

Machine Learning Analysis of the Impact of Silver Nitrate and Silver Nanoparticles on Wheat (Triticum aestivum L.): Callus Induction, Plant Regeneration, and DNA Methylation

 ,
,  ,
,  , , ,
, , ,  , and
, and 
Abstract
:1. Introduction
2. Results
2.1. Effects of Silver Nitrate and Silver Nanoparticles on In Vitro Parameters
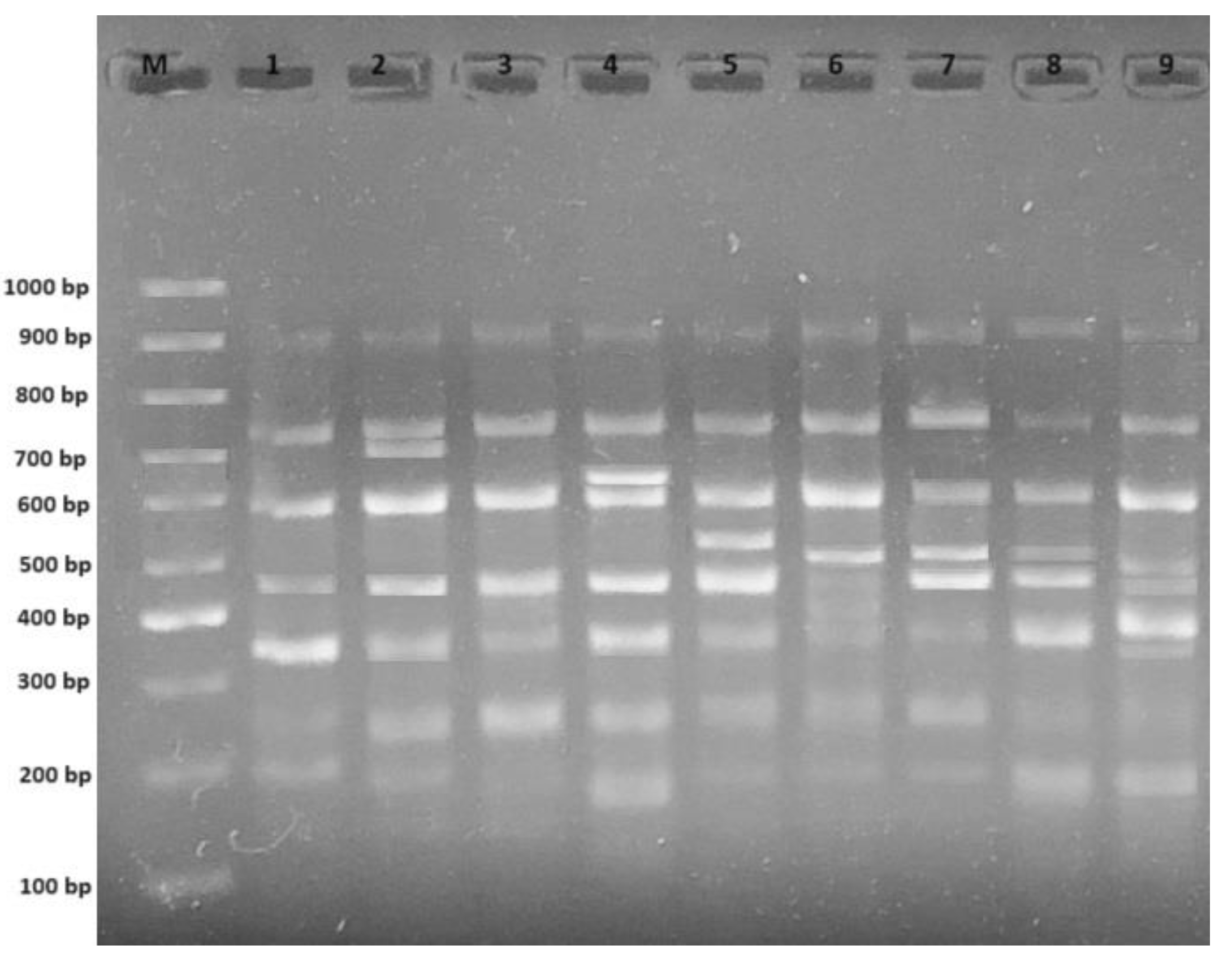
2.2. RAPD Analysis
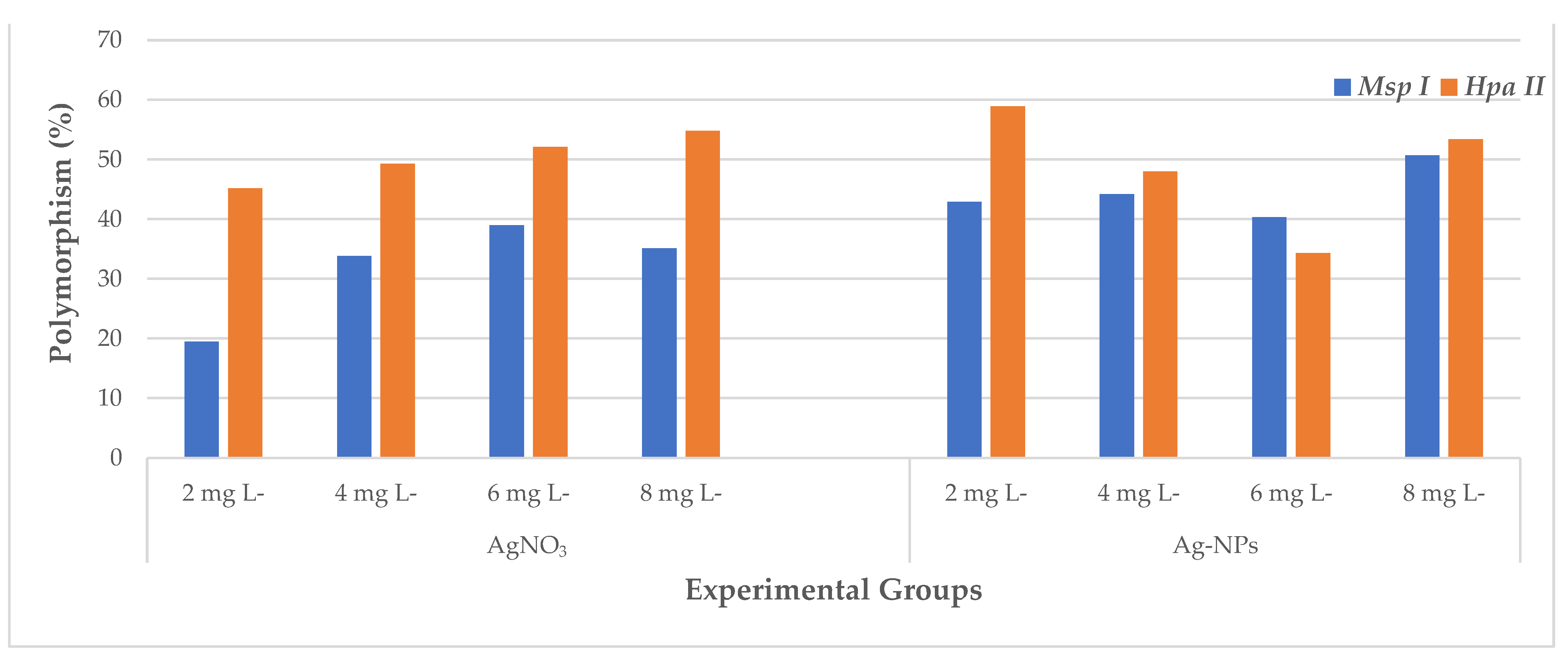
2.3. CRED-RA Analysis
2.4. Machine Learning (ML) Analysis
3. Discussion
4. Materials and Methods
4.1. Synthesis of Silver Nanoparticles (Ag-NPs)
4.2. Plant Material
4.3. In Vitro Conditions
4.4. Molecular Assays
4.4.1. Isolation of Genomic DNA
4.4.2. RAPD and CRED-RA PCR Assays
4.4.3. Genetics Analysis
4.5. Modeling Using Machine Learning Algorithms
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Thorpe, T. History of plant tissue culture. Plant Cell Cult. Protoc. 2012, 877, 9–27. [Google Scholar]
- Kumar, V.; Parvatam, G.; Ravishankar, G.A. AgNO3: A potential regulator of ethylene activity and plant growth modulator. Electron. J. Biotechnol. 2009, 12, 8–9. [Google Scholar] [CrossRef]
- Neves, M.; Correia, S.; Canhoto, J. Ethylene Inhibition Reduces De Novo Shoot Organogenesis and Subsequent Plant Development from Leaf Explants of Solanum betaceum Cav. Plants 2023, 12, 1854. [Google Scholar] [CrossRef] [PubMed]
- Neves, M.; Correia, S.; Cavaleiro, C.; Canhoto, J. Modulation of Organogenesis and Somatic Embryogenesis by Ethylene: An Overview. Plants 2021, 10, 1208. [Google Scholar] [CrossRef] [PubMed]
- Kumari, M.; Mukherjee, A.; Chandrasekaran, N. Genotoxicity of silver nanoparticles in Allium cepa. Sci. Total Environ. 2009, 407, 5243–5246. [Google Scholar] [CrossRef] [PubMed]
- Prem Kumar, G.; Sivakumar, S.; Siva, G.; Vigneswaran, M.; Senthil Kumar, T.; Jayabalan, N. Silver nitrate promotes high-frequency multiple shoot regeneration in cotton (Gossypium hirsutum L.) by inhibiting ethylene production and phenolic secretion. In Vitro Cell. Dev. Biol. 2016, 52, 408–418. [Google Scholar] [CrossRef]
- Arigita, L.; González, A.; Tamés, R.S. Influence of CO2 and sucrose on photosynthesis and transpiration of Actinidia deliciosa explants cultured in vitro. Physiol. Plant. 2002, 115, 166–173. [Google Scholar] [CrossRef]
