
Transcriptome Analysis Identifies Novel Genes Associated with Low-Temperature Seed Germination in Sweet Corn
 ,
,  , and
, and
Abstract
:1. Introduction
2. Results
2.1. Inbred Line L282 Showed Better Germination Resistance to Low Temperature Than L693
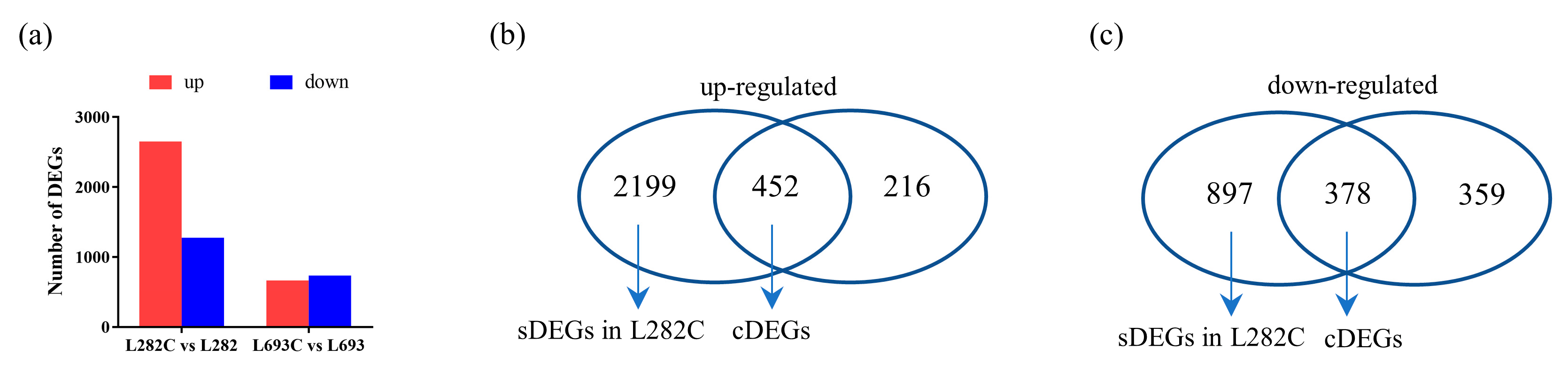
2.2. Transcriptome Analysis of L282 and L693 Seeds Germinated at Normal and Low Temperatures
2.3. Identification of Common DEGs at Low Temperature
2.4. Identification of L282 Specifically Expressed Cold-Responsive DEGs
3. Discussion
3.1. Improving the Germination Capacity of Sweet Corn Seeds at Low Temperature Is Crucial for Sweet Corn Production
3.2. Transcriptome Profiling with Respect to Sweet Corn Kernels’ Response to Low Temperature
3.3. The Genes Related to Plasma Membrane Contribute to Low-Temperature Tolerance of sh2 Sweet Corn
4. Materials and Methods
4.1. Plant Materials and Trait Evaluation
4.2. RNA Extraction and Sequencing
4.3. Sequence Data Analysis
4.4. Cluster Analysis and Functional Annotation Enrichment Analysis
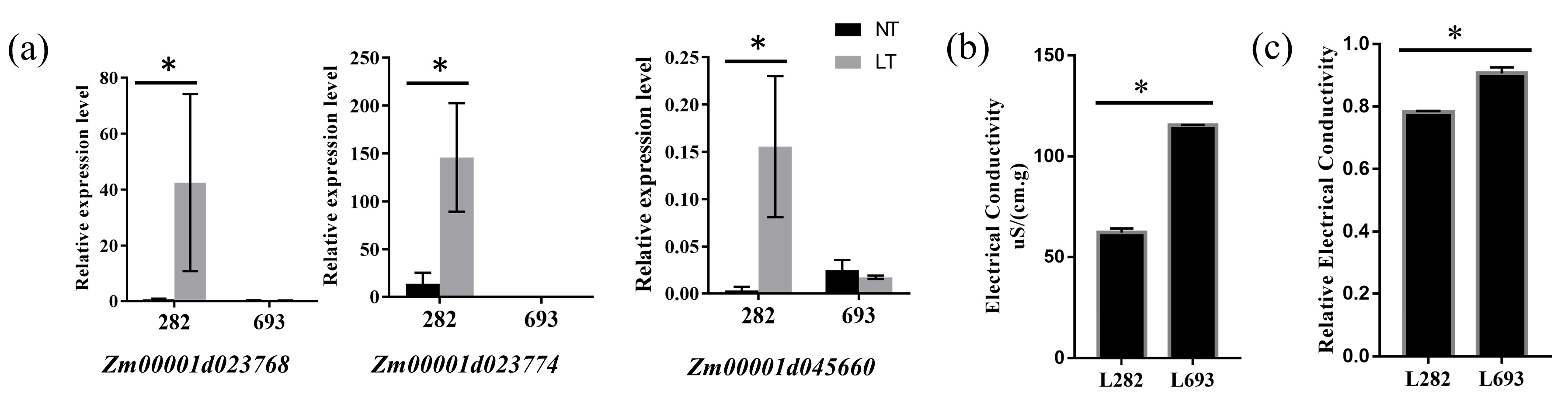
4.5. Quantitative RT-PCR Analysis
4.6. Electric Conductivity Measurement
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Revilla, P.; Anibas, C.M.; Tracy, W.F. Sweet corn research around the world 2015–2020. Agronomy 2021, 11, 534. [Google Scholar] [CrossRef]
- Hu, Y.; Colantonio, V.; Müller, B.S.F.; Leach, K.A.; Nanni, A.; Finegan, C.; Wang, B.; Baseggio, M.; Newton, C.J.; Juhl, E.M.; et al. Genome assembly and population genomic analysis provide insights into the evolution of modern sweet corn. Nat. Commun. 2021, 12, 1227. [Google Scholar] [CrossRef] [PubMed]
- Tracy, W.F.; Shuler, S.L.; Dodson-Swenson, H. The use of endosperm genes for sweet corn improvement: A review of developments in endosperm genes in sweet corn since the seminal publication in Plant Breeding Reviews, Volume 1, by Charles Boyer and Jack Shannon (1984). Plant Breed. Rev. 2019, 43, 215–241. [Google Scholar]
- The Report of the Dietary Guidelines Advisory Committee on the Dietary Guidelines for Americans, 2015, to the Secretary of Agriculture and the Secretary of Health and Human Services; USDA/HHS: Washington, DC, USA, 2015.
- Boyer, C.D.; Shannon, J.C. The use of endosperm genes for sweet corn improvement. Plant Breed. Rev. 1984, 1, 139–161. [Google Scholar]
- Hannah, L.C.; Nelson, O.E. Characterization of ADP-glucose pyrophosphorylase from shrunken-2 and brittle-2 mutants of maize. Biochem. Genet. 1976, 14, 547–560. [Google Scholar] [CrossRef] [PubMed]
- Creech, R.G. Genetic control of carbohydrate synthesis in maize endosperm. Genetics 1965, 52, 1175–1186. [Google Scholar] [CrossRef]
