

Synergistic Effects of Rhizobacteria and Salicylic Acid on Maize Salt-Stress Tolerance

 , , ,
, , ,  ,
,  ,
,
Abstract
:1. Introduction
2. Results
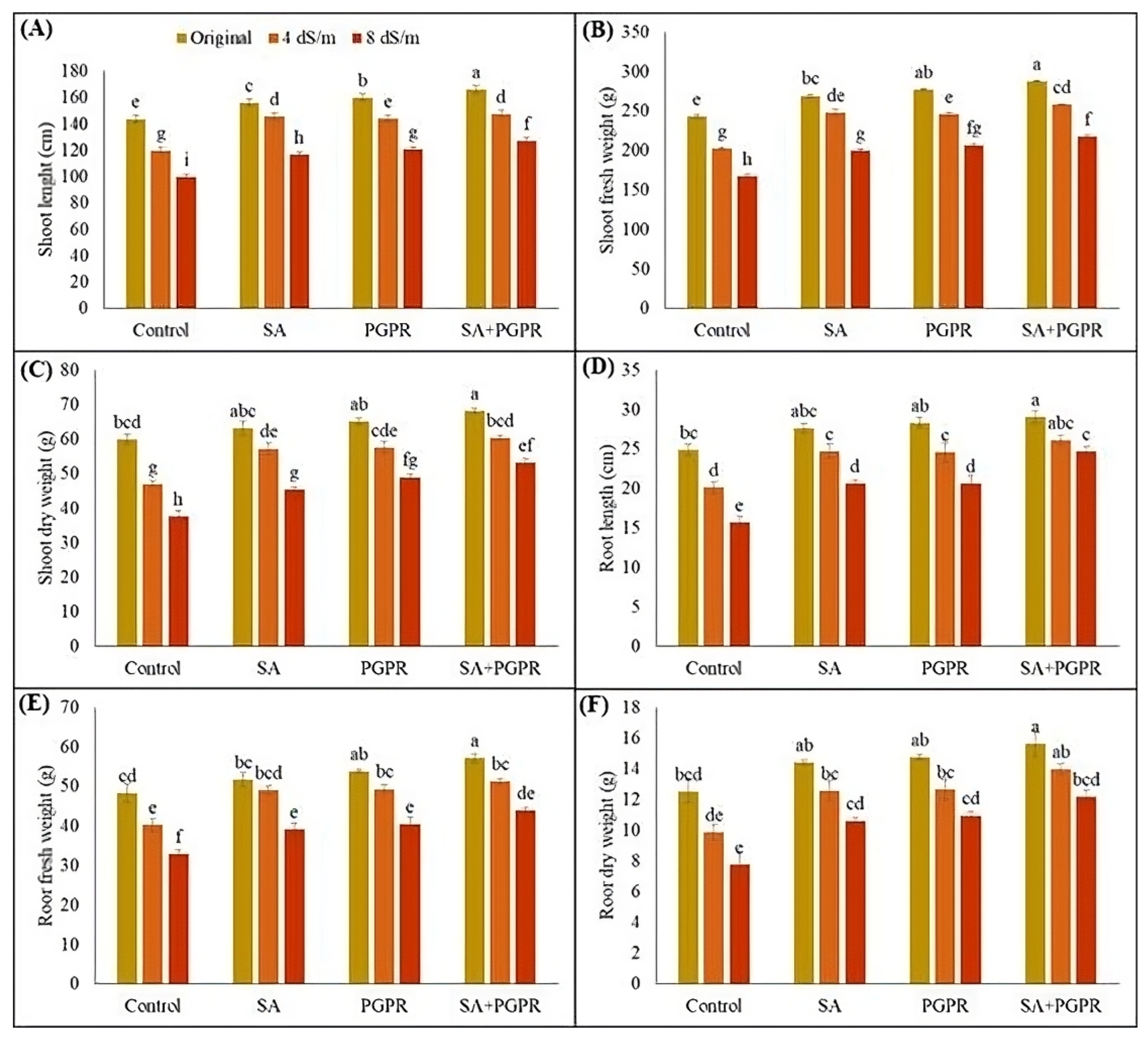
2.1. Effects on Shoot and Root Attributes of Maize
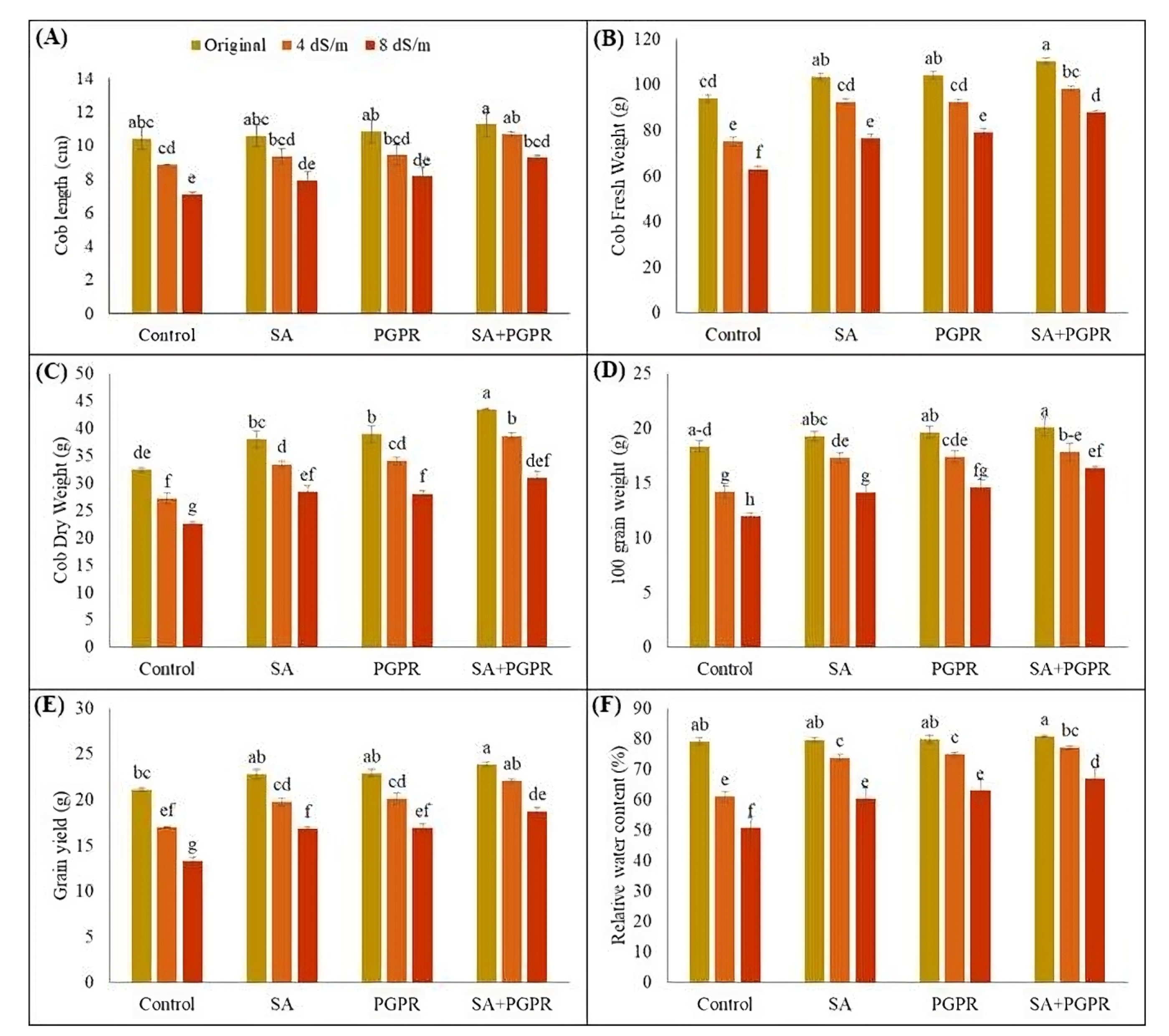
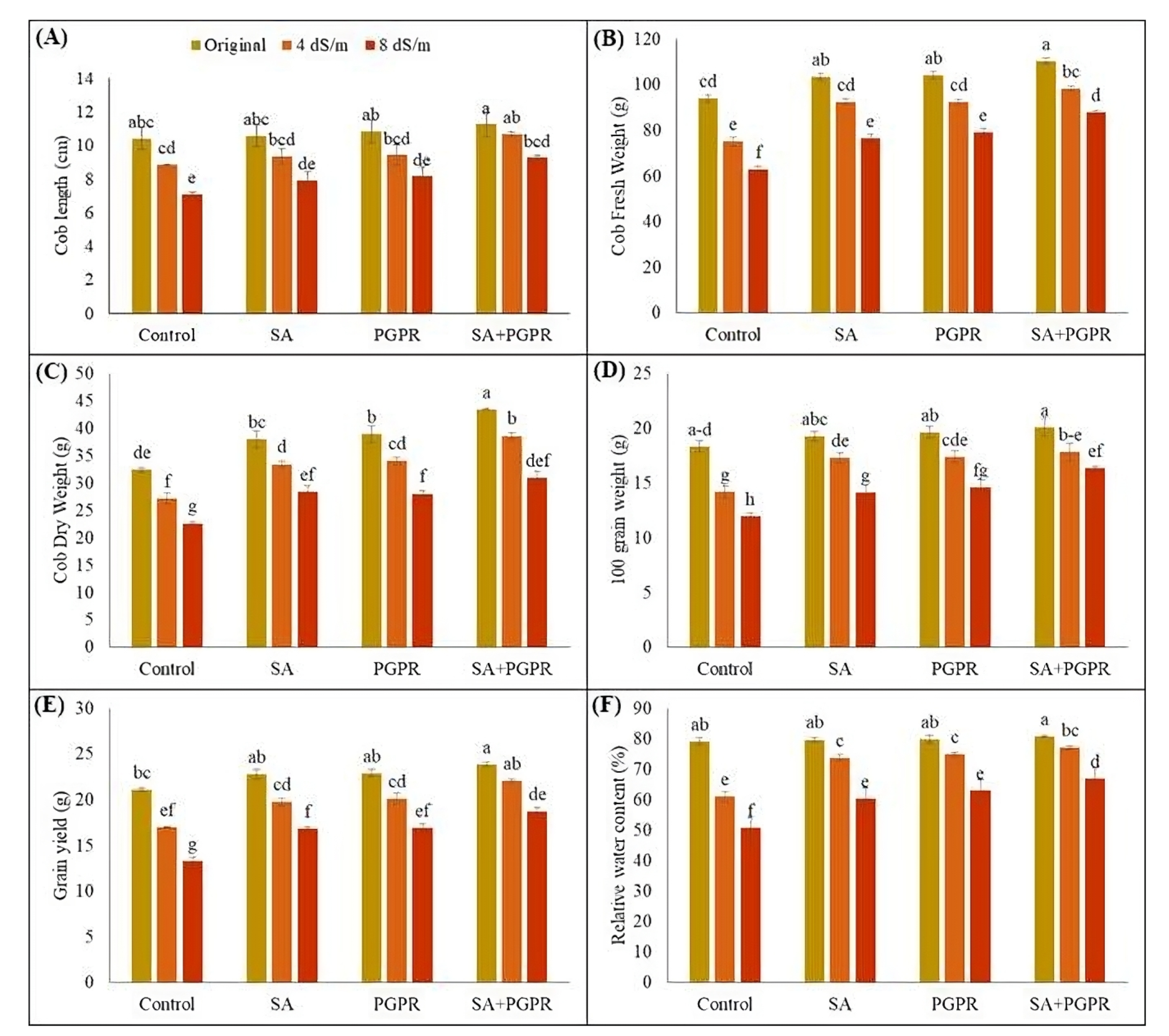
2.2. Effect on Maize Yield and Yield-Related Attributes and Relative Water Contents
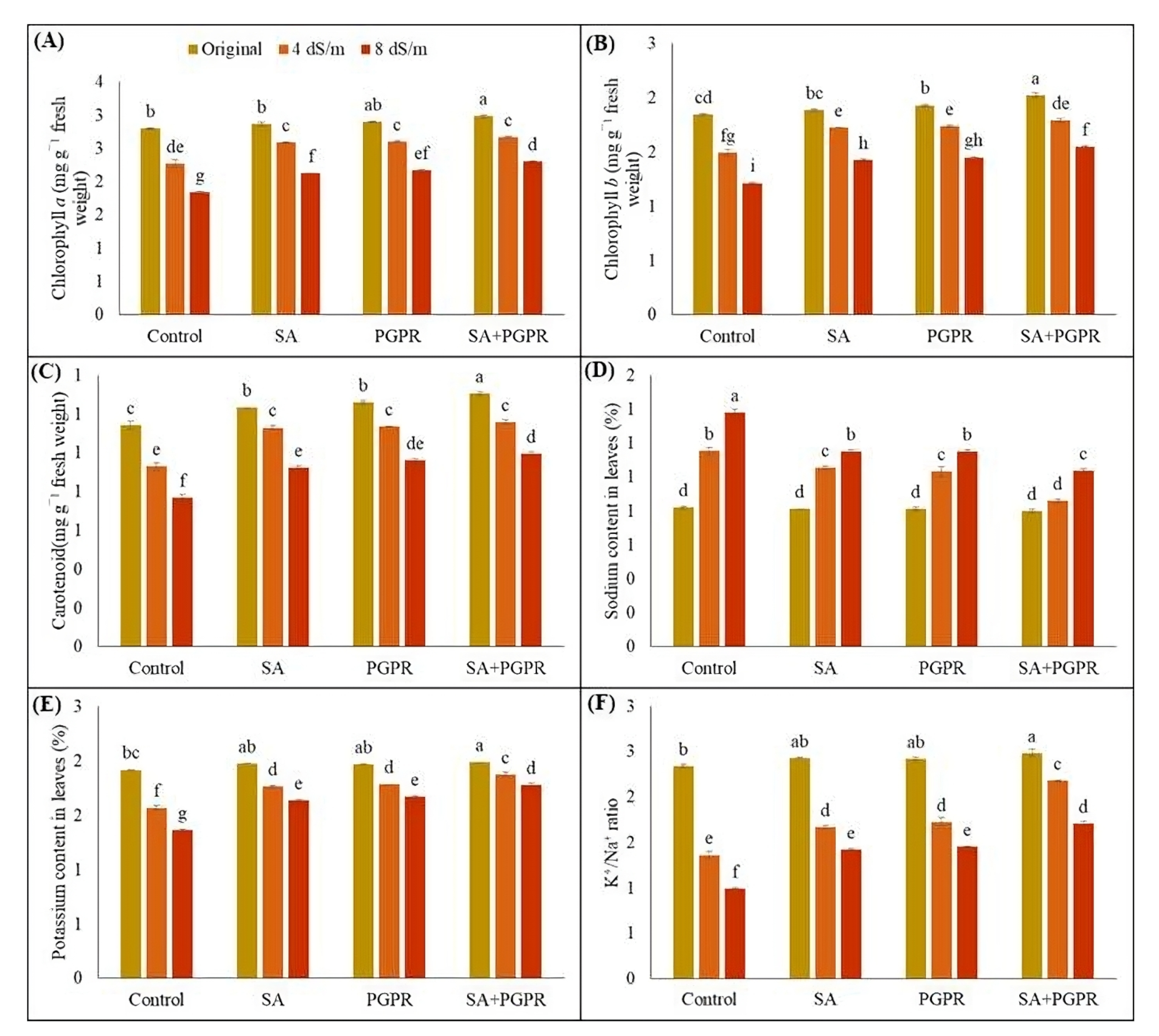
2.3. Effects on Photosynthesis Pigments and Ionic Content of Maize Plants
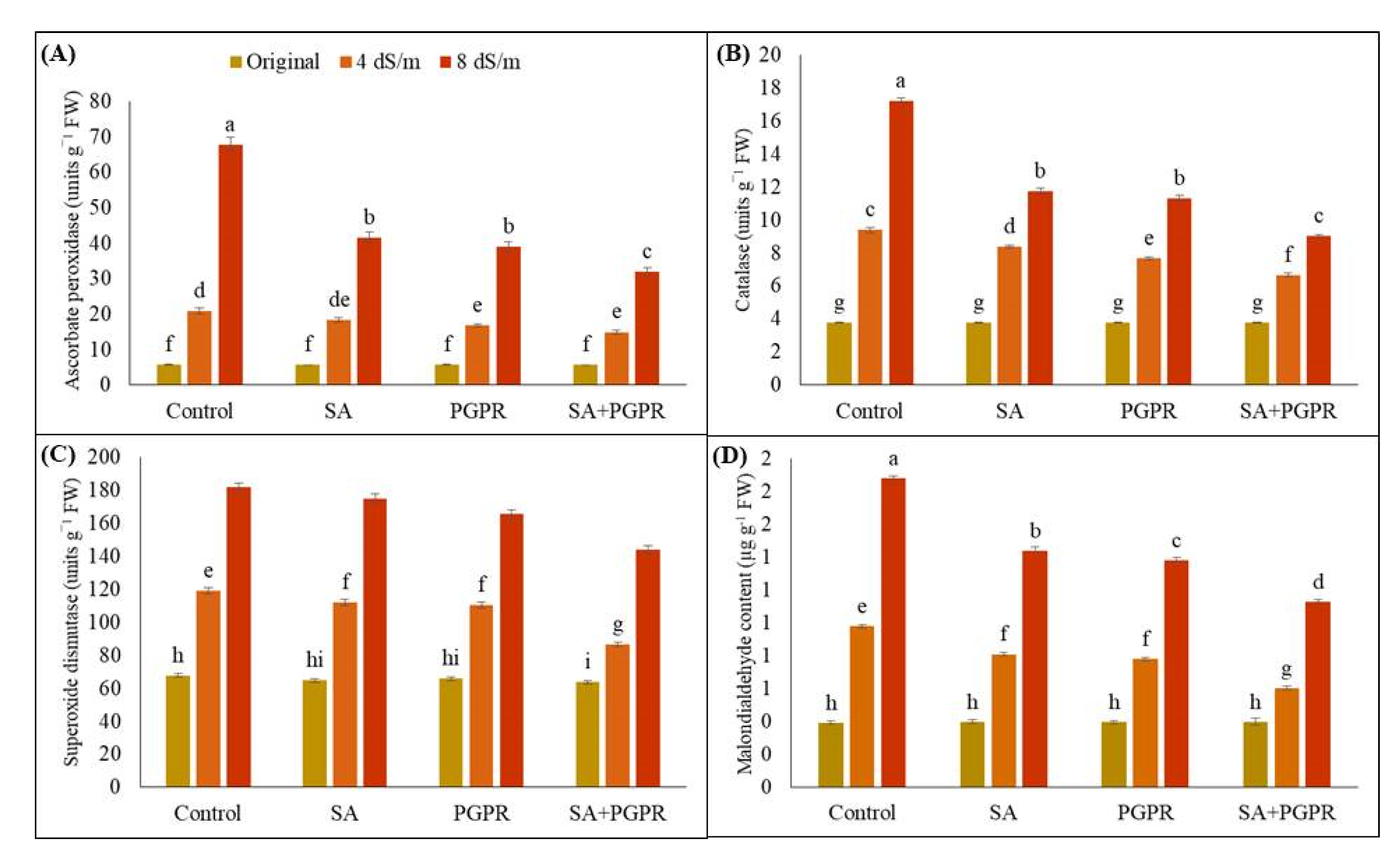
2.4. Effects on Antioxidant and Oxidant System of Maize Plant
2.5. Pearson Correlation Analysis
3. Discussion
4. Materials and Methods
4.1. Volumetric Moisture Contents (%)
4.2. Soil pH and Electrical Conductivity (EC)
4.3. Soil Organic Matter (OM)
4.4. Nutrients Concentration from Soil
4.5. Textural Determination
4.6. Surface Sterilization of Seeds
4.7. Acquisition of Salt-Tolerant PPGP Strain
4.8. Preparation of Inoculum
4.9. Seeds Inoculation
4.10. Pot Experiment
4.11. Determination of Relative Water Content
4.12. Estimation of Photosynthetic Pigments
4.13. Extraction of Antioxidant and Oxidant Activities
4.14. Growth and Yield Parameters
4.15. Sodium and Potassium Determination
4.16. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Diacono, M.; Montemurro, F. Effectiveness of Organic Wastes as Fertilizers and Amendments in Salt-Affected Soils. Agriculture 2015, 5, 221–230. [Google Scholar] [CrossRef] [Green Version]
- Parihar, P.; Singh, S.; Singh, R.; Singh, V.P.; Prasad, S.M. Effect of Salinity Stress on Plants and its Tolerance Strategies: A Review. Environ. Sci. Pollut. Res. 2015, 22, 4056–4075. [Google Scholar] [CrossRef] [PubMed]
- Kaya, C.; Ashraf, M.; Dikilitas, M.; Tuna, A.L. Alleviation of Salt Stress-Induced Adverse Effects on Maize Plants by Exogenous Application of Indoleacetic Acid (IAA) and Inorganic Nutrients—A Field Trial. Aust. J. Crop Sci. 2013, 7, 249–254. [Google Scholar]
- Hussain, S.; Zhang, J.; Zhong, C.; Zhu, L.; Cao, X.; Yu, S.; Bohr, J.A.; Hu, J.; Jin, Q. Effects of Salt Stress on Rice Growth, Development Characteristics, and the Regulating Ways: A Review. J. Integr. Agric. 2017, 16, 2357–2374. [Google Scholar] [CrossRef] [Green Version]
- Siddiqui, M.N.; Mostofa, M.G.; Akter, M.M.; Srivastava, A.K.; Sayed, M.A.; Hasan, M.S.; Tran, L.S.P. Impact of Salt-Induced Toxicity on Growth and Yield-Potential of Local Wheat Cultivars: Oxidative Stress and Ion Toxicity are among the Major Determinants of Salt Tolerant Capacity. Chemosphere 2017, 187, 385–394. [Google Scholar] [CrossRef] [PubMed]
- Razzaq, A.; Ali, A.; Safdar, L.B.; Zafar, M.M.; Rui, Y.; Shakeel, A.; Shaukat, A.; Ashraf, M.; Gong, W.; Yuan, Y. Salt stress induces physiochemical alterations in rice grain composition and quality. J. Food Sci. 2020, 85, 14–20. [Google Scholar] [CrossRef]
- Machado, R.M.A.; Serralheiro, R.P. Soil Salinity: Effect on Vegetable Crop Growth. Management Practices to Prevent and Mitigate Soil Salinization. Horticulturae 2017, 3, 30. [Google Scholar] [CrossRef] [Green Version]