- Arigita, L.; Sánchez Tamés, R.; González, A. 1-Methylcyclopropene and ethylene as regulators of in vitro organogenesis in kiwi explants. Plant Growth Regul. 2003, 40, 59–64. [Google Scholar] [CrossRef]
- Gong, Y.; Gao, F.; Tang, K.; Debergh, P. In vitro high frequency direct root and shoot regeneration in sweet potato using the ethylene inhibitor silver nitrate. S. Afr. J. Bot. 2005, 71, 110–113. [Google Scholar] [CrossRef]
- Venkatachalam, P.; Malar, S.; Thiyagarajan, M.; Indiraarulselvi, P.; Geetha, N. Effect of phycochemical coated silver nanocomplexes as novel growth-stimulating compounds for plant regeneration of Alternanthera sessilis L. J. Appl. Phycol. 2017, 29, 1095–1106. [Google Scholar] [CrossRef]
- Kumar, M.; Muthusamy, A.; Kumar, V.; Bhalla-Sarin, N. In Vitro Plant Breeding towards Novel Agronomic Traits: Biotic and Abiotic Stress Tolerance; Springer: Berlin/Heidelberg, Germany, 2019. [Google Scholar]
- Bergeson, L.L.; Cole, M.F. Regulatory implications of nanotechnology. Biol. Interact. 2014, 315, 1–12. [Google Scholar] [CrossRef]
- Ghormade, V.; Deshpande, M.V.; Paknikar, K.M. Perspectives for nano-biotechnology enabled protection and nutrition of plants. Biotechnol. Adv. 2011, 29, 792–803. [Google Scholar] [CrossRef]
- Duhan, J.S.; Kumar, R.; Kumar, N.; Kaur, P.; Nehra, K.; Duhan, S. Nanotechnology: The new perspective in precision agriculture. Biotechnol. Rep. 2017, 15, 11–23. [Google Scholar] [CrossRef] [PubMed]
- Raliya, R.; Saharan, V.; Dimkpa, C.; Biswas, P. Nanofertilizer for precision and sustainable agriculture: Current state and future perspectives. J. Agric. Food Chem. 2017, 66, 6487–6503. [Google Scholar] [CrossRef] [PubMed]
- Prasad, R.; Bhattacharyya, A.; Nguyen, Q.D. Nanotechnology in sustainable agriculture: Recent developments, challenges, and perspectives. Front. Microbio. 2017, 8, 1014. [Google Scholar] [CrossRef] [PubMed]
- Beer, C.; Foldbjerg, R.; Hayashi, Y.; Sutherland, D.S.; Autrup, H. Toxicity of silver nanoparticles—Nanoparticle or silver ion? Toxicol. Lett. 2012, 208, 286–292. [Google Scholar] [CrossRef]
