


The Highly Embryogenic Brassica napus DH4079 Line Is Recalcitrant to Agrobacterium-Mediated Genetic Transformation

Cell Biology Group-COMAV Institute, Universitat Politècnica de València, 46022 Valencia, Spain
*
Authors to whom correspondence should be addressed.
†
These authors contributed equally to this work.
‡
Present address: Instituto de Biología Molecular y Celular de Plantas (UPV-CSIC), 46022 Valencia, Spain.
Plants 2023, 12(10), 2008; https://doi.org/10.3390/plants12102008
Submission received: 22 April 2023
/
Revised: 12 May 2023
/
Accepted: 14 May 2023
/
Published: 17 May 2023
(This article belongs to the Special Issue Genetics and Epigenetics in the Brassicaceae)
Abstract
:Brassica napus is a species of high agronomic interest, used as a model to study different processes, including microspore embryogenesis. The DH4079 and DH12075 lines show high and low embryogenic response, respectively, which makes them ideal to study the basic mechanisms controlling embryogenesis induction. Therefore, the availability of protocols for genetic transformation of these two backgrounds would help to generate tools to better understand this process. There are some reports in the literature showing the stable transformation of DH12075. However, no equivalent studies in DH4079 have been reported to date. We explored the ability of DH4079 plants to be genetically transformed. As a reference to compare with, we used the same protocols to transform DH12075. We used three different protocols previously reported as successful for B. napus stable transformation with Agrobacterium tumefaciens and analyzed the response of plants. Whereas DH12075 plants responded to genetic transformation, DH4079 plants were completely recalcitrant, not producing any single regenerant out of the 1784 explants transformed and cultured. Additionally, an Agrobacterium rhizogenes transient transformation assay was performed on both lines, and only DH12075, but no DH4079 seedlings, responded to A. rhizogenes infection. Therefore, we propose that the DH4079 line is recalcitrant to Agrobacterium-mediated transformation.
1. Introduction
Genetic transformation consists of the introduction of a foreign DNA molecule into a genome, generating a genetically modified organism. The first reports on plant genetic transformation date from 1983 and used Agrobacterium tumefaciens as a transformation vector to introduce foreign DNA into the genomes of different plant species [1]. Among the different technologies available for plant genetic transformation, Agrobacterium-mediated transformation is nowadays the most used approach. The use of genetically modified crops has increased both plant productivity and farmer profits, while reducing the use of pesticides, among other advantages [2]. Indeed, a total of 438 genetically modified plant events have been approved worldwide [3]. More recently, the impact of biotechnology on agriculture has been enhanced by different genome editing techniques, such as zinc-finger nucleases, transcription activator-like effector nucleases (TALENs), and clustered regularly interspaced short palindromic repeats (CRISPR) and CRISPR-associated protein 9 (Cas9). The use of CRISPR/Cas9 technology for genome editing implies the previous generation of genetically modified plants [4]. On the other hand, the use of genetic transformation to generate gain and loss-of-function lines in different model and crop species has helped us to understand gene function and constitutes a powerful biotechnological tool for basic and applied plant genetic and molecular research [5]. Therefore, the availability of efficient plant transformation protocols for different crops and plant species is crucial for the improvement of crop performance for agriculture, as well as for the generation of new biotechnological research tools.
Microspore embryogenesis is a process whereby the male gametophyte deviates from its original developmental pathway and is induced to develop as a haploid embryo whose genome can then be doubled to become a doubled haploid (DH), fully homozygous individual [6]. This process constitutes a powerful biotechnological tool for both basic research [7,8] and applied plant breeding [9]. Brassica napus is a model species for the study of this experimental pathway [10,11] and the processes involved in its in vitro induction [12,13,14,15,16]. This is due to the high potential of the microspores of some genotypes, such as the DH4079 line, to become induced to embryogenesis [11,17]. In parallel, other lines such as the DH12075 line, consistently show a low response to the same embryogenesis-inductive conditions [17]. Thus, B. napus is an interesting target species to use to combine transformation and microspore embryogenesis protocols in order to develop DH individuals from haploid-transformed microspores, thereby fixing the transgene in homozygosis and avoiding the occurrence of hemizygous regenerants [11]. The development of successful transformation protocols for B. napus lines with different embryogenic competence would greatly help to develop biotechnological tools to study the cellular and molecular basis of this morphogenic process.
The type of explant tissue, the selection marker used and the genotype have been reported among the main factors affecting the efficiency of stable B. napus transformation [18,19]. Some of the first attempts to transform B. napus were based on the use of cotyledon petioles as explants [20]. Afterwards, other effective protocols have been developed, also based on the use of cotyledons [21] or other explants such as stem portions [22], protoplasts [23] or hypocotyl sections [24]. Among the different selection markers available, kanamycin has been commonly used as an effective selection agent in B. napus transformation, whereas the use of BASTA has been less frequent [25]. The third major factor is the genotype. In other members of the brassicaceae family, such as B. oleracea, there are protocols to stably transform with Agrobacterium different genotypes of crops such as cauliflower and broccoli, among others, with low genotype dependence [26]. However, different B. napus genotypes show different responses to Agrobacterium-mediated transformation [24]. Several works reported the successful stable transformation of the low embryogenesis-responsive lines Westar [20,22] and DH12075 [27,28], whereas to our knowledge, no transgenic lines have been described so far for the highly embryogenic DH4079 line. Whether DH4079 is extremely recalcitrant to Agrobacterium-mediated transformation, or simply no transformed DH4079 lines have been reported yet, is unknown.
In this work, we aimed to determine an optimal protocol for Agrobacterium-mediated stable transformation of the B. napus DH4079 line using different conditions, including different plant explants, plasmids, Agrobacterium strains and incubation times. As references to compare with, we also attempted stable transformation of the DH12075 line with the same protocols and analyzed transient transformation mediated by Agrobacterium rhizogenes in DH4079 and DH12075 seedlings. Our results point to a genetic recalcitrance of the DH4079 line but not of the DH12075 line, which was possible to transform with both Agrobacterium species. These results make these B. napus lines convenient models to study the mechanisms of recalcitrance to Agrobacterium-mediated transformation in plants.
2. Results and Discussion
In this work, we used three protocols for stable transformation and regeneration (namely B, Z and A, Figure 1) previously described for B. napus with some modifications [21,24]. These protocols were assayed using the YC3.6-Bar, PM-YC3.6-LTI6b and pCRISPR plasmids.
Hypocotyl and cotyledon explants were excised from B. napus donor plants (Table 1). In total, we excised and cultured 1784 explants of the DH4079 line and 1077 explants of the DH12075 line, the latter of which was used as a reference (control) of a previously transformed B. napus line [27,28]. Each protocol was tested in the two B. napus lines, transforming them with different plasmids depending on the case. We first assessed the efficiency of plant regeneration of the three protocols used in this study (Table 1, “Plants/explant”), and then the number of regenerated explants successfully transformed (Table 1, “Positive plants/explant”). We found that the genotype and the plasmid type had no significant effect on the rate of plant regeneration, since none or very few regenerants were obtained from both lines and transformation events using protocols B and Z. However, all lines and transformation events produced regenerants with protocol A. Protocol A was found to be significantly better than protocol B in terms of plant regeneration, estimated as the number of regenerated plants per explant (Table 2). The reasons for such better performance are discussed next.
2.1. The Selective Agents Have a Critical Role in Plant Regeneration
Protocol B was the only protocol that did not produce any plant (either transformed or not) per explant, while protocols Z and A were able to regenerate plants from both DH4079 and DH12075 lines (Table 1). Regeneration was seriously compromised with protocol B. Interestingly, the only difference between protocols B and A is the time point of the addition of the selective agent to the culture medium (Figure 1). In protocol A, the selective agent was added to SOM medium, whereas in protocol B it was added earlier, to SRM medium, and this appeared to be crucial for the final fate of explants. We used a kanamycin concentration of 50 mg/L since it was previously described to combine moderate rates of escapes and regeneration for successful B. napus transformation [22]. When 50 mg/L of kanamycin was used in SRM medium (for explants transformed with the PM-YC3.6-LTI6b plasmid), few adventitious roots and leaves were produced from the calli after one month of culture. After two months, all the calli from both DH4079 and DH12075 lines turned creamy or brown and arrested their growth, showing clear signs of necrosis in some cases (Figure 2A,B). No developed shoots were observed in any case. When SRM medium was supplemented with 10 mg/L of BASTA (for explants transformed with the YC3.6-bar plasmid), the regeneration capacity of the calli was completely inhibited. Neither adventitious organs nor shoots emerged from the DH4079 or DH12075 calli and after two months of culture, all callus tissue was dead (Figure 2C,D). Thus, the use of selective agents in these conditions prevented shoot regeneration before callus death.
