
Development and Evaluation of Zinc and Iron Nanoparticles Functionalized with Plant Growth-Promoting Rhizobacteria (PGPR) and Microalgae for Their Application as Bio-Nanofertilizers
 , , , , , and
, , , , , and 
Abstract
:
1. Introduction
2. Results
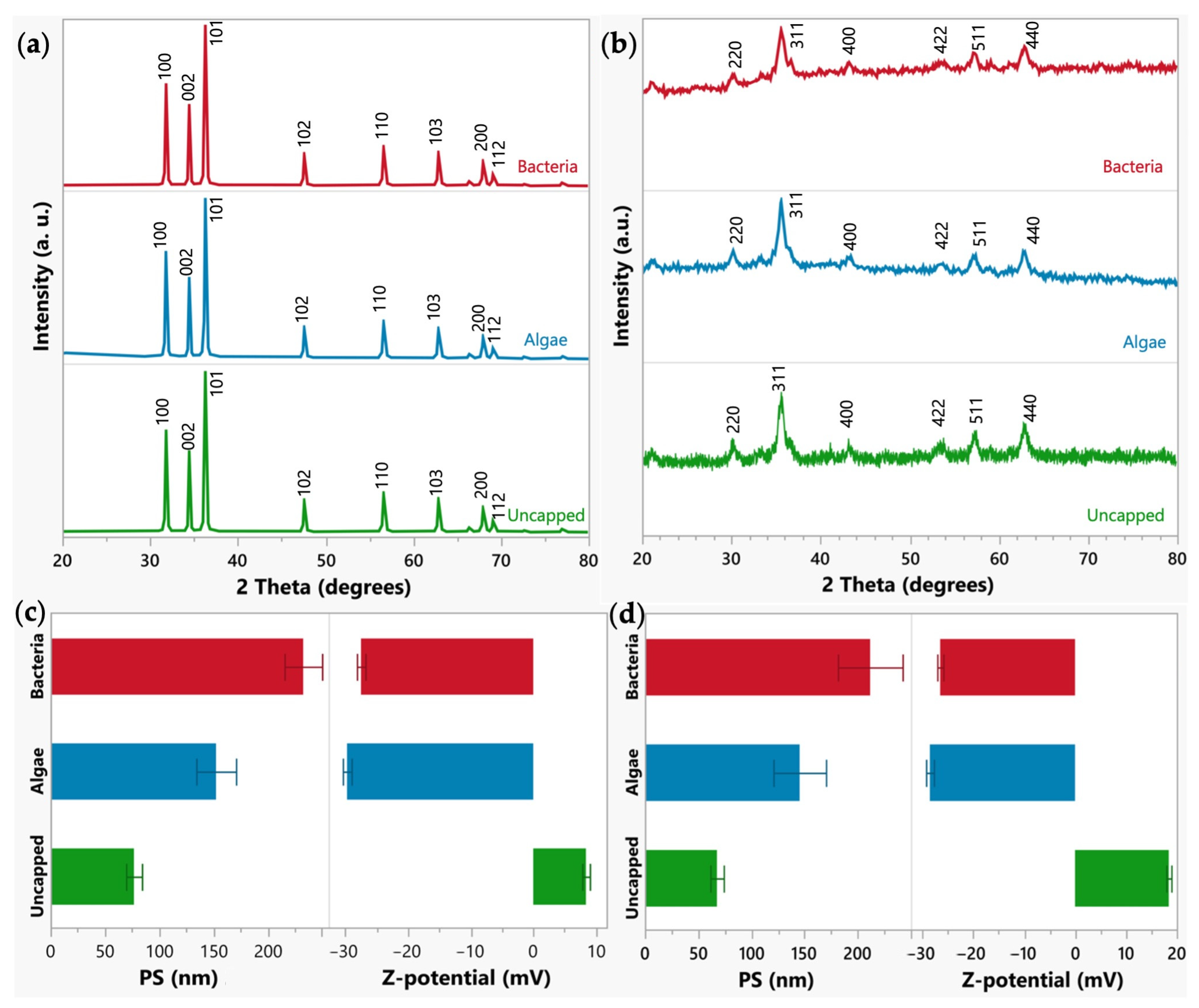
2.1. Synthesis and Characterization of Zinc and Iron Nanoparticles
2.1.1. Crystal Structure and Size Conformation of the Zinc and Iron NPs
2.1.2. Size (nm) and Zeta Potential (mV) of the NPs
2.1.3. Surface Morphology and Elemental Composition of the Uncapped Zinc and Iron Nanoparticles
2.2. Evaluation of the Plant and Microbial Growth Promotion/Inhibition Effects of the Zinc and Iron Nanoformulations
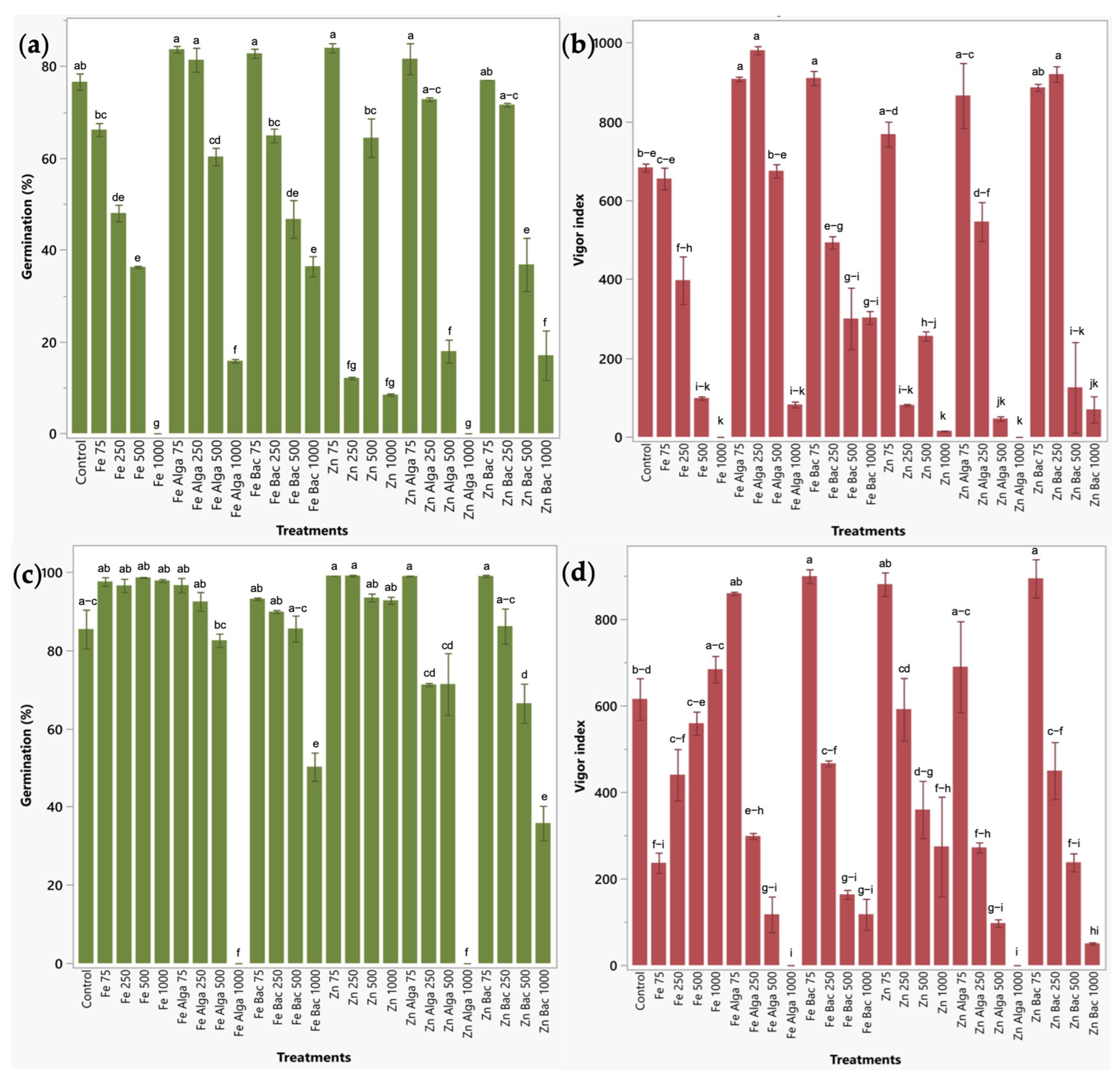
2.2.1. Effect of Zinc and Iron Bio-Nanoformulations on the Germination of Two Relevant Food-Crops
2.2.2. Impact of Bio-Nanoformulations on the Growth of Soil Microorganisms
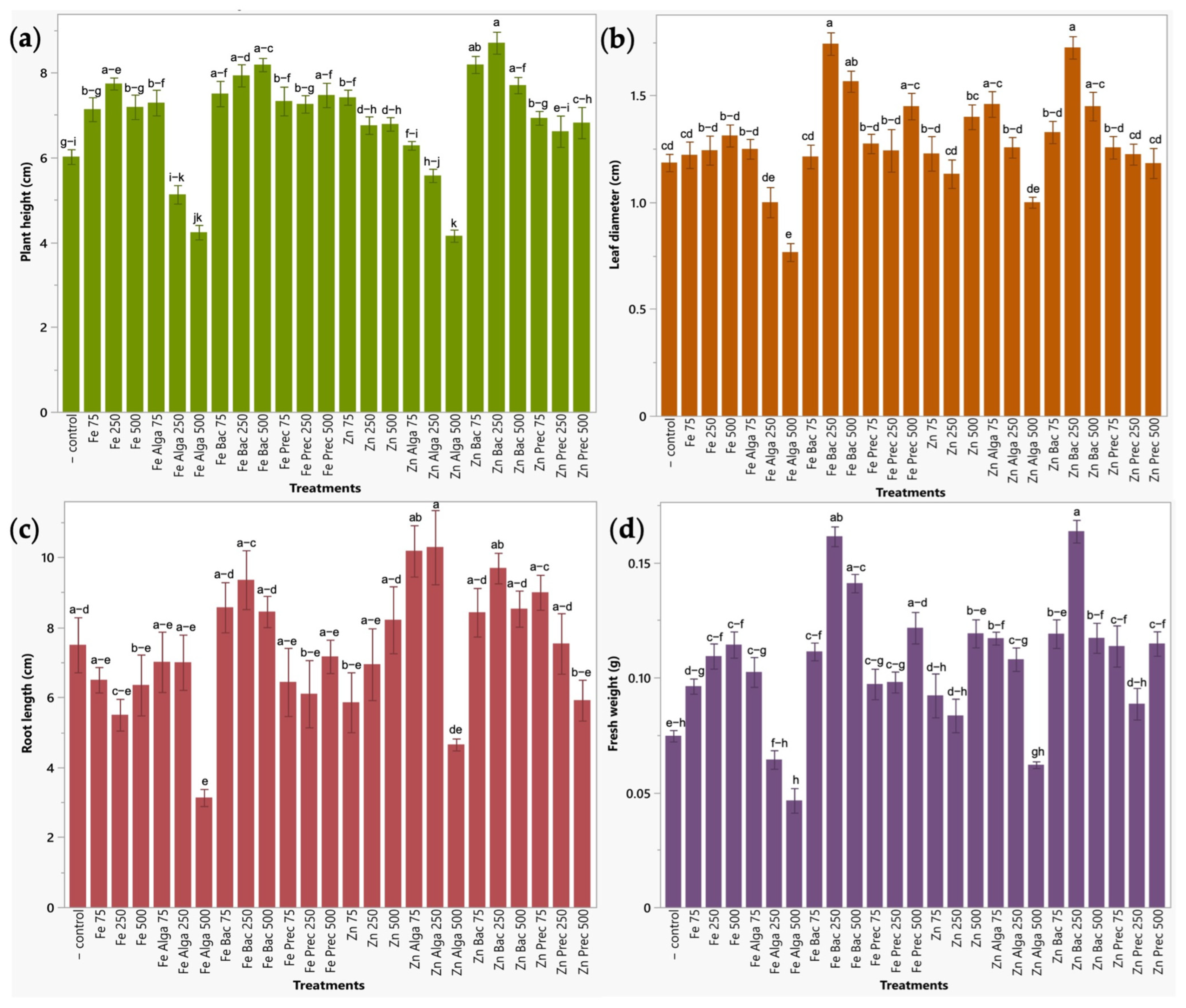
2.2.3. Early Plant Response to Bio-Nanoformulations in a Seedbed Assay
3. Discussion
4. Materials and Methods
4.1. Chemicals and Plant Material
4.2. Synthesis of Zinc and Iron Nanoparticles
4.3. Functionalization of Nanoparticles with Organic Substrates
4.3.1. Preparation of Biological Substrates
4.3.2. Surface Capping of Zinc and Iron Nanoparticles
4.4. Characterization of Zinc and Iron Nanoparticles
4.4.1. X-ray Diffraction (XRD)
4.4.2. Dynamic Light Scattering (DLS)
4.4.3. Electron Microscopy
4.5. In Vivo Evaluation of Zinc and Iron Nanoformulations
4.5.1. Effects on Seed Germination of Relevant Food Crops
4.5.2. Effects on the Growth of Soil Microorganisms
4.5.3. Impact on Early Plant Development in a Seedbed Assay
4.5.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Assunção, A.G.; Cakmak, I.; Clemens, S.; González-Guerrero, M.; Nawrocki, A.; Thomine, S. Micronutrient homeostasis in plants for more sustainable agriculture and healthier human nutrition. J. Exp. Bot. 2022, 73, 1789–1799. [Google Scholar] [CrossRef]
- Dhaliwal, S.S.; Sharma, V.; Shukla, A.K.; Verma, V.; Kaur, M.; Shivay, Y.S.; Nisar, S.; Gaber, A.; Brestic, M.; Barek, V.; et al. Biofortification—A frontier novel approach to enrich micronutrients in field crops to encounter the nutritional security. Molecules 2022, 27, 1340. [Google Scholar] [CrossRef]
- Saraiva, R.; Ferreira, Q.; Rodrigues, G.C.; Oliveira, M. Nanofertilizer use for adaptation and mitigation of the agriculture/Climate change dichotomy effects. Climate 2023, 11, 129. [Google Scholar] [CrossRef]
- Azam, M.; Bhatti, H.N.; Khan, A.; Zafar, L.; Iqbal, M. Zinc oxide nano-fertilizer application (foliar and soil) effect on the growth, photosynthetic pigments and antioxidant system of maize cultivar. Biocatal. Agric. Biotechnol. 2022, 42, 102343. [Google Scholar] [CrossRef]
- Ciurli, A.; Giagnoni, L.; Pastorelli, R.; Sega, D.; Zamboni, A.; Renella, G.; Varanini, Z. A novel P nanofertilizer has no impacts on soil microbial communities and soil microbial activity. Appl. Soil Ecol. 2022, 178, 104570. [Google Scholar] [CrossRef]
- Guardiola-Márquez, C.E.; Santos-Ramírez, M.T.; Segura-Jiménez, M.E.; Figueroa-Montes, M.L.; Jacobo-Velázquez, D.A. Fighting obesity-related micronutrient deficiencies through biofortification of agri-food crops with sustainable fertilization practices. Plants 2022, 11, 3477. [Google Scholar] [CrossRef]
- Salama, D.M.; Abd El-Aziz, M.E.; Shaaban, E.A.; Osman, S.A.; Abd El-Wahed, M.S. The impact of nanofertilizer on agro-morphological criteria, yield, and genomic stability of common bean (Phaseolus vulgaris L.). Sci. Rep. 2022, 12, 18552. [Google Scholar] [CrossRef]
- Verma, K.K.; Song, X.P.; Joshi, A.; Rajput, V.D.; Singh, M.; Sharma, A.; Singh, R.K.; Li, D.M.; Arora, J.; Minkina, T.; et al. Nanofertilizer possibilities for healthy soil, water, and food in future: An overview. Front. Plant Sci. 2022, 13, 865048. [Google Scholar] [CrossRef]