- Akram, M.S.; Shahid, M.; Tariq, M.; Azeem, M.; Javed, M.T.; Saleem, S.; Riaz, S. Deciphering Staphylococcus sciuri SAT-17 Mediated Anti-Oxidative Defense Mechanisms and Growth Modulations in Salt Stressed Maize (Zea mays L.). Front. Microbiol. 2016, 7, 867. [Google Scholar] [CrossRef]
- Syed, A.; Sarwar, G.; Shah, S.H.; Muhammad, S. Soil Salinity Research in 21st Century in Pakistan: Its Impact on Availability of Plant Nutrients, Growth and Yield of Crops. Commun. Soil Sci. Plant Anal. 2021, 52, 183–200. [Google Scholar] [CrossRef]
- Meena, M.D.; Yadav, R.K.; Narjary, B.; Yadav, G.; Jat, H.S.; Sheoran, P.; Meena, M.K.; Antil, R.S.; Meena, B.L.; Singh, H.V.; et al. Municipal Solid Waste (MSW): Strategies to Improve Salt Affected Soil Sustainability: A Review. Waste Manag. 2019, 84, 38–53. [Google Scholar] [CrossRef]
- Delgado, A.; Gómez, J.A. The Soil. Physical, Chemical and Biological Properties, 1st ed.; Springer International Publishing: Cham, Switzerland, 2016; pp. 15–26. [Google Scholar]
- Wang, D.; Wang, H.; Han, B.; Wang, B.; Guo, A.; Zheng, D.; Liu, C.; Chang, L.; Peng, M.; Wang, X. Sodium instead of potassium and chloride is an important macronutrient to improve leaf succulence and shoot development for halophyte Sesuvium portulacastrum. Plant Physiol. Biochem. 2012, 51, 53–62. [Google Scholar] [CrossRef]
- Kataria, S.; Verma, S.K. Salinity Stress Responses and Adaptive Mechanisms in Major Glycophytic Crops: The Story So Far. In Salinity Responses and Tolerance in Plants, Volume 1: Targeting Sensory, Transport and Signaling Mechanisms; Ahmad, P., Azooz, M.M., Prasad, M.N.V., Eds.; Springer: Cham, Switzerland, 2018; pp. 1–39. [Google Scholar]
- Tahjib-Ul-Arif, M.; Siddiqui, M.N.; Sohag, A.A.M.; Sakil, M.A.; Rahman, M.M.; Polash, M.A.S.; Mostofa, M.G.; Tran, L.S.P. Salicylic Acid-Mediated Enhancement of Photosynthesis Attributes and Antioxidant Capacity Contributes to Yield Improvement of Maize Plants under Salt Stress. J. Plant Growth Regul. 2018, 37, 1318–1330. [Google Scholar] [CrossRef]
- Marriboina, S.; Attipalli, R.R. Hydrophobic Cell-Wall Barriers and Vacuolar Sequestration of Na+ Ions Are among the Key Mechanisms Conferring High Salinity Tolerance in a Biofuel Tree Species, Pongamia pinnata L. Pierre. Environ. Exp. Bot. 2020, 171, 103949. [Google Scholar] [CrossRef]
- Sachdev, S.; Ansari, S.A.; Ansari, M.I. ROS Generation in Plant Cells Orchestrated by Stress. In Reactive Oxygen Species in Plants: The Right Balance; Gupta, D.K., Corpas, F.J., Eds.; Springer Nature Singapore Pte Ltd.: Singapore, 2023; pp. 23–43. [Google Scholar]
- Acosta-Motos, J.R.; Ortuño, M.F.; Bernal-Vicente, A.; Diaz-Vivancos, P.; Sanchez-Blanco, M.J.; Hernandez, J.A. Plant Responses to Salt Stress: Adaptive Mechanisms. Agronomy 2017, 7, 18. [Google Scholar] [CrossRef] [Green Version]
- Polash, M.A.S.; Sakil, M.A.; Hossain, M.A. Plants Responses and Their Physiological and Biochemical Defense Mechanisms against Salinity: A Review. Trop. Plant Res. 2019, 6, 250–274. [Google Scholar] [CrossRef]
- Zhao, C.; Zhang, H.; Song, C.; Zhu, J.K.; Shabala, S. Mechanisms of Plant Responses and Adaptation to Soil Salinity. Innovation 2020, 1, 100017. [Google Scholar] [CrossRef] [PubMed]
