
Pedoclimatic Conditions Influence the Morphological, Phytochemical and Biological Features of Mentha pulegium L.
 , , and
, , and 
Abstract
:1. Introduction
2. Results
2.1. Micromorphological Analyses
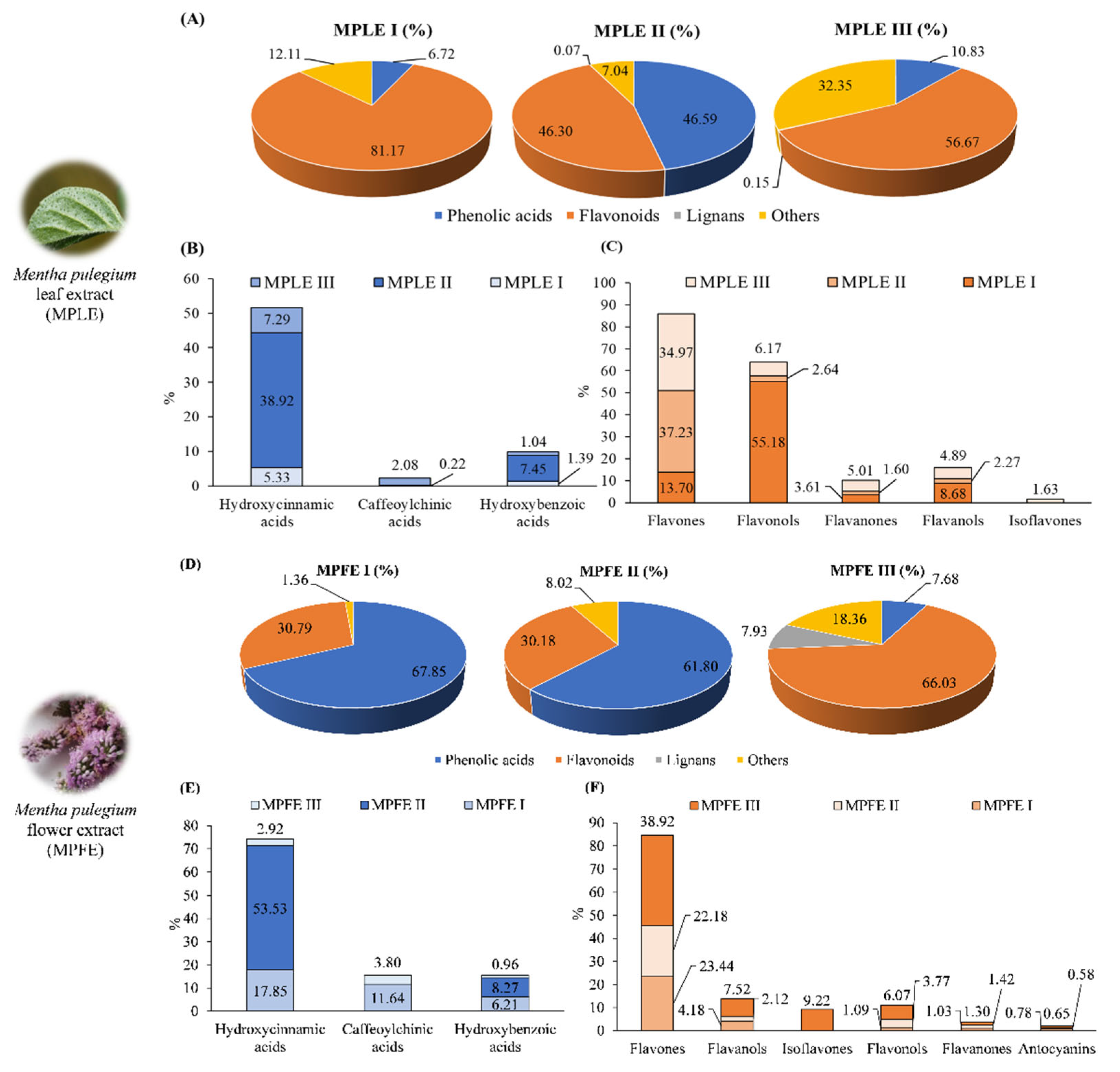
2.2. Phytochemical Analyses
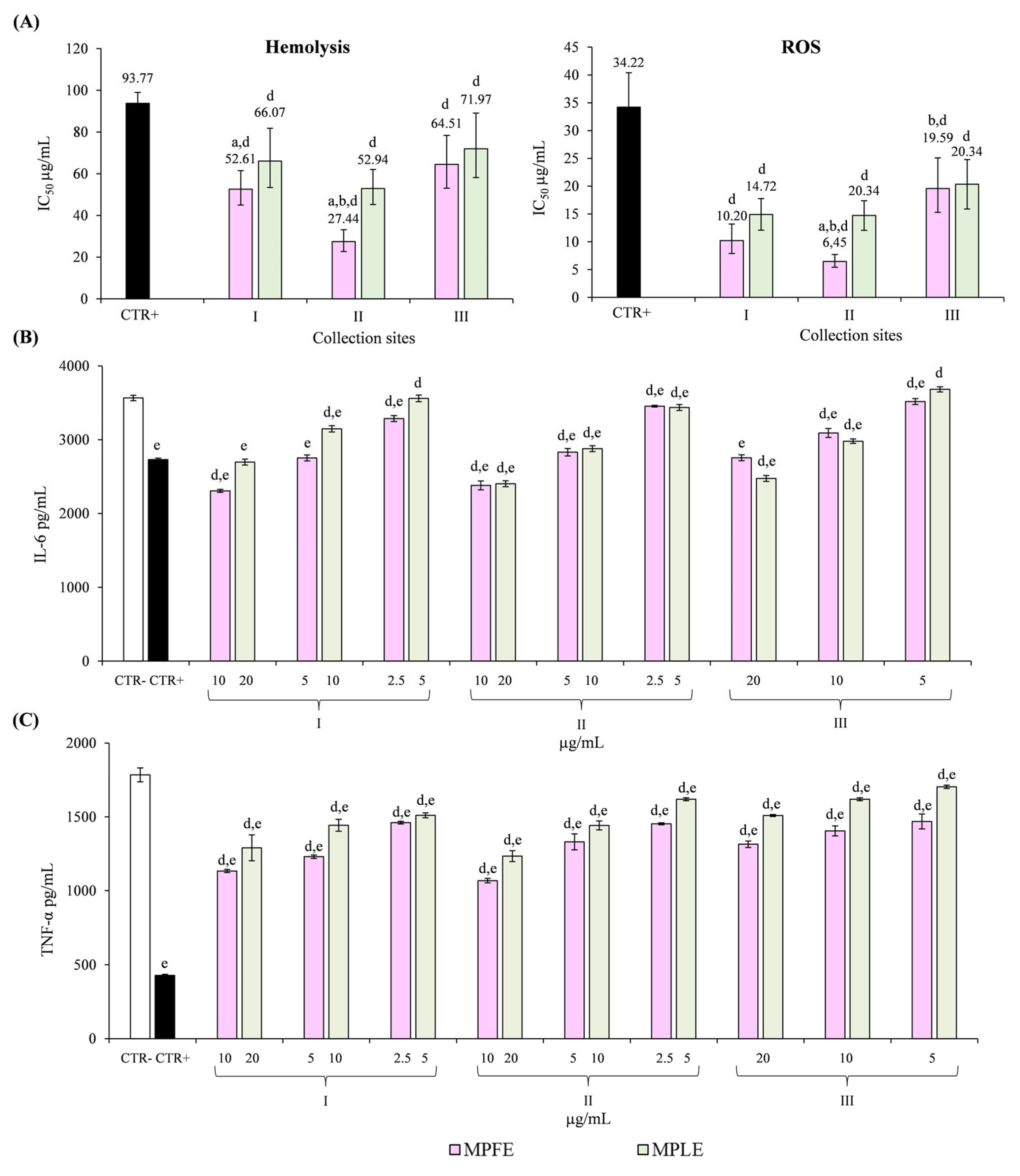
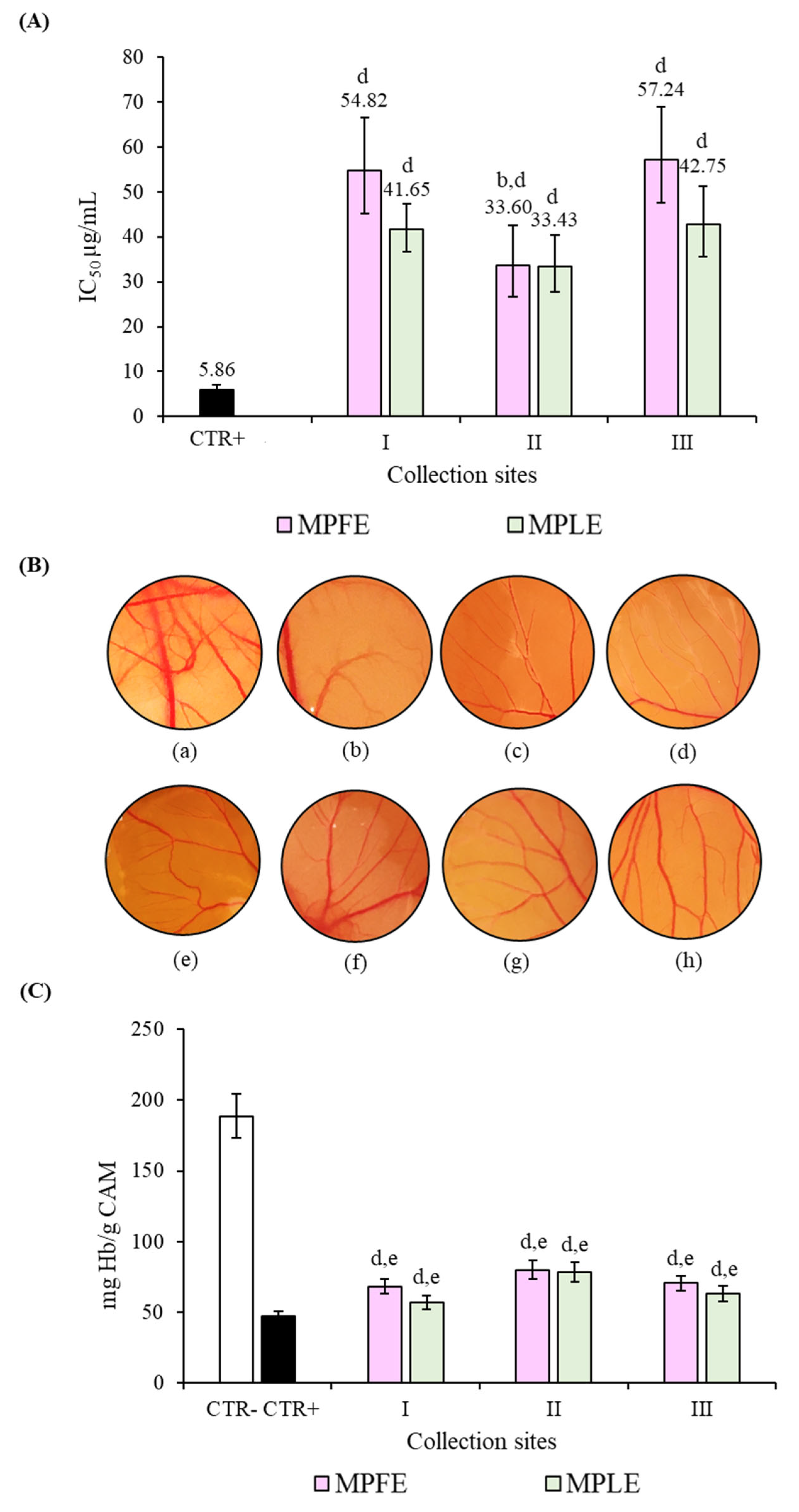
2.3. Biological Properties
3. Discussion
4. Materials and Methods
4.1. Plant Collection
4.2. Chemicals
4.3. Micromorphological Analyses
4.3.1. Light Microscopy
4.3.2. Scanning Electron Microscopy
4.4. Preparation of Hydroalcoholic Extracts
4.5. Phytochemical Screening
4.5.1. Total Phenolics
4.5.2. Total Flavonoids
4.5.3. Flavanols
4.6. Phytochemical Characterization by LC-DAD-ESI-MS Analysis
4.7. In Vitro Cell-Free Assays for Determination of Antioxidant and Anti-Inflammatory Properties
4.7.1. DPPH Assay
4.7.2. FRAP Assay
4.7.3. TEAC Assay
4.7.4. ORAC Assay
4.7.5. Bovine Serum Albumin (BSA) Denaturation Assay
4.7.6. Protease Inhibition Assay
4.8. In Vitro Cell-Based Assays for Determination of Antioxidant and Anti-Inflammatory Properties
4.8.1. Heat-Induced Hemolysis Assay
4.8.2. Scavenging Activity against Intracellular ROS
4.8.3. Evaluation of Anti-Inflammatory Activity on Peripheral Blood Mononuclear Cell
4.9. Chick Chorioallantoic Membrane (CAM) Assay for Determination of Antiangiogenic Properties
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tafrihi, M.; Imran, M.; Tufail, T.; Gondal, T.A.; Caruso, G.; Sharma, S.; Sharma, R.; Atanassova, M.; Atanassov, L.; Valere Tsouh Fokou, P.; et al. The wonderful activities of the genus Mentha: Not only antioxidant properties. Molecules 2021, 26, 1118. [Google Scholar] [CrossRef] [PubMed]
- Salehi, B.; Stojanović-Radić, Z.; Matejić, J.; Sharopov, F.; Antolak, H.; Kręgiel, D.; Sen, S.; Sharifi-Rad, M.; Acharya, K.; Sharifi-Rad, R.; et al. Plants of genus Mentha: From farm to food factory. Plants 2018, 7, 70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gulluce, M.F.; Shain, M.; Sokmen, H.; Ozer, D.; Daferera, A.; Sokmen, M.; Polissiou, M.; Adiguzel, A.; Ozkan, H. Antimicrobial and antioxidant properties of the essential oils and methanol extract from Mentha longifolia L. spp. longifolia. Food Chem. 2007, 103, 1449–1456. [Google Scholar] [CrossRef]
- Anwar, F.; Abbas, A.; Mehmood, T.; Gilani, A.H.; Rehman, N.U. Mentha: A genus rich in vital nutra-pharmaceuticals—A review. Phytother. Res. 2019, 33, 2548–2570. [Google Scholar] [CrossRef] [PubMed]
- Rahimifard, N.; Hajimehdipoor, H.; Hedayati, M.; Bagheri, O.; Pishehvar, H.; Ajani, Y. Cytotoxic effects of essential oils and extracts of some Mentha species on Vero, Hela and Hep2 cell lines. J. Med. Plants 2010, 9, 88–92. [Google Scholar]
- Patti, F.; Palmioli, A.; Vitalini, S.; Bertazza, L.; Redaelli, M.; Zorzan, M.; Rubin, B.; Mian, C.; Bertolini, C.; Iacobone, M.; et al. Anticancer effects of wild mountain Mentha longifolia extract in adrenocortical tumor cell models. Front. Pharmacol. 2020, 10, 1647. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, L.; Póvoa, O.; Teixeira, G.; Figueiredo, A.C.; Moldão, M.; Monteiro, A. Trichomes micromorphology and essential oil variation at different developmental stages of cultivated and wild growing Mentha pulegium L. populations from Portugal. Ind. Crops Prod. 2013, 43, 692–700. [Google Scholar] [CrossRef]
- Kumar, P.; Mishra, S.; Malik, A.; Satya, S. Insecticidal properties of Mentha species: A review. Ind. Crops Prod. 2011, 34, 802–817. [Google Scholar] [CrossRef]
- Nickavar, B.; Jabbareh, F. Analysis of the essential oil from Mentha pulegium and identification of its antioxidant constituents. J. Essent. Oil Bear. Plants 2018, 21, 223–229. [Google Scholar] [CrossRef]
- El-Gazar, A.A.; Emad, A.M.; Ragab, G.M.; Rasheed, D.M. Mentha pulegium L. (Pennyroyal, Lamiaceae) extracts impose abortion or fetal-mediated toxicity in pregnant rats; evidenced by the modulation of pregnancy hormones, MiR-520, MiR-146a, TIMP-1 and MMP-9 protein expressions, inflammatory state, certain related signaling pathways, and metabolite profiling via UPLC-ESI-TOF-MS. Toxins 2022, 14, 347. [Google Scholar]
- Edris, A.E. Pharmaceutical and therapeutic potentials of essential oils and their individual volatile constituents: A review. Phytother. Res. 2007, 21, 308–323. [Google Scholar] [CrossRef]