- Banerjee, S.; Roy, P.; Nandi, S.; Roy, S. Advanced biotechnological strategies towards the development of crops with enhanced micronutrient content. Plant Growth Regul. 2023, 100, 355–371. [Google Scholar] [CrossRef]
- El-Ramady, H.; Prokisch, J.; El-Baily, S.; Elasawi, T.H.; Elmahrouk, M.; Omara, A.E.D.; Elsakhawy, T.A.; Amer, M.M.; Brevik, E. Biological nanofertilizer for horticultural crops: A diagrammatic mini-review. Env. Biodivers. Soil Secur. 2022, 6, 339–348. [Google Scholar] [CrossRef]
- Manivannan, N.; Aswathy, S.; Malaikozhundan, B.; Boopathi, T. Nano-zinc oxide synthesized using diazotrophic Azospirillum improves the growth of mung bean, Vigna radiata. Int. Nano Lett. 2021, 11, 405–415. [Google Scholar] [CrossRef]
- Elbasuney, S.; El-Sayyad, G.S.; Attia, M.S.; Abdelaziz, A.M. Ferric oxide colloid: Towards green nano-fertilizer for tomato plant with enhanced vegetative growth and immune response against fusarium wilt disease. J. Inorg. Organomet. Polym. Mater. 2022, 32, 4270–4283. [Google Scholar] [CrossRef] [PubMed]
- Jaiswal, D.K.; Krishna, R.; Chouhan, G.K.; de Araujo Pereira, A.P.; Ade, A.B.; Prakash, S.; Verma, J.P. Bio-fortification of minerals in crops: Current scenario and future prospects for sustainable agriculture and human health. J. Plant Growth Regul. 2022, 98, 5–22. [Google Scholar] [CrossRef]
- El-Bialy, S.M.; El-Mahrouk, M.E.; Elesawy, T.; Omara, A.E.D.; Elbehiry, F.; El-Ramady, H.; Áron, B.; Prokisch, J.; Brevik, E.C.; Solberg, S.Ø. Biological nanofertilizers to enhance growth potential of strawberry seedlings by boosting photosynthetic pigments, plant enzymatic antioxidants, and nutritional status. Plants 2023, 12, 302. [Google Scholar] [CrossRef] [PubMed]
- Shalaby, T.A.; El-Bialy, S.M.; El-Mahrouk, M.E.; Omara, A.E.D.; El-Beltagi, H.S.; El-Ramady, H. Acclimatization of in vitro banana seedlings using root-applied bio-nanofertilizer of copper and selenium. Agronomy 2022, 12, 539. [Google Scholar] [CrossRef]
- Nongbet, A.; Mishra, A.K.; Mohanta, Y.K.; Mahanta, S.; Ray, M.K.; Khan, M.; Baek, K.-H.; Chakrabartty, I. Nanofertilizers: A smart and sustainable attribute to modern agriculture. Plants 2022, 11, 2587. [Google Scholar] [CrossRef]
- Guardiola-Márquez, C.E.; Jacobo-Velázquez, D.A. Potential of enhancing anti-obesogenic agriceuticals by applying sustainable fertilizers during plant cultivation. Front. Sustain. Food Syst. 2022, 6, 1034521. [Google Scholar] [CrossRef]
- De La Torre-Roche, R.; Cantu, J.; Tamez, C.; Zuverza-Mena, N.; Hamdi, H.; Adisa, I.O.; Elmer, W.; Gardea-Torresdey, J.; White, J.C. Seed biofortification by engineered nanomaterials: A pathway to alleviate malnutrition? J. Agric. Food Chem. 2020, 68, 12189–12202. [Google Scholar] [CrossRef]
- El-Ramady, H.; Abdalla, N.; Sári, D.; Ferroudj, A.; Muthu, A.; Prokisch, J.; Fawzy, Z.F.; Brevik, E.C.; Solberg, S.Ø. Nanofarming: Promising solutions for the future of the global agricultural industry. Agronomy 2023, 13, 1600. [Google Scholar] [CrossRef]
- Guardiola-Márquez, C.E.; Santos-Ramírez, M.T.; Figueroa-Montes, M.L.; Valencia-de los Cobos, E.O.; Stamatis-Félix, I.J.; Navarro-López, D.E.; Jacobo-Velázquez, D.A. Identification and characterization of beneficial soil microbial strains for the formulation of biofertilizers based on native plant growth-promoting microorganisms isolated from northern Mexico. Plants 2023, 12, 3262. [Google Scholar] [CrossRef]
- Mącik, M.; Gryta, A.; Frąc, M. Biofertilizers in agriculture: An overview on concepts, strategies and effects on soil microorganisms. Adv. Agron. 2020, 162, 31–87. [Google Scholar] [CrossRef]
- Rahman, M.M.; Hosano, N.; Hosano, H. Recovering microalgal bioresources: A review of cell disruption methods and extraction technologies. Molecules 2022, 27, 2786. [Google Scholar] [CrossRef]
