fruit trees. Plant Dis. 1980, 64, 662–664. [Google Scholar] [CrossRef]
- Arakawa, M.; Nakamura, H.; Uetake, Y.; Matsumoto, N. Presence and distribution of double-stranded RNA elements in the white root rot fungus Rosellinia necatrix. Mycoscience 2002, 43, 21–26. [Google Scholar] [CrossRef]
- Melgarejo Nárdiz, P.; García-Jiménez, J.; Jordá Gutiérrez, M.C.; López González, M.M.; Andrés Yebes, M.F.; Duran-Vila, N. Patógenos de Plantas Descritos en España, 2nd ed.; Ministerio de Medio Ambiente y Medio Rural y Marino: Madrid, Spain, 2010. [Google Scholar]
- González-Domínguez, E.; Pérez-Sierra, A.; Álvarez, L.A.; León, M.; Abad-Campos, P.; Armengol, J.; García-Jiménez, J. Agentes fúngicos presentes en plantaciones de nísperos (Eriobotrya japonica Lindl.) con síntomas de decaimiento en la provincia de Alicante. Boletín Sanid. Veg. -Plagas 2009, 35, 453–467. [Google Scholar]
- López-Herrera, C.J. Podredumbres radiculares del aguacate en la Costa del Sol. Años 1987-88. In Estudios de Fitopatología; Moral, J., Ed.; SEFDGIEA: Badajoz, Spain, 1989; pp. 172–176. [Google Scholar]
- Arjona-Girona, I.; López-Herrera, C.J. First report of Rosellinia necatrix causing white root rot in mango trees in Spain. Plant Dis. 2018, 102, 2639. [Google Scholar] [CrossRef]
- Armengol, J.; Vicent, A.; León, M.; Berbegal, M.; Abad-Campos, P.; García-Jiménez, J. Analysis of population structure of Rosellinia necatrix on Cyperus esculentus by mycelial compatibility and inter-simple sequence repeats (ISSR). Plant Pathol. 2010, 59, 179–185. [Google Scholar] [CrossRef]
- Guillaumin, J.J. Le pourridié. Phytoma 1986, 19, 20–23. [Google Scholar]
- Martínez-Ferri, E.; Moreno-Ortega, G.; van den Berg, N.; Pliego, C. Mild water stress-induced priming enhance tolerance to Rosellinia necatrix in susceptible avocado rootstocks. BMC Plant Biol. 2019, 19, 458. [Google Scholar] [CrossRef]
- López-Herrera, C.J.; Pérez-Jiménez, R.M.; Zea-Bonilla, T.; Basallote-Ureba, M.J.; Melero-Vara, J.M. Soil solarization in established avocado trees for control of Dematophora necatrix. Plant Dis. 1998, 82, 1088–1092. [Google Scholar] [CrossRef] [Green Version]
- López-Herrera, C.J.; Pérez-Jiménez, R.M.; Basallote-Ureba, M.J.; Zea-Bonilla, T.; Melero-Vara, J.M. Loss of viability of Dematophora necatrix in solarized soils. Eur. J. Plant Pathol. 1999, 105, 571–576. [Google Scholar] [CrossRef]
- Sztejnberg, A.; Freeman, S.; Chet, I.; Katan, J. Control of Rosellinia necatrix in soil and in apple orchard by solarization and Trichoderma harzianum. Plant Dis. 1987, 71, 365–369. [Google Scholar] [CrossRef]
- Ruano-Rosa, D.; López-Herrera, C.J. Evaluation of Trichoderma spp. as biocontrol agents against avocado white root rot. Biol. Control 2009, 51, 66–71. [Google Scholar] [CrossRef]
- Ruano-Rosa, D.; Del Moral-Navarrete, L.; Lopez-Herrera, C.J. Selection of Trichoderma spp. isolates antagonistic to Rosellinia necatrix. Span. J. Agric. Res. 2010, 8, 1084–1097. [Google Scholar] [CrossRef]