- Maqsood, M.F.; Shahbaz, M.; Zulfiqar, U.; Saman, R.U.; Rehman, A.; Naz, N.; Akram, M.; Haider, F.U. Enhancing Wheat Growth and Yield through Salicylic Acid-Mediated Regulation of Gas Exchange, Antioxidant Defense, and Osmoprotection under Salt Stress. Stresses 2023, 3, 27. [Google Scholar] [CrossRef]
- Nudrat, A.A.; Fahad, S.; Muhammad, A. Ascorbic Acid-a Potential Oxidant Scavenger and Its Role in Plant Development and Abiotic Stress Tolerance. Front. Plant Sci. 2017, 8, 613. [Google Scholar]
- Kaya, C.; Ugurlar, F.; Ashraf, M.; Ahmad, P. Salicylic Acid Interacts with Other Plant Growth Regulators and Signal Molecules in Response to Stressful Environments in Plants. Plant Physiol. Biochem. 2023, 196, 431–443. [Google Scholar] [CrossRef]
- Sharma, A.; Kohli, S.K.; Khanna, K.; Ramakrishnan, M.; Kumar, V.; Bhardwaj, R.; Brestic, M.; Skalicky, M.; Landi, M.; Zheng, B. Salicylic Acid: A Phenolic Molecule with Multiple Roles in Salt-Stressed Plants. J. Plant Growth Regul. 2023. [Google Scholar] [CrossRef]
- Kusvuran, S.; Yilmaz, U.D. Ameliorative Role of Salicylic Acid in the Growth, Nutrient Content, and Antioxidative Responses of Salt-Stressed Lettuce. Acta Sci. Pol. Hortorum Cultus 2023, 22, 75–85. [Google Scholar] [CrossRef]
- Ghassemi-Golezani, K.; Farhadi, N. The Efficacy of Salicylic Acid Levels on Photosynthetic Activity, Growth, and Essential Oil Content and Composition of Pennyroyal Plants under Salt Stress. J. Plant Growth Regul. 2021, 41, 1953–1965. [Google Scholar] [CrossRef]
- Youssef, S.M.; López-Orenes, A.; Ferrer, M.A.; Calderón, A.A. Foliar Application of Salicylic Acid Enhances the Endogenous Antioxidant and Hormone Systems and Attenuates the Adverse Effects of Salt Stress on Growth and Yield of French Bean Plants. Horticulturae 2023, 9, 75. [Google Scholar] [CrossRef]
- Omidi, M.; Khandan-Mirkohi, A.; Kafi, M.; Zamani, Z.; Ajdanian, L.; Babaei, M. Biochemical and Molecular Responses of Rosa damascena Mill. cv. Kashan to Salicylic Acid under Salinity Stress. BMC Plant Biol. 2022, 22, 373. [Google Scholar] [CrossRef]
- Talaat, N.B.; Todorova, D. Antioxidant Machinery and Glyoxalase System Regulation Confers Salt Stress Tolerance to Wheat (Triticum aestivum L.) Plants Treated with Melatonin and Salicylic Acid. J. Soil Sci. Plant Nutr. 2022, 22, 3527–3540. [Google Scholar]
- Etesami, H.; Adl, S.M. Plant Growth-Promoting Rhizobacteria (PGPR) and Their Action Mechanisms in Availability of Nutrients to Plants. In Phyto-Microbiome in Stress Regulation; Springer: Singapore, 2020; pp. 147–203. [Google Scholar]
- Das, P.P.; Singh, K.R.; Nagpure, G.; Mansoori, A.; Singh, R.P.; Ghazi, I.A.; Kumar, A.; Singh, J. Plant-Soil-Microbes: A Tripartite Interaction for Nutrient Acquisition and Better Plant Growth for Sustainable Agricultural Practices. Environ. Res. 2022, 214, 113821. [Google Scholar] [CrossRef] [PubMed]
- Chandran, H.; Meena, M.; Swapnil, P. Plant Growth-Promoting Rhizobacteria as a Green Alternative for Sustainable Agriculture. Sustainability 2021, 13, 10986. [Google Scholar] [CrossRef]