- Käferböck, A.; Smetana, S.; de Vos, R.; Schwarz, C.; Toepfl, S.; Parniakov, O. Sustainable extraction of valuable components from Spirulina assisted by pulsed electric fields technology. Algal Res. 2020, 48, 101914. [Google Scholar] [CrossRef]
- Etape, E.P.; Foba-Tendo, J.; Ngolui, L.J.; Namondo, B.V.; Yollande, F.C.; Nguimezong, M.B.N. Structural characterization and magnetic properties of undoped and Ti-doped ZnO nanoparticles prepared by modified oxalate route. J. Nanomater. 2018, 2018, 9072325. [Google Scholar] [CrossRef]
- Aliahmad, M.; Nasiri Moghaddam, N. Synthesis of maghemite (γ-Fe2O3) nanoparticles by thermal-decomposition of magnetite (Fe3O4) nanoparticles. Mater. Sci.-Pol. 2013, 31, 264–268. [Google Scholar] [CrossRef]
- Mahmoud, A.W.M.; Rashad, H.M.; Esmail, S.E.; Alsamadany, H.; Abdeldaym, E.A. Application of silicon, zinc, and zeolite nanoparticles—A tool to enhance drought stress tolerance in coriander plants for better growth performance and productivity. Plants 2023, 12, 2838. [Google Scholar] [CrossRef]
- Zulfiqar, F.; Navarro, M.; Ashraf, M.; Akram, N.A.; Munné-Bosch, S. Nanofertilizer use for sustainable agriculture: Advantages and limitations. Plant Sci. 2019, 289, 110270. [Google Scholar] [CrossRef]
- Behl, T.; Kaur, I.; Sehgal, A.; Singh, S.; Sharma, N.; Bhatia, S.; Al-Harrasi, A.; Bungau, S. The dichotomy of nanotechnology as the cutting edge of agriculture: Nano-farming as an asset versus nanotoxicity. Chemosphere 2022, 288, 132533. [Google Scholar] [CrossRef]
- Yusefi-Tanha, E.; Fallah, S.; Rostamnejadi, A.; Pokhrel, L.R. Zinc oxide nanoparticles (ZnONPs) as a novel nanofertilizer: Influence on seed yield and antioxidant defense system in soil grown soybean (Glycine max Cv. Kowsar). Sci. Total Environ. 2020, 738, 140240. [Google Scholar] [CrossRef]
- LaGrow, A.P.; Besenhard, M.O.; Hodzic, A.; Sergides, A.; Bogart, L.K.; Gavriilidis, A.; Thanh, N.T.K. Unravelling the growth mechanism of the co-precipitation of iron oxide nanoparticles with the aid of synchrotron X-Ray diffraction in solution. Nanoscale 2019, 11, 6620–6628. [Google Scholar] [CrossRef]
- Afzal, S.; Sharma, D.; Singh, N.K. Eco-friendly synthesis of phytochemical-capped iron oxide nanoparticles as nano-priming agent for boosting seed germination in rice (Oryza sativa L.). Environ. Sci. Pollut. Res. 2021, 28, 40275–40287. [Google Scholar] [CrossRef] [PubMed]
- Singh, J.; Kumar, S.; Alok, A.; Upadhyay, S.K.; Rawat, M.; Tsang, D.C.; Bolan, N.; Kim, K.H. The potential of green synthesized zinc oxide nanoparticles as nutrient source for plant growth. J. Clean. Prod. 2019, 214, 1061–1070. [Google Scholar] [CrossRef]
- Asmat-Campos, D.; López-Medina, E.; Montes de Oca-Vásquez, G.; Gil-Rivero, E.; Delfín-Narciso, D.; Juárez-Cortijo, L.; Villena-Zapata, L.; Gurreonero-Fernández, J.; Rafael-Amaya, R. ZnO Nanoparticles obtained by green synthesis as an alternative to improve the germination characteristics of L. esculentum. Molecules 2022, 27, 2343. [Google Scholar] [CrossRef]
- Itroutwar, P.D.; Kasivelu, G.; Raguraman, V.; Malaichamy, K.; Sevathapandian, S.K. Effects of biogenic zinc oxide nanoparticles on seed germination and seedling vigor of maize (Zea mays). Biocatal. Agric. Biotechnol. 2020, 29, 101778. [Google Scholar] [CrossRef]
- Iannone, M.F.; Groppa, M.D.; de Sousa, M.E.; van Raap, M.B.F.; Benavides, M.P. Impact of magnetite iron oxide nanoparticles on wheat (Triticum aestivum L.) development: Evaluation of oxidative damage. Environ. Exp. Bot. 2016, 131, 77–88. [Google Scholar] [CrossRef]
- Li, J.; Hu, J.; Ma, C.; Wang, Y.; Wu, C.; Huang, J.; Xing, B. Uptake, translocation and physiological effects of magnetic iron oxide (γ-Fe2O3) nanoparticles in corn (Zea mays L.). Chemosphere 2016, 159, 326–334. [Google Scholar] [CrossRef]
- Montoya-Giraldo, Y.A.; Portela-Dávila, A.R.; Susunaga-Gómez, D.; Urquijo-Morales, J.P.; Velásquez-Torres, Á.A.; Villanueva-Mejía, D.F. In vitro germination and growth of maize and lettuce seeds exposed to iron oxide magnetic nanoparticles. Dyna 2023, 90, 9–13. [Google Scholar] [CrossRef]