In protocol Z, cotyledons of the DH4079 and DH12075 lines were used as explants. With this protocol, the callus formation and growth were more limited than with protocol B, but clear differences in terms of regeneration were observed between explants exposed to different selective agents. After one month growing in selection medium with 50 mg/L of kanamycin, explants from the PM-YC3.6-LTI6b transformation developed the first shoots (Figure 3A) and after two months, resistant (green) and susceptible (purple) shoots were observed (Figure 3B). However, explants grown in selection medium supplemented with 5 mg/L of BASTA (for YC3.6-bar explants) did not develop any shoots. After one month of culture, visible signs of necrosis appeared in the explants (Figure 3C) and after two months, all cotyledon explants died (Figure 3D). Thus, the use of kanamycin allowed for plant regeneration from explants, whereas the use of BASTA promoted their death.
The conditions of use of the selective agent in culture media has been reported as one of the main factors affecting transformation efficiency [19]. Indeed, in the B. napus cv. Westar, an increase in selection conditions from 50 to 100 mg/L of kanamycin resulted in a reduction of non-transformed regenerated shoots (escape shoots), but the regeneration rate dropped down from 19% to 13% [22]. However, kanamycin concentration did not seem to be a problem in our case, since for protocol A, regenerants were obtained with this concentration in both protocols B and Z. Instead, the problem in protocol B seemed to be the time point of addition of the selective agent. A similar effect of kanamycin in plant regeneration has already been described in other species, such as grapevine [29], carrot [30] and cotton [31]. A similar scenario appears to occur with the use of BASTA in protocol B. However, BASTA was toxic in protocol Z. In general, herbicides have not been commonly used as selective agents as much as antibiotics due to the difficulty of establishing efficient concentrations which permit tissue regeneration and transformed plant selection [32], as it appears to occur in our B. napus explant transformations using BASTA in the selective medium. Such an inhibitory effect of BASTA in plant regeneration has also been described at even lower concentrations in watermelon [33] and peach [34]. Therefore, the addition of the selective agent (either kanamycin or BASTA) in the early steps of the protocol, when organogenesis is not yet initiated, results in an arrest of growth and organogenic differentiation, as well as the use of BASTA at both 5 (protocol Z) and 10 mg/L (protocol B).
2.2. The DH12075 Line, but Not DH4079, Can Be Genetically Transformed Using Protocol A
The protocol showing the highest plant-regeneration ratio was protocol A (Table 1 and Table 2), where inoculation consisted of the submersion of hypocotyl explants in the Agrobacterium suspension. With protocol A, calli were produced at the cuttings of hypocotyl explants after two weeks of culture (Figure 4A). Upon individualization of the calli, the first evidence of shoot formation was visible after approximately four weeks of culture (Figure 4B, arrowheads) and after six weeks, clearly visible, well-formed shoots were visible (Figure 4C, arrowheads).
Plants were produced from both DH4079 and DH12075 lines transformed with YC3.6-Bar, PM-YC3.6-LTI6b and pCRISPR. As opposed to the other protocols used, no differences were observed in terms of regeneration efficiency between the use of kanamycin and BASTA. After three weeks in SOM medium supplemented with carbenicillin and the corresponding selective agent, some plants turned whitish and stopped growing (Figure 4D, arrows), indicating sensitivity to the selective agent. Others were able to survive in the presence of the selective agent and kept growing green and vigorous, regenerating plantlets (Figure 4D, arrowheads). These results confirmed that it is possible to regenerate plantlets from B. napus DH4079 and DH12075 lines using protocol A, whose regeneration ability did not depend on the explant genotype or the type of selective agent used.
To assess whether regenerated green, growing plantlets incorporated the plasmid, they were genotyped by PCR with different primer pair combinations (see Materials and methods). None of the regenerated plantlets from the DH4079 line tested positive for PCR (Figure 4E), whereas few plantlets regenerated from the DH12075-excised explants were genotyped as positive (Figure 4F), being able to grow to fully regenerated transgenic plants (Figure 4G,H), which resulted in a calculated efficiency of 0.65% in terms of PCR-positive plants regenerated per explant. In all the transformation and regeneration events performed, there was a difference between the number of total and PCR-positive plants per explant, revealing the presence of escapes (Table 1), defined as plants resistant to selective agents but not transformed with the corresponding construct. This was particularly high in the case of the YC3.6-Bar experiments (2.48 plants per explant, but not a single PCR-positive plant). We speculate that this high occurrence of escapes could be due to not sufficiently restrictive concentrations of the corresponding selective agent, as also reported in other backgrounds [22]. Notwithstanding this, we were unable to identify any single transformed DH4079 regenerant, which confirms the extreme recalcitrance of this DH line. Together, these results show that the B. napus DH lines DH12075 and DH4079 exhibit a remarkably low transformation efficiency. In particular, the DH4079 line is recalcitrant to Agrobacterium-mediated transformation using the protocols described so far, including the modifications presented in this work.
Traditionally, the DH4079 background has been considered as recalcitrant for transformation [24]. Indeed, despite the enormous practical applicability that the development of DH4079 lines transformed with different genetic markers would have for the study of microspore embryogenesis, no DH4079 transgenic lines have been reported up to date. Out of the 1784 explants excised from DH4079 plants and transformed, we were not able to identify any single transgenic plant. This contrasts with the 13.4% of transformation efficiency previously reported for 62 DH4079 explants transformed and regenerated using the same protocol [24]. The reasons for such discrepancy are difficult to elucidate. Possible explanations could be the use of DH4079 plant material that was somehow different, or the residual presence of A. tumefaciens in the plant material used to analyze the efficiency of transformation. To avoid these potential problems, we are confident in using the highly embryogenic B. napus DH4079 line, since we routinely perform microspore cultures with microspores isolated from plants of this line [35] and confirm their high embryogenic response. Moreover, plant samples for PCR were excised from leaves of regenerated plants at the 2–3 true leaf stage upon plant acclimation, avoiding possible Agrobacterium dragging within the sample. We used the same protocol previously published by Maheshwari et al. [24] adapted to our experimental conditions, and obtained similar regeneration rates. Overall, we obtained with such protocol a transformation efficiency for the DH12075 line similar to that previously described for the same line [36]. Therefore, we strongly believe that our data (both the successful transformation of DH12075 and the unsuccessful transformation of DH4079) are robust and consistent. The remarkably different amounts of DH4079 explants transformed, cultured and analyzed in both cases (1784 in this work vs. 62 in Maheshwari et al. [24]) support this notion. Therefore, we postulate that the DH4079 line of B. napus is recalcitrant to A. tumefaciens-mediated genetic transformation.
2.3. The B. napus DH4079 Line Is Also Recalcitrant to Transient A. rhizogenes Transformation
Different methods of transient plant transformation have been tried in B. napus, including A. tumefaciens transformation of microspore-derived embryos [37] or microprojectile bombardment of isolated microspores combined with A. tumefaciens incubation [38], but not infection with Agrobacterium rhizogenes. A. rhizogenes, a gram-negative, soil-borne bacterium, naturally harbors large Ri plasmids that contain genes that favor infection of plant tissues and transference of their DNA into host plant cells [39]. However, as opposed to Ti plasmids from A. tumefaciens, Ri plasmids from A. rhizogenes induce the formation of hairy roots in the infected plant tissue [39]. Thus, genetic manipulation of these plasmids has allowed to establish protocols for transient plant transformation in different species, including B. napus [40]. We used this method to evaluate whether our B. napus lines show a response similar or different from that of A. tumefaciens stable transformation. We infected the hypocotyls of one-week-old entire plants of both DH12075 and DH4079 lines with A. rhizogenes and analyzed the occurrence of morphogenic processes as a consequence of infection after 3–4 weeks. In DH12075, we found plants with no specific response to infection, plants producing a callus at the site of wounding for infection, and plants developing large hairy roots at the site of infection, whereas DH4079 only presented plants with no specific response or with callus production (Figure 5A). The percentage of plants producing calli was similar for both lines, ranging between ~10–15% (Figure 5B), which may indicate a plant response to wounding rather than to bacterial infection. However, there was a remarkable difference in root production upon infection, which revolved around 50% for DH12075, but was null for DH4079 (Figure 5B). These results confirmed the recalcitrance of DH4079 to Agrobacterium transformation, not only with A. tumefaciens but also with A. rhizogenes.