- Arjona-Girona, I.; López-Herrera, C.J. Study of a new biocontrol fungal agent for avocado white root rot. Biol. Control 2018, 117, 6–12. [Google Scholar] [CrossRef]
- Arjona-López, J.M.; López-Herrera, C.J. Entoleuca sp. infected by mycoviruses as potential biocontrol agents of avocado white root rot. Eur. J. Plant Pathol. 2021, 159, 409–420. [Google Scholar] [CrossRef]
- Ruano-Rosa, D.; Lopez-Herrera, C.J. Biocontrol de la podredumbre blanca del aguacate con aislados no-patogénicos de Rosellinia necatrix. In Proceedings of the XIII Congreso de la Sociedad Española de Fitopatología, Murcia, Spain, 18–22 September 2006; Montesinos, E., Ed.; p. 377. [Google Scholar]
- Pal, J.; Sharma, S.K.; Devi, S.; Sharma, R.; Raj, H.; Karn, M.; Verma, S.; Vedukola, P.R.; Sharma, A. Screening, identification, and colonization of fungal root endophytes against Dematophora necatrix: A ubiquitous pathogen of fruit trees. Egypt. J. Biol. Pest Control 2020, 30, 112. [Google Scholar] [CrossRef]
- Cazorla, F.M.; Duckett, S.B.; Bergström, E.T.; Noreen, S.; Odijk, R.; Lugtenberg, B.J.J.; Thomas-Oates, J.E.; Bloemberg, G.V. Biocontrol of avocado Dematophora root rot by antagonistic Pseudomonas fluorescens PCL1606 correlates with the production of 2-hexyl 5-propyl resorcinol. Mol. Plant-Microbe Interact. 2006, 19, 418–428. [Google Scholar] [CrossRef] [Green Version]
- Pliego, C.; Ramos, C.; de Vicente, A.; Cazorla, F.M. Screening for candidate bacterial biocontrol agents against soilborne fungal plant pathogens. Plant Soil 2011, 340, 505–520. [Google Scholar] [CrossRef] [Green Version]
- Tienda, S.; Vida, C.; Lagendijk, E.; de Weert, S.; Linares, I.; González-Fernández, J.; Guirado, E.; de Vicente, A.; Cazorla, F.M. Soil application of a formulated biocontrol rhizobacterium, Pseudomonas chlororaphis PCL1606, induces soil suppressiveness by impacting specific microbial communities. Front. Microbiol. 2020, 11, 1874. [Google Scholar] [CrossRef]
- Kanematsu, S.; Arakawa, M.; Oikawa, Y.; Onoue, M.; Osaki, H.; Nakamura, H.; Ikeda, K.; Kuga-Uetake, Y.; Nitta, H.; Sasaki, A.; et al. A Reovirus causes hypovirulence of Rosellinia necatrix. Phytopathology 2004, 94, 561–568. [Google Scholar] [CrossRef] [Green Version]
- Chiba, S.; Salaipeth, L.; Lin, Y.-H.; Sasaki, A.; Kanematsu, S.; Suzuki, N. A novel bipartite double-stranded RNA mycovirus from the white root rot fungus Rosellinia necatrix: Molecular and biological characterization, taxonomic considerations, and potential for biological control. J. Virol. 2009, 83, 12801–12812. [Google Scholar] [CrossRef] [Green Version]
- Arjona-López, J.M.; Telengech, P.; Suzuki, N.; López-Herrera, C.J. A moderate level of hypovirulence conferred by a hypovirus in the avocado white root rot fungus, Rosellinia necatrix. Fungal Biol. 2021, 125, 69–76. [Google Scholar] [CrossRef]
- Ruano-Rosa, D.; Cazorla, F.M.; Bonilla, N.; Martín-Pérez, R.; De Vicente, A.; López-Herrera, C.J. Biological control of avocado white root rot with combined applications of Trichoderma spp. and rhizobacteria. Eur. J. Plant Pathol. 2014, 138, 751–762. [Google Scholar] [CrossRef]