- Rai-Kalal, P.; Jajoo, A. Priming with zinc oxide nanoparticles improve germination and photosynthetic performance in wheat. Plant Physiol. Biochem. 2021, 160, 341–351. [Google Scholar] [CrossRef]
- Sharma, D.; Afzal, S.; Singh, N.K. Nanopriming with phytosynthesized zinc oxide nanoparticles for promoting germination and starch metabolism in rice seeds. J. Biotechnol. 2021, 336, 64–75. [Google Scholar] [CrossRef]
- Tovar, G.I.; Briceño, S.; Suarez, J.; Flores, S.; González, G. Biogenic synthesis of iron oxide nanoparticles using Moringa oleifera and chitosan and its evaluation on corn germination. Environ. Nanotechnol. Monit. Manag. 2020, 14, 100350. [Google Scholar] [CrossRef]
- Yang, Y.; Li, R.; Wang, L.; Bi, H.; Peng, Z.; Li, J.; Guo, C.; Bi, Y.; Lai, Y.; Guo, D. Enhanced germination and growth of alfalfa with seed presoaking and hydroponic culture in Fe2O3 magnetic nanoparticles. J. Nanomater. 2023, 2023, 9783977. [Google Scholar] [CrossRef]
- Domínguez-Castillo, C.; Alatorre-Cruz, J.M.; Castañeda-Antonio, D.; Munive, J.A.; Guo, X.; López-Olguín, J.F.; Fuentes-Ramírez, L.E.; Carreño-López, R. Potential seed germination-enhancing plant growth-promoting rhizobacteria for restoration of Pinus chiapensis ecosystems. J. For. Res. 2021, 32, 2143–2153. [Google Scholar] [CrossRef]
- Hardiansyah, M.Y.; Musa, Y.; Jaya, A.M. Bio-priming seeds with PGPR of bamboo rhizosphere in cocoa (Theobroma cacao L.) seeds germination. Int. J. Sci. Res. Biol. Sci. 2020, 7, 11–18. [Google Scholar]
- Sedefoglu, N.; Zalaoglu, Y.; Bozok, F. Green synthesized ZnO nanoparticles using Ganoderma lucidum: Characterization and in vitro nanofertilizer effects. J. Alloys Compd. 2022, 918, 165695. [Google Scholar] [CrossRef]
- Alavi, M.; Nokhodchi, A. Synthesis and modification of bio-derived antibacterial Ag and ZnO nanoparticles by plants, fungi, and bacteria. Drug Discov. Today 2021, 26, 1953–1962. [Google Scholar] [CrossRef] [PubMed]
- Gudkov, S.V.; Burmistrov, D.E.; Serov, D.A.; Rebezov, M.B.; Semenova, A.A.; Lisitsyn, A.B. Do iron oxide nanoparticles have significant antibacterial properties? Antibiotics 2021, 10, 884. [Google Scholar] [CrossRef] [PubMed]
- Gudkov, S.V.; Burmistrov, D.E.; Serov, D.A.; Rebezov, M.B.; Semenova, A.A.; Lisitsyn, A.B. A mini review of antibacterial properties of ZnO nanoparticles. Front. Phys. 2021, 9, 641481. [Google Scholar] [CrossRef]
- Pillai, A.M.; Sivasankarapillai, V.S.; Rahdar, A.; Joseph, J.; Sadeghfar, F.; Rajesh, K.; Kyzas, G.Z. Green synthesis and characterization of zinc oxide nanoparticles with antibacterial and antifungal activity. J. Mol. Struct. 2020, 1211, 128107. [Google Scholar] [CrossRef]
- Ogunyemi, S.O.; Abdallah, Y.; Zhang, M.; Fouad, H.; Hong, X.; Ibrahim, E.; Masum, M.M.I.; Hossain, A.; Mo, J.; Li, B. Green synthesis of zinc oxide nanoparticles using different plant extracts and their antibacterial activity against Xanthomonas oryzae pv. oryzae. Artif. Cells Nanomed. Biotechnol. 2019, 47, 341–352. [Google Scholar] [CrossRef]
- Wei, X.; Cao, P.; Wang, G.; Liu, Y.; Song, J.; Han, J. CuO, ZnO, and γ-Fe2O3 nanoparticles modified the underground biomass and rhizosphere microbial community of Salvia miltiorrhiza (Bge.) after 165-day exposure. Ecotoxicol. Environ. Saf. 2021, 217, 112232. [Google Scholar] [CrossRef]
- Szabo, R.; Bodolea, C.; Mocan, T. Iron, copper, and zinc homeostasis: Physiology, physiopathology, and nanomediated applications. Nanomaterials 2021, 11, 2958. [Google Scholar] [CrossRef] [PubMed]
- Lv, J.; Christie, P.; Zhang, S. Uptake, translocation, and transformation of metal-based nanoparticles in plants: Recent advances and methodological challenges. Environ. Sci. Nano 2019, 6, 41–59. [Google Scholar] [CrossRef]