- Garwood, D.L.; McArdle, F.J.; Vanderslice, S.F.; Shannon, J.C. Postharvest carbohydrate transformations and processed quality of high sugar maize genotypes. J. Am. Soc. Hortic. Sci. 1976, 101, 400–404. [Google Scholar] [CrossRef]
- Carey, E.E.; Rhodes, A.M.; Dickinson, D.B. Post-harvest levels of sugars and sorbitol in sugary enhancer (su se) and sugary (su Se) maize. Hortic. Sci. 1982, 17, 241–242. [Google Scholar]
- Juvik, J.A.; Yousef, G.G.; Han, T.; Tadmor, Y.; Azanza, F.; Tracy, W.F.; Barzur, A.; Rocheford, T.R. QTL influencing kernel chemical composition and seedling stand establishment in sweet corn with the shrunken2 and sugary enhancer1 endosperm mutations. J. Am. Soc. Hortic. Sci. 2003, 128, 864–875. [Google Scholar] [CrossRef] [Green Version]
- Douglass, S.K.; Juvik, J.A.; Splittstoesser, W.E. Sweet corn seedling emergence and variation in kernel carbohydrates reserves. Seed Sci. Technol. 1993, 21, 433–445. [Google Scholar]
- Pairochteerakul, P.; Jothityangkoon, D.; Ketthaisong, D.; Simla, S.; Lertrat, K.; Suriharn, B. Seed germination in relation to total sugar and starch in endosperm mutant of sweet corn genotypes. Agronomy 2018, 8, 299. [Google Scholar] [CrossRef] [Green Version]
- Juvik, J.A.; Jangulo, M.C.; Headrick, J.M.; Pataky, J.K.; Tracy, W.F. Changes in characteristics of kernels in a population of shrunken-2 maize selected for improved field emergence and increased kernel weight. J. Am. Soc. Hortic. Sci. 1993, 118, 135–140. [Google Scholar] [CrossRef]
- Lertrat, K.; Pulam, T. Breeding for increased sweetness in sweet corn. Int. J. Plant Breed. 2007, 1, 27–30. [Google Scholar]
- Gong, F.; Yang, L.; Tai, F.; Hu, X.; Wang, W. “Omics” of maize stress response for sustainable food production: Opportunities and challenges. OMICS 2014, 18, 714–732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, Y.L.; Shi, Y.T.; Yang, S.H. Advances and challenges in uncovering cold tolerance regulatory mechanisms in plants. New Phytol. 2019, 222, 1690–1704. [Google Scholar] [CrossRef] [PubMed]
- Kong, X.P.; Pan, J.W.; Zhang, M.Y.; Xing, X.; Zhou, Y.; Liu, Y.; Li, D.P.; Li, D.Q. ZmMKK4, a novel group C mitogen-activated protein kinase kinase in maize (Zea mays), confers salt and cold tolerance in transgenic Arabidopsis. Plant Cell Environ. 2011, 34, 1291–1303. [Google Scholar] [CrossRef]
- Jiang, H.F.; Shi, Y.T.; Liu, J.Y.; Li, Z.; Fu, D.Y.; Wu, S.F.; Li, M.Z.; Yang, Z.J.; Shi, Y.L.; Lai, J.S.; et al. Natural polymorphism of ZmICE1 contributes to amino acid metabolism that impacts cold tolerance in maize. Nat. Plants 2022, 8, 1176–1190. [Google Scholar] [CrossRef]