A recent work reported the formation of hairy roots from both DH12075 and DH4079 when infected with a modified A. tumefaciens strain carrying a Ri hairy-root-inducing plasmid [41]. Hairy roots developed from 97% and 42% of the infected DH12075 and DH4079 seedlings, respectively, but transgenic plants were only possible to regenerate from transformed and cultured DH12075 roots [41]. This transformation approach using engineered A. tumefaciens with Ri plasmid is essentially different from the protocols used in the present work and may open an alternative approach to produce transgenic B. napus lines upon plant regeneration from transformed roots in different genotypes, including DH12075. However, it also failed in achieving stable transformation of DH4079. These observations, together with the work presented in this manuscript, support the notion that the DH4079 line is extremely recalcitrant to Agrobacterium transformation.
2.4. A Possible Relationship between Recalcitrance to Genetic Transformation and Doubled Haploidy?
Within the Brassica genus, efficiencies of stable transformation are remarkably variable, ranging between 0.59–1.56% for wucai (B. campestris) [42], 2.2–10.83% for Chinese cabbage (B. rapa ssp. Pekinensis) [43,44], 2.7–6.4% for broccoli (B. oleracea var. italica) [45], ~7% for B. juncea [46], 7–13.6% for B. oleraceae var Botrytis [47,48] and as high as 32.5–45% for cabbage (B. oleracea subsp. capitata), depending on the explant type (shoot tips and hypocotyls, respectively) [49]. As with B. napus, the efficiency reported by different works was highly dependent on the genetic background used. In commercial cultivars such as Oscar and RK7, an efficiency of up to 67% was reported [50]. In cv. Westar, a range of transformation efficiency of 7–33%, depending on in vitro conditions, was reported [22,50]. Interestingly, when fully homozygous DH plants derived from Westar were used in similar experimental conditions, the transformation efficiency dropped down to 0.3–3% [36]. Whether these differences are due to different specific experimental conditions, or to allele fixation derived from the process of chromosome doubling inherent to DH production [6], is not known. However, the transformation efficiency reported in our work for DH12075, a Westar-derived DH line, was 0.65%, which fits within the range of efficiencies reported for DH backgrounds [36]. This supports the hypothesis that recalcitrance to transformation could be influenced by the degree of gene fixation in a partially allogamous species such as B. napus.
Finally, it is also worth mentioning that DH4079, in addition to being recalcitrant to transformation, is one of the backgrounds most responsive to induction of microspore embryogenesis for DH production [11]. In parallel, some of the B. napus backgrounds where genetic transformation has been proven efficient or at least possible, such as Westar and DH12075 [41], are also known to show a very low or null response to induction of microspore embryogenesis [51,52]. Interestingly, it is also known that Arabidopsis thaliana, a model species for genetic studies where genetic transformation is very well developed and widely used for decades, is extremely recalcitrant to microspore embryogenesis, with no successful reports of induction of such morphogenic process published to date. The scenario in a tomato plant, another plant model species, is similar: transformation has been successfully achieved, but its extreme recalcitrance to microspore embryogenesis is widely acknowledged [53,54,55]. Thus, it is tempting to speculate about a possible inverse relationship between the ability for genetic transformation and the response to induction of microspore embryogenesis. This could be an interesting hypothesis to elucidate in future research.
3. Materials and Methods
3.1. Plant Material
Seedlings from two different B. napus DH lines were used as donors of explants: the DH4079 line, derived from cv. Topas [11], and the DH12075 line, derived from a cross between cvs. Westar × Cresor.
3.2. Constructs
Three plasmids were used in this experiment (Figure 6A). On the one hand, we used YC3.6 and PM-YC3.6-LTI6b, two cameleon calcium sensor constructs that confer BASTA and kanamycin resistance, respectively, kindly provided by Dr. Jörg Kudla [56]. On the other hand, we used pCRISPR, a GoldenBraid-based construction for CRISPR/Cas9 genome editing [57] containing a CaMV 35S promoter, Cas9 CDS, and Nos Terminator cassette, assembled in the pDGB3-alpha1 binary plasmid containing a cassette for kanamycin for plant selection.
3.3. Management and Transformation of Escherichia coli and Agrobacterium Strains
Chemically competent Escherichia coli One Shot™ ccdB Survival™ 2 T1R cells were prepared from cells growing under agitation and 37 °C in 50 mL Psi medium (Table 3) until the DO600 reached 0.4–0.5. Bacterial culture was cooled down and centrifuged (5 min, 5000× g, 4 °C). The pellet was resuspended in 20 mL of cold Transformation Buffer I (TBI; Table 3) and incubated on ice for 15 min. Next, the bacterial suspension was centrifuged (10 min, 2000× g, 4 °C) and the resulting pellet was resuspended in 2 mL of Transformation Buffer II (TBII; Table 3) and incubated in ice for 15 min. Finally, 50 µL aliquots were kept at −80 °C. For transformation, aliquots were thawed and 5 µL (~1 µg) of DNA was added. Tubes were incubated for 60 min on ice, 30 s at 42 °C and then on ice for 2 min. Next, 250 µL of autoclaved SOC medium (Table 3) were added and tubes were incubated at 37 °C for 1 h in a shaker at 225 rpm. From each tube, 25–100 µL were plated in commercial LB-Agar Miller medium (VWR Life Science) plates supplemented with the corresponding bacterial selective agents (Table 4) and incubated at 37 °C. Saturated E. coli liquid LB cultures were used to perform plasmid extractions with the NZYMiniprep kit (NZYtech).
Agrobacterium tumefaciens strain LBA4404 carrying the helper vir plasmid pAL4404 (ElectroMAX™ A. tumefaciens LBA4404 Cells, Invitrogen™, Waltham, MA, USA) was used for plant transformations. Electrocompetent A. tumefaciens cells were prepared by culturing a single colony in liquid YM medium (Table 3) for 2 days at 28–30 °C. One mL of culture was diluted 1:100 in liquid YM medium. The dilution was incubated at 28–30 °C in agitation until DO600 ranged 0.5–0.7. The Agrobacterium culture was centrifuged (15 min, 4 °C, 6000× g) and the pellet was washed with 30 mL of cold sterile MilliQ water. Centrifugation (same conditions) and washing was repeated twice. Finally, cells were resuspended in 1 mL of cold 10% (v/v) glycerol and 20 µL aliquots were stored at −80 °C. For transformation of A. tumefaciens LBA4404 competent cells, each aliquot was thawed, 1 µL (~200 ng) of plasmid was added and the mixture was pipetted into the electroporation cuvette. The mixture was electroporated at 2.0 kV, 200 Ω y 25 µF using an Electro Cell Manipulator 600 electroporator from BTX (Holliston, MA, USA). After electroporation, 1 mL of autoclaved liquid YM medium was added to the micro tube. The content was transferred to a 15 mL tube and incubated at 225 rpm and 28 °C for 3 h. Then, 50–100 µL were plated in solid YM (Table 3) medium with the corresponding plasmid-specific selective agents (Table 4). Plates were incubated at 28 °C for 2–3 days until colonies were observed. Preparation of Agrobacterium rhizogenes strain 15834 competent cells and transformation with pCRISPR by electroporation was performed as described above for A. tumefaciens, but using MGL liquid medium (Table 3).