- Bhavya, K.; Geetha, A. Plant Growth Promoting Rhizobacteria. Adv. Agric. Sci. 2021, 61, 87. [Google Scholar]
- Wang, F.; Zhao, H.; Yu, C.; Tang, J.; Wu, W.; Yang, Q. Determination of the Geographical Origin of Maize (Zea mays L.) Using Mineral Element Fingerprints. J. Sci. Food Agric. 2020, 100, 1294–1300. [Google Scholar]
- Elhakem, A. Salicylic Acid Ameliorates Salinity Tolerance in Maize by Regulation of Phytohormones and Osmolytes. Plant Soil Environ. 2020, 66, 533–541. [Google Scholar] [CrossRef]
- Singh, A.; Radzhput, V.D.; Rawat, S.; Singh, A.K.; Bind, A.; Singh, A.K.; Chernikova, N.P.; Voloshina, M.S.; Lobzenko, I.P. Monitoring Soil Salinity and Recent Advances in Mechanism of Salinity Tolerance in Plants. Biogeosyst. Tech. 2020, 7, 66–86. [Google Scholar]
- Ali, M.; Afzal, S.; Parveen, A.; Kamran, M.; Javed, M.R.; Abbasi, G.H.; Malik, Z.; Riaz, M.; Ahmad, S.; Chattha, M.S.; et al. Silicon Mediated Improvement in the Growth and Ion Homeostasis by Decreasing Na+ Uptake in Maize (Zea mays L.) Cultivars Exposed to Salinity Stress. Plant Physiol. Biochem. 2021, 158, 208–218. [Google Scholar] [CrossRef] [PubMed]
- Ondrasek, G.; Rathod, S.; Manohara, K.K.; Gireesh, C.; Anantha, M.S.; Sakhare, A.S.; Parmar, B.; Yadav, B.K.; Bandumula, N.; Raihan, F.; et al. Salt Stress in Plants and Mitigation Approaches. Plants 2022, 11, 717. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Singh, P.; Dhillon, G.P.S.; Sharma, S.; Singh, B.; Gill, R.I.S. Differential Impacts of Soil Salinity and Water Logging on Eucalyptus Growth and Carbon Sequestration under Mulched vs. Unmulched Soils in South-Western Punjab, India. Plant Soil 2023, 482, 401–425. [Google Scholar] [CrossRef]
- Ahmad, P.; Abdel Latef, A.A.; Hashem, A.; AbdAllah, E.F.; Gucel, S.; Tran, L.S.P. Nitric Oxide Mitigates Salt Stress by Regulating Levels of Osmolytes and Antioxidant Enzymes in Chickpea. Front. Plant Sci. 2016, 7, 347. [Google Scholar] [CrossRef] [Green Version]
- Aneela, R.İ.A.Z.; Rafique, M.; Aftab, M.; Qureshi, M.A.; Javed, H.; Mujeeb, F.; Akhtar, S. Mitigation of Salinity in Chickpea by Plant Growth Promoting Rhizobacteria and Salicylic Acid. Eurasian J. Soil Sci. 2019, 8, 221–228. [Google Scholar]
- Khan, N.; Bano, A.; Curá, J.A. Role of Beneficial Microorganisms and Salicylic Acid in Improving Rainfed Agriculture and Future Food Safety. Microorganisms 2020, 8, 1018. [Google Scholar] [CrossRef]
- Pasternak, T.; Groot, E.P.; Kazantsev, F.V.; Teale, W.; Omelyanchuk, N.; Kovrizhnykh, V.; Palme, K.; Mironova, V.V. Salicylic Acid Affects Root Meristem Patterning via Auxin Distribution in a Concentration-Dependent Manner. Plant Physiol. 2019, 180, 1725–1739. [Google Scholar] [CrossRef] [PubMed]
- Pii, Y.; Mimmo, T.; Tomasi, N.; Terzano, R.; Cesco, S.; Crecchio, C. Microbial Interactions in the Rhizosphere: Beneficial Influences of Plant Growth-Promoting Rhizobacteria on Nutrient Acquisition Process. A Review. Biol. Fertil. Soils 2015, 51, 403–415. [Google Scholar] [CrossRef]