- Arjona-López, J.M.; Tienda, S.; Arjona-Girona, I.; Cazorla, F.M.; López-Herrera, C.J. Combination of low concentrations of fluazinam and antagonistic rhizobacteria to control avocado white root rot. Biol. Control 2019, 136, 103996. [Google Scholar] [CrossRef]
- Ruano-Rosa, D.; Arjona-Girona, I.; López-Herrera, C.J. Integrated control of avocado white root rot combining low concentrations of fluazinam and Trichoderma spp. Crop Prot. 2017, 112, 363–370. [Google Scholar] [CrossRef]
- Arjona-López, J.M.; López-Herrera, C.J. Control of avocado white root rot using non-pathogenic Rosellinia necatrix isolates combined with low concentration of fluazinam. BioControl 2020, 65, 247–255. [Google Scholar] [CrossRef]
- Behdad, E. The influence of several new systemic fungicides on Rosellinia necatrix (Hart.) Berl. Iran. J. Plant Pathol. 1976, 12, 40–41. [Google Scholar]
- Kanadani, G.; Date, H.; Nasu, H. Effect of fluazinam soil-drench on white root rot of grapevine. Jpn. J. Phytopathol. 1998, 64, 139–141. [Google Scholar] [CrossRef] [Green Version]
- Nitta, H.; Hatamoto, M.; Kurihisa, H. Control of white root rot on Japanese pear using dazomet micro-granules. Bull. Hiroshima Pref. Agr. Res. Cent. 2002, 72, 25–34. [Google Scholar]
- López-Herrera, C.J.; Zea-Bonilla, T. Effects of benomyl, carbendazim, fluazinam and thiophanate methyl on white root rot of avocado. Crop Prot. 2007, 26, 1186–1192. [Google Scholar] [CrossRef]
- Arjona-López, J.M.; Capote, N.; Melero-Vara, J.M.; López-Herrera, C.J. Control of avocado white root rot by chemical treatments with fluazinam in avocado orchards. Crop Prot. 2020, 131, 105100. [Google Scholar] [CrossRef]
- Carvalho, F.P. Pesticides, environment, and food safety. Food Energy Secur. 2017, 6, 48–60. [Google Scholar] [CrossRef]
- Bakırcı, G.T.; Acay, D.B.Y.; Bakırcı, F.; Ötleş, S. Pesticide residues in fruits and vegetables from the Aegean region, Turkey. Food Chem. 2014, 160, 379–392. [Google Scholar] [CrossRef]
- Gullino, M.L.; Kuijpers, L.A.M. Social and political implications of managing plant diseases with restricted fungicides in Europe. Annu. Rev. Phytopathol. 1994, 32, 559–581. [Google Scholar] [CrossRef]
- EU Law—EUR-Lex. 2022. Access to European Union Law. Available online: https://eur-lex.europa.eu/homepage.html (accessed on 25 August 2022).
- EU Pesticide Database. 2022. Available online: https://food.ec.europa.eu/plants/pesticides/eu-pesticides-database_en (accessed on 26 August 2022).
- Registro de Productos Fitosanitarios, Ministerio de Agricultura, Pesca y Alimentación. 2022. Available online: https://www.mapa.gob.es/es/agricultura/temas/sanidad-vegetal/productos-fitosanitarios/registro-productos/ (accessed on 26 August 2022).
- Farm to Fork Strategy, European Commission. 2022. Available online: https://food.ec.europa.eu/horizontal-topics/farm-fork-strategy_en (accessed on 20 September 2022).
