
Wheat Crop under Waterlogging: Potential Soil and Plant Effects
by
 , , , and
, , , and
Isabel P. Pais
1,2,*,
Rita Moreira
1,
José N. Semedo
1,2,
José C. Ramalho
2,3 ,
,
Fernando C. Lidon
2,4,
José Coutinho
2,5,
Benvindo Maçãs
2,5 and
Paula Scotti-Campos
1,2 1
Instituto Nacional de Investigação Agrária e Veterinária, I.P., Quinta do Marquês, Av. República, 2784-505 Oeiras, Portugal
2
GeoBioTec Research Center, Faculdade de Ciências e Tecnologia, Campus da Caparica, Universidade Nova de Lisboa, 2829-516 Caparica, Portugal
3
PlantStress & Biodiversity Lab, Centro de Estudos Florestais (CEF), Instituto Superior Agronomia (ISA), Universidade de Lisboa (ULisboa), Quinta do Marquês, Av. República, 2784-505 Oeiras, Portugal
4
Earth Sciences Department, Faculdade de Ciências e Tecnologia, Campus da Caparica, Universidade Nova de Lisboa, 2829-516 Caparica, Portugal
5
Instituto Nacional de Investigação Agrária e Veterinária, I.P., Estrada Gil Vaz, Ap. 6, 7350-901 Elvas, Portugal
*
Author to whom correspondence should be addressed.
Plants 2023, 12(1), 149; https://doi.org/10.3390/plants12010149
Submission received: 25 November 2022
/
Revised: 22 December 2022
/
Accepted: 23 December 2022
/
Published: 28 December 2022
(This article belongs to the Collection Feature Papers in Plant Physiology and Metabolism)
Abstract
:Inundation, excessive precipitation, or inadequate field drainage can cause waterlogging of cultivated land. It is anticipated that climate change will increase the frequency, intensity, and unpredictability of flooding events. This stress affects 10–15 million hectares of wheat every year, resulting in 20–50% yield losses. Since this crop greatly sustains a population’s food demands, providing ca. 20% of the world’s energy and protein diets requirements, it is crucial to understand changes in soil and plant physiology under excess water conditions. Variations in redox potential, pH, nutrient availability, and electrical conductivity of waterlogged soil will be addressed, as well as their impacts in major plant responses, such as root system and plant development. Waterlogging effects at the leaf level will also be addressed, with a particular focus on gas exchanges, photosynthetic pigments, soluble sugars, membrane integrity, lipids, and oxidative stress.
1. Introduction
Waterlogging is one of the main abiotic stresses affecting crop productivity worldwide [1]. However, due to climate change, the number of flooding events has increased in recent decades, as a result of more severe and unpredictable rainfall [2,3]. Overall waterlogging affects 10 to 15 million ha of wheat cultivated area, causing annual yield losses of 20 to 50% [4]. Global warming will likely boost extreme climatic phenomena, extending the risk of flooding also to new areas [5].
Water plays a vital role in plant growth, being crucial in plants’ interaction with the environment [6,7]. However, excess water in the soil may cause changes in soil properties such as reduced soil oxygen availability, disrupting its diffusion to plant tissues [8] and leading to hypoxic/anoxic conditions [9]. The effects of waterlogging depend on a number of variables, including the depth and duration of waterlogging [10], the crop’s developmental stage [11,12], and the weather conditions [10].
In a waterlogged environment, oxygen deficiency can limit root growth and ultimately lead to root death. In such conditions, energy-dependent processes such as water and nutrient absorption and transport to the shoot are impaired, compromising plant growth and development [10,13,14,15] as well as final yield [15,16,17]. In addition, decreases in leaf nitrogen content, leaf water potential, stomatal conductance, CO2 assimilation rate and photosynthesis, as well as accelerated leaf chlorosis and senescence may also be observed [15,17]. The down-regulation of the photosynthetic machinery in waterlogging sensitive plants results in the excessive production of reactive oxygen species (ROS) causing severe oxidative damage and degradation of cellular structures, interfering with normal metabolism [10,18]. ROS cause lipoperoxidation phenomena resulting in membrane injury, protein degradation, enzyme inactivation, damage to nucleic acids and, eventually, cell death [19,20,21,22].
Wheat is the third most cultivated cereal crop, supplying approximately 20% of the world’s energy and protein requirements [23]. In 2020, wheat global production reached a productivity of ca. 3.5 t ha−1 [24]. Taking into account predictable population growth and climate change scenarios, it is necessary to increase wheat productivity [23], and ensure production stability, two essential components of food security.
We will address some issues related to waterlogging effects in wheat crops, namely soil redox potential, pH, nutrient availability, and electrical conductivity, as well as their impacts in the root system and plant development. We will also focus on major leaf responses related to photosynthetic activity and membrane integrity under oxidative stress conditions.
2. Main Waterlogging Effects in Soil
Waterlogging can occur whenever soil moisture levels go beyond the field capacity. In such scenarios, excess water saturates the soil pores in the presence of a very slim water layer on the soil surface, or even in its absence [12]. This abiotic stress has a negative effect on the majority of terrestrial plants, limiting crop yield. Changes in the physical, chemical, electrochemical, and biological properties of the soil (Table 1) can lead to a decrease in root biomass, hampering vegetative development [10,25,26] and inducing plant organs senescence [27,28].
In a waterlogged soil, all pores are filled with water, changing the ideal solid:pore material (50:50) and soil:air volume (75:25) ratios, which have implications for the plant’s physiological performance [29]. The atmosphere contains 21% oxygen, while the soil should have a concentration of at least 10% [30]. In cultivated soil, the dissolved [O2] is around 0.23 mol m−3, whereas in waterlogged areas, the level is less than 0.05 mmol m−3 [31]. Under normal conditions, the soil atmosphere is rich in CO2 and deficient in O2 [29] due to the aerobic respiration of roots and microorganisms. However, good aeration enables rapid O2 intake and CO2 output, providing an adequate amount of oxygen to meet plant nutritional requirements and allow its development [29]. When waterlogging is established, gas exchanges between soil and atmosphere almost stop as gas diffusion in water is 104 times slower than in air [32]. Plant roots respiration and microbial activity use the oxygen trapped in the soil, promoting a hypoxia/anoxic situation in the rhizosphere [9], resulting in both insufficient O2 levels and toxic CO2 concentrations in soil.
Significant changes in redox potential (Eh) can be caused by soil waterlogging. Eh is the most important physicochemical parameter for the evaluation of the oxidation or reduction level of a flooded soil. In general, soil [O2] is inversely proportional to the Eh. In flooded soils, oxygen deficiency leads to biological reduction processes and a decrease in Eh. According to Søndergaard [33], Eh will be approximately 250 mV and 0 mV for [O2] values of ca. 10% and 1–2%, respectively. Under optimal aeration conditions, cultivated soils have Eh values between +300 and +500 mV [34], with +400 to +450 mV apparently being ideal [35]. In plants roots, the change from aerobic to anaerobic metabolism occurs when [O2] < 1% [33]. In waterlogged soils, Eh can reach values between −300 and +100 mV [35], explaining why growth decreases dramatically in plants susceptible to this stress.
The optimal pH range for the vast majority of cultivated plants is 6.5 to 7 and it is considered ideal for nutrient availability. Nonetheless, values between 5.5 and 8 still provide favorable growth conditions [35]. In waterlogged soils, pH tends toward neutrality, with increases in acidic soils and decreases in alkaline ones [35,36]. Soil pH strongly influences the solubility, mobility, and bioavailability of nutrients and potentially toxic elements, which in turn affects their uptake by plants [37]. Soil acidity is associated with Mo, P, Mg, and Ca deficiency. At a low pH, other elements such as Al, Mn, and Fe may become more available while Al, Fe, and Mn may reach toxic levels to plants [38]. Alkaline soils, on the other hand, are typically deficient in Co and Zn and show less P availability since Ca can bind to it. In these conditions plants tend to be underdeveloped, depicting poor growth and lower yield [10,39,40,41,42].
Nitrogen (N), an essential element for plant growth and one of the most crucial yield-limiting nutrients, is typically extracted from the soil in the inorganic forms of ammonium (NH4+) or nitrate (NO3−). In addition to the amount of nitrogen available in the soil, the form in which it is present can influence plant physiological and metabolic processes, such as nutrient uptake, enzyme activity, photosynthesis and respiration rate, water balance, and signaling pathways [43,44]. Waterlogging significantly reduces gas diffusion, leading to an increase in NH4+ in the soil. Although this ion is an intermediary in numerous metabolic reactions, when it is the only source of N, it may induce a strong inhibition of K uptake, an essential nutrient that is also involved in several important plant physiological processes [44]. Wheat grows preferentially on NO3− nutrition. In waterlogged soils, substantial decreases (15–20%) in wheat growth and yield were reported [10], possibly due high NH4+ levels.
Electrical conductivity (EC) is a good indicative of soil quality [45], as it correlates with the concentration of NO3−, sulphate (SO42−), NH4+, K, S, and Cl, as well as the soil’s nutrient availability. Significant changes in EC can be caused by soil waterlogging. At the onset of that stress, soil EC tends to increase, reach its maximum, and then decrease to stable values. This rise is due to the mobilization of Fe2+ and Mn2+, the accumulation of NH4+, HCO3, and RCOO-, as well as the displacement of cations adsorbed on colloids by Fe2+, Mn2+, and NH4+ [46]. Due to nutrient leaching, the soil may be less fertile after a flood [47]. Low EC values may indicate nutrient deficiency and, if less than 0.10 dS m−1, soil deterioration [22].