- Riaz, U.; Murtaza, G.; Anum, W.; Samreen, T.; Sarfraz, M.; Nazir, M.Z. Plant Growth-Promoting Rhizobacteria (PGPR) as Biofertilizers and Biopesticides. In Microbiota and Biofertilizers: A Sustainable Continuum for Plant and Soil Health; Egamberdieva, D., Ahmad, P., Eds.; Springer: Cham, Switzerland, 2021; pp. 181–196. [Google Scholar]
- Tariq, M.; Noman, M.; Ahmed, T.; Hameed, A.; Manzoor, N.; Zafar, M. Antagonistic Features Displayed by Plant Growth Promoting Rhizobacteria (PGPR): A Review. J. Plant Sci. Phytopathol. 2017, 1, 038–043. [Google Scholar]
- Verma, P.P.; Shelake, R.M.; Das, S.; Sharma, P.; Kim, J.Y. Plant Growth-Promoting Rhizobacteria (PGPR) and Fungi (PGPF): Potential Biological Control Agents of Diseases and Pests. In Microbial Interventions in Agriculture and Environment: Vol 1: Research Trends, Priorities and Prospects; Singh, D.P., Singh, H.B., Prabha, R., Dubey, N.K., Eds.; Springer: Singapore, 2019; pp. 281–311. [Google Scholar]
- Khan, A.; Sayyed, R.Z.; Seifi, S. Rhizobacteria: Legendary Soil Guards in Abiotic Stress Management. In Plant Growth Promoting Rhizobacteria for Sustainable Stress Management: Volume 1: Rhizobacteria in Abiotic Stress Management; Singh, D.P., Singh, H.B., Prabha, R., Dubey, N.K., Eds.; Springer Nature: Singapore, 2019; pp. 327–343. [Google Scholar]
- Ahluwalia, O.; Singh, P.C.; Bhatia, R. A Review on Drought Stress in Plants: Implications, Mitigation and the Role of Plant Growth Promoting Rhizobacteria. Resources 2021, 5, 32. [Google Scholar] [CrossRef]
- Allakhverdiev, S.I. Optimising Photosynthesis for Environmental Fitness. Funct. Plant Biol. 2020, 47, iii–vii. [Google Scholar] [CrossRef]
- Barickman, C.; Kopsell, D.A.; Sams, C. Abscisic Acid Increases Carotenoid and Chlorophyll Concentrations in Leaves and Fruit of Two Tomato Genotypes. J. Am. Soc. Hortic. Sci. 2014, 139, 261–266. [Google Scholar] [CrossRef] [Green Version]
- Nigam, B.; Dubey, R.S.; Rathore, D. Protective Role of Exogenously Supplied Salicylic Acid and PGPB (Stenotrophomonas sp.) on Spinach and Soybean Cultivars Grown under Salt Stress. Sci. Hortic. 2022, 293, 110654. [Google Scholar] [CrossRef]
- Jamshidi Jam, B.; Shekari, F.; Andalibi, B.; Fotovat, R.; Jafarian, V.; Dolatabadian, A. The Effects of Salicylic Acid and Silicon on Safflower Seed Yield, Oil Content, and Fatty Acids Composition under Salinity Stress. Silicon 2023, 15, 4081–4094. [Google Scholar] [CrossRef]
- Khan, M.I.R.; Palakolanu, S.R.; Chopra, P.; Rajurkar, A.B.; Gupta, R.; Iqbal, N.; Maheshwari, C. Improving Drought Tolerance in Rice: Ensuring Food Security through Multi-Dimensional Approaches. Physiol. Plant. 2021, 172, 645–668. [Google Scholar] [CrossRef]
- Shabala, S.; Pottosin, I. Regulation of Potassium Transport in Plants under Hostile Conditions: Implications for Abiotic and Biotic Stress Tolerance. Physiol. Plant. 2014, 151, 257–279. [Google Scholar] [CrossRef]
- Ahmad, P.; Hashem, A.; Abd-Allah, E.F.; Alqarawi, A.A.; John, R.; Egamberdieva, D. Role of Trichoderma harzianum in Mitigating NaCl Stress in Indian Mustard (Brassica juncea L.) through Antioxidative Defense System. Front. Plant Sci. 2015, 6, 868. [Google Scholar] [CrossRef] [Green Version]