- Al-Rajhi, A.M.H.; Qanash, H.; Almuhayawi, M.S.; Al Jaouni, S.K.; Bakri, M.M.; Ganash, M.; Salama, H.M.; Selim, S.; Abdelghany, T.M. Molecular interaction studies and phytochemical characterization of Mentha pulegium L. constituents with multiple biological utilities as antioxidant, antimicrobial, anticancer and anti-hemolytic agents. Molecules 2022, 27, 4824. [Google Scholar] [CrossRef]
- Parsania, M.; Rezaee, M.B.; Monavari, S.H.; Jaimand, K.; Mousavi-Jazayeri, S.M.; Razazian, M.; Nadjarha, M.H. Antiviral screening of four plant extracts against acyclovir resistant herpes simplex virus type-1. Pak. J. Pharm. Sci. 2017, 30, 1407–1411. [Google Scholar]
- Mahboubi, M.; Haghi, G. Antimicrobial activity and chemical composition of Mentha pulegium L. essential oil. J. Ethnopharmacol. 2008, 119, 325–327. [Google Scholar] [CrossRef]
- Barchan, A.; Bakkali, M.; Arakrak, A.; Laglaoui, A. Effet antibactérien et anti-biofilm de trois espèces de Mentha: Mentha spicata, Mentha pulegium et Mentha piperita. Phytothérapie 2016, 14, 88–96. [Google Scholar] [CrossRef]
- Gruenwald, J.; Brendler, T.; Jaenicke, C. PDR for Herbal Medicines; Medical Economic Company Inc.: Montvale, NJ, USA, 2000. [Google Scholar]
- Teixeira, B.; Marques, A.; Ramos, C.; Batista, I.; Serrano, C.; Matos, O.; Neng, N.R.; Nogueira, J.M.; Saraiva, J.A.; Nunes, M.L. European pennyroyal (Mentha pulegium) from Portugal: Chemical composition of essential oil and antioxidant and antimicrobial properties of extracts and essential oil. Ind. Crops Prod. 2012, 36, 81–87. [Google Scholar] [CrossRef]
- LiverTox: Clinical and Research Information on Drug-Induced Liver Injury. Bethesda (MD): National Institute of Diabetes and Digestive and Kidney Diseases; Pennyroyal Oil. 2012. Available online: https://www.ncbi.nlm.nih.gov/books/NBK548673/ (accessed on 14 November 2022).
- Caputo, L.; Cornara, L.; Raimondo, F.M.; De Feo, V.; Vanin, S.; Denaro, M.; Trombetta, D.; Smeriglio, A. Mentha pulegium L.: A plant underestimated for its toxicity to be recovered from the perspective of the circular economy. Molecules 2021, 26, 2154. [Google Scholar] [CrossRef]
- Gordon, P.; Khojasteh, S.C. A decades-long investigation of acute metabolism-based hepatotoxicity by herbal constituents: A case study of pennyroyal oil. Drug Metab. Rev. 2014, 47, 12–20. [Google Scholar] [CrossRef]
- Taamalli, A.; Arráez-Román, D.; Abaza, L.; Iswaldi, I.; Fernandez-Gutierrez, A.; Zarrouk, M.; SeguraCarretero, A. LC-MS-based metabolite profiling of methanolic extracts from the medicinal and aromatic species Mentha pulegium and Origanum majorana. Phytochem. Anal. 2015, 26, 320–330. [Google Scholar] [CrossRef]
- Bota, V.B.; Neamtu, A.-A.; Olah, N.-K.; Chișe, E.; Burtescu, R.F.; Pripon Furtuna, F.R.; Nicula, A.-S.; Neamtu, C.; Maghiar, A.-M.; Ivănescu, L.-C.; et al. A comparative analysis of the anatomy, phenolic profile, and antioxidant capacity of Tussilago farfara L. Vegetative Organs. Plants 2022, 11, 1663. [Google Scholar] [CrossRef]
- Brahmi, F.; Lounis, N.; Mebarakou, S.; Guendouze, N.; Yalaoui-Guellal, D.; Madani, K.; Boulekbache-Makhlouf, L.; Duez, P. Impact of Growth sites on the phenolic contents and antioxidant activities of three Algerian Mentha Species (M. pulegium L., M. rotundifolia (L.) Huds., and M. spicata L.). Front. Pharmacol. 2022, 13, 886337. [Google Scholar] [CrossRef] [PubMed]
- Appel, H.M.; Governor, H.L.; D’ascenzo, M.; Siska, E.; Schultz, J.C. Limitations of Folin assays of foliar phenolics in ecological studies. J. Chem. Ecol. 2001, 27, 761–778. [Google Scholar] [CrossRef] [PubMed]
- Sànchez-Rangel, J.C.; Benavides, J.; Heredia, J.B.; Cisneros-Zevallos, L.; Jacobo-Velázquez, D.A. The Folin-Ciocalteu assay revisited: Improvement of its specificity for total phenolic content determination. Anal. Methods 2013, 5, 5990–5999. [Google Scholar] [CrossRef]
- Martins, G.R.; Monteiro, A.F.; do Amaral, F.R.L.; da Silva, A.S. A validated Folin-Ciocalteu method for total phenolics quantification of condensed tannin-rich açaí (Euterpe oleracea Mart.) seeds extract. J. Food Sci. Technol. 2021, 58, 4693–4702. [Google Scholar] [CrossRef] [PubMed]
- Metcalfe, C.R.; Chalk, L. Anatomy of the Dicotyledons: Wood Structure and Conclusion of the General Introduction, 2nd ed.; Clarendon Press: Oxford, NY, USA, 1983; Volume 2, pp. 82–97. [Google Scholar]
- Iwashina, T.; Ootani, S.; Hayashi, K. Determination of minor flavonol-glycosides and suger free flavonols in the tepals of several species of Cereoicideae (Cactaceae). Bot. Mag. 1986, 99, 53–62. [Google Scholar] [CrossRef]