- Graham, J.; Feichtenberger, E. Citrus Phytophthora diseases: Management challenges and successes. J. Citrus Pathol. 2015, 2, 1–11. [Google Scholar]
- Lee, S.B.; Ko, K.; Aldwinckle, H.S. Resistance of selected Malus germplasm to Rosellinia necatrix. J. Am. Pomol. Soc. 2000, 54, 219–228. [Google Scholar]
- Choi, B.-H.; Kim, C.-S.; Jeong, Y.-J.; Park, I.-H.; Han, S.-G.; Yoon, T.-M. Resistance evaluatoon of G, CG, or M series apple rootstocks to soil-borne diseases (Phytophthora root rot, white root rot, and southern blight) and woolly apple aphid. Hortic. Sci. Technol. 2021, 39, 167–174. [Google Scholar] [CrossRef]
- Barceló-Muñoz, A.; Zea-Bonilla, T.; Jurado-Valle, I.; Imbroada-Solano, I.; Vidoy-Mercado, I.; Pliego-Alfaro, F.; López-Herrera, C.J. Programa de selección de portainjertos de aguacate tolerantes a la podredumbre blanca causada por Rosellinia necatrix en el sur de España (1995–2007). In Proceedings of the VI World Avocado Congress (Actas VI Congreso Mundial del Aguacate), Viña Del Mar, Chile, 12–16 November 2007; pp. 1–8. [Google Scholar]
- Pérez Jiménez, R.M.; Zea Bonilla, T.; Imbroda Solano, I.; Pliego-Alfaro, F.; López Herrera, C.J.; Barceló-Muñoz, A. Selección de portainjertos de aguacate tolerantes a la podredumbre blanca causada por Rosellinia necatrix. In Proceedings of the V World Avocado Congress (Actas V Congreso Mundial del Aguacate), Granada-Málaga, Spain, 19–24 October 2003; pp. 537–541. [Google Scholar]
- Mansoori, B.; Dorostkar, M. Reactions of some grape cultivars to Dematophora necatrix. Vitis 2008, 47, 231–233. [Google Scholar]
- Sztejnberg, A.; Jabareen, H. Dematophora root rot disease in persimmon and studies on resistance of rootstocks to the disease. Alon Hanotea 1985, 39, 757–762. [Google Scholar]
- Sztejnberg, A.; Jabareen, H. Studies of resistance of persimmon rootstocks to Dematophora root rot. Phytoparasitica 1986, 14, 240. [Google Scholar]
- Tallón Vila, C.I. Biotechnology applied to the genetic improvement of citrus rootstocks. In Development of a Protocol for Micropropagation and Adventitious Regeneration for Use in Generating Salt Toleran Mutant Lines; Universidad de Murcia: Murcia, Spain, 2015. [Google Scholar]
- Bové, J.M. Huanglongbing: A destructive, newly-emerging, century-old disease of citrus. J. Plant Pathol. 2006, 88, 7–37. [Google Scholar]
- Jagoueix, S.; Bove, J.M.; Garnier, M. The phloem-limited bacterium of greening disease of citrus is a member of the α subdivision of the Proteobacteria. Int. J. Syst. Evol. Microbiol. 1994, 44, 379–386. [Google Scholar] [CrossRef] [Green Version]
- Texeira, D.C.; Ayres, J.; Kitajima, E.W.; Danet, L.; Jagoueix-Eveillard, S.; Saillard, C.; Bové, J.M. First report of a Huanglongbing-like disease of citrus in São Paulo State, Brazil and association of a new Liberibacter Species, “Candidatus Liberibacter americanus”, with the disease. Plant Dis. 2005, 89, 107. [Google Scholar] [CrossRef]
- McClean, A.P.D.; Oberholzer, P.C.J. Citrus psylla, a vector of the greening disease of sweet orange. South Afr. J. Agric. Sci. 1965, 8, 297–298. [Google Scholar]
- Capoor, S.P.; Rao, D.G.; Viswanath, S.M. Diaphorina citri Kuway., a vector of the greening disease of citrus in India. Indian J. Agric. Sci. 1967, 37, 572–579. [Google Scholar]