- Karlidag, H.; Yildirim, E.; Turan, M. Salicylic Acid Ameliorates the Adverse Effect of Salt Stress on Strawberry. Sci. Agric. 2009, 66, 180–187. [Google Scholar] [CrossRef]
- Mangalassery, S.; Dayal, D.; Kumar, A.; Bhatt, K.; Nakar, R.; Kumar, A.; Singh, J.P.; Misra, A.K. Pattern of Salt Accumulation and Its Impact on Salinity Tolerance in Two Halophyte Grasses in Extreme Saline Desert in India. Agronomy 2017, 7, 81. [Google Scholar]
- Maas, E.V. Salt Tolerance of Plants. In CRC Handbook of Plant Science in Agriculture Volume II; CRC Press: Boca Raton, FL, USA, 2019; pp. 57–76. [Google Scholar]
- Shahid, M.A.; Sarkhosh, A.; Khan, N.; Balal, R.M.; Ali, S.; Rossi, L.; Gómez, C.; Mattson, N.; Nasim, W.; Garcia-Sanchez, F. Insights into the Physiological and Biochemical Impacts of Salt Stress on Plant Growth and Development. Agronomy 2020, 10, 938. [Google Scholar] [CrossRef]
- El-Katony, T.M.; El-Bastawisy, Z.M.; El-Ghareeb, S.S. Timing of Salicylic Acid Application Affects the Response of Maize (Zea mays L.) Hybrids to Salinity Stress. Heliyon 2019, 5, e01547. [Google Scholar] [CrossRef] [Green Version]
- Cai, Z.Q.; Gao, Q. Comparative physiological and biochemical mechanisms of salt tolerance in five contrasting highland quinoa cultivars. BMC Plant Biol. 2020, 20, 70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahanger, M.A.; Agarwal, R.M. Salinity stress induced alterations in antioxidant metabolism and nitrogen assimilation in wheat (Triticum aestivum L.) as influenced by potassium supplementation. Plant Physiol. Biochem. 2017, 115, 449–460. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, P.; Alyemeni, M.N.; Ahanger, M.A.; Egamberdieva, D.; Wijaya, L.; Alam, P. Salicylic acid (SA) induced alterations in growth, biochemical attributes and antioxidant enzyme activity in faba bean (Vicia faba L.) seedlings under NaCl toxicity. Russ. J. Plant Physiol. 2018, 65, 104–114. [Google Scholar] [CrossRef]
- Jayakannan, M.; Bose, J.; Babourina, O.; Rengel, Z.; Shabala, S. Salicylic Acid in Plant Salinity Stress Signalling and Tolerance. Plant Growth Regul. 2015, 76, 25–40. [Google Scholar] [CrossRef]
- Ilangumaran, G.; Smith, D.L. Plant Growth Promoting Rhizobacteria in Amelioration of Salinity Stress: A Systems Biology Perspective. Front. Plant Sci. 2017, 8, 1768. [Google Scholar] [CrossRef] [Green Version]
- Elhindi, K.M.; Al-Amri, S.M.; Abdel-Salam, E.M.; Al-Suhaibani, N.A. Effectiveness of Salicylic Acid in Mitigating Salt-Induced Adverse Effects on Different Physio-Biochemical Attributes in Sweet Basil (Ocimum basilicum L.). J. Plant Nutr. 2017, 40, 908–919. [Google Scholar] [CrossRef]
- Paul, D.; Lade, H. Plant-Growth-Promoting Rhizobacteria to Improve Crop Growth in Saline Soils: A Review. Agron. Sustain. Dev. 2014, 34, 737–752. [Google Scholar] [CrossRef]
- Souana, K.; Taïbi, K.; Abderrahim, L.A.; Amirat, M.; Achir, M.; Boussaid, M.; Mulet, J.M. Salt-Tolerance in Vicia faba L. Is Mitigated by the Capacity of Salicylic Acid to Improve Photosynthesis and Antioxidant Response. Sci. Hortic. 2020, 273, 109641. [Google Scholar] [CrossRef]