- Rastogi, A.; Zivcak, M.; Sytar, O.; Kalaji, H.M.; He, X.; Mbarki, S.; Brestic, M. Impact of metal and metal oxide nanoparticles on plant: A critical review. Front. Chem. 2017, 5, 78. [Google Scholar] [CrossRef]
- Hong, J.; Wang, C.; Wagner, D.C.; Gardea-Torresdey, J.L.; He, F.; Rico, C.M. Foliar application of nanoparticles: Mechanisms of absorption, transfer, and multiple impacts. Environ. Sci. Nano 2021, 8, 1196–1210. [Google Scholar] [CrossRef]
- Lin, D.; Xing, B. Phytotoxicity of nanoparticles: Inhibition of seed germination and root growth. Environ. Pollut. 2007, 150, 243–250. [Google Scholar] [CrossRef]
- Koç, E.; Karayiğit, B. Assessment of biofortification approaches used to improve micronutrient-dense plants that are a sustainable solution to combat hidden hunger. J. Soil Sci. Plant Nutr. 2022, 22, 475–500. [Google Scholar] [CrossRef] [PubMed]
- Miri, A.; Khatami, M.; Sarani, M. Biosynthesis, magnetic and cytotoxic studies of hematite nanoparticles. J. Inorg. Organomet. Polym. Mater. 2020, 30, 767–774. [Google Scholar] [CrossRef]
- Mydeen, S.S.; Kumar, R.R.; Kottaisamy, M.; Vasantha, V.S. Biosynthesis of ZnO nanoparticles through extract from Prosopis juliflora plant leaf: Antibacterial activities and a new approach by rust-induced photocatalysis. J. Saudi Chem. Soc. 2020, 24, 393–406. [Google Scholar] [CrossRef]
- Puglisi, I.; Barone, V.; Fragalà, F.; Stevanato, P.; Baglieri, A.; Vitale, A. Effect of microalgal extracts from Chlorella vulgaris and Scenedesmus quadricauda on germination of Beta vulgaris seeds. Plants 2020, 9, 675. [Google Scholar] [CrossRef]
- Vasantharaj, S.; Sathiyavimal, S.; Senthilkumar, P.; LewisOscar, F.; Pugazhendhi, A. Biosynthesis of iron oxide nanoparticles using leaf extract of Ruellia tuberosa: Antimicrobial properties and their applications in photocatalytic degradation. J. Photochem. Photobiol. B Biol. 2019, 192, 74–82. [Google Scholar] [CrossRef]
- Raliya, R.; Biswas, P.; Tarafdar, J.C. TiO2 nanoparticle biosynthesis and its physiological effect on mung bean (Vigna radiata L.). Biotechnol. Rep. 2015, 5, 22–26. [Google Scholar] [CrossRef] [PubMed]
- Khanam, J.; Ahmed, M.F.; Zaman, S.K.M.; Sharmin, N.; Ahmed, S. Synthesis of nano-sized magnetic iron oxide by a simple and facile co-precipitation method. Bangladesh J. Sci. Ind. Res. 2022, 57, 67–76. [Google Scholar] [CrossRef]
- Sheikholeslami, Z.; Kebria, D.Y.; Qaderi, F. Application of γ-Fe2O3 nanoparticles for pollution removal from water with visible light. J. Mol. Liq. 2020, 299, 112118. [Google Scholar] [CrossRef]
- Kotresh, M.G.; Patil, M.K.; Inamdar, S.R. Reaction temperature based synthesis of ZnO nanoparticles using co-precipitation method: Detailed structural and optical characterization. Optik 2021, 243, 167506. [Google Scholar] [CrossRef]
- Adam, R.E.; Pozina, G.; Willander, M.; Nur, O. Synthesis of ZnO nanoparticles by co-precipitation method for solar driven photodegradation of Congo red dye at different pH. Photonics Nanostruct. Fundam. Appl. 2018, 32, 11–18. [Google Scholar] [CrossRef]
- Nemer, G.; Louka, N.; Vorobiev, E.; Salameh, D.; Nicaud, J.M.; Maroun, R.G.; Koubaa, M. Mechanical cell disruption technologies for the extraction of dyes and pigments from microorganisms: A review. Fermentation 2021, 7, 36. [Google Scholar] [CrossRef]
- Mata, S.N.; de Souza Santos, T.; Cardoso, L.G.; Andrade, B.B.; Duarte, J.H.; Costa, J.A.V.; de Souza, C.O.; Druzian, J.I. Spirulina sp. LEB 18 cultivation in a raceway-type bioreactor using wastewater from desalination process: Production of carbohydrate-rich biomass. Bioresour. Technol. 2020, 311, 123495. [Google Scholar] [CrossRef]
- Hamouda, R.A.; Hussein, M.H.; Elhadary, A.M.A.; Abuelmagd, M.A. Extruded polysaccharide/protein matrix from Arthrospira platensis cultures mediated silver nanoparticles biosynthesis and capping. Appl. Nanosci. 2020, 10, 3839–3855. [Google Scholar] [CrossRef]