- Yamamoto, P.T.; Felippe, M.R.; Garbim, L.F.; Coelho, J.H.C.; Ximenes, N.L.; Martins, E.C.; Leite, A.P.R.; Sousa, M.C.; Abrahão, D.P.; Braz, J.D. Diaphorina citri (Kuwayama) (Hemiptera: Psyllidae): Vector of the bacterium Candidatus Liberibacter americanus. In Proceedings of the Huanglongbing-Greening International Workshop, Ribeirão Prêto, Brazil, 16–20 July 2006; Pietersen, G., Le Roux, H.F., Eds.; Citrus Research International: Ribeiro Preto, Brazil, 2006; p. 96. [Google Scholar]
- Ajene, I.J.; Khamis, F.; Mohammed, S.; Rasowo, B.; Ombura, F.L.; Pietersen, G.; van Asch, B.; Ekesi, S. First report of field population of Trioza erytreae carrying the Huanglongbing-associated pathogen, Candidatus Liberibacter asiaticus, in Ethiopia. Plant Dis. 2019, 103, 1766. [Google Scholar] [CrossRef]
- Pérez-Otero, R.; Mansilla, J.P.; Del Estal, P. Detección de la psila africana de los cítricos, Trioza erytreae (Del Guercio, 1918) (Hemiptera: Psylloidea: Triozidae), en la Península Ibérica. Arq. Entomolóxicos 2015, 13, 119–122. [Google Scholar]
- Arenas-Arenas, F.J.; Duran-Vila, N.; Quinto, J.; Hervalejo, Á. Is the presence of Trioza erytreae, vector of huanglongbing disease, endangering the Mediterranean citrus industry? Survey of its population density and geographical spread over the last years. J. Plant Pathol. 2018, 100, 567–574. [Google Scholar] [CrossRef]
- Arenas-Arenas, F.J.; Duran-Vila, N.; Quinto, J.; Hervalejo, Á. Geographic spread and inter-annual evolution of populations of Trioza erytreae in the Iberian Peninsula. J. Plant Pathol. 2019, 101, 1151–1157. [Google Scholar] [CrossRef]
- DGAV. Direção-Geral de Alimentação e Veterinária. 2022. Available online: https://www.dgav.pt/ (accessed on 29 August 2022).
- EPPO. European and Mediterranean Plant Protection Organization. 2022. Available online: https://gd.eppo.int/ (accessed on 5 October 2022).
- Aparicio-Durán, L.; Arjona-López, J.M.; Hervalejo, A.; Calero-Velázquez, R.; Arenas-Arenas, F.J. Preliminary findings of new Citrus rootstocks potentially tolerant to foot rot caused by Phytophthora. Horticulturae 2021, 7, 389. [Google Scholar] [CrossRef]
- Aparicio-Durán, L.; Gmitter, F.G., Jr.; Arjona-López, J.M.; Calero-Velázquez, R.; Hervalejo, Á.; Arenas-Arenas, F.J. Water-stress influences on three new promising HLB-tolerant Citrus rootstocks. Horticulturae 2021, 7, 336. [Google Scholar] [CrossRef]
- Graham, J.H.; Timmer, L.W. Phytophthora diseases of citrus. In Plant Diseases of International Importance. Diseases of Fruit Crops; Kumar, J., Chaube, H.S., Singh, U.S., Mukhopadhyay, A.N., Eds.; Prentice Hall: Hoboken, NJ, USA, 1992; Volume 3, pp. 250–269. [Google Scholar]
- Savage, E.M.; Gardner, F.E. The Troyer and Carrizo citranges. Calif. Citrogr. 1965, 50, 112–116. [Google Scholar]
- McCarty, C.D.; Bitter, W.P.; Cole, D.A. Comparisons between Troyer and Carrizo citrange. Citrograph 1974, 59, 294–310. [Google Scholar]
- Bowman, K.D.; Joubert, J. Citrus rootstocks. In The Genus Citrus; Talon, M., Caruso, M., Gmitter, F.G., Eds.; Elsevier Inc.: Amsterdam, The Netherlands, 2020; pp. 105–127. ISBN 9780128121634. [Google Scholar]
- Citrus Variety Collection. University of California Riverside. 2022. Available online: https://citrusvariety.ucr.edu/ (accessed on 30 August 2022).