- Li, Z.Y.; Fu, D.Y.; Wang, X.; Zeng, R.; Zhang, X.; Tian, J.G.; Zhang, S.S.; Yang, X.H.; Tian, F.; Lai, J.S.; et al. The transcription factor bZIP68 negatively regulates cold tolerance in maize. Plant Cell 2022, 34, 2833–2851. [Google Scholar] [CrossRef]
- Zeng, R.; Li, Z.Y.; Shi, Y.T.; Fu, D.Y.; Pan, L.; Cheng, J.K.; Jiang, C.F.; Yang, S.H. Natural variation in a type—A response regulator confers maize chilling tolerance. Nat. Commun. 2021, 12, 4713. [Google Scholar] [CrossRef]
- Li, X.H.; Hu, H.R.; Hu, X.M.; Wang, G.H.; Du, X.M.; Li, L.; Wang, F.; Fu, J.J.; Wang, G.H.; Wang, J.H.; et al. Transcriptome analysis of near-isogenic lines provides novel insights into genes associated with seed low-temperature germination ability in Maize (Zea mays L.). Plants 2022, 11, 887. [Google Scholar] [CrossRef]
- Mao, J.H.; Yu, Y.T.; Yang, J.; Li, G.K.; Li, C.Y.; Qi, X.T.; Wen, T.X.; Hu, J.G. Comparative transcriptome analysis of sweet corn seedlings under low-temperature stress. Crop J. 2017, 5, 396–406. [Google Scholar] [CrossRef]
- Steponkus, P.L. Role of the plasma membrane in freezing injury and cold acclimation. Annu. Rev. Plant Physiol. 1984, 35, 543–584. [Google Scholar] [CrossRef]
- Gu, R.L.; Huang, R.; Jia, G.Y.; Yuan, Z.P.; Ren, L.S.; Li, L.; Wang, J.H. Effect of mechanical threshing on damage and vigor of maize seed threshed at different moisture contents. J. Integr. Agric. 2019, 18, 1571–1578. [Google Scholar] [CrossRef]
- Rajjou, L.; Duval, M.; Gallardo, K.; Catusse, J.; Bally, J.; Job, C.; Job, D. Seed germination and vigor. Annu. Rev. Plant Biol. 2012, 63, 507–533. [Google Scholar] [CrossRef] [Green Version]
- Finch-Savage, W.E.; Bassel, G.W. Seed vigor and crop establishment: Extending performance beyond adaptation. J. Exp. Bot. 2016, 67, 567–591. [Google Scholar] [CrossRef]
- Kuş, E. Evaluation of some operational parameters of a vacuum single-seed planter in maize sowing. J. Agric. Sci. 2021, 27, 327–334. [Google Scholar]
- Revilla, P.; Malvar, R.A.; Cartea, M.E.; Butrón, A.; Ordás, A. Inheritance of cold tolerance at emergence and during early season growth in maize. Crop Sci. 2000, 40, 1579–1585. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Pan, S.S.; Guo, G.Y.; Gu, Q.Q.; Pan, R.H.; Guan, Y.J.; Hu, J. Preparation of a thermoresponsive maize seed coating agent using polymer hydrogel for chilling resistance and anti-counterfeiting. Prog. Org. Coat. 2020, 139, 105452. [Google Scholar] [CrossRef]
- Douds, D.D.; Wilson, D.O.; Seidel, R.; Ziegler-Ulsh, C. A method to minimize the time needed for formation of mycorrhizas in sweet corn seedlings for outplanting using AM fungus inoculum. produced on-farm. Sci. Hortic. 2016, 203, 62–68. [Google Scholar] [CrossRef]