3. Plant Responses to Waterlogging
Plants response to waterlogging is highly dependent on a number of factors, such as the depth and duration of exposure and the plant developmental stage [11,41,48,49], among others.
Several authors have identified the wheat reproductive phases, including the stem elongation stage to anthesis and post-anthesis, as the plants’ most sensitive time to waterlogging stress [11,14,50]. Wu et al. [49] found the greatest negative impact at the seedling stage while Pampana et al. [12] observed no differences at the three and four leaf stages. Ding et al. [51] reported wheat yield reductions by 9 to 15% when waterlogging was imposed at the seedling, jointing, and tillering stages and decreased leaf area by 10% and 29% at anthesis and at the milk-ripe stage, respectively. At the tillering stage, reduced spike and grain numbers were reported, whereas waterlogging at booting decreases grain weight [49]. The highest tolerance to waterlogging was found in the period after anthesis followed by the jointing stage [14].
Under natural conditions, waterlogging depths can change, influencing the severity of plant damage. Depending on how deeply water penetrates the soil, total or partial waterlogging can be considered. Total waterlogging occurs when water is present from the soil’s bottom to its surface, affecting all plant organs below ground. On the other hand, partial waterlogging occurs when water does not reach the soil surface and only affects a portion of the root systems [9]. According to Malik et al. [28] bread wheat tillering was reduced by 24, 45, and 62% when the water level was 20, 10, and 0 cm below the soil surface, with decreases of 39, 58, and 73% (respectively) in length of adventitious root main axes per plant.
The duration of waterlogging events has a significant effect on the damage caused to crops. Overall, the longer the duration, the greater the negative effects in plants [52,53]. Reported impacts of waterlogging include changes in plant photosynthesis, respiration, transpiration, and antioxidative system, enhanced organs senescence and reduced accumulation and remobilization of photosynthetic products, that ultimately result in decreased yield components such as spikes number, kernels per spike, and kernel weight [4,10,13,14,52,54]. Lower soil O2 availability is the primary cause of observed negative effects [10,55,56], but anatomical, morphological, or physiological changes may help to mitigate the effects of such deficiency in plants [57].
3.1. Roots
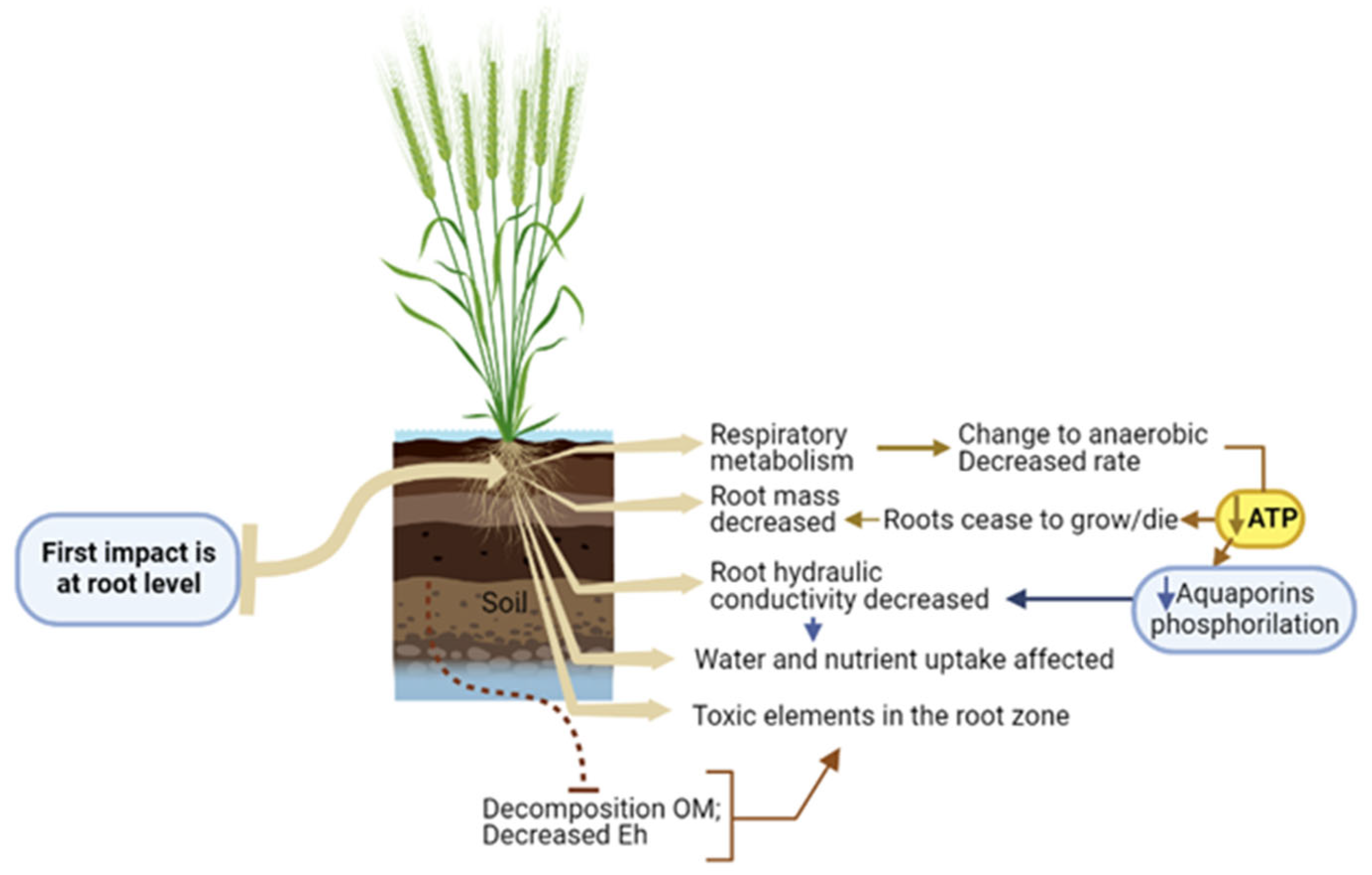
Roots are essential organs for water and nutrients uptake, photoassimilates storage, anchorage, mechanical support, and rhizosphere interface [58,59]. To accomplish this, roots require energy from cellular respiration [60]. Waterlogging conditions primarily affect the plant at the root level, where the initial responses occur [61] (Figure 1).
Root damage causes severe shoot growth impairments [10,15,62]. Arrested root growth and root death significantly decrease seminal root dry mass [10,15]. Under waterlogging conditions low [O2] in the rhizosphere enhances anaerobic respiration [63], resulting in lower ATP production [64]. Energy deficit at the root level impairs aquaporins phosphorylation, which regulates cell water flux, compromising water and nutrient absorption due to a drastic reduction in root hydraulic conductivity [15,65]. Additionally, the drop of soil Eh may increase the availability of Mn2+ and Fe2+ to toxic levels and their accumulation in the roots. Organic acids and other potentially toxic metabolites produced by decomposition of organic matter in anoxic soils may increase in saturated soils. Anaerobic respiration can result in accumulation of lactic acid, ethanol, aldehydes, among others, and in ROS production, namely hydrogen peroxide, which can also cause cellular damage [31,66]. In rice, organic acid toxicity inhibits root respiration, reduces nutrient uptake, increases suberization and/or lignification of outer cell walls, and can cause root occlusions [67]. In barley, adverse effects of organic acids on K+ fluxes in roots were reported [68]. Endogenously-produced CO2 and ethylene can also adversely affect roots, as pH in root cells can became more acidic due to excess CO2, and high ethylene can inhibit root extension [10,32,69].
Some plants tolerant to hypoxia (low [O2]) or anoxia (no O2) can develop morphological adaptations to compensate for the lack of O2 in the root zone [28,70]. Adventitious root formation from the stem or branches is a common response [70] that promotes gas transport, as well as nutrient and water uptake, during waterlogging, significantly enhancing plant survival and productivity [71]. These roots are able to take up and transport O2, making it available to submerged roots [72]. According to Wiengweera and Greenway [73], in wheat plants subjected to waterlogging, adventitious roots absorbed more efficiently P and K than seminal roots. Root architecture can also differ in wheat genotypes, leading to different root distributions in the soil [74]. In flooded areas, shallower root systems may be beneficial in oxygen uptake, since upper soil layers usually have a higher O2 concentration than deeper ones [75,76].
Under waterlogging stress, ethylene accumulation can also activate programmed cell death of root cortical cells, inducing the formation of aerenchyma in adventitious roots. Aerenchyma development in wheat can boost plant tolerance and survival under conditions of waterlogging and oxygen deprivation [77], since it increases energy availability and reduces hypoxic stress [72,78]. This specialized parenchymal tissue has interconnected, large, gas-filled intercellular spaces that provide a low-resistance pathway, hence enhancing gas diffusion between the roots and shoot [72,77]. Furthermore, aerenchyma can discharge CO2 and toxic volatile substances from submerged tissues [31]. However, the internal O2 movement to the apex, which enables root expansion, has a limited extent, and adventitious root growth does not fully compensate seminal root loss. In response to waterlogging, the root:shoot ratio in wheat can decrease from 0.4 to 0.2, reflecting the stronger decrease of root dry mass (up to 62%) when compared with shoot (up to 33%) [10].