- Bulut, O.; Akın, D.; Sönmez, Ç.; Öktem, A.; Yücel, M.; Öktem, H.A. Phenolic compounds, carotenoids, and antioxidant capacities of a thermo-tolerant Scenedesmus sp. (Chlorophyta) extracted with different solvents. J. Appl. Phycol. 2019, 31, 1675–1683. [Google Scholar] [CrossRef]
- Chaudhuri, S.K.; Malodia, L. Biosynthesis of zinc oxide nanoparticles using leaf extract of Calotropis gigantea: Characterization and its evaluation on tree seedling growth in nursery stage. Appl. Nanosci. 2017, 7, 501–512. [Google Scholar] [CrossRef]
- Azeez, L.; Lateef, A.; Adejumo, A.L.; Adeleke, J.T.; Adetoro, R.O.; Mustapha, Z. Adsorption behaviour of rhodamine B on hen feather and corn starch functionalized with green synthesized silver nanoparticles (AgNPs) mediated with cocoa pods extracts. Chem. Afr. 2020, 3, 237–250. [Google Scholar] [CrossRef]
- Vivekanandhan, S.; Schreiber, M.; Mason, C.; Mohanty, A.K.; Misra, M. Maple leaf (Acer sp.) extract mediated green process for the functionalization of ZnO powders with silver nanoparticles. Colloids Surf. B 2014, 113, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Farghaly, F.A.; Nafady, N.A. Green synthesis of silver nanoparticles using leaf extract of Rosmarinus officinalis and its effect on tomato and wheat plants. J. Agric. Sci. 2015, 7, 277. [Google Scholar] [CrossRef]
- Salih, A.M.; Al-Qurainy, F.; Khan, S.; Tarroum, M.; Nadeem, M.; Shaikhaldein, H.O.; Gaafar, A.R.Z.; Alfarraj, N.S. Biosynthesis of zinc oxide nanoparticles using Phoenix dactylifera and their effect on biomass and phytochemical compounds in Juniperus procera. Sci. Rep. 2021, 11, 19136. [Google Scholar] [CrossRef] [PubMed]
- Mazumder, J.A.; Khan, E.; Perwez, M.; Gupta, M.; Kumar, S.; Raza, K.; Sardar, M. Exposure of biosynthesized nanoscale ZnO to Brassica juncea crop plant: Morphological, biochemical and molecular aspects. Sci. Rep. 2020, 10, 8531. [Google Scholar] [CrossRef]
- Radini, I.A.; Hasan, N.; Malik, M.A.; Khan, Z. Biosynthesis of iron nanoparticles using Trigonella foenum-graecum seed extract for photocatalytic methyl orange dye degradation and antibacterial applications. J. Photochem. Photobiol. B Biol. 2018, 183, 154–163. [Google Scholar] [CrossRef]
- Boutchuen, A.; Zimmerman, D.; Aich, N.; Masud, A.M.; Arabshahi, A.; Palchoudhury, S. Increased plant growth with hematite nanoparticle fertilizer drop and determining nanoparticle uptake in plants using multimodal approach. J. Nanomater. 2019, 2019, 6890572. [Google Scholar] [CrossRef]
- García-López, J.I.; Niño-Medina, G.; Olivares-Sáenz, E.; Lira-Saldivar, R.H.; Barriga-Castro, E.D.; Vázquez-Alvarado, R.; Rodríguez-Salinas, P.A.; Zavala-García, F. Foliar application of zinc oxide nanoparticles and zinc sulfate boosts the content of bioactive compounds in habanero peppers. Plants 2019, 8, 254. [Google Scholar] [CrossRef]
- Palmqvist, N.G.M.; Bejai, S.; Meijer, J.; Seisenbaeva, G.A.; Kessler, V.G. Nano titania aided clustering and adhesion of beneficial bacteria to plant roots to enhance crop growth and stress management. Sci. Rep. 2015, 5, 10146. [Google Scholar] [CrossRef]
- Szymanski, M.; Dobrucka, R. Evaluation of phytotoxicity of bimetallic Ag/Au nanoparticles synthesized using Geum urbanum L. J. Inorg. Organomet. Polym. Mater. 2021, 31, 2459–2470. [Google Scholar] [CrossRef]
- Win, T.T.; Khan, S.; Bo, B.; Zada, S.; Fu, P. Green synthesis and characterization of Fe3O4 nanoparticles using Chlorella-K01 extract for potential enhancement of plant growth stimulating and antifungal activity. Sci. Rep. 2021, 11, 21996. [Google Scholar] [CrossRef] [PubMed]
- Acharya, P.; Jayaprakasha, G.K.; Crosby, K.M.; Jifon, J.L.; Patil, B.S. Nanoparticle-mediated seed priming improves germination, growth, yield, and quality of watermelons (Citrullus lanatus) at multi-locations in Texas. Sci. Rep. 2020, 10, 5037. [Google Scholar] [CrossRef] [PubMed]