- Aleza, P.; Forner-Giner, M.A.; Del-Pino, Á. El panorama varietal y los nuevos patrones. Análisis de la situación actual. In Una Hoja de Ruta Para la Citricultura Española; García Álvarez-Coque, J.M., Moltó García, E., Eds.; Cajamar Caja Rural: Almería, Spain, 2020; pp. 151–166. ISBN 978-84-95531-49-0. [Google Scholar]
- Arjona-Girona, I.; Ariza-Fernández, T.; López-Herrera, C.J. Contribution of Rosellinia necatrix toxins to avocado white root rot. Eur. J. Plant Pathol. 2017, 148, 109–117. [Google Scholar] [CrossRef]
- Kunwar, S.; Grosser, J.; Gmitter, F.G.; Castle, W.S.; Albrecht, U. Field performance of ‘Hamlin’ orange trees grown on various rootstocks in Huanglongbing-endemic conditions. HortScience 2021, 56, 244–253. [Google Scholar] [CrossRef]
- Florida Citrus Rootstock Selection Guide, 4th Edition. 2022. Available online: https://crec.ifas.ufl.edu/extension/citrus_rootstock/tables.html (accessed on 7 October 2022).
- Project 18-029C. Evaluation of Citrus Rootstocks Response to HLB in Large-Scale Existing Field Trials. 2022. Available online: https://slideplayer.com/slide/17630075/ (accessed on 7 October 2022).
- López-Herrera, C.J.; Pérez-Jiménez, R.M.; Barceló-Muñoz, A.; Zea-Bonilla, T. Evaluación de patrones de aguacate por su tolerancia a la podredumbre blanca. Rev. Chapingo Ser. Hortic. 1999, 5, 267–270. [Google Scholar]
- Zumaquero, A.; Martínez-Ferri, E.; Matas, A.J.; Reeksting, B.; Olivier, N.A.; Pliego-Alfaro, F.; Barceló, A.; van den Berg, N.; Pliego, C. Rosellinia necatrix infection induces differential gene expression between tolerant and susceptible avocado rootstocks. PLoS ONE 2019, 14, e0212359. [Google Scholar] [CrossRef] [Green Version]
- Oficina Española de Variedades Vegetales, Registro de Variedades, Ministerio de Agricultura, Pesca y Alimentación. 2022. Available online: https://www.mapa.gob.es/es/agricultura/temas/medios-de-produccion/semillas-y-plantas-de-vivero/registro-de-variedades/ (accessed on 29 August 2022).