3.2. Shoot Development and Productivity
Water uptake by the roots and transpiration from the leaves allow plants growth but, when subjected to waterlogging, susceptible genotypes may present severe impairments of some key metabolic activities, such as photosynthesis, respiration and transpiration. The detrimental effect on these processes causes severe energy deficiency, poor growth, and enhanced leaf/organs senescence, decreasing the accumulation and remobilization of photoassimilates and hence grain yield [15,64,79] (Figure 2).
Several authors report that waterlogging significantly inhibits the growth of aerial parts of the plants, mostly due to a reduced leaf elongation rate, resulting in smaller leaves, but also due to a decrease in the number of tillers and tillers impaired development [10,16,28,62,80,81]. According to Malik et al. [15], the imposition of 3–21 days of waterlogging on 3-week-old plants, followed by 21 or 7 days of recovery, decreased the shoot mass by 43% to 72%, in comparison to well-drained plants over the same period. Herzog et al. [10] reported a 67% reduction in average shoot dry mass of wheat plants grown in waterlogged soil. This decline, together with the overall wilting of the plant and the senescence of the basal leaves, significantly reduces the area available for light absorption and limits photoassimilation. In bread wheat, chlorotic dry mass increased from 8–21% in non-waterlogged plants to 33–70% in 14-days-waterlogged plants [82]. During the recovery period this trend was maintained, with control plants exhibiting lower chlorotic dry mass values (14–36% and 18–43% at 7- and 14-days recovery, respectively) than treated plants (37–95% and 49–100% for the same recovery periods).
3.3. Impacts in Physiological Performance
3.3.1. Gas Exchanges and Sugars Metabolism
Changes in respiration and photosynthesis (Pn) are caused by the high sensitivity of photosynthetic processes to stressful conditions [20,83], frequently used as indicators of productivity under stress conditions. In plants susceptible to waterlogging, physiological activities are drastically reduced and may cause cell death, whereas tolerant plants depict less severe effects or even an improvement in the response of some parameters [28].
As a result of impaired root function, a reduction in shoot physiological performance may occur in wheat plants subjected to waterlogging [10,17,84]. Stomatal closure, reduced transpiration, and photosynthesis inhibition are typical responses to this environmental stress [10,17,84]. Stomatal conductance (gs) is one of the primary factors influencing photosynthesis [85], with a significant effect on photosynthetic rates under waterlogged conditions [10,28]. Stomatal closure and (gs) decrease allow a down-regulation of leaf transpiration [10]. However, this also lowers internal CO2 concentration (Ci), which in turn limits carbon fixation, reducing photosynthesis and increasing respiration, negatively affecting plant production [28,86]. However, photosynthetic rates can also decrease due to non-stomatal factors such as chlorophyll degradation and decreased chlorophyll synthesis, resulting in leaf senescence and yellowing [10,57,80]. Damage to the photosystem II by ROS, decreased photosynthetic enzyme activities, and low nitrogen content exacerbated the decline in photosynthetic activity and the detrimental effects of waterlogging [10,57,80,87]. Through Pn, plants convert carbon dioxide and water into sugars, which are their principal source of energy for different cellular activities [88]. Waterlogging can lead to changes in Pn and respiration, with consequences for sugar metabolism and energetic balance [89]. Lower Pn rates reported at the onset of waterlogging may be related to the accumulation of sugars in the leaves rather than to stomatal closure [10,15]. The accumulation of sugars in leaves of waterlogged wheat plants has been reported [10,15,63].
At the onset of waterlogging, sugars accumulation may result from root hypoxia, which inhibits the rapid growth of both the root and the aboveground portion of the plant. Under such conditions, sugar production in the leaves exceeds its consumption [10,15,28]. Concurrently, constraints in the root system diminish the phloem transport capacity of the roots [10,15], which also contributes to the accumulation of photoassimilates in the leaves. The resulting sugar overproduction, together with a reduced ability for phloem transport in hypoxic roots, leads to further decrease in Pn as a negative feedback of carbohydrate accumulation [10].
In long-term flooding, plants experience energy and carbohydrate deprivation due to reduced photosynthesis and aerobic respiration. Therefore, the initial carbohydrate reserves may be a crucial factor in tolerance to hypoxia/anoxia, as the plant must utilize its stored glucose reserves to maintain metabolic activity under anaerobic conditions [89,90]. Additionally, sugars are also involved in plant stress responses and adaptation, contributing to the stabilization of membrane structures and maintenance of cell turgor through osmotic adjustment and osmoprotection [91].
3.3.2. Chlorophylls and Carotenoids
Photosynthetic pigments are fundamental molecules in the photosynthetic process, their primary function being light absorption and the production of reducing compounds [31,92]. Changes in pigment content and composition have a direct effect on the photosynthetic rate. Chlorophylls are essential for the conversion of light radiation into chemical energy. They are strictly associated with photosynthetic efficiency and, therefore, with plants growth and environmental adaptability [31,92]. Chlorophyll a is present at the reaction centers of both photosystems (PSI and PSII), whereas chlorophyll b is the most important accessory light-absorbing pigment in light-harvesting complexes.
Several authors observed decreases in chlorophyll content with waterlogging. At the emergence stage, a 10-day’s stress induced chlorophyll reductions between 15 and 33% in four wheat genotypes [93]. At the tillering stage, reductions from 41% to 61% were observed in six wheat varieties subjected to 28 days waterlogging [80].
Carotenoids have several functions in plant metabolism. As photosynthetic pigments, they are accessory antenna molecules, harvesting and transferring light energy to chlorophylls during the photosynthetic process. Carotenoids also play an important role in oxidative stress tolerance, contributing to protect the photosynthetic apparatus by scavenging ROS and repressing lipid peroxidation [20]. Waterlogging can affect the concentration of carotenoid pigments and several studies reported their reduction in wheat-susceptible plants [57,81,94]. However, in tolerant genotypes, the amount remained high [95]. Overall, the decline in carotenoid content is more severe in plants subjected to longer waterlogging periods. At the tillering stage, carotenoid contents decreases by 11–15%, 16–38%, and 29–67% after 7, 14, and 21 days of waterlogging, respectively [94]. The same study reported that 14 days of waterlogging at elongation stage caused a more severe carotenoid lowering (32–49%), highlighting different effects according to crop development phase.
3.3.3. Chlorophyll Fluorescence
In the light-harvesting antenna, light photons are captured by chlorophyll and partially (ca. 2%) re-emitted as fluorescence [96]. Chlorophyll fluorescence is a reliable and sensitive tool to assess light-harvesting efficiency in plants [97,98], that complements information obtained through gas exchanges. Under stress conditions, this parameter can decrease, allowing a quantitative comparison of the stress responses, and indirectly providing information on leaf photosynthetic performance.
Maximum quantum efficiency of PSII (Fv/Fm) evaluates the proportion of functional PSII reaction centers. Reductions in this ratio can indicate damage to the photosynthetic apparatus which that may result in Pn decreases [48]. In wheat, declines in Fv/Fm ratio have been reported due to the imposition of waterlogging, indicating impairment of PSII [10,49,80] and consequently, a decreased use-efficiency of captured photon energy [48].
3.3.4. Membrane Integrity and the Role of Lipid Composition
The ability to maintain membrane integrity under stressful conditions ensures cellular compartmentalization and the functioning of metabolic processes, being determinant to protoplasmic tolerance [83]. Waterlogging, as well as other biotic and/or abiotic stresses, can lead to changes at the membrane level, with structural impacts that affect membrane permeability, assessed through increased electrolyte leakage from cells [83,99,100,101], that may reflect severe membrane damage and low survival ability [93,102]. Therefore, membrane stability is frequently used as an indicator of tolerance or susceptibility to environmental stresses [83,103].
Lipids are fundamental plant macromolecules playing key roles in membranes structure, energy storage, and metabolic signaling [104,105,106,107]. In response to abiotic stresses, qualitative and quantitative changes may occur in of the lipid matrix compositions, such remodeling plasticity being crucial for maintenance of membrane integrity [99] significantly contributing to its functionality [66,107]. Hypoxia/anoxia can induce changes in membrane lipids, and it was shown that tolerant plants could increase the degree of unsaturation of membrane lipids, and also enhances lipids biosynthesis under such conditions [102,106].