- Iwashina, T.; Ootani, S.; Hayashi, K. On the pigmentation spherical bodies and crystals in tepals of Cactaceous species in reference to the nature of betalains or flavonols. Bot. Mag. 1988, 101, 175–184. [Google Scholar] [CrossRef]
- Inoue, T.; Yoshinaga, A.; Takabe, K.; Yoshioka, T.; Ogawa, K.; Sakamoto, M.; Azuma, J.; Honda, Y. In situ detection and identification of hesperidin crystals in satsuma mandarin (Citrus unshiu) peel cells. Phytochem. Anal. 2015, 26, 105–110. [Google Scholar] [CrossRef] [Green Version]
- Smeriglio, A.; Cornara, L.; Denaro, M.; Barreca, D.; Burlando, B.; Xiao, J.; Trombetta, D. Antioxidant and cytoprotective activities of an ancient Mediterranean citrus (Citrus lumia Risso) albedo extract: Microscopic observations and polyphenol characterization. Food Chem. 2019, 279, 347–355. [Google Scholar] [CrossRef]
- Cornara, L.; D’Arrigo, C.; Pioli, F.; Borghesi, B.; Bottino, C.; Patrone, E.; Mariotti, M.G. Micromorphological investigation on the leaves of the rock samphire (Crithmum maritimum L.): Occurrence of hesperidin and diosmin crystals. Plant Biosyst. 2009, 143, 283–292. [Google Scholar] [CrossRef]
- Ramelet, A.A. Clinical benefits of daflon 500 mg in the most severe stages of chronic venous insufficiency. Angiology 2001, 52, S49–S56. [Google Scholar] [CrossRef]
- Farnsworth, N.R.; Cordell, G.A. A review of some biologically active compounds isolated from plants as reported in the 1974–1975 literature. Lloydia 1976, 39, 420–455. [Google Scholar]
- El Shafae, A.M.; El Domiaty, M.M. Improved LC methods for the determination of diosmin and/or hesperidin in plant extracts and pharmaceutical formulations. J. Pharm. Biomed. Anal. 2001, 26, 539–545. [Google Scholar] [CrossRef]
- Chevallier, A. Encyclopedia of Herbal Medicine: 550 Herbs and Remedies for Common Ailments, third edition; Dorling Kindersley Ltd.: New York, NY, USA, 2016. [Google Scholar]
- Solereder, H. Systematische Anatomie der Dicotyledonen; Verlag Von Ferdinand Enke: Stuttgart, Germany, 1908; Volume II. [Google Scholar]
- Metcalfe, C.R.; Chalk, L. Anatomy of the Dicotyledons: Leaves, Stem, and Wood in Relation to Taxonomy with Notes on Economic Uses; Clarendon Press: Oxford, NY, USA, 1950. [Google Scholar]
- Hegnauer, R. Hydrophyllaceae. In Chemotaxonomie der Pflanzen; Springer: Basel, Switzerland, 1966; Volume 19, pp. 271–274. [Google Scholar]
- Kar, A. Pharmacognosy and Pharmacobiotechnology; New Age International P. Ltd.: New Delhi, India, 2003. [Google Scholar]
- Bonzani, N.E.; Costaguta, M.; Barboza, G.E. Estudios anatómicos en especies de Mentha (Fam. Lamiaceae) de Argentina. Arnaldoa 2007, 14, 77–96. [Google Scholar]
- Bonzani, N.E. Estudios farmacobotánicos, fitoquímicos y farmacológicos en especies de Mentha de Argentina. In Tesis de Magister en Ciencias Químicas; Universidad Nacional de Córdoba: Córdoba, Argentina, 2004; p. 185. [Google Scholar]
- Taarit, M.B.; Msaada, K.; Hosni, K.; Hammami, M.; Kchouk, M.E.; Marzouk, B. Plant growth, essential oil yield and composition of sage (Salvia officinalis L.) fruits cultivated under salt stress conditions. Ind. Crops Prod. 2009, 30, 333–337. [Google Scholar] [CrossRef]
- Valifard, M.; Mohsenzadeh, S.; Kholdebarin, B.; Rowshan, V. Effects of salt stress on volatile compounds, total phenolic content, and antioxidant activities of Salvia mirzayanii. S. Afr. J. Bot. 2014, 93, 92–97. [Google Scholar] [CrossRef]
- Zrig, A.; Tounekti, T.; Hegab, M.M.; Ali, S.O.; Khemira, H. Essential oils, amino acids and polyphenols changes in salt-stressed Thymus vulgaris exposed to open–field and shade enclosure. Ind. Crops Prod. 2016, 91, 223–230. [Google Scholar] [CrossRef]
- Bistgani, Z.E.; Hashemi, M.; Da Costa, M.; Craker, L.; Maggi, F.; Morshedloo, M.R. Effect of salinity stress on the physiological characteristics, phenolic compounds and antioxidant activity of Thymus vulgaris L. and Thymus daenensis Celak. Ind. Crops Prod. 2019, 135, 311–320. [Google Scholar] [CrossRef]
- Soheilikhah, Z.; Modarresi, M.; Karimi, N.; Movafegh, A. Qualitative and quantitative analysis of diosmin content of hyssop (Hyssopus officinalis) in response to salinity stress. Heliyon 2021, 7, e08228. [Google Scholar] [CrossRef]
- Brullo, S.; Brullo, C.; Sciandrello, S.; Tavilla, G.; Cambria, S.; Tomaselli, V.; Ilardi, V.; Giusso del Galdo, G.; Minissale, P. The plant communities of the class isoëto-nanojuncetea in Sicily. Plants 2022, 11, 1214. [Google Scholar] [CrossRef]
- Sanchez, D.H.; Siahpoosh, M.R.; Roessner, U.; Udvardi, M.; Kopka, J. Plant metabolomics reveals conserved and divergent metabolic responses to salinity. Physiol. Plant. 2008, 132, 209–219. [Google Scholar] [CrossRef]