- Brasili, E.; Bavasso, I.; Petruccelli, V.; Vilardi, G.; Valletta, A.; Dal Bosco, C.; Gentili, A.; Pasqua, G.; Di Palma, L. Remediation of hexavalent chromium contaminated water through zero-valent iron nanoparticles and effects on tomato plant growth performance. Sci. Rep. 2020, 10, 1920. [Google Scholar] [CrossRef]
- Kornarzyński, K.; Sujak, A.; Czernel, G.; Wiącek, D. Effect of Fe3O4 nanoparticles on germination of seeds and concentration of elements in Helianthus annuus L. under constant magnetic field. Sci. Rep. 2020, 10, 8068. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.J.; Chung, S.G.; Lee, S.H.; Choi, J.W. Relation of microbial biomass to counting units for Pseudomonas aeruginosa. Afr. J. Microbiol. Res. 2012, 6, 4620–4622. [Google Scholar] [CrossRef]
- Balouiri, M.; Sadiki, M.; Ibnsouda, S.K. Methods for in vitro evaluating antimicrobial activity: A review. J. Pharm. Anal. 2016, 6, 71–79. [Google Scholar] [CrossRef]
- Saboor, A.; Ali, M.A.; Danish, S.; Ahmed, N.; Fahad, S.; Datta, R.; Ansari, M.J.; Nasif, O.; Rahman, M.H.U.; Glick, B.R. Effect of arbuscular mycorrhizal fungi on the physiological functioning of maize under zinc-deficient soils. Sci. Rep. 2021, 11, 18468. [Google Scholar] [CrossRef] [PubMed]
- Noolu, V.; Mudavath, H.; Pillai, R.J.; Yantrapalli, S.K. Permanent deformation behaviour of black cotton soil treated with calcium carbide residue. Constr. Build. Mater. 2019, 223, 441–449. [Google Scholar] [CrossRef]
- Bokhari, A.; Essack, M.; Lafi, F.F.; Andres-Barrao, C.; Jalal, R.; Alamoudi, S.; Razali, R.; Alzubaidy, H.; Shah, K.H.; Siddique, S.; et al. Bioprospecting desert plant Bacillus endophytic strains for their potential to enhance plant stress tolerance. Sci. Rep. 2019, 9, 18154. [Google Scholar] [CrossRef]
- Jiménez-Gómez, A.; Flores-Félix, J.D.; García-Fraile, P.; Mateos, P.F.; Menéndez, E.; Velázquez, E.; Rivas, R. Probiotic activities of Rhizobium laguerreae on growth and quality of spinach. Sci. Rep. 2018, 8, 295. [Google Scholar] [CrossRef]
- Kuan, K.B.; Othman, R.; Abdul Rahim, K.; Shamsuddin, Z.H. Plant growth-promoting rhizobacteria inoculation to enhance vegetative growth, nitrogen fixation and nitrogen remobilisation of maize under greenhouse conditions. PLoS ONE 2016, 11, e0152478. [Google Scholar] [CrossRef] [PubMed]
- Afanador-Barajas, L.N.; Navarro-Noya, Y.E.; Luna-Guido, M.L.; Dendooven, L. Impact of a bacterial consortium on the soil bacterial community structure and maize (Zea mays L.) cultivation. Sci. Rep. 2021, 11, 13092. [Google Scholar] [CrossRef] [PubMed]
- Magotra, S.; Bhagat, N.; Ambardar, S.; Ali, T.; Hurek, B.R.; Hurek, T.; Verma, P.K.; Vakhlu, J. Field evaluation of PGP Bacillus sp. strain D5 native to Crocus sativus, in traditional and non traditional areas, and mining of PGP genes from its genome. Sci. Rep. 2021, 11, 5454. [Google Scholar] [CrossRef]
- Bononi, L.; Chiaramonte, J.B.; Pansa, C.C.; Moitinho, M.A.; Melo, I.S. Phosphorus-solubilizing Trichoderma spp. from Amazon soils improve soybean plant growth. Sci. Rep. 2020, 10, 2858. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Assay | Objective | Treatments |
|---|---|---|
| Seed germination test | To determine if the biological agents used to formulate bio-nanofertilizers or the NP concentration significantly affect the germination of two important annual food-crops. | Twenty-four treatments corresponding to two micronutrients (iron and zinc), three capping conditions (uncapped or capped with bacteria (Bac) or microalgae (Alga)), and four concentrations (75, 250, 500, 1000 μg mL−1): Zn Bac 75, Zn Bac 250, Zn Bac 500, Zn Bac 1000, Zn Alga 75, Zn Alga 250, Zn Alga 500, Zn Alga 1000, Zn 75, Zn 250, Zn 500, Zn 1000, Fe Bac 75, Fe Bac 250, Fe Bac 500, Fe Bac 1000, Fe Alga 75, Fe Alga 250, Fe Alga 500, Fe Alga 1000, Fe 75, Fe 250, Fe 500, Fe 1000. Control (water-irrigated plants). |