3.4. Explant Stable Transformation and Plant Regeneration
Three A. tumefaciens stable transformation protocols were assayed (Figure 1), two (B and A) based on [24] and one (protocol Z) based on [21]. For all protocols, bacterial inoculums were prepared by centrifuging (5975× g, 10 h, 4 °C) liquid cultures grown at 28 °C in liquid YM medium (Table 3) supplemented with 50 mg/L rifampicin and appropriate bacterial selective agents (Table 4) until reaching an approximate OD600 = 1.0. The bacterial pellets were resuspended in liquid MS medium with 1% sucrose until reaching a OD600 ranging 0.15–0.2, and 100 µM acetosyringone was added. To produce the explant-donor seedlings, B. napus seeds of the DH4079 and DH12075 lines were surface-sterilized for 15 min in a 70% ethanol +0.1% Triton X-100 solution, followed by 15 min in 10% bleach solution and three rinses with distilled water. Seeds were then germinated in Germination Medium (GM; Table 5).
In protocols A and B, hypocotyl explants of 0.5–1 cm were excised from 7-day-old seedlings and pre-cultured in Callus Induction Medium (CIM; Table 5) for 5 days. After 3–5 days, explants were inoculated with the corresponding A. tumefaciens culture. Pre-cultured explants were submerged in the bacterial suspension, blotted on sterile paper and transferred to CIM medium for 72 h at 25 °C under reduced light conditions by covering the culture with a piece of white paper to reduce light intensity to dim light. After co-cultivation with A. tumefaciens, explants were transferred to CIM medium supplemented with 500 mg/L of carbenicillin and cultured for 2 weeks at 25 °C under dim light. The regenerated calli were cut into pieces and cultured for 4–6 weeks in Shoot Regeneration Medium (SRM; Table 5) for protocol A, and for protocol B in SRM medium supplemented with 50 mg/L of kanamycin as plant selection agent for PM-YC3.6-LTI6b transformation and 10 mg/L of BASTA (amonium glufosinate) for YC3.6-Bar. The developing shoot buds were transferred to Shoot Outgrowth Medium (SOM; Table 5) supplemented with plant selection agents as described above. Shoots longer than 2 cm were transferred to Root Regeneration Medium (RRM; Table 5) supplemented with plant selection agents as described above.
For protocol Z, cotyledon explants were obtained by cutting the petiole base from 5- to 7-day-old seedlings. Excised cotyledons were inoculated by dipping the petiole base for 3–5 s in liquid MS + V medium + 1% sucrose and the A. tumefaciens preparation (OD600 = 0.2 supplemented with 100 µM acetosyringone). Then, explants were inoculated by inserting 1–2 mm of the petiole base in solid Co-Cultivation Medium (CCM; Table 5). Plates were kept at 25 °C for 72 h under dim light. Then, explants were transferred to Selection Medium (SM; Table 5) supplemented with plant selection agents (15 mg/L of kanamycin for PM-YC3.6-LTI6b and CRISPRp transformations and 5 mg/L of BASTA for YC3.6-Bar) for at least four weeks, subculturing them to fresh SM every week. The regenerating shoots produced were transferred to Rooting Medium (RM; Table 5) supplemented with plant selection agents as described above.
For genotyping of regenerated plants, when putative transgenic plants developed 2–3 true leaves, 1 cm2 pieces of leaf tissue were excised and DNA was extracted on TNES extraction buffer (100 mM Tris Buffer, 100 mM EDTA, 250 mM NaCl, pH 8) and precipitated with 1 volume of isopropanol at room temperature for 2 min. The pellet was resuspended in 50 µL MilliQ water. The primer pairs used to genotype regenerated plants from transformations with PM-YC3.6-LTI6b, YC3.6-Bar and pCRISPR are shown in Figure 6B, and their respective sequences in Table 6. All PCRs were performed by adding 1 µM primers to the Mix MZY Taq II 2x Green Master Mix (NZYtech, Lisbon, Portugal), with annealing temperature of 59 °C and extension time of 40 s.
3.5. A. rhizogenes-Mediated Transient Transformation
For transient hairy root transformation of B. napus with Agrobacterium rhizogenes, the protocol was adapted from Ron et al. [26]. Briefly, 3 mL of saturated A. rhizogenes culture on MGL liquid medium supplemented with kanamycin (50 mg/mL) was centrifuged at 5000× g, 4 °C for 15 min, and the obtained pellet was resuspended in 200 µL of fresh MGL liquid medium. The resulting concentrated bacterial suspension was used to infect 10-day-old B. napus seedlings sowed and grown as described above and cultured on solid MS media (0.8% plant agar) under sterile conditions. For A. rhizogenes infection, a sterile needle tip was impregnated with the concentrated bacterial suspension, and two soft, little wounds were made along the stem of each seedling. Plantlets were then cultured in vitro for 3–4 additional weeks under the same conditions, and the numbers of roots produced were then registered.
3.6. Statistical Analysis
Plant regeneration data were analyzed using the StatGraphics software, v18.1.13. The Kruskal–Wallis test with p ≤ 0.05 was used to detect statistically significant differences in plant regeneration and transformation rates.
Author Contributions
Conceptualization: J.M.S.-S. and R.M.; funding acquisition: J.M.S.-S. and R.M.; investigation: A.C.-S., R.M. and R.P.; methodology: A.C.-S. and R.M.; data curation: A.C.-S. and R.M.; formal analysis: A.C.-S., R.M. and J.M.S.-S.; project administration: J.M.S.-S. and R.M.; supervision: J.M.S.-S. and R.M.; writing—original draft preparation: A.C.-S., J.M.S.-S. and R.M.; writing—review and editing: A.C.-S., J.M.S.-S. and R.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by MCIN/AEI/10.13039/501100011033, grant number PID2020-115763RB-I00 to JMSS and by Generalitat Valenciana, grant number CDEIGENT 2018/023 to RM. ACS is recipient of a PhD contract (FPU17/00715) from the Spanish Ministerio de Educación.
Data Availability Statement
All data supporting the findings of this study are available within the paper.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- van Montagu, M. It Is a Long Way to GM Agriculture. Ann. Rev. Plant. Biol. 2011, 62, 1–23. [Google Scholar] [CrossRef]
- Klümper, W.; Qaim, M. A Meta-Analysis of the Impacts of Genetically Modified Crops. PLoS ONE 2014, 9, e111629. [Google Scholar] [CrossRef]
- ISAAA. GM Approval Database. Available online: https://www.isaaa.org/gmapprovaldatabase (accessed on 23 May 2022).