Lipid remodeling influences the fluidity, integrity, and permeability of plant cell membranes through changes in the composition of lipid classes, the lengths of their carbon skeletons, or the saturation of their fatty acids [106,107]. Hypoxia treatment significantly altered the lipid composition of wheat, with tolerant genotypes exhibiting more efficient lipid remodeling, allowing the bilayer structure of membranes to be preserved during hypoxia stress [107]. Hypoxia reduced the phospholipids phosphatidylcholine (PC) and phosphatidylethanolamine (PE), but the PC:PE ratio increased in the tolerant genotype, thereby limiting the synthesis of non-bilayer membrane phases and conserving fluidity. Non-susceptible plants exhibited considerable increases in phosphatidylglycerol (PG) and phosphatidic acid (PA) as a result of hypoxia. Despite differences in PE content across tolerant and susceptible genotypes, hypoxia-induced alterations followed a similar pattern, suggesting that PE had no contribution to hypoxia tolerance. Several authors have found changes in the glycolipids monogalactosyldiacylglycerol (MGDG) and digalactosyldiacylglycerol (DGDG) in response to flooding [106]. After 4 days of exposure to hypoxia, Xu (2019) [107] observed a 31.6% and 20% decrease in MGDG concentrations in sensitive and tolerant genotypes, respectively. DGDG content was unchanged by hypoxia in the sensitive genotype, whereas in the tolerant genotype, an increase of 25.3% was seen after 2 days of treatment, followed by a drop of 31.1% on the fourth day of hypoxia. Changes in glycolipids can have a significant effect on plants’ tolerance to waterlogging since MGDG is essential for photosynthetic reactions and DGDG is essential for maintaining the maximum efficiency of photosynthetic electron flow by altering PSI and PSII activity [107].
3.3.5. Oxidative Stress
Although ROS are a normal product of plant cell metabolism, biotic and abiotic stresses are often accompanied by oxidative stress, which is characterized by an increase in intracellular ROS. ROS can be divided into free radicals (superoxide radical (O2•−), hydroxyl radical (OH•), perhydroxy radical (HO2•)) and non-radicals (singlet oxygen (1O2), hydrogen peroxide (H2O2)) [31]. When accumulated in mesophyll cells, their strong oxidizing activity can lead to lipoperoxidation and degradation of membrane lipids, and cause oxidative damage to proteins and DNA, resulting in severe cell injuries [22,31,108]. Carbon–carbon double bonds in lipids are preferential targets for ROS, meaning that cell membranes are rich in polyunsaturated fatty acids (PUFA), particularly abundant in chloroplasts, and are extremely susceptible to lipoperoxidation [109,110]. In waterlogging-susceptible plants, the downregulation of the photosynthetic machinery leads to excessive ROS generation within the leaf. Rapid chlorosis of basal leaves precedes premature leaf senescence caused by the remobilization of nitrogen to younger leaves [10,111]. Decreased chlorophyll content in the remaining leaves is an indicator of oxidative stress [88]. Excess water can increase ROS content many-fold higher than normal growing conditions, causing severe oxidative damage to plant cells [18]. This rise suggests the presence of lipoperoxidation events [112] and is commonly associated with greater concentrations of malonyldialdehyde (MDA), one of several lipid oxidation products [109]. Genotypes showing lower MDA levels under stress may be more resistant to oxidative stress [110]. ROS accumulation also causes a significant disruption to plant ionic homeostasis, directly influencing the functioning of various cation [113] and anion channels [114]. Wheat plants can overcome oxidative stress through activation of antioxidative defense systems involving enzymatic and non-enzymatic mechanisms to neutralize excessive ROS and reduce the extent of oxidative damage [1,18,19,20,54].
By removing, neutralizing, or scavenging ROS and their intermediates, antioxidant enzymes such as superoxide dismutase (SOD), catalase (CAT), glutathione reductase (GR), and ascorbate peroxidase (APX) disrupt the cascades of uncontrolled oxidation, converting ROS into harmless compounds [90]. Non-enzymatic antioxidants, such as reduced glutathione (GSH), ascorbic acid (AsA), carotenoids, and tocopherols, play a crucial role in membrane stabilization and cellular components protection. Several reports of increased antioxidant enzyme activity as a result of waterlogging, have also been reported in tolerant wheat plants [31,94].
3.4. Waterlogging Effects in Yield Components
Wheat is considered susceptible to waterlogging, and decreases in grain yield per plant have been widely reported under such stress [12,52,57,62,81]. Waterlogging for 30 days throughout sowing, seedling, flowering, and grain-filling reduced grain yield by 50–70% due to poor seed set and fewer spikes per unit area [52,115]. At the tillering stage, 21 days of waterlogging resulted in lower yield with value reductions from 37% to 60% [94]. In waterlogging-susceptible genotypes, reductions were also observed in the number of spikes per plant [116], the number of grains per spike [4], and the thousand kernel weight [117].
The emission, development, and survival of tillers are crucial features since they are directly proportional to the number of spikes per unit area [118], which has a straight effect on wheat yield [28]. Studies with different wheat genotypes subjected to waterlogging during the tillering phase revealed that a reduction in the number of tillers emitted was not always accompanied by a reduction in the number of fertile ones, indicating a good strategy of the plant to maintain production to cope with energy deficit [28,62,81]. If the number of fertile tillers remains unaltered, the decline in productivity results from a small contribution of fertile tillers to the formation of the final yield [118]. Yield decreases under stress were associated with reduced production [15,28] due to low tillers survival, fewer and smaller fertile tillers, and smaller grains [119,120,121]. A significant decrease in seed number per spike (2.0 to 78.8%) with increasing time of waterlogging periods (5 to 50 days), with the lowest value at the 50th day and the highest seed number per spike in control plants [52]. Ding et al., (2020) [51] reported decreases in single-spike yield (9%) and in kernels per spikelet (5%) in waterlogged wheat, although no changes were observed concerning spikelet fertility and spikelets per spike. The same trend was found by Alizadeh-Vaskasi (2018) [94], who reported reductions in kernels-per-spike and single-spike yield in wheat plants subjected to waterlogging at tillering and elongation stages.
3.5. Genetic Responses to Waterlogging
Adverse environments, such as waterlogged soil, may induce several changes at morphological and physiological levels in plants. To enhance waterlogging tolerance, a vast number of stress response genes are activated and essential functional proteins are synthesized [122,123]. Wei et al., (2021) [124] observed a considerable down-regulation of photosynthesis-related genes (e.g., PsbQ, PsbO, and petF) and light-harvesting chlorophyll protein complex genes (e.g., LHCB1, LHCB3, LHCB5, LHCA1, and LHCA4) in two wheat genotypes in response to waterlogging. Borrego-Benjumea et al., (2020) [125] observed the participation of genes involved in multiple metabolic pathways at the root level of barley, including glucose and nitrogen metabolisms. The down-regulation of genes involved in ROS detoxification, nitrogen, and amino acid metabolism was also identified in the same study. In wheat, Tong et al., (2021) [126] reported an enhanced expression of RBOH (Respiratory Burst Oxidase Homolog), which regulates ROS accumulation. As the mechanism of ROS production/scavenging is essential for the regulation of aerenchyma development in roots, genes implicated in this mechanism have been considered candidates for waterlogging tolerance [126]. Same authors also found multiple Quantitative Trait Locus (QTLs) for waterlogging tolerance features related to root fresh biomass (QRfbio.ua-1B-WGH), shoot fresh biomass (QSfbio.ua-1B-WGH), chlorophyll content (QSpadpost.ua-1B-WF and QSpad.ua-1D.5), and germination rate index (GRI-7A). The knowledge of the genetic pathways linked with this stress can be of great value to the molecular breeding of waterlogging-tolerant wheat.
4. Conclusions
The increased frequency and intensity of extreme weather events, such as waterlogging episodes, resulting from global warming, is one of the challenges to maintaining/improve wheat yield. Plant growth and development processes under waterlogging depend on morphological, physiological, and biochemical adaptation, and on the gene regulation modulating such traits. Identifying key traits underlying tolerance responses and understanding their roles in adaptation to waterlogging will contribute to develop more adapted wheat plants and to boost wheat yield and grain quality under a changing climate.
Author Contributions
Conceptualization, I.P.P. and P.S.-C.; writing—original draft preparation, I.P.P.; writing—review and editing, I.P.P., P.S.-C., R.M., J.N.S. and J.C.R.; supervision, P.S.-C., F.C.L., J.C. and B.M. All authors have read and agreed to the published version of the manuscript.
Funding
This work received funding from Fundação para a Ciência e a Tecnologia, I.P. (FCT), Portugal, through the research units UIDP/04035/2020 (GeoBioTec), and UIDB/00239/2020 (CEF).
Data Availability Statement
Not applicable.