- Grosser, J.; Graham, J.; Hoyte, A. Continued development of rootstocks tolerant of the Phytophthora-Diaprepes complex via greenhouse screening. In Proceedings of the Florida State Horticultural Society, Palm Beach County, FL, USA, 3–5 June 2007; Volume 120, pp. 103–109. [Google Scholar]
- Grosser, J.W.; Omar, A.A.; Gmitter, J.A.; Syvertsen, J.P. Salinity tolerance of ‘Valencia’ orange trees on allotetraploid rootstocks. In Proceeding of the Florida State Horticultural Society, Delray Beach, FL, USA, 3–5 June 2012; Volume 125, pp. 50–55. [Google Scholar]
- Grosser, J.W. Citrus Rootstock Named “UFR-1”. U.S. Patent 2015/0237782 P1, 20 August 2015. [Google Scholar]
- Grosser, J.W. Citrus Rootstock Named “UFR-5”. U.S. Patent PP27,298 P3, 25 October 2016. [Google Scholar]
- Grosser, J.W. Citrus Rootstock Named “UFR-6”. U.S. Patent 2015/0237780 P1, 20 August 2015. [Google Scholar]
- Arjona-López, J.M.; Capote, N.; López-Herrera, C.J. Improved real-time PCR protocol for the accurate detection and quantification of Rosellinia necatrix in avocado orchards. Plant Soil 2019, 443, 605–612. [Google Scholar] [CrossRef]
- Schena, L.; Nigro, F.; Ippolito, A. Identification and detection of Rosellinia necatrix by conventional and real-time Scorpion-PCR. Eur. J. Plant Pathol. 2002, 108, 355–366. [Google Scholar] [CrossRef]
- Campbell, C.L.; Madden, L.V. Temporal analysis of epidemics I: Descriptions and comparisons of disease progress curve. In Introduction to Plant Disease Epidemiology; Campbell, C.L., Madden, L.V., Eds.; Wiley: New York, NY, USA, 1990; pp. 161–202. [Google Scholar]
- Vincent, J.M. Distortion of fungal hyphæ in the presence of certain inhibitors. Nature 1947, 159, 850. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; ISBN 3-900051-07-0. Available online: http://www.r-project.org/ (accessed on 29 June 2022).
- Steel, R.G.D.; Torrie, J.H. Principles and Procedures of Statistics: With Special Reference to the Biological Sciences; McGraw-Hill Book Company, Inc.: New York, NY, USA, 1960. [Google Scholar]
- De Mendiburu, F. Statistical Procedures for Agricultural Research; Package “Agricolae”, Version 1.4-4; Comprehensive R archive network; Institute for Statistics and Mathematics: Vienna, Austria, 2013. [Google Scholar]
- Wickham, H. Data Analysis. In ggplot2. Use R! Springer: Cham, Switzerland, 2016; pp. 189–201. [Google Scholar]


| Effect of R. necatrix | ||
|---|---|---|
| Rootstock | Inoculated | Control (Non-Inoculated) |
| Carrizo citrange | 540.36 ± 29.44 defg | 816.03 ± 14.34 a |
| B11R3T27 | 558.89 ± 24.89 def | 733.27 ± 17.68 b |
| B11R3T53 | 488.93 ± 23.00 ghi | 558.39 ± 19.64 def |
| B11R5T49 | 563.07 ± 25.43 de | 652.77 ± 17.09 c |
| B11R5T60 | 432.57 ± 35.77 ij | 443.48 ± 32.64 hij |
| B11R5T64 | 415.45 ± 31.65 j | 438.08 ± 30.48 hij |
| CL-5146 | 253.37 ± 15.02 l | 472.50 ± 19.64 ghij |
| Orange-14 | 413.12 ± 23.05 j | 577.84 ± 15.70 d |
| UFR-1 | 503.04 ± 21.78 efgh | 724.23 ± 16.21 b |