- Kulus, D.; Tymoszuk, A. Gold nanoparticles affect the cryopreservation efficiency of in vitro-derived shoot tips of bleeding heart. Plant Cell Tissue Organ Cult. (PCTOC) 2021, 146, 297–311. [Google Scholar] [CrossRef]
- Zhang, X.-F.; Liu, Z.-G.; Shen, W.; Gurunathan, S. Silver nanoparticles: Synthesis, characterization, properties, applications, and therapeutic approaches. Int. J. Mol. Sci. 2016, 17, 1534. [Google Scholar] [CrossRef]
- Parzymies, M. Nano-silver particles reduce contaminations in tissue culture but decrease regeneration rate and slows down growth and development of Aldrovanda vesiculosa explants. Appl. Sci. 2021, 11, 3653. [Google Scholar] [CrossRef]
- Bello-Bello, J.J.; Spinoso-Castillo, J.L.; Arano-Avalos, S.; Martínez-Estrada, E.; Arellano-García, M.E.; Pestryakov, A.; Toledano-Magaña, Y.; García-Ramos, J.C.; Bogdanchikova, N. Cytotoxic, genotoxic, and polymorphism effects on Vanilla planifolia Jacks ex Andrews after long-term exposure to Argovit® silver nanoparticles. Nanomaterials 2018, 8, 754. [Google Scholar] [CrossRef]
- Pérez-Caselles, C.; Alburquerque, N.; Faize, L.; Bogdanchikova, N.; García-Ramos, J.C.; Rodríguez-Hernández, A.G.; Pestryakov, A.; Burgos, L. How to get more silver? Culture media adjustment targeting surge of silver nanoparticle penetration in apricot tissue during in vitro micropropagation. Horticulturae 2022, 8, 855. [Google Scholar] [CrossRef]
- Rahmawati, M.; Mahfud, C.; Risuleo, G.; Jadid, N. Nanotechnology in plant metabolite improvement and in animal welfare. Appl. Sci. 2022, 12, 838. [Google Scholar] [CrossRef]
- Reynolds, M.P.; Braun, H.-J. Wheat Improvement. In Wheat Improvement Food Security in a Changing Climate; Reynolds, M.P., Braun, H.-J., Eds.; Springer: Cham, Switzerland, 2022; pp. 3–15. ISBN 9783030906733. [Google Scholar]
- Huertas-García, A.B.; Tabbita, F.; Alvarez, J.B.; Sillero, J.C.; Ibba, M.I.; Rakszegi, M.; Guzmán, C. Genetic variability for grain components related to nutritional quality in spelt and common wheat. J. Agric. Food Chem. 2023, 71, 10598–10606. [Google Scholar] [CrossRef] [PubMed]