Acknowledgments
The authors thank the research center (GeoBioTec) UIDB/04035/2020 for support facilities.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bailey-Serres, J.; Voesenek, L.A.C.J. Flooding stress: Acclimations and genetic diversity. Ann. Rev. Plant Biol. 2008, 59, 313–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- IPCC. Climate Change 2021: The Physical Science Basis—Summary for the Policymakers (Working Group I). In Climate Change 2021: The Physical Science Basis; Masson-Delmotte, V., Zhai, P., Pirani, A., Connors, S.L., Péan, C., Chen, Y., Goldfarb, L., Gomis, M.I., Matthews, J.B.R., Berger, S., Eds.; Cambridge University Press: Cambridge, UK, 2021. [Google Scholar]
- Pais, I.P.; Reboredo, F.H.; Ramalho, J.C.; Pessoa, M.F.; Lidon, F.C.; Silva, M.M. Potential impacts of climate change on agriculture-A review. Emir. J. Food Agric. 2020, 32, 397–407. [Google Scholar] [CrossRef]
- Hossain, A.; Uddin, S.N. Mechanisms of waterlogging tolerance in wheat: Morphological and metabolic adaptations under hypoxia or anoxia. Aust. J. Crop Sci. 2011, 5, 1094–1101. [Google Scholar]
- Feyen, L.; Dankers, R.; Bódis, K. Fluvial flood risk in Europe in present and future climates. Clim. Change 2012, 112, 47–62. [Google Scholar] [CrossRef]
- Gornall, J.; Betts, R.; Burke, E.; Clark, R.; Camp, J.; Willett, K.; Wiltshire, A. Implications of climate change for agricultural productivity in the early twenty-first century. Phil. Trans. R. Soc. B 2010, 365, 2973–2989. [Google Scholar] [CrossRef]
- Yu, Q.; Li, L.; Luo, Q.; Eamus, D.; Xu, S.; Chen, C.; Wang, E.; Liu, J.; Nielsen, D.C. Year patterns of climate impact on wheat yields. Int. J. Climatol. 2014, 34, 518–528. [Google Scholar] [CrossRef]
- Lee, S.C.; Mustroph, A.; Sasidharan, R.; Vashisht, D.; Pedersen, O.; Oosumi, T.; Voesenek, L.A.C.J.; Bailey-Serres, J. Molecular characterization of the submergence response of the Arabidopsis thaliana ecotype Columbia. New Phytol. 2011, 190, 457–471. [Google Scholar] [CrossRef]
- Sasidharan, R.; Bailey-Serres, J.; Ashikari, M.; Atwell, B.J.; Colmer, T.D.; Fagerstedt, K.; Fukao, T.; Geigenberger, P.; Hebelstrup, K.H.; Hill, R.D.; et al. Community recommendations on terminology and procedures used in flooding and low oxygen stress research. New Phytol. 2017, 214, 1403–1407. [Google Scholar] [CrossRef] [Green Version]
- Herzog, M.; Striker, G.G.; Colmer, T.D.; Pedersen, O. Mechanisms of waterlogging tolerance in wheat—A review of root and shoot physiology. Plant Cell Environ. 2016, 39, 1068–1086. [Google Scholar] [CrossRef]
- De San Celedonio, R.P.; Abeledo, L.G.; Miralles, D.J. Identifying the critical period for waterlogging on yield and its components in wheat and barley. Plant Soil 2014, 378, 265–277. [Google Scholar] [CrossRef]
- Pampana, S.; Masoni, A.; Arduini, I. Grain yield of durum wheat as affected by waterlogging at tillering. Cereal Res. Commun. 2016, 44, 706–716. [Google Scholar] [CrossRef]
- Araki, H.; Hossain, M.A.; Takahashi, T. Waterlogging and hypoxia have permanent effects on wheat root growth and respiration. J. Agron. Crop Sci. 2012, 198, 264–275. [Google Scholar] [CrossRef]
- Araki, H.; Hamada, A.; Hossain, M.A.; Takahashi, T. Waterlogging at jointing and/or after anthesis in wheat induces early leaf senescence and impairs grain filling. Field Crops Res. 2012, 137, 27–36. [Google Scholar] [CrossRef]
- Malik, A.I.; Colmer, T.D.; Lambers, H.; Setter, T.L.; Schortemeyer, M. Short-term waterlogging has long-term effects on the growth and physiology of wheat. New Phytol. 2002, 153, 225–236. [Google Scholar] [CrossRef]
- Dickin, E.; Wright, D. The effects of winter waterlogging and summer drought on the growth and yield of winter wheat (Triticum aestivum L.). Eur. J. Agron. 2008, 28, 234–244. [Google Scholar] [CrossRef]
- Zheng, C.; Jiang, D.; Liu, F.; Dai, T.; Jing, Q.; Cao, W. Effects of salt and waterlogging stresses and their combination on leaf photosynthesis, chloroplast ATP synthesis, and antioxidant capacity in wheat. Plant Sci. 2009, 176, 575–582. [Google Scholar] [CrossRef]
- Lal, M.; Kumari, A.; Pooja; Sheokand, S. Reactive oxygen species, reactive nitrogen species and oxidative metabolism under waterlogging stress. In Reactive Oxygen, Nitrogen and Sulfur Species in Plants: Production, Metabolism, Signaling and Defense Mechanisms; Hasanuzzaman, M., Fotopoulos, V., Nahar, K., Fujita, M., Eds.; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2019; pp. 777–812. [Google Scholar] [CrossRef]
- Bali, A.S.; Sidhu, G.P.S. Abiotic stress-induced oxidative stress in wheat. In Wheat Production in Changing Environments; Hasanuzzaman, M., Nahar, K., Hossain, M.A., Eds.; Springer: Singapore, 2019; pp. 225–239. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Mahmud, J.A.; Nahar, K.; Anne, T.I.; Inafuku, M.; Oku, H.; Fujita, M. Responses, adaptation, and ROS metabolism in plants exposed to waterlogging stress. In Reactive Oxygen Species and Antioxidant Systems in Plants: Role and Regulation under Abiotic Stress, 1st ed.; Khan, M.I.R., Khan, N.A., Eds.; Springer: Singapore, 2017; pp. 257–281. [Google Scholar] [CrossRef]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. J. Bot. 2012, 2012, 1–26. [Google Scholar] [CrossRef]
- Hawkesford, M.J.; Araus, J.L.; Park, R.; Calderini, D.; Miralles, D.; Shen, T.; Zhang, J.; Parry, M.A.J. Prospects of doubling global wheat yields. Food Energy Secur. 2013, 2, 34–48. [Google Scholar] [CrossRef]
- FAOSTAT. 2020 Food and Agriculture Organization of the United Nations, Statistics Division. Available online: http://www.fao.org/faostat/en/#data/ (accessed on 2 November 2022).
- De San Celedonio, R.P.; Abeledo, L.G.; Brihet, J.M.; Miralles, D.J. Waterlogging affects leaf and tillering dynamics in wheat and barley. J. Agron. Crop Sci. 2016, 202, 409–420. [Google Scholar] [CrossRef]
- Pierret, A.; Doussan, C.; Capowiez, Y.; Bastardie, F.; Pagès, L. Root functional architecture: A framework for modeling the interplay between roots and soil. Vadose Zone J. 2007, 6, 269–281. [Google Scholar] [CrossRef]
- Jimenez, J.C.; Moreno, L.P.; Magnitskiy, S. Respuesta de las plantas a estres por inundacion. Rev. Colomb. Cienc. Hort. 2012, 6, 96–109. [Google Scholar] [CrossRef] [Green Version]
- Malik, A.I.; Colmer, T.D.; Lambers, H.; Schortemeyer, M. Wheat in response to different depths of waterlogging. Austr. J. Plant Physiol. 2001, 28, 1121–1131. [Google Scholar]
- Morales-Olmedo, M.; Ortiz, M.; Sellés, G. Effects of transient soil waterlogging and its importance for rootstock selection. Chil. J. Agric. Res. 2015, 75, 45–56. [Google Scholar] [CrossRef] [Green Version]
- Goud, E.L.; Singh, J.; Kummar, P. Climate change and their impact on global food production. In Microbiome under Changing Climate, Implications and Solutions; Kumar, A., Singh, J., Ferreira, L.F.R., Eds.; Woodhead Publishing: Sawston, UK, 2022; pp. 415–436. [Google Scholar] [CrossRef]
- Pan, J.; Sharif, R.; Xu, X.; Chen, X. Mechanisms of waterlogging tolerance in plants: Research progress and prospects. Front. Plant Sci. 2021, 11, 627331. [Google Scholar] [CrossRef]
- Greenway, H.; Armstrong, W.; Colmer, T.D. Conditions leading to high CO2 (>5 kPa) in waterlogged-flooded soils and possible effects on root growth and metabolism. Ann. Bot. 2006, 98, 9–32. [Google Scholar] [CrossRef] [Green Version]
- Søndergaard, M. Redox Potential Definitions and General Aspects. In Encyclopedia of Inland Waters; Elsevier: Amsterdam, The Netherlands, 2009; pp. 852–859. [Google Scholar]
- Macías, F.; Arbestain, M.C. Soil carbon sequestration in a changing global environment. Mitig. Adapt. Strateg. Glob. Change 2010, 15, 511–529. [Google Scholar] [CrossRef]
- Husson, O. Redox potential (Eh) and pH as drivers of soil/plant/microorganism systems: A transdisciplinary overview pointing to integrative opportunities for agronomy. Plant Soil 2013, 362, 389–417. [Google Scholar] [CrossRef]
- Parent, C.; Capelli, N.; Berger, A.; Crèvecoeur, M.; Dat, J. An overview of plant responses to soil waterlogging. Plant Stress 2008, 2, 20–27. [Google Scholar]
- Förstner, U. Land contamination by metals: Global scope and magnitude of problem. In Metal Speciation and Contamination of Soil; Allen, H.E., Huang, C.P., Bailey, G.W., Bowers, A.R., Eds.; Lewis Publishers: Boca Raton, FL, USA, 1995; pp. 1–33. Available online: https://www.researchgate.net/publication/234079750 (accessed on 2 November 2022).