- Fowler, S.; Thomashow, M.F. Arabidopsis transcriptome profiling indicates that multiple regulatory pathways are activated during cold acclimation in addition to the CBF cold response pathway. Plant Cell 2002, 14, 1675–1690. [Google Scholar] [CrossRef] [Green Version]
- Ma, Q.; Dai, X.; Xu, Y.; Guo, J.; Liu, Y.; Chen, N.; Xiao, J.; Zhang, D.; Xu, Z.; Zhang, X.; et al. Enhanced tolerance to chilling stress in OsMYB3R-2 transgenic rice is mediated by alteration in cell cycle and ectopic expression of stress genes. Plant Physiol. 2009, 150, 244–256. [Google Scholar] [CrossRef] [PubMed]
- Die, J.V.; Arora, R.; Rowland, L.J. Global patterns of protein abundance during the development of cold hardiness in Blueberry. Environ. Exp. Bot. 2016, 124, 11–21. [Google Scholar] [CrossRef]
- Li, H.; Yue, H.; Xie, J.; Bu, J.; Li, L.; Xin, X.; Zhao, Y.; Zhang, H.; Yang, L.; Wang, J.; et al. Transcriptomic profiling of the high-vigour maize (Zea mays L.) hybrid variety response to cold and drought stresses during seed germination. Sci. Rep. 2021, 11, 19345. [Google Scholar] [CrossRef] [PubMed]
- Maxfield, F.R. Plasma membrane microdomains. Curr. Opin. Cell Biol. 2002, 14, 483–487. [Google Scholar] [CrossRef] [PubMed]
- de la Serna, J.B.; Schütz, G.J.; Eggeling, C.; Cebecauer, M. There is no simple model of the plasma membrane organization. Front. Cell Dev. Biol. 2016, 4, 106. [Google Scholar]
- Grecco, H.E.; Schmick, M.; Bastiaens, P.I.H. Signaling from the living plasma membrane. Cell 2011, 144, 897–909. [Google Scholar] [CrossRef] [Green Version]
- Uemura, M.; Tominaga, Y.; Nakagawara, C.; Shigematsu, S.; Minami, A.; Kawamura, Y. Responses of the plasma membrane to low temperatures. Physiol. Plant. 2006, 126, 81–89. [Google Scholar] [CrossRef]
- Kawamura, Y.; Uemura, M. Mass spectrometric approach for identifying putative plasma membrane proteins of Arabidopsis leaves associated with cold acclimation. Plant J. 2003, 36, 141–154. [Google Scholar] [CrossRef]
- Minami, A.; Fujiwara, M.; Furuto, A.; Fukao, Y.; Yamashita, T.; Kamo, M.; Kawamura, Y.; Uemura, M. Alterations in detergent-resistant plasma membrane microdomains in Arabidopsis thaliana during cold acclimation. Plant Cell Physiol. 2009, 50, 341–359. [Google Scholar] [CrossRef] [Green Version]
- Rahman, A.; Kawamura, Y.; Maeshima, M.; Rahman, A.; Uemura, M. Plasma membrane aquaporin members PIPs act in concert to regulate cold acclimation and freezing tolerance responses in Arabidopsis thaliana. Plant Cell Physiol. 2020, 61, 787–802. [Google Scholar] [CrossRef]
- Hirano, S.; Sasaki, K.; Osaki, Y.; Tahara, K.; Takahashi, H.; Takemiya, A.; Kodama, Y. The localization of phototropin to the plasma membrane defines a cold-sensing compartment in Marchantia polymorpha. PNAS Nexus 2022, 1, pgac030. [Google Scholar] [CrossRef]