- Hua, K.; Zhang, J.; Botella, J.R.; Ma, C.; Kong, F.; Liu, B.; Zhu, J.-K. Perspectives on the Application of Genome-Editing Technologies in Crop Breeding. Mol. Plant. 2019, 12, 1047–1059. [Google Scholar] [CrossRef]
- Hopp, H.E.; Spangenberg, G.; Herrera-Estrella, L. Editorial: Plant Transformation. Front. Plant Sci. 2022, 13, 876671. [Google Scholar] [CrossRef] [PubMed]
- Seguí-Simarro, J.M.; Jacquier, N.M.A.; Widiez, T. Overview of in vitro and in vivo doubled haploid technologies. In Doubled Haploid Technology, 1st ed.; Seguí-Simarro, J.M., Walker, J.M., Eds.; Methods in Molecular Biology; Springer Science + Business Media, LLC: New York, NY, USA, 2021; Volume 1, pp. 3–22. [Google Scholar]
- Jouannic, S.; Champion, A.; Seguí-Simarro, J.M.; Salimova, E.; Picaud, A.; Tregear, J.; Testillano, P.; Risueño, M.C.; Simanis, V.; Kreis, M.; et al. The protein kinases AtMAP3Kε1 and BnMAP3Kε1 are functional homologues of S. pombe cdc7p and may be involved in cell division. Plant J. 2001, 26, 637–649. [Google Scholar] [CrossRef] [PubMed]
- Satpute, G.; Long, H.; Seguí-Simarro, J.M.; Risueño, M.C.; Testillano, P.S. Cell architecture during gametophytic and embryogenic microspore development in Brassica napus. Acta Physiol. Plant. 2005, 27, 665–674. [Google Scholar] [CrossRef]
- Weyen, J. Applications of doubled haploids in plant breeding and applied research. In Doubled Haploid Technology, 1st ed.; Seguí-Simarro, J.M., Walker, J.M., Eds.; Methods in Molecular Biology; Humana Press: New York, NY, USA, 2021; Volume 1, pp. 23–39. [Google Scholar]
- Custers, J.B.M.; Cordewener, J.H.G.; Fiers, M.A.; Maassen, B.T.H.; van Lookeren-Campagne, M.M.; Liu, C.M. Androgenesis in Brassica: A model system to study the initiation of plant embryogenesis. In Current Trends in the Embryology of Angiosperm; Bhojwani, S.S., Soh, W.Y., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2001; pp. 451–470. [Google Scholar]
- Ferrie, A.; Möllers, C. Haploids and doubled haploids in Brassica spp. for genetic and genomic research. Plant Cell Tissue Organ Cult. 2011, 104, 375–386. [Google Scholar] [CrossRef]
- Rivas-Sendra, A.; Corral-Martínez, P.; Porcel, R.; Camacho-Fernández, C.; Calabuig-Serna, A.; Seguí-Simarro, J.M. Embryogenic competence of microspores is associated with their ability to form a callosic, osmoprotective subintinal layer. J. Exp. Bot. 2019, 70, 1267–1281. [Google Scholar] [CrossRef]
- Rivas-Sendra, A.; Calabuig-Serna, A.; Seguí-Simarro, J.M. Dynamics of calcium during in vitro microspore embryogenesis and in vivo microspore development in Brassica napus and Solanum melongena. Front. Plant Sci. 2017, 8, 1177. [Google Scholar] [CrossRef]
- Parra-Vega, V.; Corral-Martínez, P.; Rivas-Sendra, A.; Seguí-Simarro, J.M. Induction of embryogenesis in Brassica napus microspores produces a callosic subintinal layer and abnormal cell walls with altered levels of callose and cellulose. Front. Plant Sci. 2015, 6, 1018. [Google Scholar] [CrossRef]
- Parra-Vega, V.; Corral-Martínez, P.; Rivas-Sendra, A.; Seguí-Simarro, J.M. Formation and excretion of autophagic plastids (plastolysomes) in Brassica napus embryogenic microspores. Front. Plant Sci. 2015, 6, 94. [Google Scholar] [CrossRef]
- Corral-Martínez, P.; Parra-Vega, V.; Seguí-Simarro, J.M. Novel features of Brassica napus embryogenic microspores revealed by high pressure freezing and freeze substitution: Evidence for massive autophagy and excretion-based cytoplasmic cleaning. J. Exp. Bot. 2013, 64, 3061–3075. [Google Scholar] [CrossRef] [PubMed]
- Corral-Martínez, P.; Camacho-Fernández, C.; Mir, R.; Seguí-Simarro, J.M. Doubled haploid production in high- and low-response genotypes of rapeseed (Brassica napus) through isolated microspore culture. In Doubled Haploid Technology, 1st ed.; Seguí-Simarro, J.M., Walker, J.M., Eds.; Methods in Molecular Biology; Springer Science + Business Media, LLC: New York, NY, USA, 2021; Volume 2, pp. 129–144. [Google Scholar]
- Dai, C.; Li, Y.; Li, L.; Du, Z.; Lin, S.; Tian, X.; Li, S.; Yang, B.; Yao, W.; Wang, J. An efficient Agrobacterium-mediated transformation method using hypocotyl as explants for Brassica napus. Mol. Breed. 2020, 40, 1–13. [Google Scholar] [CrossRef]
- Bhalla, P.L.; Singh, M.B. Agrobacterium-mediated transformation of Brassica napus and Brassica oleracea. Nat. Protoc. 2008, 3, 181–189. [Google Scholar] [CrossRef] [PubMed]
- Moloney, M.M.; Walker, J.M.; Sharma, K.K. High efficiency transformation of Brassica napus using Agrobacterium vectors. Plant Cell Rep. 1989, 8, 238–242. [Google Scholar] [CrossRef]
- Sparrow, P.A.C.; Dale, P.J.; Irwin, J.A. Brassica oleracea. In Agrobacterium Protocols; Wang, K., Ed.; Humana Press: Totowa, NJ, USA, 2006; pp. 417–426. [Google Scholar]
- Fry, J.; Barnason, A.; Horsch, R.B. Transformation of Brassica napus with Agrobacterium tumefaciens based vectors. Plant Cell Rep. 1987, 6, 321–325. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Sonntag, K.; Rudloff, E.; Han, J. Production of fertile transgenic Brassica napus by Agrobacterium-mediated transformation of protoplasts. Plant Breed. 2005, 124, 1–4. [Google Scholar] [CrossRef]
- Maheshwari, P.; Selvaraj, G.; Kovalchuk, I. Optimization of Brassica napus (canola) explant regeneration for genetic transformation. New Biotechnol. 2011, 29, 144–155. [Google Scholar] [CrossRef]
- Gocal, G.F. Gene editing in Brassica napus for basic research and trait development. Vitr. Cell. Dev. Biol.-Plant 2021, 57, 731–748. [Google Scholar] [CrossRef]
- Sheng, X.; Yu, H.; Wang, J.; Shen, Y.; Gu, H. Establishment of a stable, effective and universal genetic transformation technique in the diverse species of Brassica oleracea. Front. Plant Sci. 2022, 13, 1021669. [Google Scholar] [CrossRef]
- Wu, L.; El-Mezawy, A.; Shah, S. A seed coat outer integument-specific promoter for Brassica napus. Plant Cell Rep. 2011, 30, 75–80. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Beaith, M.; Chalifoux, M.; Ying, J.; Uchacz, T.; Sarvas, C.; Griffiths, R.; Kuzma, M.; Wan, J.; Huang, Y. Shoot-specific down-regulation of protein farnesyltransferase (α-subunit) for yield protection against drought in canola. Mol. Plant. 2009, 2, 191–200. [Google Scholar] [CrossRef]
- Colby, S.M.; Meredith, C.P. Kanamycin sensitivity of cultured tissues of Vitis. Plant Cell Rep. 1990, 9, 237–240. [Google Scholar] [CrossRef]
- Hardegger, M.; Sturm, A. Transformation and regeneration of carrot (Daucus carota L.). Mol. Breed. 1998, 4, 119–127. [Google Scholar] [CrossRef]
- Zhang, B.-H.; Liu, F.; Liu, Z.-H.; Wang, H.-M.; Yao, C.-B. Effects of kanamycin on tissue culture and somatic embryogenesis in cotton. Plant Growth Regul. 2001, 33, 137–149. [Google Scholar] [CrossRef]
- Khuong, T.T.H.; Crété, P.; Robaglia, C.; Caffarri, S. Optimisation of tomato Micro-tom regeneration and selection on glufosinate/Basta and dependency of gene silencing on transgene copy number. Plant Cell Rep. 2013, 32, 1441–1454. [Google Scholar] [CrossRef]