- Blanco, J.; Lafuente, D.; Gómez, M.; García, T.; Domingo, J.L.; Sánchez, D.J. Polyvinyl pyrrolidone-coated silver nanoparticles in a human lung cancer cell: Time-and dose-dependent influence over p53 and caspase-3 protein expression and epigenetic effects. Arch. Toxicol. 2017, 91, 651–666. [Google Scholar] [CrossRef] [PubMed]
- Brzóska, K.; Grądzka, I.; Kruszewski, M. Silver, gold, and iron oxide nanoparticles alter miRNA expression but do not affect DNA methylation in HepG2 cells. Materials. 2019, 12, 1038. [Google Scholar] [CrossRef] [PubMed]
- Mytych, J.; Zebrowski, J.; Lewinska, A.; Wnuk, M. Prolonged effects of silver nanoparticles on p53/p21 pathway-mediated proliferation, DNA damage response, and methylation parameters in HT22 hippocampal neuronal cells. Mol. Neurobiol. 2017, 54, 1285–1300. [Google Scholar] [CrossRef]
- Smolkova, B.; Miklikova, S.; Kajabova, V.H.; Babelova, A.; El Yamani, N.; Zduriencikova, M.; Fridrichova, I.; Zmetakova, I.; Krivulcik, T.; Kalinkova, L. Global and gene specific DNA methylation in breast cancer cells was not affected during epithelial-to-mesenchymal transition in vitro. Neoplasma 2016, 609. [Google Scholar] [CrossRef]
- Haliloğlu, K.; Türkoğlu, A.; Balpınar, Ö.; Nadaroğlu, H.; Alaylı, A.; Poczai, P. Effects of Zinc, Copper and Iron Oxide Nanoparticles on Induced DNA Methylation, Genomic Instability and LTR Retrotransposon Polymorphism in Wheat (Triticum aestivum L.). Plants 2022, 11, 2193. [Google Scholar] [CrossRef]
- Pejam, F.; Ardebili, Z.O.; Ladan-Moghadam, A.; Danaee, E. Zinc oxide nanoparticles mediated substantial physiological and molecular changes in tomato. PloS ONE 2021, 16, e0248778. [Google Scholar] [CrossRef]
- Rajaee Behbahani, S.; Iranbakhsh, A.; Ebadi, M.; Majd, A.; Ardebili, Z.O. Red elemental selenium nanoparticles mediated substantial variations in growth, tissue differentiation, metabolism, gene transcription, epigenetic cytosine DNA methylation, and callogenesis in bittermelon (Momordica charantia); an in vitro experiment. PloS ONE 2020, 15, e0235556. [Google Scholar] [CrossRef]
- Sotoodehnia-Korani, S.; Iranbakhsh, A.; Ebadi, M.; Majd, A.; Ardebili, Z.O. Selenium nanoparticles induced variations in growth, morphology, anatomy, biochemistry, gene expression, and epigenetic DNA methylation in Capsicum annuum; an in vitro study. Environ. Pollut. 2020, 265, 114727. [Google Scholar] [CrossRef] [PubMed]