- Guo, X.; Xu, H.; Chong, K. Cold signal shuttles from membrane to nucleus. Mol. Cell 2017, 66, 7–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eom, J.; Chen, L.; Sosso, D.; Julius, B.T.; Lin, I.W.; Qu, X.; Braun, D.; Frommer, W. SWEETs, transporters for intracellular and intercellular sugar translocation. Curr. Opin. Plant Biol. 2015, 25, 53–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Julius, B.; Leach, K.; Tran, T.; Mertz, R.; Braun, D. Sugar transporters in plants: New insights and discoveries. Plant Cell Physiol. 2017, 58, 1442–1460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phukan, U.; Jeena, G.S.; Tripathi, V.; Shukla, R.K. MaRAP2-4, a waterlogging-responsive ERF from Mentha, regulates bidirectional sugar transporter AtSWEET10 to modulate stress response in Arabidopsis. Plant Biotechnol. J. 2018, 16, 221–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, J.; Sun, M.; Ma, Q.; Kang, H.; Liu, Y.; Hao, Y.; You, C. MdSWEET17, a sugar transporter in apple, enhances drought tolerance in tomato. J. Integr. Agric. 2019, 18, 2041–2051. [Google Scholar] [CrossRef]
- Wang, L.; Yao, L.; Hao, X.; Li, N.; Qian, W.; Yue, C.; Ding, C.; Zeng, J.; Yang, Y.; Wang, X. Tea plant SWEET transporters: Expression profiling, sugar transport, and the involvement of CsSWEET16 in Modifying Cold Tolerance in Arabidopsis. Plant Mol. Biol. 2018, 96, 577–592. [Google Scholar] [CrossRef]
- Klemens, P.A.W.; Patzke, K.; Deitmer, J.; Spinner, L.; Le Hir, R.; Bellini, C.; Bedu, M.; Chardon, F.; Krapp, A.; Neuhaus, H.E. Overexpression of the vacuolar sugar Carrier AtSWEET16 modifies germination, growth, and stress tolerance in Arabidopsis. Plant Physiol. 2013, 163, 1338–1352. [Google Scholar] [CrossRef] [Green Version]
- Huang, D.; Chen, Y.; Liu, X.; Ni, D.; Bai, L.; Qin, Q. Genome-wide identification and expression analysis of the SWEET gene family in daylily (Hemerocallis fulva) and functional analysis of HfSWEET17 in response to cold stress. BMC Plant Biol. 2022, 22, 211. [Google Scholar] [CrossRef]
- Nagalakshmi, U.; Waern, K.; Snyder, M. RNA-Seq: A method for comprehensive transcriptome analysis. Curr. Protoc. Mol. Biol. 2010, 89, 4–11. [Google Scholar] [CrossRef]
- Trapnell, C.; Pachter, L.; Salzberg, S.L. TopHat: Discovering splice junctions with RNA-Seq. Bioinformatics 2009, 25, 1105–1111. [Google Scholar] [CrossRef] [PubMed]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—A Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Love, M.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2012. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Name | Line | Germination Condition | Rep | Total Reads | Rate of Total Mapped Reads (%) | Rate of Uniquely Mapped Reads (%) | Num. of Expressed Genes | Rate of Expressed Genes (%) |