- Ganasan, K.; Huyop, F. The sensitivity of plant tissue culture and plant cell of Citrullus lanatus cv. Round dragon against BASTA. Int. J. Agric. Res. 2010, 5, 11–18. [Google Scholar] [CrossRef]
- Ricci, A.; Sabbadini, S.; Prieto, H.; Padilla, I.M.; Dardick, C.; Li, Z.; Scorza, R.; Limera, C.; Mezzetti, B.; Perez-Jimenez, M. Genetic transformation in peach (Prunus persica L.): Challenges and ways forward. Plants 2020, 9, 971. [Google Scholar] [CrossRef]
- Camacho-Fernández, C.; Seguí-Simarro, J.M.; Mir, R.; Boutilier, K.; Corral-Martínez, P. Cell Wall Composition and Structure Define the Developmental Fate of Embryogenic Microspores in Brassica napus. Front. Plant Sci. 2021, 12, 737139. [Google Scholar] [CrossRef]
- Kazan, K.; Curtis, M.D.; Goulter, K.C.; Manners, J.M. Agrobacterium tumefaciens-mediated transformation of double haploid canola (Brassica napus) lines. Funct. Plant Biol. 1997, 24, 97–102. [Google Scholar] [CrossRef]
- Dubas, E.; Moravčíková, J.; Libantová, J.; Matušíková, I.; Benková, E.; Żur, I.; Krzewska, M. The influence of heat stress on auxin distribution in transgenic B. napus microspores and microspore-derived embryos. Protoplasma 2014, 251, 1077–1087. [Google Scholar] [CrossRef] [PubMed]
- Abdollahi, M.R.; Corral-Martinez, P.; Mousavi, A.; Salmanian, A.H.; Moieni, A.; Seguí-Simarro, J.M. An efficient method for transformation of pre-androgenic, isolated Brassica napus microspores involving microprojectile bombardment and Agrobacterium-mediated transformation. Acta Physiol. Plant. 2009, 31, 1313–1317. [Google Scholar] [CrossRef]
- Moore, L.; Warren, G.; Strobel, G. Involvement of a plasmid in the hairy root disease of plants caused by Agrobacterium rhizogenes. Plasmid 1979, 2, 617–626. [Google Scholar] [CrossRef]
- Christey, M.C.; Sinclair, B.K. Regeneration of transgenic kale (Brassica oleracea var. acephala), rape (B. napus) and turnip (B. campestris var. rapifera) plants via Agrobacterium rhizogenes mediated transformation. Plant Sci. 1992, 87, 161–169. [Google Scholar] [CrossRef]
- Jedličková, V.; Mácová, K.; Štefková, M.; Butula, J.; Staveníková, J.; Sedláček, M.; Robert, H.S. Hairy root transformation system as a tool for CRISPR/Cas9-directed genome editing in oilseed rape (Brassica napus). Front. Plant Sci. 2022, 13, 919290. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Zeng, F.; Wang, J.; Ye, X.; Zhu, S.; Yuan, L.; Hou, J.; Wang, C. Transgenic Wucai (Brassica campestris L.) produced via Agrobacterium-mediated anther transformation in planta. Plant Cell Rep. 2019, 38, 577–586. [Google Scholar] [CrossRef]
- Park, B.-J.; Liu, Z.; Kanno, A.; Kameya, T. Genetic improvement of Chinese cabbage for salt and drought tolerance by constitutive expression of a B. napus LEA gene. Plant Sci. 2005, 169, 553–558. [Google Scholar] [CrossRef]
- Li, X.; Li, H.; Zhao, Y.; Zong, P.; Zhan, Z.; Piao, Z. Establishment of a simple and efficient Agrobacterium-mediated genetic transformation system to Chinese Cabbage (Brassica rapa L. ssp. pekinensis). Hortic. Plant J. 2021, 7, 117–128. [Google Scholar] [CrossRef]
- Chen, L.-F.O.; Hwang, J.-Y.; Charng, Y.-Y.; Sun, C.-W.; Yang, S.-F. Transformation of broccoli (Brassica oleracea var. italica) with isopentenyltransferase gene via Agrobacterium tumefaciens for post-harvest yellowing retardation. Mol. Breed. 2001, 7, 243–257. [Google Scholar] [CrossRef]
- Naeem, I.; Munir, I.; Durrett, T.P.; Iqbal, A.; Aulakh, K.S.; Ahmad, M.A.; Khan, H.; Khan, I.A.; Hussain, F.; Shuaib, M. Feasible regeneration and agro bacterium-mediated transformation of Brassica juncea with Euonymus alatus diacylglycerol acetyltransferase (EaDAcT) gene. Saudi J. Biol. Sci. 2020, 27, 1324–1332. [Google Scholar] [CrossRef]
- Bhalla, P.L.; Smith, N. Agrobacterium tumefaciens-mediated transformation of cauliflower, Brassica oleracea var. botrytis. Mol. Breed. 1998, 4, 531–541. [Google Scholar] [CrossRef]
- Chakrabarty, R.; Viswakarma, N.; Bhat, S.; Kirti, P.; Singh, B.; Chopra, V. Agrobacterium-mediated transformation of cauliflower: Optimization of protocol and development of Bt-transgenic cauliflower. J. Biosci. 2002, 27, 495–502. [Google Scholar] [CrossRef] [PubMed]
- Rafat, A.; Abd Aziz, M.; Abd Rashid, A.; Abdullah, S.N.A.; Kamaladini, H.; Sirchi, M.T.; Javadi, M. Optimization of Agrobacterium tumefaciens-mediated transformation and shoot regeneration after co-cultivation of cabbage (Brassica oleracea subsp. capitata) cv. KY Cross with AtHSP101 gene. Sci. Hort. 2010, 124, 1–8. [Google Scholar] [CrossRef]
- Zhang, Y.; Singh, M.B.; Swoboda, I.; Bhalla, P.L. Agrobacterium-mediated transformation and generation of male sterile lines of Australian canola. Aust. J. Agric. Res. 2005, 56, 353–361. [Google Scholar] [CrossRef]
- Malik, M.R.; Wang, F.; Dirpaul, J.; Zhou, N.; Hammerlindl, J.; Keller, W.; Abrams, S.R.; Ferrie, A.M.R.; Krochko, J.E. Isolation of an embryogenic line from non-embryogenic Brassica napus cv. Westar through microspore embryogenesis. J. Exp. Bot. 2008, 59, 2857–2873. [Google Scholar] [CrossRef]
- Corral-Martinez, P.; Siemons, C.; Horstman, A.; Angenent, G.C.; de Ruijter, N.; Boutilier, K. Live Imaging of embryogenic structures in Brassica napus microspore embryo cultures highlights the developmental plasticity of induced totipotent cells. Plant Reprod. 2020, 33, 143–158. [Google Scholar] [CrossRef]
- Seguí-Simarro, J.M.; Nuez, F. Embryogenesis induction, callogenesis, and plant regeneration by in vitro culture of tomato isolated microspores and whole anthers. J. Exp. Bot. 2007, 58, 1119–1132. [Google Scholar] [CrossRef]
- Seguí-Simarro, J.M. Androgenesis in solanaceae. In In Vitro Embryogenesis; Germanà, M.A., Lambardi, M., Eds.; Methods in Molecular Biology; Springer Science + Business Media: New York, NY, USA, 2016; Volume 1359, pp. 209–244. [Google Scholar]
- Seguí-Simarro, J.M.; Nuez, F. Meiotic metaphase I to telophase II is the most responsive stage of microspore development for induction of androgenesis in tomato (Solanum lycopersicum). Acta Physiol. Plant. 2005, 27, 675–685. [Google Scholar] [CrossRef]
- Krebs, M.; Held, K.; Binder, A.; Hashimoto, K.; Den Herder, G.; Parniske, M.; Kudla, J.; Schumacher, K. FRET-based genetically encoded sensors allow high-resolution live cell imaging of Ca2+ dynamics. Plant J. 2012, 69, 181–192. [Google Scholar] [CrossRef]
- Vazquez-Vilar, M.; Bernabé-Orts, J.M.; Fernandez-Del-Carmen, A.; Ziarsolo, P.; Blanca, J.; Granell, A.; Orzaez, D. A modular toolbox for gRNA-Cas9 genome engineering in plants based on the GoldenBraid standard. Plant Methods 2016, 12, 10. [Google Scholar] [CrossRef]
Figure 1.
Scheme of the different steps of the three transformation protocols (B, A and Z) used in this work. See text for further details.
Figure 1.
Scheme of the different steps of the three transformation protocols (B, A and Z) used in this work. See text for further details.