- Chiappero, J.; Cappellari, L.d.R.; Sosa Alderete, L.G.; Palermo, T.B.; Banchio, E. Plant growth promoting rhizobacteria improve the antioxidant status in Mentha piperita grown under drought stress leading to an enhancement of plant growth and total phenolic content. Ind. Crops Prod. 2019, 139, 111553. [Google Scholar] [CrossRef]
- Farhadi, N.; Ghassemi-Golezani, K. Physiological changes of Mentha pulegium in response to exogenous salicylic acid under salinity. Sci. Hortic. 2020, 267, 109325. [Google Scholar] [CrossRef]
- de Medeiros Gomes, J.; Cahino Terto, M.V.; Golzio do Santos, S.; Sobral da Silva, M.; Fechine Tavares, J. Seasonal variations of polyphenols content, sun protection factor and antioxidant activity of two Lamiaceae species. Pharmaceutics 2021, 13, 110. [Google Scholar] [CrossRef] [PubMed]
- Chrysargyris, A.; Papakyriakou, E.; Petropoulos, S.A.; Tzortzakis, N. The combined and single effect of salinity and copper stress on growth and quality of Mentha spicata plants. J. Hazard. Mater. 2019, 368, 584–593. [Google Scholar] [CrossRef] [PubMed]
- Eftekhari, A.; Khusro, A.; Ahmadian, E.; Dizaj, S.M.; Hasanzadeh, A.; Cucchiarini, M. phytochemical and nutra-pharmaceutical attributes of Mentha spp.: A comprehensive review. Arab. J. Chem. 2021, 14, 103106. [Google Scholar] [CrossRef]
- Fatiha, B.; Didier, H.; Naima, G.; Khodir, M.; Martin, K.; Léocadie, K.; Caroline, S.; Mohamed, C.; Pierre, D. Phenolic composition, in vitro antioxidant effects and tyrosinase inhibitory activity of three Algerian Mentha species: M. spicata (L.), M. pulegium (L.) and M. rotundifolia (L.) Huds (Lamiaceae). Ind. Crops Prod. 2015, 74, 722–730. [Google Scholar] [CrossRef]
- Bahadori, M.B.; Zengin, G.; Bahadori, S.; Dinparast, L.; Movahhedin, N. Phenolic composition and functional properties of wild mint (Mentha longifolia var. Calliantha (Stapf) Briq.). Int. J. Food Prop. 2018, 21, 183–193. [Google Scholar] [CrossRef] [Green Version]
- Ćavar Zeljković, S.; Šišková, J.; Komzáková, K.; De Diego, N.; Kaffková, K.; Tarkowski, P. Phenolic compounds and biological activity of selected Mentha species. Plants 2021, 10, 550. [Google Scholar] [CrossRef]
- Pistelli, L.; Tonelli, M.; Pellegrini, E.; Cotrozzi, L.; Pucciariello, C.; Trivellini, A.; Lorenzini, G.; Nali, C. Accumulation of rosmarinic acid and behaviour of ros processing systems in Melissa officinalis L. under heat stress. Ind. Crops Prod. 2019, 138, 111469. [Google Scholar] [CrossRef]
- Vafadar, F.; Amooaghaie, R.; Ehsanzadeh, P.; Ghanadian, M. Salinity stress alters ion homeostasis, antioxidant activities and the production of rosmarinic acid, luteolin and apigenin in Dracocephalum kotschyi Boiss. Biologia 2020, 75, 2147–2158. [Google Scholar] [CrossRef]
- Sharma, A.; Shahzad, B.; Rehman, A.; Bhardwaj, R.; Landi, M.; Zheng, B. Response of phenylpropanoid pathway and the role of polyphenols in plants under abiotic stress. Molecules 2019, 24, 2452. [Google Scholar] [CrossRef] [Green Version]
- Bautista, I.; Boscaiu, M.; Lidón, A.; Llinares, J.V.; Lull, C.; Donat, M.; Mayoral, O.; Vicente, O. Environmentally induced changes in antioxidant phenolic compounds levels in wild plants. Acta Physiol. Plant. 2016, 38, 9. [Google Scholar] [CrossRef]
- Kaulen, H.; Schell, J.; Kreuzaler, F. Light-induced expression of the chimeric chalcone synthase-NPTII gene in tobacco cells. EMBO J. 1986, 5, 1–8. [Google Scholar] [CrossRef]
- Koes, R.E.; Spelt, C.E.; Mol, J.N. The chalcone synthase multigene family of petunia hybrida (V30): Differential, light-regulated expression during flower development and uv light induction. Plant Mol. Biol. 1989, 12, 213–225. [Google Scholar] [CrossRef]
- Dolzhenko, Y.; Bertea, C.M.; Occhipinti, A.; Bossi, S.; Maffei, M.E. UV-B Modulates the interplay between terpenoids and flavonoids in peppermint (Mentha x piperita L.). J. Photochem. Photobiol. B Biol. 2010, 100, 67–75. [Google Scholar] [CrossRef]
- Park, Y.J.; Baek, S.A.; Choi, Y.; Kim, J.K.; Park, S.U. Metabolic profiling of nine Mentha species and prediction of their antioxidant properties using chemometrics. Molecules 2019, 24, 258. [Google Scholar] [CrossRef] [Green Version]
- Moshrefi-Araghi, A.; Nemati, H.; Azizi, M.; Moshtaghi, N.; Shoor, M. Association of genetic structure and diversity in iranian wild germplasms of Mentha longifolia L. based on phenotypical, biochemical, and molecular markers. Chem. Biodivers. 2021, 18, e2001044. [Google Scholar] [CrossRef]