- Liu, K.; Harrison, M.T.; Shabala, S.; Meinke, H.; Ahmed, I.; Zhang, Y.; Tian, X.; Zhou, M. The state of the art in modeling waterlogging impacts on plants: What do we know and what do we need to know. Earth’s Future 2020, 8, e2020EF001801. [Google Scholar] [CrossRef]
- Mantri, N.; Patade, V.; Penna, S.; Ford, R.; Pang, E. Abiotic stress responses in plants: Present and future. In Abiotic Stress Responses in Plants; Ahmad, P., Prasad, M., Eds.; Springer: New York, NY, USA, 2012; pp. 1–20. [Google Scholar] [CrossRef]
- Prasad, R.P.J.F. Soil Acidity. In Soil Fertility Management for Sustainable Agriculture, 1st ed.; CRC Press: Boca Raton, FL, USA, 1997; pp. 67–92. [Google Scholar]
- Setter, T.L.; Waters, I.; Sharma, S.K.; Singh, K.N.; Kulshreshtha, N.; Yaduvanshi, N.P.S.; Ram, P.C.; Singh, B.N.; Rane, J.; McDonald, G.; et al. Review of wheat improvement for waterlogging tolerance in Australia and India: The importance of anaerobiosis and element toxicities associated with different soils. Ann. Bot. 2009, 103, 221–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, K.; Yu, L.; Xu, J.; Brookes, P.C. pH, nitrogen mineralization, and KCl-extractable aluminum as affected by initial soil pH and rate of vetch residue application: Results from a laboratory study. J. Soils Sediments 2014, 14, 1513–1525. [Google Scholar] [CrossRef]
- Guo, S.; Zhou, Y.; Shen, Q.; Zhang, F. Effect of ammonium and nitrate nutrition on some physiological processes in higher plants—Growth, photosynthesis, photorespiration, and water relations. Plant Biol. 2007, 9, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Jia, Y.; Chen, H.; Zhang, L.; Yang, J.; Zhang, J.; Hu, X.; Ye, X.; Li, Y.; Zhou, Y. Growth, photosynthesis, and nutrient uptake in wheat are affected by differences in nitrogen levels and forms and potassium supply. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bünemann, E.K.; Mäder, P.; Wohlfahrt, J.; Brussaard, L.; Bongiorno, G.; de Goede, R.; Geissen, V.; Fleskens, L.; Sukkel, W.; Bai, Z.; et al. Concepts and Indicators of Soil Quality—A Review; Report 04; ISQPAPER: Wageningen, The Netherlands, 2016; Available online: www.iSQAPER-project.eu (accessed on 2 November 2022).
- Camargo, F.A.D.O.; Santos, G.D.A.; Zonta, E. Alterações eletroquímicas em solos inundados. Ciência Rural 1999, 29, 171–180. [Google Scholar] [CrossRef] [Green Version]
- Santos, A.B.; Fageria, N.K.; Zimmermann, F.J.P. Atributos químicos do solo afetado pelo manejo da água e do fertilizante potássico na cultura de arroz irrigado. Rev. Brasil. Eng. Agrícola Ambient. 2002, 6, 12–16. [Google Scholar] [CrossRef]
- Shao, G.C.; Lan, J.J.; Yu, S.E.; Liu, N.; Guo, R.Q.; She, D.L. Photosynthesis and growth of winter wheat in response to waterlogging at different growth stages. Photosynthetica 2013, 51, 429–437. [Google Scholar] [CrossRef]
- Wu, X.; Tang, Y.; Li, C.; Wu, C.; Huang, G. Chlorophyll fluorescence and yield responses of winter wheat to waterlogging at different growth stages. Plant Prod. Sci. 2015, 18, 284–294. [Google Scholar] [CrossRef]
- Setter, T.L.; Waters, I. Review of prospects for germplasm improvement for waterlogging tolerance in wheat, barley and oats. Plant Soil 2003, 253, 1–34. [Google Scholar] [CrossRef]
- Ding, J.; Liang, P.; Wu, P.; Zhu, M.; Li, C.; Zhu, X.; Gao, D.; Chen, Y.; Guo, W. Effects of waterlogging on grain yield and associated traits of historic wheat cultivars in the middle and lower reaches of the Yangtze River, China. Field Crops Res. 2020, 246, 107695. [Google Scholar] [CrossRef]
- Olgun, M.; Kumlay, A.M.; Adiguzel, M.C.; Caglar, A. The effect of waterlogging in wheat (T. aestivum L.). Acta Agric. Scand. Sect. B Soil Plant Sci. 2008, 58, 193–198. [Google Scholar] [CrossRef]
- Tian, L.X.; Zhang, Y.C.; Chen, P.L.; Zhang, F.F.; Li, J.; Yan, F.; Dong, Y.; Feng, B.L. How does the waterlogging regime affect crop yield? A global meta-analysis. Front. Plant Sci. 2021, 12, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Cai, J.; Liu, F.; Jiang, D.; Dai, T.; Cao, W. Generation and scavenging of reactive oxygen species in wheat flag leaves under combined shading and waterlogging stress. Funct. Plant Biol. 2012, 39, 71–81. [Google Scholar] [CrossRef]
- Colmer, T.D.; Voesenek, L.A.C.J. Flooding tolerance: Suites of plant traits in variable environments. Funct. Plant Biol. 2009, 36, 665–681. [Google Scholar] [CrossRef]
- Voesenek, L.A.C.J.; Bailey-Serres, J. Flood adaptive traits and processes: An overview. New Phytol. 2015, 206, 57–73. [Google Scholar] [CrossRef] [PubMed]
- Ploschuk, R.A.; Miralles, D.J.; Colmer, T.D.; Ploschuk, E.L.; Striker, G.G. Waterlogging of winter crops at early and late stages: Impacts on leaf physiology, growth and yield. Front. Plant Sci. 2018, 871, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hendel, E.; Bacher, H.; Oksenberg, A.; Walia, H.; Schwartz, N.; Peleg, Z. Deciphering the genetic basis of wheat seminal root anatomy uncovers ancestral axial conductance alleles. Plant Cell Environ. 2021, 44, 1921–1934. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, H.; Pradal, C. Root phenotyping: Important and minimum information required for root modeling in crop plants. Breed. Sci. 2021, 71, 109–116. [Google Scholar] [CrossRef]
- Fagerstedt, K.V.; Blokhina, O.B.; Pucciariello, C.; Perata, P. Flooding tolerance mechanisms in roots. In Plant Roots: The Hidden Half, 4th ed.; Eshel, A., Beeckman, T., Eds.; CRC Press Taylor & Francis Group: Boca Raton, FL, USA, 2013; Chapter 32; 32p. [Google Scholar] [CrossRef]
- Chen, Y.; Palta, J.; Prasad, P.V.V.; Siddique, K.H.M. Phenotypic variability in bread wheat root systems at the early vegetative stage. BMC Plant Biol. 2020, 20, 1–16. [Google Scholar] [CrossRef]
- Robertson, D.; Zhang, H.; Palta, J.A.; Colmer, T.; Turner, N.C. Waterlogging affects the growth, development of tillers, and yield of wheat through a severe, but transient, N deficiency. Crop Pasture Sci. 2009, 60, 578–586. [Google Scholar] [CrossRef]
- Huang, B.; Johnson, J.W. Root respiration and carbohydrate status of two wheat genotypes in response to hypoxia. Ann. Bot. 1995, 75, 427–432. [Google Scholar] [CrossRef]
- Gibbs, J.; Greenway, H. Mechanisms of anoxia tolerance in plants. I. Growth, survival and anaerobic catabolism. Funct. Plant Biol. 2003, 30, 1–47. [Google Scholar] [CrossRef] [PubMed]
- Tan, X.; Xu, H.; Khan, S.; Equiza, M.A.; Lee, S.H.; Vaziriyeganeh, M.; Zwiazek, J.J. Plant water transport and aquaporins in oxygen-deprived environments. J. Plant Physiol. 2018, 227, 20–30. [Google Scholar] [CrossRef] [PubMed]
- Blokhina, O.; Virolainen, E.; Fagerstedt, K.V. Antioxidants, oxidative damage and oxygen deprivation stress: A review. Ann. Bot. 2003, 91, 179–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colmer, T.D.; Kotula, L.; Malik, A.I.; Takahashi, H.; Konnerup, D.; Nakazono, M.; Pedersen, O. Rice acclimation to soil flooding: Low concentrations of organic acids can trigger a barrier to radial oxygen loss in roots. Plant Cell Environ. 2019, 42, 2183–2197. [Google Scholar] [CrossRef] [PubMed]
- Pang, J.; Cuin, T.; Shabala, L.; Zhou, M.; Mendham, N.; Shabala, S. Effect of secondary metabolites associated with anaerobic soil conditions on ion fluxes and electrophysiology in barley roots. Plant Physiol. 2007, 145, 266–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, B.; Johnson, J.W.; Box, J.E.; NeSmith, D.S. Root characteristics and hormone activity of wheat in response to hypoxia and ethylene. Crop Sci. 1997, 37, 812–818. [Google Scholar] [CrossRef]
- Ayi, Q.; Zeng, B.; Liu, J.; Li, S.; van Bodegom, P.M.; Cornelissen, J.H.C. Oxygen absorption by adventitious roots promotes the survival of completely submerged terrestrial plants. Ann. Bot. 2016, 118, 675–683. [Google Scholar] [CrossRef]