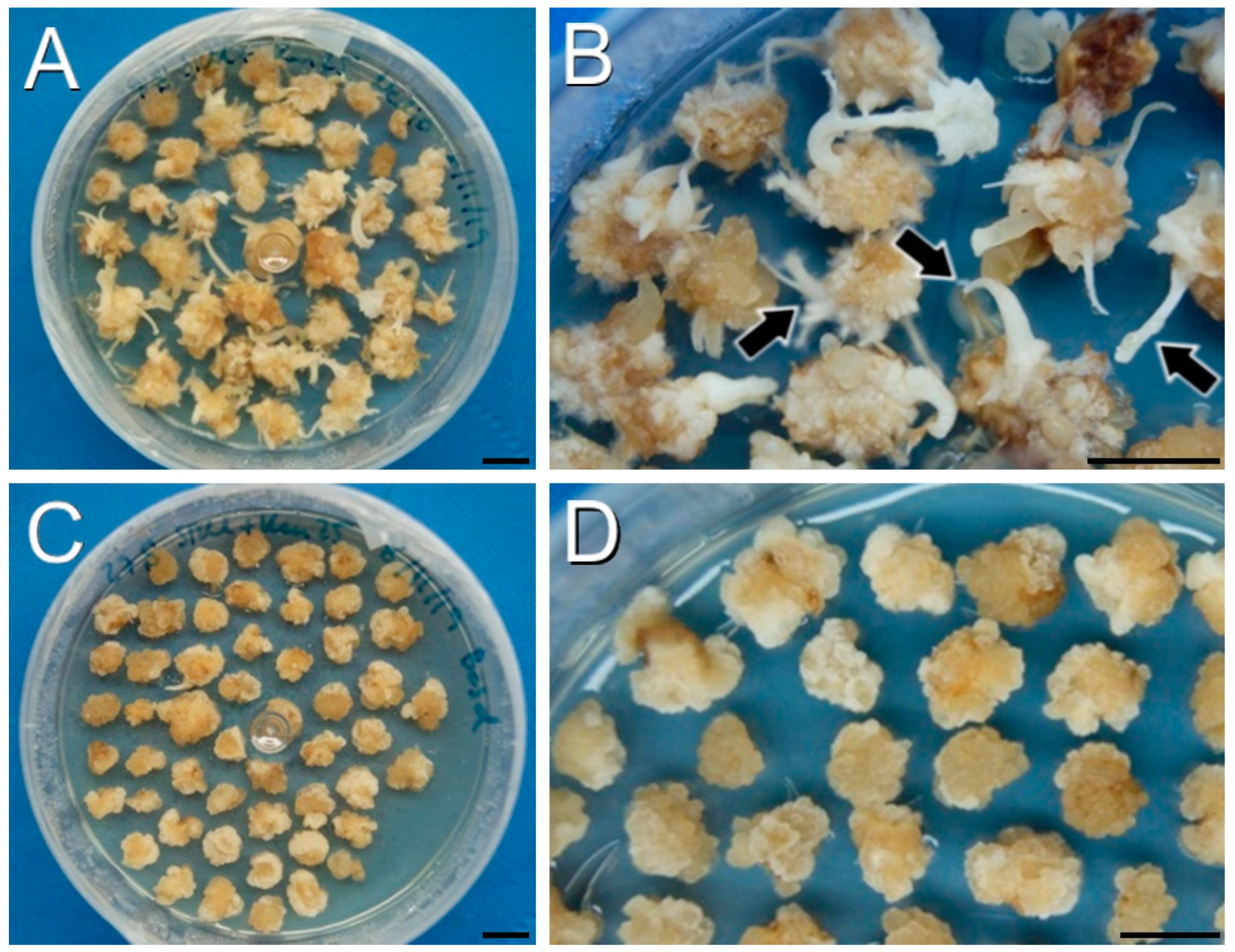
Figure 2.
Callus production and growth using protocol B after one month. (A,B): DH4079 calli in SRM supplemented with 50 mg/L of kanamycin. Note the presence of adventitious organs (arrowheads in (B) in some of these calli. (C,D): DH12075 calli in SRM supplemented with 10 mg/L of BASTA. (D): Amplified image where the absence of regeneration is clear. Bars: 1 cm.
Figure 2.
Callus production and growth using protocol B after one month. (A,B): DH4079 calli in SRM supplemented with 50 mg/L of kanamycin. Note the presence of adventitious organs (arrowheads in (B) in some of these calli. (C,D): DH12075 calli in SRM supplemented with 10 mg/L of BASTA. (D): Amplified image where the absence of regeneration is clear. Bars: 1 cm.

Figure 3.
Callus production and growth using protocol Z. (A): Cotyledons after one month in selection medium (50 mg/L of kanamycin) showing a susceptible, purple shoot (arrow). (B): Cotyledons after two months in selection medium (50 mg/L kanamycin) showing resistant, green shoots (arrowhead) and susceptible, purple shoots (arrow). (C,D): Cotyledons after one (C) and two months (D) in selection medium with 5 mg/L of BASTA. Bars: 1 cm.
Figure 3.
Callus production and growth using protocol Z. (A): Cotyledons after one month in selection medium (50 mg/L of kanamycin) showing a susceptible, purple shoot (arrow). (B): Cotyledons after two months in selection medium (50 mg/L kanamycin) showing resistant, green shoots (arrowhead) and susceptible, purple shoots (arrow). (C,D): Cotyledons after one (C) and two months (D) in selection medium with 5 mg/L of BASTA. Bars: 1 cm.

Figure 4.
Callus production and plant regeneration using protocol A. (A): Explants after 2 weeks in CIM + carbenicillin. (B): Calli after 4 weeks in SRM + carbenicillin. Note the occurrence of some green shoots on the callus surface (arrowheads). (C): Shoots (arrowheads) emerging from calli after 6 weeks in SRM + carbenicillin. (D): Green, regenerated plantlets resistant to the selective agent (black arrowheads) and white, arrested shoots sensitive to the selective agent (white arrows) after three weeks in SOM + carbenicillin + kanamycin. (E,F): PCR genotyping of green, transformed plants of the DH4079 (E) and DH12075 lines (F), using YC3.6 Fw and GFP 5′ Rv primers. Plasmid DNA was used as PCR positive control (+) and water was used as negative control (-). Note the presence of a PCR-positive DNA sample in (F), lane 5. (G,H): Green regenerated transgenic in vitro (G) and acclimated (H) plant of the DH12075 line. Bars: 1 cm.
Figure 4.
Callus production and plant regeneration using protocol A. (A): Explants after 2 weeks in CIM + carbenicillin. (B): Calli after 4 weeks in SRM + carbenicillin. Note the occurrence of some green shoots on the callus surface (arrowheads). (C): Shoots (arrowheads) emerging from calli after 6 weeks in SRM + carbenicillin. (D): Green, regenerated plantlets resistant to the selective agent (black arrowheads) and white, arrested shoots sensitive to the selective agent (white arrows) after three weeks in SOM + carbenicillin + kanamycin. (E,F): PCR genotyping of green, transformed plants of the DH4079 (E) and DH12075 lines (F), using YC3.6 Fw and GFP 5′ Rv primers. Plasmid DNA was used as PCR positive control (+) and water was used as negative control (-). Note the presence of a PCR-positive DNA sample in (F), lane 5. (G,H): Green regenerated transgenic in vitro (G) and acclimated (H) plant of the DH12075 line. Bars: 1 cm.

Figure 5.
Response of B. napus DH12075 and DH4079 to transient A. rhizogenes infection. (A): The infected DH12075 plants (left) show in some cases no specific response, and in others callus formation (arrowhead) or formation of adventitious roots (arrow). In contrast, DH4079 plants (right) only show either no specific response in some plants or just callus formation (arrowhead) in others. (B): Comparison of the percentages of root and callus formation in DH12075 and DH4079. Bar: 1 cm.
Figure 5.
Response of B. napus DH12075 and DH4079 to transient A. rhizogenes infection. (A): The infected DH12075 plants (left) show in some cases no specific response, and in others callus formation (arrowhead) or formation of adventitious roots (arrow). In contrast, DH4079 plants (right) only show either no specific response in some plants or just callus formation (arrowhead) in others. (B): Comparison of the percentages of root and callus formation in DH12075 and DH4079. Bar: 1 cm.

Figure 6.
Diagrams of the vectors and primers used for B. napus transformation. (A). Vectors used for the optimization of B. napus transformation protocol: YC3.6-Bar, PM-YC3.6-LTI6b and CRISPR/Cas9 recombinant plasmid (pCRISPR). LB: Left Border; RB: Right Border; Ag7: gene7; mas: mannopine synthase; UBQ10: UBIQUITIN10; nos: nopalin synthase; rbcS: ribulose bisphosphate carboxylase oxygenase small subunit. (B). Scheme of the primers (black arrows) used to genotype the regenerated plants and their amplification products. The first scheme shows the primer combinations used for cameleon constructs (YC3.6-Bar and PM-YC3.6-LTI6b), and the second scheme shows the primers used for CRISPR/Cas9 constructs.
Figure 6.
Diagrams of the vectors and primers used for B. napus transformation. (A). Vectors used for the optimization of B. napus transformation protocol: YC3.6-Bar, PM-YC3.6-LTI6b and CRISPR/Cas9 recombinant plasmid (pCRISPR). LB: Left Border; RB: Right Border; Ag7: gene7; mas: mannopine synthase; UBQ10: UBIQUITIN10; nos: nopalin synthase; rbcS: ribulose bisphosphate carboxylase oxygenase small subunit. (B). Scheme of the primers (black arrows) used to genotype the regenerated plants and their amplification products. The first scheme shows the primer combinations used for cameleon constructs (YC3.6-Bar and PM-YC3.6-LTI6b), and the second scheme shows the primers used for CRISPR/Cas9 constructs.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Summary of the results of DH4079 and SH12075 B. napus transformation with the B, Z and A protocols.