- Kim, G.D.; Park, Y.S.; Jin, Y.H.; Cheung-Seog, P. Production and applications of rosmarinic acid and structurally related compounds. Appl. Microbiol. Biotechnol. 2015, 99, 2083–2092. [Google Scholar] [CrossRef]
- Zhao, J.; Xu, L.; Jin, D.; Xin, Y.; Tian, L.; Wang, T.; Zhao, D.; Wang, Z.; Wang, J. Rosmarinic acid and related dietary supplements: Potential applications in the prevention and treatment of cancer. Biomolecules 2022, 12, 1410. [Google Scholar] [CrossRef]
- Fletcher, R.S.; Slimmon, T.; Kott, L.S. Environmental factors affecting the accumulation of rosmarinic acid in spearmint (Mentha spicata L.) and peppermint (Mentha piperita L.). Open Agric. J. 2010, 4, 10–16. [Google Scholar] [CrossRef]
- Nath, L.R.; Gorantla, J.N.; Joseph, S.M.; Antony, J.; Thankachan, S.; Menon, D.B.; Sankar, S.; Lankalapalli, R.S.; Anto, R.J. Kaempferide, the most active among the four flavonoids isolated and characterized from Chromolaena odorata, induces apoptosis in cervical cancer cells while being pharmacologically safe. RSC Adv. 2015, 5, 100912–100922. [Google Scholar] [CrossRef]
- Eguchi, H.; Matsunaga, T.; Endo, S.; Ichihara, K.; Ikari, A. Kaempferide enhances chemosensitivity of human lung adenocarcinoma A549 cells mediated by the decrease in phosphorylation of akt and claudin-2 expression. Nutrients 2020, 12, 1190. [Google Scholar] [CrossRef] [PubMed]
- Elhady, S.S.; Eltamany, E.E.; Shaaban, A.E.; Bagalagel, A.A.; Muhammad, Y.A.; El-Sayed, N.M.; Ayyad, S.N.; Ahmed, A.A.M.; Elgawish, M.S.; Ahmed, S.A. Jaceidin flavonoid isolated from chiliadenus montanus attenuates tumor progression in mice via VEGF inhibition: In vivo and in silico studies. Plants 2020, 9, 1031. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, T.P.; Feder, N.; McCully, M.E. Polychromatic staining of plant cell walls by toluidine blue O. Protoplasma 1964, 59, 368–373. [Google Scholar] [CrossRef]
- O’Brien, T.P.; McCully, M.E. The Study of Plant Structure: Principles and Selected Methods; Termarcarphi Pty. Ltd.: Melbourne, Australia, 1981. [Google Scholar]
- Chieco, C.; Rotondi, A.; Morrone, L.; Rapparini, F.; Baraldi, R. An ethanol-based fixation method for anatomical and micro-morphological characterization of leaves of various tree species. Biotech. Histochem. 2013, 88, 109–119. [Google Scholar] [CrossRef]
- Smeriglio, A.; De Francesco, C.; Denaro, M.; Trombetta, D. Prickly pear betalain-rich extracts as new promising strategy for intestinal inflammation: Plant complex vs. main isolated bioactive compounds. Front. Pharmacol. 2021, 12, 722398. [Google Scholar] [CrossRef]
- Monforte, M.T.; Smeriglio, A.; Germanò, M.P.; Pergolizzi, S.; Circosta, C.; Galati, E.M. Evaluation of antioxidant, antiinflammatory, and gastroprotective properties of Rubus fruticosus L. fruit juice. Phytother. Res. 2018, 32, 1404–1414. [Google Scholar] [CrossRef]
- Danna, C.; Bazzicalupo, M.; Ingegneri, M.; Smeriglio, A.; Trombetta, D.; Burlando, B.; Cornara, L. Anti-inflammatory and wound healing properties of leaf and rhizome extracts from the medicinal plant Peucedanum ostruthium (L.) W. D. J. Koch. Molecules 2022, 27, 4271. [Google Scholar] [CrossRef]
- Smeriglio, A.; Bonasera, S.; Germanò, M.P.; D’Angelo, V.; Barreca, D.; Denaro, M.; Monforte, M.T.; Galati, E.M.; Trombetta, D. Opuntia ficus-indica (L.) Mill. fruit as source of betalains with antioxidant, cytoprotective, and anti-angiogenic properties. Phytother. Res. 2019, 33, 1526–1537. [Google Scholar] [CrossRef]
- Occhiuto, C.; Aliberto, G.; Ingegneri, M.; Trombetta, D.; Circosta, C.; Smeriglio, A. Comparative evaluation of the nutrients, phytochemicals, and antioxidant activity of two hempseed oils and their byproducts after cold pressing. Molecules 2022, 27, 3431. [Google Scholar] [CrossRef]
- Bazzicalupo, M.; Burlando, B.; Denaro, M.; Barreca, D.; Trombetta, D.; Smeriglio, A.; Cornara, L. Polyphenol characterization and skin-preserving properties of hydroalcoholic flower extract from Himantoglossum robertianum (Orchidaceae). Plants 2019, 8, 502. [Google Scholar] [CrossRef] [Green Version]
- Bellocco, E.; Barreca, D.; Laganà, G.; Calderaro, A.; El Lekhlifi, Z.; Chebaibi, S.; Smeriglio, A.; Trombetta, D. Cyanidin-3-O-galactoside in ripe pistachio (Pistacia vera L. variety Bronte) hulls: Identification and evaluation of its antioxidant and cytoprotective activities. J. Funct. Foods 2016, 27, 376–385. [Google Scholar] [CrossRef]