- Steffens, B.; Steffen-Heins, A.; Sauter, M. Reactive oxygen species mediate growth and death in submerged plants. Front. Plant Sci. 2013, 4, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Steffens, B.; Rasmussen, A. The physiology of adventitious roots. Plant Physiol. 2016, 170, 603–617. [Google Scholar] [CrossRef] [Green Version]
- Wiengweera, A.; Greenway, H. Performance of seminal and nodal roots of wheat in stagnant solution: K+ and P uptake and effects of increasing O2 partial pressures around the shoot on nodal root elongation. J. Exp. Bot. 2004, 55, 2121–2129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oyanagi, A.; Kiribuchi-Otobe, C.; Yanagisawa, T.; Miura, S.; Kobayashi, H.; Muranaka, S. Growth and grain yield of wheat experimental lines with deep and shallow root system in wet paddy fields. Jpn. J. Crop Sci. 2004, 73, 300–308. [Google Scholar] [CrossRef]
- Omori, F.; Mano, Y. QTL mapping of root angle in F2 populations from maize ‘B73’ × teosinte ‘Zea luxurians’. Plant Root 2007, 1, 57–65. [Google Scholar] [CrossRef] [Green Version]
- Pais, I.P.; Moreira, R.; Semedo, J.N.; Reboredo, F.H.; Lidon, F.C.; Coutinho, J.; Maçãs, B.; Scotti-Campos, P. Phenotypic Diversity of Seminal Root Traits in Bread Wheat Germplasm from Different Origins. Plants 2022, 11, 2842. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, T.; Watanabe, K.; Fukazawa, A.; Mori, H.; Abe, F.; Kawaguchi, K.; Oyanagi, A.; Nakazono, M. Ethylene and reactive oxygen species are involved in root aerenchyma formation and adaptation of wheat seedlings to oxygen-deficient conditions. J. Exp. Bot. 2014, 65, 261–273. [Google Scholar] [CrossRef] [Green Version]
- Striker, G.G.; Insausti, P.; Grimoldi, A.A.; Vega, A.S. Trade-off between root porosity and mechanical strength in species with different types of aerenchyma. Plant Cell Environ. 2007, 30, 580–589. [Google Scholar] [CrossRef]
- De San Celedonio, R.P.; Abeledo, L.G.; Miralles, D.J. Physiological traits associated with reductions in grain number in wheat and barley under waterlogging. Plant Soil 2018, 429, 469–481. [Google Scholar] [CrossRef]
- Amri, M.; El Ouni, M.H.; Salem, M.B. Waterlogging affect the development, yield and components, chlorophyll content and chlorophyll fluorescence of six bread wheat genotypes (Triticum aestivum L.). Bulg. J. Agric. Sci. 2014, 20, 647–657. [Google Scholar]
- Collaku, A.; Harrison, S.A. Losses in wheat due to waterlogging. Crop Sci. 2002, 42, 444–450. [Google Scholar] [CrossRef]
- Pais, I.P.; Moreira, R.; Semedo, J.N.; Reboredo, F.H.; Lidon, F.C.; Maçãs, B.; Scotti-Campos, P. Effects of waterlogging on growth and development of bread wheat genotypes. Biol. Life Sci. Forum 2022, 11, 38. [Google Scholar] [CrossRef]
- Scotti-Campos, P.; Semedo, J.N.; Pais, I.; Oliveira, M.; Passarinho, J.; Ramalho, J.C. Heat tolerance of Portuguese old bread wheat varieties. Emir. J. Food Agric. 2014, 26, 170–179. [Google Scholar] [CrossRef] [Green Version]
- Bhagat, K.P.; Kumar, R.A.; Ratnakumar, P.; Kumar, S.; Bal, S.K.; Agrawal, P.K. Photosynthesis and associated aspects under abiotic stresses environment. In Approaches to Plant Stress and Their Management; Gaur, R.K., Sharma, P., Eds.; Springer: Delhi, India, 2014; pp. 191–205. [Google Scholar] [CrossRef]
- Medrano, H.; Escalona, J.M.; Bota, J.; Gulías, J.; Flexas, J. Regulation of photosynthesis of C3 plants in response to progressive drought: Stomatal conductance as a reference parameter. Ann. Bot. 2002, 89, 895–905. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.D.; Li, J.C.; Wei, F.Z.; Wang, C.Y.; Zhang, Y.; Sun, G. Effects of nitrogen spraying on the post-anthesis stage of winter wheat under waterlogging stress. Acta Physiol. Plant. 2014, 36, 207–216. [Google Scholar] [CrossRef]
- Langan, P.; Bernád, V.; Walsh, J.; Henchy, J.; Khodaeiaminjan, M.; Mangina, E.; Negrão, S. Phenotyping for waterlogging tolerance in crops: Current trends and future prospects. J. Exp. Bot. 2022, 73, 5149–5169. [Google Scholar] [CrossRef]
- Singh, J.; Thakur, J.K. Photosynthesis and abiotic stress in plants. In Biotic and Abiotic Stress Tolerance in Plant; Vats, E., Ed.; Springer: Berlin/Heidelberg, Germany, 2018; pp. 27–46. [Google Scholar] [CrossRef]
- Fukao, T.; Barrera-Figueroa, B.E.; Juntawong, P.; Peña-Castro, J.M. Submergence and waterlogging stress in plants: A review highlighting research opportunities and understudied aspects. Front. Plant Sci. 2019, 10, 1–24. [Google Scholar] [CrossRef]
- Sharma, S.K.; Kulshreshtha, N.; Kumar, A.; Yaduvanshi, N.P.S.; Singh, M.; Prasad, K.R.K.; Basak, N. Waterlogging effects on elemental composition of wheat genotypes in sodic soils. J. Plant Nutr. 2018, 41, 1252–1262. [Google Scholar] [CrossRef]
- Sharma, A.; Shahzad, B.; Kumar, V.; Kohli, S.K.; Sidhu, G.P.S.; Bali, A.S.; Handa, N.; Kapoor, D.; Bhardwaj, R.; Zheng, B. Phytohormones regulate accumulation of osmolytes under abiotic stress. Biomolecules 2019, 9, 285. [Google Scholar] [CrossRef] [Green Version]
- Anjum, S.A.; Xie, X.; Wang, L.; Saleem, M.F.; Man, C.; Lei, W. Morphological, physiological and biochemical responses of plants to drought stress. Afr. J. Agric. Res. 2011, 6, 2026–2032. [Google Scholar] [CrossRef]
- Yadav, V.K.; Kajla, M.; Singh, S.P.; Singh, A.K.; Yadav, R.K.; Dwivedi, A.K. Effect of waterlogging tolerance in wheat (Tritium aestivum L.) at ear emergence stage on growth, biochemical and yield parameters in sodic soil. J. App. Nat. Sci. 2015, 7, 949–954. [Google Scholar] [CrossRef] [Green Version]
- Alizadeh-Vaskasi, F.; Pirdashti, H.; Cherati Araei, A.; Saadatmand, S. Waterlogging effects on some antioxidant enzymes activities and yield of three wheat promising lines. Acta Agric. Slov. 2018, 111, 621–631. [Google Scholar] [CrossRef] [Green Version]
- Ozcubukcu, S.; Ergun, N. Effects of waterlogging and nitric oxide on chlorophyll and carotenoid pigments of wheat. J. Food Agric. Environ. 2013, 11, 2319–2323. [Google Scholar] [CrossRef]
- Cessna, S.; Demmig-Adams, B.; Adams, W.W. Exploring photosynthesis and plant stress using inexpensive chlorophyll fluorometers. J. Nat. Resour. Life Sci. Educ. 2010, 39, 22–30. [Google Scholar] [CrossRef]
- Lazár, D. Chlorophyll a fluorescence induction. Biochim. Biophys. Acta 1999, 1412, 1–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lazár, D. The polyphasic chlorophyll a fluorescence rise measured under high intensity of exciting light. Funct. Plant Biol. 2006, 33, 9–30. [Google Scholar] [CrossRef]
- Dias, A.S.; Barreiro, M.G.; Campos, P.S.; Ramalho, J.C.; Lidon, F.C. Wheat cellular membrane thermotolerance under heat stress. J. Agron. Crop Sci. 2010, 196, 100–108. [Google Scholar] [CrossRef]
- Scotti-Campos, P.; Semedo, J.N.; Pais, I.P.; Oliveira, M.; Passarinho, J.; Santos, M.; Almeida, A.S.; Costa, A.R.; Pinheiro, N.; Bagorro, C.; et al. Physiological responses to drought in four developed Triticum aestivum groups. Emir. J. Food Agric. 2015, 27, 178–185. [Google Scholar] [CrossRef]
- Shabala, S. Physiological and cellular aspects of phytotoxicity tolerance in plants: The role of membrane transporters and implications for crop breeding for waterlogging tolerance. New Phytol. 2011, 190, 289–298. [Google Scholar] [CrossRef] [Green Version]
- Blokhina, O.; Fagerstedt, K.V. Oxidative metabolism, ROS and NO under oxygen deprivation. Plant Physiol. Biochem. 2010, 48, 359–373. [Google Scholar] [CrossRef]
- Demidchik, V.; Straltsova, D.; Medvedev, S.S.; Pozhvanov, G.A.; Sokolik, A.; Yurin, V. Stress-induced electrolyte leakage: The role of K+-permeable channels and involvement in programmed cell death and metabolic adjustment. J. Exp. Bot. 2014, 65, 1259–1270. [Google Scholar] [CrossRef]