Table 1.
Summary of the results of DH4079 and SH12075 B. napus transformation with the B, Z and A protocols.
| Genotype | Protocol | Plasmid | Resistance | Explants | Plants/Explant | Percentage of Positive Plants |
|---|---|---|---|---|---|---|
| 4079 | B | YC3.6-Bar | BASTA | 296 | 0.00 | 0% |
| 4079 | B | PM-YC3.6-LTI6b | Kanamycin | 266 | 0.00 | 0% |
| 12075 | B | YC3.6-Bar | BASTA | 285 | 0.00 | 0% |
| 12075 | B | PM-YC3.6-LTI6b | Kanamycin | 403 | 0.00 | 0% |
| 4079 | Z | YC3.6-Bar | BASTA | 138 | 0.00 | 0% |
| 4079 | Z | PM-YC3.6-LTI6b | Kanamycin | 143 | 0.02 | 0% |
| 12075 | Z | YC3.6-Bar | BASTA | 98 | 0.00 | 0% |
| 12075 | Z | PM-YC3.6-LTI6b | Kanamycin | 138 | 0.46 | 0% |
| 4079 | A | YC3.6-Bar | BASTA | 358 | 2.48 | 0% |
| 4079 | A | PM-YC3.6-LTI6b | Kanamycin | 361 | 0.64 | 0% |
| 4079 | A | CRISPR | Kanamycin | 222 | 0.12 | 0% |
| 12075 | A | CRISPR | Kanamycin | 153 | 0.19 | 3.4% |
Table 2.
Effect of the protocols on the regeneration capacity of plants. Different letters indicate statistically significant differences according to LSD test (p ≤ 0.05).
Table 2.
Effect of the protocols on the regeneration capacity of plants. Different letters indicate statistically significant differences according to LSD test (p ≤ 0.05).
| Protocol | Plants/Explant |
|---|---|
| B | 0.00 ± 0.00 b |
| Z | 0.12 ± 0.06 ab |
| A | 1.10 ± 0.12 a |
Table 3.
Composition of the different culture media used for management and transformation of bacterial strains. TBI: Transformation Buffer I; TBII: Transformation Buffer II.
Table 3.
Composition of the different culture media used for management and transformation of bacterial strains. TBI: Transformation Buffer I; TBII: Transformation Buffer II.
| Psi | TBI | TBII | SOC | Liquid YM | Solid YM | MGL | |
|---|---|---|---|---|---|---|---|
| Potassium acetate (mM) | 30 | ||||||
| RbCl2 (mM) | 100 | 10 | |||||
| CaCl2·2H2O (mM) | 10 | 75 | |||||
| MnCl2·4H2O (mM) | 50 | ||||||
| MOPS (mM) | 10 | ||||||
| MgCl2 (mM) | 52.63 | 10 | |||||
| NaCl (mM) | 10 | 1.7 | 1.7 | 85.5 | |||
| KCl (mM) | 2.5 | ||||||
| MgSO4 (mM) | 10 | ||||||
| MgSO4·7H2O (mM) | 0.8 | 0.8 | 0.4 | ||||
| Glucose (mM) | 20 | ||||||
| K2HPO4·3H2O (mM) | 2.2 | 2.2 | 1.4 | ||||
| Tryptone (%) | 2 | 2 | 0.5 | ||||
| Yeast extract (%) | 0.5 | 0.5 | 0.04 | 0.04 | 0.25 | ||
| Mannitol (%) | 1 | 1 | 0.5 | ||||
| Glycerol (% v/v) | 15 | 15 | |||||
| L-glutamic acid (g/L) | 1 | ||||||
| Biotin (mg/L) | 1 | ||||||
| Bacteriological agar (%) | 1 | ||||||
| Rifampicin (mg/L) | 50 | ||||||
| pH | 7.6 | 5.8 | 6.5 | 7 | 7 | 7 | 7 |
Table 4.
Plasmid selection agents used for bacterial transformation.
| PM-YC3.6-LTI6b | YC3.6-Bar | CRISPRp | |
|---|---|---|---|
| Kanamycin | 50 mg/L | 50 mg/L | |
| Spectinomycin | 100 mg/L | ||
| Streptomycin | 100 mg/L |
Table 5.
Composition of the different culture media used for explant transformation and plant regeneration. 2,4-D: 2,4-dichlorophenoxyacetic acid; BAP: 6-benzylaminopurine; CCM: Co-Cultivation Medium; CIM: Callus Induction Medium; GM: germination medium; MS + V: Murashige and Skoog basal medium including vitamins (Duchefa Biochemie); RRM: Root Regeneration Medium; SM: Selection medium; SOM: Shoot Outgrowth Medium; SRM: Shoot Regeneration Medium.
Table 5.
Composition of the different culture media used for explant transformation and plant regeneration. 2,4-D: 2,4-dichlorophenoxyacetic acid; BAP: 6-benzylaminopurine; CCM: Co-Cultivation Medium; CIM: Callus Induction Medium; GM: germination medium; MS + V: Murashige and Skoog basal medium including vitamins (Duchefa Biochemie); RRM: Root Regeneration Medium; SM: Selection medium; SOM: Shoot Outgrowth Medium; SRM: Shoot Regeneration Medium.
| GM | CIM | SRM | SOM | RRM | CCM | SM | RM | |
|---|---|---|---|---|---|---|---|---|
| MS + V (g/L) | 4.6 | 4.6 | 4.6 | 4.6 | 2.3 | 4.6 | 4.6 | 2.3 |
| MES (g/L) | 2.5 | 2.5 | 2.5 | 2.5 | ||||
| Myo-inositol (g/L) | 1 | 1 | 1 | 1 | ||||
| AgNO3 (mg/L) | 5 | 5 | ||||||
| 2,4-D (mg/L) | 1 | |||||||
| BAP (mg/L) | 5 | 0.05 | 2 | 2 | ||||
| Sucrose (g/L) | 20 | 30 | 30 | 30 | 10 | 30 | 30 | 10 |
| Plant agar (g/L) | 8 | 8 | 8 | 8 | 8 | 8 | 8 | 8 |
| Carbenicillin (mg/L) | 500 | 500 | 100 | 500 | 500 | |||
| pH | 5.8 | 5.8 | 5.8 | 5.8 | 5.8 | 5.8 | 5.8 | 5.8 |
Table 6.
Oligonucleotides used as primers (Fw: Forward primer; Rv: Reverse primer).
| Oligonucleotide | Sequence (5′ to 3′) |
|---|---|
| 35S 3′ Fw | GATGACGCACAATCCCACTATCC |
| hCas9_3330 Rv | GCAGAATGGCGTCTGACAGG |
| YC3.6 Fw | TAAACGGCCACAGGTTCAGC |
| YC3.6 Rv | CGATCACATGGTCCTGCTGGA |
| GFP5′ Rv | GCGACGTAAACGGCCACAAGTTCAG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Calabuig-Serna, A.; Mir, R.; Porcel, R.; Seguí-Simarro, J.M. The Highly Embryogenic Brassica napus DH4079 Line Is Recalcitrant to Agrobacterium-Mediated Genetic Transformation. Plants 2023, 12, 2008. https://doi.org/10.3390/plants12102008
AMA Style
Calabuig-Serna A, Mir R, Porcel R, Seguí-Simarro JM. The Highly Embryogenic Brassica napus DH4079 Line Is Recalcitrant to Agrobacterium-Mediated Genetic Transformation. Plants. 2023; 12(10):2008. https://doi.org/10.3390/plants12102008
Chicago/Turabian StyleCalabuig-Serna, Antonio, Ricardo Mir, Rosa Porcel, and Jose M. Seguí-Simarro. 2023. "The Highly Embryogenic Brassica napus DH4079 Line Is Recalcitrant to Agrobacterium-Mediated Genetic Transformation" Plants 12, no. 10: 2008. https://doi.org/10.3390/plants12102008
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.