- Jing, X.; Hou, P.; Lu, Y.; Deng, S.; Li, N.; Zhao, R.; Sun, J.; Wang, Y.; Han, Y.; Lang, T.; et al. Overexpression of Copper/Zinc Superoxide Dismutase from Mangrove Kandelia candel in Tobacco Enhances Salinity Tolerance by the Reduction of Reactive Oxygen Species in Chloroplast. Front. Plant Sci. 2015, 6, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaur, N.; Dhawan, M.; Sharma, I.; Pati, P.K. Interdependency of Reactive Oxygen Species Generating and Scavenging System in Salt Sensitive and Salt Tolerant Cultivars of Rice. BMC Plant Biol. 2016, 16, 131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, C.; Zu, C.; Lu, D.; Zheng, Q.; Shen, J.; Wang, H.; Li, D. Effect of Exogenous Selenium Supply on Photosynthesis, Na+ Accumulation and Antioxidative Capacity of Maize (Zea mays L.) under Salinity Stress. Sci. Rep. 2017, 7, 42039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noreen, S.; Sultan, M.; Akhter, M.S.; Shah, K.H.; Ummara, U.; Manzoor, H.; Ulfat, M.; Alyemeni, M.N.; Ahmad, P. Foliar Fertigation of Ascorbic Acid and Zinc Improves Growth, Antioxidant Enzyme Activity and Harvest Index in Barley (Hordeum vulgare L.) Grown under Salt Stress. Plant Physiol. Biochem. 2021, 158, 244–254. [Google Scholar] [CrossRef]
- Moustafa-Farag, M.; Najeeb, U.; Yang, J.; Hu, Z.; Fang, Z.M. Nitric Oxide Protects Carbon Assimilation Process of Watermelon from Boron-Induced Oxidative Injury. Plant Physiol. Biochem. 2017, 111, 166–173. [Google Scholar] [CrossRef]
- Sarkar, A.; Ghosh, P.K.; Pramanik, K.; Mitra, S.; Soren, T.; Pandey, S.; Maiti, T.K. A Halotolerant Enterobacter sp. Displaying ACC Deaminase Activity Promotes Rice Seedling Growth under Salt Stress. Res. Microbiol. 2018, 169, 20–32. [Google Scholar] [CrossRef]
- García-Caparrós, P.; Hasanuzzaman, M.; Lao, M.T. Oxidative Stress and Antioxidant Defense in Plants under Salinity. In Reactive Oxygen, Nitrogen and Sulfur Species in Plants: Production, Metabolism, Signaling and Defense Mechanisms; Gupta, D.K., Palma, J.M., Corpas, F.J., Eds.; Springer International Publishing: Cham, Switzerland, 2019; pp. 291–309. [Google Scholar]
- Azarabadi, S.R.; Abdollahi, H.; Torabi, M.; Salehi, Z.; Nasiri, J. ROS Generation, Oxidative Burst and Dynamic Expression Profiles of ROS-Scavenging Enzymes of Superoxide Dismutase (SOD), Catalase (CAT) and Ascorbate Peroxidase (APX) in Response to Erwinia amylovora in Pear (Pyrus communis L.). Eur. J. Plant Pathol. 2017, 147, 279–294. [Google Scholar] [CrossRef]
- Rajput, V.D.; Singh, R.K.; Verma, K.K.; Sharma, L.; Quiroz-Figueroa, F.R.; Meena, M.; Gour, V.S.; Minkina, T.; Sushkova, S.; Mandzhieva, S. Recent Developments in Enzymatic Antioxidant Defence Mechanism in Plants with Special Reference to Abiotic Stress. Biology 2021, 10, 267. [Google Scholar] [CrossRef]
- Salinity Laboratory Staff. Diagnosis and Improvement of Saline and Alkali Soils; Agriculture Handbook; U.S. Department of Agriculture: Washington, DC, USA, 1954; Volume 60. [Google Scholar]
- Walkley, A.; Black, I.A. An Examination of the Degtjareff Method for Determining Soil Organic Matter, and a Proposed Modification of the Chromic Acid Titration Method. Soil Sci. 1934, 37, 29–38. [Google Scholar] [CrossRef]