| Soil microbial growth evaluation | To elucidate the nanotoxicity effects of different concentrations of nanoformulations on soil microorganisms. | Eighteen treatments resulting from two micronutrients (iron and zinc), three capping conditions (uncapped or capped with bacteria (Bac) or microalgae (Alga)) and three concentrations (75, 250, 500 μg mL−1): Zn Bac 75, Zn Bac 250, Zn Bac 500, Zn Alga 75, Zn Alga 250, Zn Alga 500, Zn 75, Zn 250, Zn 500, Fe Bac 75, Fe Bac 250, Fe Bac 500, Fe Alga 75, Fe Alga 250, Fe Alga 500, Fe 75, Fe 250, Fe 500. Control (water-irrigated plants). |
| Early plant response in a seedbed assay | To evaluate if the nanoscale of micronutrients, their concentration, and the use of capping agents influence the early plant response of two fast-growing plant species. | Twenty-four treatments corresponding to two micronutrients (iron and zinc), three capping conditions (uncapped or capped with bacteria (Bac) or microalgae (Alga)), two metal precursors (Prec), and three concentrations (75, 250, 500 μg mL−1): Zn Bac 75, Zn Bac 250, Zn Bac 500, Zn Alga 75, Zn Alga 250, Zn Alga 500, Zn 75, Zn 250, Zn 500, Zn Prec 75, Zn Prec 250, Zn Prec 500, Fe Bac 75, Fe Bac 250, Fe Bac 500, Fe Alga 75, Fe Alga 250, Fe Alga 500, Fe 75, Fe 250, Fe 500, Fe Prec 75, Fe Prec 250, Fe Prec 500. Control (water-irrigated plants). |
| Treatment | Initial Count T0 (CFU/mL) | Final Count T1 (CFU/mL) | % of Increase/Reduction of T1 with Respect to T0 * |
|---|---|---|---|
| Zn Bac 75 | 1.93 × 108 | 2.63 × 108 | 39.57 ± 4.53 bc |
| Zn Bac 250 | 2.76 × 108 | 3.52 × 108 | 26.88 ± 1.27 de |
| Zn Bac 500 | 1.85 × 108 | 9.41 × 107 | −49.57 ± 0.79 k |
| Zn Alga 75 | 5.38 × 107 | 7.15 × 107 | 32.90 ± 0.14 cd |
| Zn Alga 250 | 5.49 × 107 | 6.75 × 107 | 23.01 ± 0.08 e |
| Zn Alga 500 | 5.39 × 107 | 3.70 × 107 | −31.05 ± 0.43 ij |
| Zn 75 | 3.76 × 107 | 8.41 × 107 | 123.87 ± 0.05 a |
| Zn 250 | 1.24 × 108 | 1.73 × 108 | 41.13 ± 2.06 b |
| Zn 500 | 4.37 × 107 | 2.15 × 107 | −51.25 ± 0.56 k |
| Fe Bac 75 | 2.01 × 108 | 2.75 × 108 | 36.10 ± 1.04 bc |
| Fe Bac 250 | 2.18 × 108 | 2.66 × 108 | 21.36 ± 1.34 e |
| Fe Bac 500 | 2.09 × 108 | 2.03 × 108 | −3.32 ± 0.30 g |
| Fe Alga 75 | 1.54 × 108 | 2.00 × 108 | 30.32 ± 0.04 d |
| Fe Alga 250 | 1.48 × 108 | 9.87 × 107 | −34.07 ± 1.04 j |
| Fe Alga 500 | 1.49 × 108 | 7.11 × 107 | −52.73 ± 0.87 k |
| Fe 75 | 1.50 × 107 | 1.66 × 107 | 10.46 ± 0.18 f |
| Fe 250 | 2.90 × 107 | 2.13 × 107 | −28.08 ± 2.36 hi |
| Fe 500 | 1.34 × 107 | 1.01 × 107 | −25.55 ± 1.33 h |
| Control | 6.21 × 107 | 6.85 × 107 | 9.95 ± 0.51 f |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guardiola-Márquez, C.E.; López-Mena, E.R.; Segura-Jiménez, M.E.; Gutierrez-Marmolejo, I.; Flores-Matzumiya, M.A.; Mora-Godínez, S.; Hernández-Brenes, C.; Jacobo-Velázquez, D.A. Development and Evaluation of Zinc and Iron Nanoparticles Functionalized with Plant Growth-Promoting Rhizobacteria (PGPR) and Microalgae for Their Application as Bio-Nanofertilizers. Plants 2023, 12, 3657. https://doi.org/10.3390/plants12203657
Guardiola-Márquez CE, López-Mena ER, Segura-Jiménez ME, Gutierrez-Marmolejo I, Flores-Matzumiya MA, Mora-Godínez S, Hernández-Brenes C, Jacobo-Velázquez DA. Development and Evaluation of Zinc and Iron Nanoparticles Functionalized with Plant Growth-Promoting Rhizobacteria (PGPR) and Microalgae for Their Application as Bio-Nanofertilizers. Plants. 2023; 12(20):3657. https://doi.org/10.3390/plants12203657
Chicago/Turabian StyleGuardiola-Márquez, Carlos Esteban, Edgar R. López-Mena, M. Eugenia Segura-Jiménez, Isaac Gutierrez-Marmolejo, Manuel A. Flores-Matzumiya, Shirley Mora-Godínez, Carmen Hernández-Brenes, and Daniel A. Jacobo-Velázquez. 2023. "Development and Evaluation of Zinc and Iron Nanoparticles Functionalized with Plant Growth-Promoting Rhizobacteria (PGPR) and Microalgae for Their Application as Bio-Nanofertilizers" Plants 12, no. 20: 3657. https://doi.org/10.3390/plants12203657