- Wang, X. Lipid signaling. Curr. Opin. Plant Biol. 2004, 7, 329–336. [Google Scholar] [CrossRef]
- Xiao, S.; Chye, M.L. New roles for acyl-CoA-binding proteins (ACBPs) in plant development, stress responses and lipid metabolism. Prog. Lipid Res. 2011, 50, 141–151. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.J.; Zhou, Y.; Chen, Q.F.; Xiao, S. New insights into the role of lipids in plant hypoxia responses. Prog. Lipid Res. 2021, 81, 101072. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Pan, R.; Zhou, M.; Xu, Y.; Zhang, W. Lipid remodelling plays an important role in wheat (Triticum aestivum) hypoxia stress. Funct. Plant Biol. 2019, 47, 58–66. [Google Scholar] [CrossRef] [PubMed]
- Baxter, A.; Mittler, R.; Suzuki, N. ROS as key players in plant stress signalling. J. Exp. Bot. 2014, 65, 1229–1240. [Google Scholar] [CrossRef]
- Ayala, A.; Muñoz, M.F.; Argüelles, S. Lipid peroxidation: Production, metabolism, and signaling mechanisms of malondialdehyde and 4-hydroxy-2-nonenal. Oxid. Med. Cell. Longev. 2014, 2014, 360438. [Google Scholar] [CrossRef] [Green Version]
- Duhan, S.; Kumari, A.; Lal, M.; Sheokand, S. Oxidative stress and antoxidant defense under combined waterlogging and salinity stresses. In Reactive Oxygen, Nitrogen and Sulfur Species in Plants: Production, Metabolism, Signaling and Defence Mechanisms; Hasanuzzaman, M., Fotopoulos, V., Nahar, K., Fujita, M., Eds.; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2019; pp. 113–142. [Google Scholar]
- Manik, S.M.N.; Pengilley, G.; Dean, G.; Field, B.; Shabala, S.; Zhou, M. Soil and crop management practices to minimize the impact of waterlogging on crop productivity. Front. Plant Sci. 2019, 10, 1–23. [Google Scholar] [CrossRef]
- Nikolaeva, M.K.; Maevskaya, S.N.; Shugaev, A.G.; Bukhov, N.G. Effect of drought on chlorophyll content and antioxidant enzyme activities in leaves of three wheat cultivars varying in productivity. Russ. J. Plant Physiol. 2010, 57, 87–95. [Google Scholar] [CrossRef]
- Shabala, S.; Shabala, L.; Barcelo, J.; Poschenrieder, C. Membrane transporters mediating root signalling and adaptive responses to oxygen deprivation and soil flooding. Plant Cell Environ. 2014, 37, 2216–2233. [Google Scholar] [CrossRef]
- Pottosin, I.; Zepeda-Jazo, I.; Bose, J.; Shabala, S. An anion conductance, the essential component of the Hydroxyl-Radical-Induced ion current in plant roots. Int. J. Mol. Sci. 2018, 19, 897. [Google Scholar] [CrossRef] [Green Version]
- Luxmoore, R.J.; Fischer, R.A.; Stolzy, L.H. Flooding and soil temperature effects on wheat during grain filling. Agron. J. 1973, 65, 361–364. [Google Scholar] [CrossRef]
- Marashi, S.K.; Chinchanikar, G.S. Effect of waterlogging periods on kernel yield and yield components of wheat (Triticum aestivum L.) cultivars. Res. Crop 2010, 5, 561–567. [Google Scholar]
- Pfeiffer, W.H.; Trethowan, R.M.; van Ginkel, M.; Ortiz-Monasterio, I.; Rajaram, S. Breeding for abiotic stress tolerance in wheat. In Abiotic Stresses: Plant Resistance through Breeding and Molecular Approaches, 1st ed.; Ashraf, M., Harris, P.J.C., Eds.; Food Product Press: New York, NY, USA, 2005; pp. 401–489. [Google Scholar]
- Valério, I.P.; Carvalho, F.I.F.; Oliveira, A.C.; Machado, A.D.A.; Benin, G.; Scheeren, P.L.; Souza, V.Q.; Hartwig, I. Desenvolvimento de afilhos e componentes do rendimento em genótipos de trigo sob diferentes densidades de semeadura. Pesqui. Agropecu. Bras. 2008, 43, 319–326. [Google Scholar] [CrossRef] [Green Version]
- Condon, A.G.; Giunta, F. Yield response of restricted-tillering wheat to transient waterlogging on duplex soils. Aust. J. Agric. Res. 2003, 54, 957–967. [Google Scholar] [CrossRef]
- Sharma, D.B.; Swarup, A. Effects of short-term flooding on growth, yield and mineral composition of wheat on sodic soil under field conditions. Plant Soil 1988, 107, 137–143. [Google Scholar] [CrossRef]
- Yaduvanshi, N.P.S.; Setter, T.L.; Sharma, S.K.; Singh, K.N.; Kulshreshtha, N. Influence of waterlogging on yield of wheat (Triticum aestivum), redox potentials, and concentrations of microelements in different soils in India and Australia. Soil Res. 2012, 50, 489–499. [Google Scholar] [CrossRef]
- Wang, X.; Huang, M.; Zhou, Q.; Cai, J.; Dai, T.B.; Cao, W.X.; Jiang, D. Physiological and proteomic mechanisms of waterlogging priming improves tolerance to waterlogging stress in wheat (Triticum aestivum L.). Environ. Exp. Bot. 2016, 132, 175–182. [Google Scholar] [CrossRef]
- Wu, J.; Wang, J.; Hui, W.; Zhao, F.; Wang, P.; Su, C.; Gong, W. Physiology of plant responses to water stress and related genes: A review. Forests 2022, 13, 324. [Google Scholar] [CrossRef]
- Wei, M.; Li, X.; Yang, R.; Li, L.; Wang, Z.; Wang, X.; Sha, A. Novel insights into genetic responses for waterlogging stress in two local wheat cultivars in Yangtze river basin. Front. Genet. 2021, 12, 681680. [Google Scholar] [CrossRef]
- Borrego-Benjumea, A.; Carter, A.; Tucker, J.R.; Yao, Z.; Xu, W.; Badea, A. Genome-wide analysis of gene expression provides new insights into waterlogging responses in Barley (Hordeum vulgare L.). Plants 2020, 9, 240. [Google Scholar] [CrossRef] [Green Version]
- Tong, C.; Hill, C.B.; Zhou, G.; Zhang, X.-Q.; Jia, Y.; Li, C. Opportunities for improving waterlogging tolerance in cereal crops—Physiological traits and genetic mechanisms. Plants 2021, 10, 1560. [Google Scholar] [CrossRef]
Figure 1.
Major effects of waterlogging occurring at root level. Figure elements created using biorender.com (accessed on 15 December 2022).
Figure 1.
Major effects of waterlogging occurring at root level. Figure elements created using biorender.com (accessed on 15 December 2022).

Figure 2.
Effects of waterlogging at shoot level. Figure elements created using biorender.com (accessed on 15 December 2022).
Figure 2.
Effects of waterlogging at shoot level. Figure elements created using biorender.com (accessed on 15 December 2022).


{kind=link}
{kind=link}
Table 1.
Effects of waterlogging on the physical, electrochemical, chemical, and biological properties of the soil. Elements of the figure were created using biorender.com (accessed on 15 November 2022).
Table 1.
Effects of waterlogging on the physical, electrochemical, chemical, and biological properties of the soil. Elements of the figure were created using biorender.com (accessed on 15 November 2022).
| Impacts in Soil Properties Due to Waterlogging | ||
|---|---|---|
 | Physical | Changes in ideal solid:pore and soil:air volume ratios; Decreased [O2]; increased [CO2]; lowered diffusion coefficient for gases. |
| Electrochemical | Decreased redox potential (Eh); Changes in soil pH and EC. | |
| Chemical | Changes in solubility, mobility, and bioavailability of nutrients and potentially toxic elements. | |
| Biological | Changes in microbial activity and in the nitrogen cycle (mineralization and immobilization of organic N). | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Pais, I.P.; Moreira, R.; Semedo, J.N.; Ramalho, J.C.; Lidon, F.C.; Coutinho, J.; Maçãs, B.; Scotti-Campos, P. Wheat Crop under Waterlogging: Potential Soil and Plant Effects. Plants 2023, 12, 149. https://doi.org/10.3390/plants12010149
AMA Style
Pais IP, Moreira R, Semedo JN, Ramalho JC, Lidon FC, Coutinho J, Maçãs B, Scotti-Campos P. Wheat Crop under Waterlogging: Potential Soil and Plant Effects. Plants. 2023; 12(1):149. https://doi.org/10.3390/plants12010149
Chicago/Turabian StylePais, Isabel P., Rita Moreira, José N. Semedo, José C. Ramalho, Fernando C. Lidon, José Coutinho, Benvindo Maçãs, and Paula Scotti-Campos. 2023. "Wheat Crop under Waterlogging: Potential Soil and Plant Effects" Plants 12, no. 1: 149. https://doi.org/10.3390/plants12010149
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.