- Smeriglio, A.; Denaro, M.; Di Gristina, E.; Mastracci, L.; Grillo, F.; Cornara, L.; Trombetta, D. Pharmacognostic approach to evaluate the micromorphological, phytochemical and biological features of Citrus lumia seeds. Food Chem. 2022, 375, 131855. [Google Scholar] [CrossRef]
- Smeriglio, A.; Denaro, M.; D’Angelo, V.; Germanò, M.P.; Trombetta, D. Antioxidant, anti-inflammatory and anti-angiogenic properties of Citrus lumia Juice. Front. Pharmacol. 2020, 11, 593506. [Google Scholar] [CrossRef]
- Smeriglio, A.; Denaro, M.; Barreca, D.; D’Angelo, V.; Germanò, M.P.; Trombetta, D. Polyphenolic profile and biological activities of black carrot crude extract (Daucus carota L. ssp. sativus var. atrorubens Alef.). Fitoterapia 2018, 124, 49–57. [Google Scholar] [CrossRef]
- Certo, G.; Costa, R.; D’Angelo, V.; Russo, M.; Albergamo, A.; Dugo, G.; Germanò, M.P. Anti-angiogenic activity and phytochemical screening of fruit fractions from Vitex agnus castus. Nat. Prod. Res. 2017, 31, 2850–2856. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Micromorphological Analysis | LC-DAD Analysis * | ||
|---|---|---|---|---|
| Abundance of crystals | mg diosmin/100 g DE a | |||
| Leaves | Flowers | Leaves | Flowers | |
| MPI | +++ | +++ | 66.60 ± 0.44 b | 27.47 ± 0.24 b |
| MPII | ++ | ++ | 65.76 ± 0.21 | 10.46 ± 0.12 c |
| MPIII | ++ | + | 65.60 ± 0.33 | 4.61 ± 0.08 |
| Sample | Total Phenolics (g GAE/100 g DE) | Flavonoids (g RE/100 g DE) | Flavan-3-ols (g CE/100 g DE) |
|---|---|---|---|
| MPLE I | 3.39 ± 0.32 a | 22.83 ± 1.42 a,b | 8.48 ± 0.23 a,b |
| MPLE II | 7.87 ± 0.72 b | 6.13 ± 0.18 b | 2.33 ± 0.11 b |
| MPLE III | 3.96 ± 0.28 | 6.54 ± 0.11 | 3.50 ± 0.21 |
| MPFE I | 11.15 ± 0.42 a,b | 23.73 ± 1.35 a,b | 2.75 ± 0.17 b |
| MPFE II | 8.87 ± 0.80 b | 6.53 ± 0.14 b | 2.86 ± 0.15 b |
| MPFE III | 5.21 ± 0.20 | 27.62 ± 2.63 | 4.75 ± 0.22 |
| n. | Compound | λmax nm | [M-H]- m/z | MPLE (Area %) | MPFE (Area %) | ||||
|---|---|---|---|---|---|---|---|---|---|
| I | II | III | I | II | III | ||||
| 1 | Riboflavine 5’-(dihydrogen phosphate) | 300 | 455 | - | 3.34 ± 0.16 b | 19.23 ± 0.56 | - | - | 7.31 ± 0.33 |
| 2 | Epicatechin hydroxybenzoate | 272 | 409 | - | - | 0.73 ± 0.03 | 1.10 ± 0.04 a | 0.29 ± 0.01 b | 0.26 ± 0.01 |
| 3 | Gallocatechin-3-gallate § | 278 | 457 | - | - | - | - | 0.70 ± 0.03 | - |
| 4 | Epigallocatechin-3-O-gallate § | 274 | 457 | 0.37 ± 0.01 | - | - | - | - | - |
| 5 | Floridin | 256,282 | 414 | - | - | 2.48 ± 0.08 | - | - | - |
| 6 | Caffeoylhydroxycitric acid | 236,326 | 369 | - | - | - | - | - | 3.68 ± 0.15 |
| 7 | Ferulic acid hexoside | 234,322 | 355 | - | - | - | 0.68 ± 0.02 | - | - |
| 8 | Catechin hexoside | 276,320 | 451 | - | 0.28 ± 0.01 | - | - | 0.13 ± 0.00 | - |
| 9 | Coumaroylquinic acid | 216,320 | 338 | 0.72 ± 0.02 | - | - | - | - | - |
| 10 | Salvianolic acid J | 276,344 | 537 | - | - | 0.42 ± 0.02 | - | - | - |
| 11 | Epicatechin methylgallate | 278 | 455 | - | - | - | 0.38 ± 0.01 | - | - |
| 12 | Malonyl-daidzin | 264,308 | 297 | - | - | 0.72 ± 0.01 | - | - | 8.3 ± 0.27 |
| 13 | Luteolin malonyl-hexoside | 263,348 | 533 | 0.88 ± 0.03 | - | - | - | - | - |
| 14 | Apigenin apiosyl-hexoside | 268,328 | 563 | - | 5.43 ± 0.17 | - | - | 3.37 ± 0.12 | - |
| 15 | Unknown | 300 | 567 | - | - | 2.98 ± 0.12 | - | - | 0.98 ± 0.03 |
| 16 | Unknown | 300 | 391 | - | - | 1.13 ± 0.03 | - | - | 1.35 ± 0.05 |
| 17 | Epitheaflagallin 3-O-gallate § | 260,294 | 551 | - | - | - | - | 0.62 ± 0.03 b | 5.35 ± 0.18 |
| 18 | Epigallocatechin § | 278 | 305 | - | - | - | 2.07 ± 0.08 | - | - |
| 19 | Gallocatechin § | 278 | 305 | 8.31 ± 0.15 a | 1.99 ± 0.07 b | 0.64 ± 0.02 | - | - | - |
| 20 | Eriodictyol-7-O-glucoside | 296,326 | 449 | - | - | - | - | 0.99 ± 0.02 | - |
| 21 | Medioresinol § | 278 | 387 | - | - | 0.15 ± 001 | - | - | - |
| 22 | Dimethylepigallocatechin gallate | 278 | 485 | - | - | 3.52 ± 0.12 | - | - | 1.91 ± 0.05 |
| 23 | Eriodictyol § | 292,328 | 287 | - | - | - | - | 0.38 ± 0.01 | - |
| 24 | Tuberonic acid hexoside | 274,304 | 387 | - | 1.17 ± 0.04 b | 0.96 ± 0.02 | 0.35 ± 0.01 a | 1.05 ± 0.03 b | 1.68 ± 0.04 |
| 25 | Methyl-galloylgalactarate | 226,268 | 375 | 2.27 ± 0.05 | - | - | - | - | - |
| 26 | Hydroxyluteolin-xyloside | - | 433 | - | - | - | - | - | 0.20 ± 0.01 |
| 27 | Trihydroxy-methoxy-MDF-glucuronide | 274,314 | 519 | - | 0.42 ± 0.02 b | 1.08 ± 0.04 | - | - | 0.47 ± 0.02 |
| 28 | Epigallocatechin digallate | 278 | 609 | - | - | - | 0.63 ± 0.02 a | 0.38 ± 0.02 | - |
| 29 | Unknown | - | 374 | - | - | - | - | - | 0.30 ± 0.02 |
| 30 | Limocitrin§ | 280,328,379 | 375 | - | - | - | - | 0.49 ± 0.03 b | 0.24 ± 0.01 |