- Waksman, S.A.; Stevens, K.R. A Critical Study of the Methods for Determining the Nature and Abundance of Soil Organic Matter. Soil Sci. 1930, 30, 97–116. [Google Scholar] [CrossRef]
- Watanabe, F.S.; Olsen, S.R. Test of an Ascorbic Acid Method for Determining Phosphorus in Water and NaHCO3 Extracts from Soil. Soil Sci. Soc. Am. J. 1965, 29, 677–678. [Google Scholar] [CrossRef]
- Moodie, C.D.; Smith, H.W.; Mcreery, R.A. Laboratory Manual for Soil Fertility; Pullman State College of Washington: Washington, DC, USA, 1959; pp. 31–39. [Google Scholar]
- Weatherley, P. Studies in the Water Relations of the Cotton Plant. I. The Field Measurement of Water Deficits in Leaves. New Phytol. 1950, 49, 81–97. [Google Scholar] [CrossRef]
- Arnon, D.T. Copper Enzyme in Isolated Chloroplasts, Polyphenaloxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asada, K.; Takahashi, M. Production and Scavenging of Active Oxygens in Chloroplasts. Photoinhibition; Elsevier: Amsterdam, The Netherlands, 1987; pp. 227–287. [Google Scholar]
- Chandlee, J.M.; Scandalios, J.G. Analysis of Variants Affecting the Catalase Developmental Program in Maize Scutellum. Theor. Appl. Genet. 1984, 69, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Giannopolitis, C.N.; Ries, S.K. Superoxide Dismutases: II. Purification and Quantitative Relationship with Water-Soluble Protein in Seedlings. Plant Physiol. 1977, 59, 315–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jambunathan, N. Determination and Detection of Reactive Oxygen Species (ROS), Lipid Peroxidation, and Electrolyte Leakage in Plants. In Plant Stress Tolerance; Sunkar, R., Ed.; Methods in Molecular Biology; Humana Press, Springer: New York, NY, USA; Dordrecht, The Netherlands; Berlin/Heidelberg, Germany; London, UK, 2010; Volume 639, pp. 291–297. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Unit | Value |
|---|---|---|
| Textural class | --------- | Sandy clay loam |
| pH | --------- | 7.71 |
| Gravimetric moisture contents | % | 34 |
| ECe | dS m−1 | 1.46 |
| Organic matter | % | 0.64 |
| Total Nitrogen | % | 0.060 |
| Available phosphorus | mg kg−1 | 6.91 |
| Extractable potassium | mg kg−1 | 134.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ali, Q.; Ahmad, M.; Kamran, M.; Ashraf, S.; Shabaan, M.; Babar, B.H.; Zulfiqar, U.; Haider, F.U.; Ali, M.A.; Elshikh, M.S. Synergistic Effects of Rhizobacteria and Salicylic Acid on Maize Salt-Stress Tolerance. Plants 2023, 12, 2519. https://doi.org/10.3390/plants12132519
Ali Q, Ahmad M, Kamran M, Ashraf S, Shabaan M, Babar BH, Zulfiqar U, Haider FU, Ali MA, Elshikh MS. Synergistic Effects of Rhizobacteria and Salicylic Acid on Maize Salt-Stress Tolerance. Plants. 2023; 12(13):2519. https://doi.org/10.3390/plants12132519
Chicago/Turabian StyleAli, Qasim, Maqshoof Ahmad, Muhammad Kamran, Sana Ashraf, Muhammad Shabaan, Babar Hussain Babar, Usman Zulfiqar, Fasih Ullah Haider, M. Ajmal Ali, and Mohamed S Elshikh. 2023. "Synergistic Effects of Rhizobacteria and Salicylic Acid on Maize Salt-Stress Tolerance" Plants 12, no. 13: 2519. https://doi.org/10.3390/plants12132519