|---|---|---|---|---|---|---|---|---|
| L282 | L282 | Normal | 1 | 22,051,805 | 88.19% | 77.64% | 20,346 | 51.39% |
| 2 | 37,564,993 | 88.85% | 78.52% | 19,998 | 50.51% | |||
| L693 | L693 | Normal | 1 | 23,130,884 | 88.83% | 78.75% | 20,143 | 50.88% |
| 2 | 23,854,290 | 90.11% | 80.55% | 20,778 | 52.48% | |||
| L282C | L282 | Cold | 1 | 22,230,484 | 89.10% | 80.10% | 21,045 | 53.16% |
| 2 | 21,280,625 | 88.62% | 78.72% | 21,067 | 53.21% | |||
| L693C | L693 | Cold | 1 | 22,958,482 | 89.93% | 73.93% | 21,161 | 53.45% |
| 2 | 35,891,174 | 89.21% | 79.68% | 21,219 | 53.60% |
| Gene | log2|FC| in L282C | FDR | log2|FC| in L693C | FDR | Annotation |
|---|---|---|---|---|---|
| Zm00001d025943 | 3.80 | 0.03 | NA | NA | fructofuranosidases |
| Zm00001d014866 | 1.27 | 0.03 | NA | NA | glycosyl transferase |
| Zm00001d031303 | 1.22 | 0.03 | NA | NA | raffinose synthases |
| Zm00001d037480 | 1.95 | 0.01 | NA | NA | raffinose synthases |
| Zm00001d017502 | 2.78 | 3.00 × 10−04 | NA | NA | trehalose 6-phosphate phosphatase |
| Zm00001d029371 | 1.06 | 0.01 | NA | NA | trehalose 6-phosphate phosphatase |
| ZmSWEET4c | 3.10 | 3.25 × 10−05 | NA | NA | sugar export transporter |
| ZmSWEET1a | 1.07 | 7.49 × 10−06 | NA | NA | sugar export transporter |
| ZmSWEET4b | 1.75 | 1.73 × 10−10 | NA | NA | sugar export transporter |
| ZmSWEET13a | 1.39 | 2.16 × 10−05 | NA | NA | sugar export transporter |
| ZmSWEET13b | 1.22 | 2.79 × 10−04 | NA | NA | sugar export transporter |
| ZmSWEET14a | 1.50 | 1.56 × 10−06 | NA | NA | sugar export transporter |
| ZmSWEET14b | 1.83 | 6.21 × 10−05 | NA | NA | sugar export transporter |
| ZmSWEET17b | 3.44 | 3.20 × 10−05 | NA | NA | sugar export transporter |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiao, Y.; Chen, M.; Zheng, N.; Xu, Z.; Zhang, J.; Hu, X.; Li, L.; Gu, R.; Du, X.; Wang, J. Transcriptome Analysis Identifies Novel Genes Associated with Low-Temperature Seed Germination in Sweet Corn. Plants 2023, 12, 159. https://doi.org/10.3390/plants12010159
Xiao Y, Chen M, Zheng N, Xu Z, Zhang J, Hu X, Li L, Gu R, Du X, Wang J. Transcriptome Analysis Identifies Novel Genes Associated with Low-Temperature Seed Germination in Sweet Corn. Plants. 2023; 12(1):159. https://doi.org/10.3390/plants12010159
Chicago/Turabian StyleXiao, Yingni, Mei Chen, Nannan Zheng, Zhuoyi Xu, Jie Zhang, Xinmin Hu, Li Li, Riliang Gu, Xuemei Du, and Jianhua Wang. 2023. "Transcriptome Analysis Identifies Novel Genes Associated with Low-Temperature Seed Germination in Sweet Corn" Plants 12, no. 1: 159. https://doi.org/10.3390/plants12010159