| UFR-5 | 339.68 ± 20.73 k | 562.60 ± 21.61 de |
| UFR-6 | 491.03 ± 34.29 fghi | 736.13 ± 27.37 b |
| 2247 × 6070-02-2 | 290.85 ± 18.99 kl | 484.22 ± 30.66 ghi |
| Effect of R. necatrix | ||
|---|---|---|
| Rootstock | Inoculated | Control (Non-Inoculated) |
| Carrizo citrange | 64.29 ± 13.78 h | 355.37 ± 15.24 efg |
| B11R3T27 | 538.29 ± 108.22 cde | 1264.55 ± 200.95 ab |
| B11R3T53 | 525.99 ± 87.03 cde | 1108.15 ± 115.55 ab |
| B11R5T49 | 457.17 ± 92.95 def | 777.96 ± 104.30 c |
| B11R5T60 | 236.20 ± 40.98 fgh | 192.99 ± 25.43 gh |
| B11R5T64 | 174.74 ± 29.94 gh | 181.17 ± 15.13 gh |
| CL-5146 | 110.77 ± 17.06 gh | 695.82 ± 117.99 cd |
| Orange-14 | 684.86 ± 130.25 cd | 1337.15 ± 173.95 a |
| UFR-1 | 292.19 ± 79.69 efgh | 1082.67 ± 66.34 ab |
| UFR-5 | 124.19 ± 28.18 gh | 676.36 ± 65.48 cd |
| UFR-6 | 297.11 ± 81.02 efgh | 674.18 ± 105.60 cd |
| 2247 × 6070-02-2 | 546.82 ± 121.05 cde | 1070.27 ± 87.79 b |
| Rootstock | FWAG ± SE | FWR ± SE | DWAG ± SE | DWR ± SE |
|---|---|---|---|---|
| Carrizo citrange | 61.74 ± 3.38 ab | 82.00 ± 1.36 ab | 40.00 ± 5.72 a | 78.46 ± 2.03 a |
| B11R3T27 | 33.73 ± 8.56 de | 77.75 ± 1.86 ab | 24.08 ± 9.66 ab | 64.40 ± 3.33 ab |
| B11R3T53 | 48.44 ± 5.02 abcd | 83.63 ± 1.84 a | 34.86 ± 4.74 ab | 75.00 ± 3.60 ab |
| B11R5T49 | 28.83 ± 9.91 de | 52.05 ± 5.81 c | 18.19 ± 7.44 bc | 45.23 ± 6.14 c |
| B11R5T60 | 5.80 ± 3.48 f | 22.26 ± 9.27 d | 2.77 ± 2.46 c | 23.39 ± 9.85 d |
| B11R5T64 | 22.53 ± 8.02 ef | 42.33 ± 6.84 c | 21.14 ± 7.90 abc | 46.80 ± 6.18 c |
| CL-5146 | 67.08 ± 3.65 a | 83.97 ± 1.64 a | 41.55 ± 6.54 a | 77.34 ± 1.73 ab |
| Orange-14 | 40.94 ± 10.86 bcde | 70.36 ± 4.12 b | 23.52 ± 10.78 abc | 63.90 ± 4.40 b |
| UFR-1 | 56.13 ± 4.71 abc | 75.64 ± 2.40 ab | 32.85 ± 5.13 ab | 69.22 ± 2.51 ab |
| UFR-5 | 67.53 ± 4.31 a | 81.21 ± 2.90 ab | 41.79 ± 5.14 a | 76.37 ± 4.51 ab |
| UFR-6 | 36.97 ± 9.94 cde | 75.45 ± 2.87 ab | 25.92 ± 9.28 ab | 67.92 ± 3.23 ab |
| 2247 × 6070-02-2 | 33.33 ± 11.08 de | 78.95 ± 6.47 ab | 25.15 ± 9.12 ab | 68.88 ± 6.17 ab |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arjona-López, J.M.; Gmitter, F.G., Jr.; Romero-Rodríguez, E.; Grosser, J.W.; Hervalejo, A.; López-Herrera, C.J.; Arenas-Arenas, F.J. Susceptibility of Novel Promising Citrus Rootstocks to White Root Rot. Plants 2022, 11, 3388. https://doi.org/10.3390/plants11233388
Arjona-López JM, Gmitter FG Jr., Romero-Rodríguez E, Grosser JW, Hervalejo A, López-Herrera CJ, Arenas-Arenas FJ. Susceptibility of Novel Promising Citrus Rootstocks to White Root Rot. Plants. 2022; 11(23):3388. https://doi.org/10.3390/plants11233388
Chicago/Turabian StyleArjona-López, Juan M., Frederick G. Gmitter, Jr., Estefanía Romero-Rodríguez, Jude W. Grosser, Aurea Hervalejo, Carlos J. López-Herrera, and Francisco J. Arenas-Arenas. 2022. "Susceptibility of Novel Promising Citrus Rootstocks to White Root Rot" Plants 11, no. 23: 3388. https://doi.org/10.3390/plants11233388