| 31 | Kaempferol 3-O-glucuronide § | 272,368 | 461 | - | - | - | - | - | 5.28 ± 0.12 |
| 32 | Luteolin 7-O-rutinoside § | 272,344 | 593 | 2.89 ± 0.03 a | 2.37 ± 0.14 b | 5.36 ± 0.25 | 2.98 ± 0.17 a | 1.65 ±0.04 b | 0.27 ± 0.02 |
| 33 | Kaempferol 3-O-rutinoside § | 266,323,364 | 593 | - | - | - | - | - | 0.55 ± 0.02 |
| 34 | Unknown | 250,294 | 559 | - | - | - | - | 0.25 ± 0.01 | - |
| 35 | Salvianolic acid G | 258,346 | 417 | 0.30 ± 0.01 a | 0.78 ± 0.04 b | 1.37 ± 0.02 | - | 2.40 ± 0.12 b | 1.05 ± 0.24 |
| 36 | Unknown | 256,326 | 639 | - | 0.34 ± 0.02 | - | - | - | - |
| 37 | Salvianolic acid K | 286,321 | 555 | - | - | - | - | - | 0.75 ± 0.02 |
| 38 | Ellagic acid acetyl pentoside | 256,326 | 475 | - | 0.31 ± 0.01 b | 0.59 ± 0.02 | - | 0.22 ± 0.01 | - |
| 39 | 5-Nonadecylresorcinol | 275 | 375 | - | - | - | - | - | 0.25 ± 0.01 |
| 40 | Quercetin acetyl hexoside | 262,354 | 505 | - | - | - | 0.96 0.03 | - | - |
| 41 | Todolactol A | 236,288 | 375 | - | - | - | - | - | 6.71 ± 0.22 |
| 42 | Limocitrol | 280,327,390 | 375 | - | 2.0 ± 0.08 b | 2.54 ± 0.08 | - | 1.51 ±0.05 | - |
| 43 | Caffeoyl-rosmarinic acid | 284,328 | 537 | - | - | - | - | 0.15 ± 0.00 | - |
| 44 | Valoneic acid dilactone | 298 | 469 | - | - | - | - | - | 0.5 ± 0.02 |
| 45 | Syringic acid hexose | 240,318 | 359 | 0.83 ± 0.04 | - | - | - | - | - |
| 46 | Unknown | 282,344 | 491 | - | 0.23 ± 0.01 | - | - | - | - |
| 47 | Quercetin 3-O-(6’’-O-malonyl-β-D-glucoside) § | 262,352 | 549 | - | - | 1.34 ± 0.03 | - | - | 0.85 ± 0.03 |
| 48 | Unknown | 300 | 442 | - | - | - | - | 0.09 ± 0.00 | - |
| 49 | Salvianolic acid I | 252,318,352 | 537 | 3.15 ± 0.05 a | 8.41 ± 0.22 | - | 6.82 ± 0.22 a | 8.76 ± 0.32 | - |
| 50 | Dihydroquercetin 3-rhamnoside § | 254,368 | 449 | - | - | 0.37 ± 0.02 | - | - | - |
| 51 | Apigenin 6,8-di-C-glucoside § | 244,266,334 | 593 | 0.72 ± 0.02 a | - | 1.35 ± 0.03 | 2.01 ± 0.12 a | - | 1.50 ± 0.05 |
| 52 | Salvianolic acid D § | 254,347 | 417 | 0.64 ± 0.01 a | 3.41 ± 0.11 b | 0.82 ± 0.02 | - | 0.67 ± 0.02 b | 1.24 ± 0.04 |
| 53 | Myricetin acetylhexoside | 240,374 | 521 | - | - | 1.28 ± 0.04 | - | - | - |
| 54 | Unknown | 248,312,364 | 513 | - | - | 0.81 ± 0.04 | - | - | - |
| 55 | Galloylgalactaric acid | 209,275 | 361 | 8.71 ± 0.11 | - | - | - | - | - |
| 56 | Daidzin § | 252,312 | 415 | - | - | 1.17 ± 0.02 | - | - | 0.92 ± 0.02 |
| 57 | Unknown | 240,310 | 559 | - | - | 0.45 ± 0.01 | - | - | - |
| 58 | 3,4-Dicaffeoylquinic acid § | 216,326 | 515 | - | - | 2.08 ± 0.13 | 11.64 ± 0.67 a | - | 2.06 ± 0.08 |
| 59 | Salvianolic acid B § | 244,285,336 | 717 | - | 0.25 ± 0.01 | - | 5.50 ± 0.22 a | 6.85 ± 0.28 | - |
| 60 | Isosalvianolic acid B | 254,285,340 | 717 | - | - | - | 0.79 ± 0.03 a | 6.41 ± 0.32 | - |
| 61 | Syringaresinol § | 240,271 | 417 | - | - | - | - | - | 1.22 ± 0.04 |
| 62 | Diosmin § | 260,350 | 607 | 3.61 ± 0.06 a | 1.23 ± 0.03 b | 3.33 ± 0.14 | 0.54 ± 0.02 a | 0.11 ± 0.00 b | 0.15 ± 0.01 |
| 63 | Unknown | 246,266,312 | 424 | - | - | - | - | 0.20 ± 0.01 | - |
| 64 | Salvianolic acid K | 250,270,345 | 555 | - | 0.61 ± 0.03 b | 2.95 ± 0.11 | - | 0.25 ± 0.01 | - |
| 65 | Unknown | 300 | 553 | - | - | 0.12 ± 0.00 | - | - | - |
| 66 | Rosmarinic acid § | 292,332 | 359 | 0.52 ± 0.02 a | 19.16 ± 0.48 b | 0.26 ± 0.01 | 24.31 ± 0.84 a | 25.52 ± 0.76 | - |
| 67 | Malonylglycitin | 256,320 | 531 | - | - | 0.46 ± 0.02 | - | - | - |
| 68 | Pentacosenylresorcinol | 300 | 457 | - | - | - | - | - | 0.63 ± 0.02 |
| 69 | Salvianolic acid E § | 286,314,350 | 717 | - | 6.08 ± 0.12 | - | 1.94 ± 0.06 a | 1.74 ± 0.04 | - |
| 70 | Unknown | 246,266,312 | 506 | - | - | 1.51 ± 0.04 | - | 0.05 ± 0.00 | - |
| 71 | Lithospermic acid § | 238,336 | 537 | - | - | - | 1.59 ± 0.04 a | 0.37 ± 0.02 | - |
| 72 | Unknown | 286,323 | 519 | - | 1.01 ± 0.02 | - | - | - | - |
| 73 | Salvianolic acid L | 286,314,340 | 717 | - | - | - | 2.33 ± 0.08 | - | - |
| 74 | Glucoerucin § | 217,236 | 419 | - | - | 0.6 ± 0.03 | - | 0.31 ± 0.01 | - |
| 75 | 3,5-Dicaffeoylquinic acid § | 216,322 | 515 | - | 0.22 ± 0.01 | - | - | - | 0.74 ± 0.03 |
| 76 | Quercetin hydroxybenzoylhexoside | 257,358 | 583 | 2.37 ± 0.04 | - | - | - | - | - |
| 77 | Unknown | 266,300,334 | 531 | 0.26 ± 0.01 | - | - | - | - | - |
| 78 | Isorhamnetin sulfate | 253,370 | 395 | - | - | 0.83 ± 0.02 | - | - | 1.23 ± 0.04 |
| 79 | Apigenin 8-C-α-L-arabinoside 6-C-β-D-glucoside § | 268,333 | 563 | - | - | 0.27 ± 0.01 | - | - | - |
| 80 | Rosmanol § | 262,288 | 345 | - | - | - | - | 5.51 ± 0.22 | - |
| 81 | Syringetin § | 284,370 | 345 | - | 0.22 ± 0.00 | - | 0.13 ± 0.01 | - | - |
| 82 | Salvianolic acid C § | 264,342 | 491 | - | 0.22 ± 0.01 b | 1.89 ± 0.04 | 6.04 ± 0.22 a | 0.19 ± 0.01 b | 0.88 ± 0.03 |
| 83 | Isorhamnetin 3-glucuronide § | 260,370 | 491 | - | 8.04 ± 0.32 | - | - | - | 0.45 ± 0.02 |
| 84 | Unknown | 222,290,362 | 581 | 0.28 ± 0.01 | - | - | - | - | - |
| 85 | Luteolin § | 254,266,346 | 285 | 3.65 ± 0.15 | - | - | - | - | - |
| 86 | Mangiferin gallate | 260,362 | 573 | - | 0.11 ± 0.00 | - | - | - | - |
| 87 | Peonidin § | 282,526 | 300 | - | - | - | 0.78 ± 0.04 a | 0.58 ± 0.04 | 0.65 ± 0.03 |
| 88 | Patuletin hexoside | 262,354 | 493 | - | 0.42 ± 0.02 | - | - | - | - |
| 89 | Jaceosidin § | 214,282,342 | 329 | - | 4.44 ± 0.12 b | 1.42 ± 0.05 | 1.35 ± 0.03 a | 3.50 ± 0.14 b | 2.6 ± 0.01 |
| 90 | Unknown | 216,286,402 | 328 | - | - | - | - | 0.09 ± 0.00 | - |
| 91 | Unknown | 288,312,398 | 598 | - | - | - | 1.01 ± 0.02 | - | - |
| 92 | Diosmetin 7-glucuronide § | 230,266,346 | 475 | - | - | - | - | 0.05 ± 0.00 | - |
| 93 | Salvigenin § | 284,334 | 327 | - | 0.37 ± 0.01 b | 0.96 ± 0.03 | 0.76 ± 0.03 | - | 1.15 ± 0.02 |
| 94 | Jaceidin Isomer A | 282,352 | 359 | 4.84 ± 0.18 a | 14.47 ± 0.58 b | 23.63 ± 1.05 | 15.73 ± 0.83 a | 13.45 ± 0.48 b | 30.79 ± 1.58 |
| 95 | Kaempferide § | 254,266,344 | 299 | 52.81 ± 0.64 | - | - | - | - | - |
| 96 | Jaceosidin Isomer I | 252,268,346 | 329 | - | 1.00 ± 0.04 b | 0.27 ± 0.01 | 0.73 ± 0.03 a | 0.21 ± 0.01 b | 1.51 ± 0.05 |
| 97 | Quercetin sulfate | 246,354 | 381 | - | - | - | - | 1.77 ± 0.05 | - |
| 98 | Jaceidin isomer II | 270,354 | 359 | - | 1.74 ± 0.03 b | 0.76 ± 0.03 | 0.64 ± 0.02 a | - | 1.40 ± 0.04 |
| 99 | Phloretin xylosyl-hexoside | 230,288 | 567 | - | - | 0.49 ± 0.02 | - | - | 0.36 ± 0.01 |
| 100 | 6-Methoxyapigenin § | 260,330 | 299 | - | 0.32 ± 0.01 | - | - | - | - |
| 101 | Unknown | 254,264,348 | 344 | - | - | 0.65 ± 0.03 | - | - | - |
| 102 | Apigenina-7-glucoside § | 268,330 | 431 | 0.72 ± 0.03 | - | - | - | - | - |
| 103 | Methyl digalloyl-β-D-hexopyranoside | 268,310,360 | 497 | 0.59 ± 0.01 | - | - | - | - | - |
| 104 | Dihydroxyphenyl galloyl-β-D-hexopyranoside | 252,290,342 | 439 | 0.56 ± 0.02 a | 7.14 ± 0.21 b | 0.45 ± 0.02 | 6.21 ± 0.02 a | 8.27 ± 0.14 b | 0.96 ± 0.02 |
| 105 | 5-Galloylquinic acid § | - | 343 | - | 0.28 ± 0.01 | - | - | - | - |
| 106 | Trachelogenin § | - | 387 | - | 0.07 ± 0.00 | - | - | - | - |
| 107 | Unknown | - | 381 | - | - | - | - | 0.06 ± 0.00 | - |
| 108 | Hexopyranuronosyl-xylose | 266,334 | 325 | - | 0.13 ± 0.00 | - | - | 0.09 ± 0.00 b | 0.69 ± 0.03 |
| 109 | Quercetin 4’-glucuronide § | 258,368 | 477 | - | - | 0.64 ± 0.02 | - | - | - |
| 110 | Unknown | 266,332 | 488 | - | - | - | - | 0.06 ± 0.00 | - |
| 111 | Epigallocatechin-caffeate | 274,344 | 467 | - | 1.56 ± 0.03 b | 4.0 ± 0.21 | - | - | - |
| 112 | Unknown | 276,340 | 675 | - | 0.43 ± 0.01 b | 0.94 ± 0.02 | - | 0.26 ± 0.01 b | 0.63 ± 0.02 |
| Sample | Collection Site | Altitude (m a.s.l) a | GPS Coordinates | T b (°C) | P c (mm/year) | Soil | Voucher |
|---|---|---|---|---|---|---|---|
| MPI | Isola Lunga- Stagnone di Marsala (Trapani) | 2.00 | 37°54′06″ N, 12°27′07″ E | 18.5 | 520 | Clayey-saline depressions | R&Sp d 05/19 |
| MPII | Castronovo di Sicilia (Palermo) | 338 | 37°40′09″ N, 13°38′54″ E | 16.5 | 700 | Clayey-moist | R&Sc e 06/19 |
| MPIII | Castellana Sicula (Palermo) | 860 | 37°45′42″ N, 13°59′73″ E | 16.0 | 800 | Clayey-moist | R&Sc e 07/19 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cornara, L.; Sgrò, F.; Raimondo, F.M.; Ingegneri, M.; Mastracci, L.; D’Angelo, V.; Germanò, M.P.; Trombetta, D.; Smeriglio, A. Pedoclimatic Conditions Influence the Morphological, Phytochemical and Biological Features of Mentha pulegium L. Plants 2023, 12, 24. https://doi.org/10.3390/plants12010024
Cornara L, Sgrò F, Raimondo FM, Ingegneri M, Mastracci L, D’Angelo V, Germanò MP, Trombetta D, Smeriglio A. Pedoclimatic Conditions Influence the Morphological, Phytochemical and Biological Features of Mentha pulegium L. Plants. 2023; 12(1):24. https://doi.org/10.3390/plants12010024
Chicago/Turabian StyleCornara, Laura, Federica Sgrò, Francesco Maria Raimondo, Mariarosaria Ingegneri, Luca Mastracci, Valeria D’Angelo, Maria Paola Germanò, Domenico Trombetta, and Antonella Smeriglio. 2023. "Pedoclimatic Conditions Influence the Morphological, Phytochemical and Biological Features of Mentha pulegium L." Plants 12, no. 1: 24. https://doi.org/10.3390/plants12010024