- Vafaie Moghadam, A.; Iranbakhsh, A.; Saadatmand, S.; Ebadi, M.; Oraghi Ardebili, Z. New insights into the transcriptional, epigenetic, and physiological responses to zinc oxide nanoparticles in Datura stramonium; potential species for phytoremediation. J. Plant Growth Reg. 2022, 41, 271–281. [Google Scholar] [CrossRef]
- Li, X.; Liu, W.; Sun, L.; Aifantis, K.E.; Yu, B.; Fan, Y.; Feng, Q.; Cui, F.; Watari, F. Effects of physicochemical properties of nanomaterials on their toxicity. J. Biomed. Mater. Res. A 2015, 103, 2499–2507. [Google Scholar] [CrossRef]
- Erturk, F.A.; Aydin, M.; Sigmaz, B.; Taspinar, M.S.; Arslan, E.; Agar, G.; Yagci, S. Effects of As2O3 on DNA methylation, genomic instability, and LTR retrotransposon polymorphism in Zea mays. Environ. Sci. Pollut. Res. 2015, 22, 18601–18606. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Rong, T.; Cao, M. Analysis of DNA methylation in different maize tissues. J. Genet. Genom. 2008, 35, 41–48. [Google Scholar] [CrossRef]
- Hossein Pour, A.; Özkan, G.; Nalci, Ö.; Haliloğlu, K. Estimation of genomic instability and DNA methylation due to aluminum (Al) stress in wheat (Triticum aestivum L.) using iPBS and CRED-iPBS analyses. Turk. J. Bot. 2019, 43, 27–37. [Google Scholar] [CrossRef]
- Orłowska, R.; Pachota, K.A.; Androsiuk, P.; Bednarek, P.T. Triticale green plant regeneration is due to DNA methylation and sequence changes affecting distinct sequence contexts in the presence of copper ions in induction medium. Cells 2021, 11, 84. [Google Scholar] [CrossRef]
- Pereira, W.J.; Pappas, M.d.C.R.; Grattapaglia, D.; Pappas, G.J., Jr. A cost-effective approach to DNA methylation detection by Methyl Sensitive DArT sequencing. PloS ONE 2020, 15, e0233800. [Google Scholar] [CrossRef]
- Wang, S.; Lv, J.; Zhang, L.; Dou, J.; Sun, Y.; Li, X.; Fu, X.; Dou, H.; Mao, J.; Hu, X. MethylRAD: A simple and scalable method for genome-wide DNA methylation profiling using methylation-dependent restriction enzymes. Open Biol. 2015, 5, 150130. [Google Scholar] [CrossRef]
- Brunner, A.L.; Johnson, D.S.; Kim, S.W.; Valouev, A.; Reddy, T.E.; Neff, N.F.; Anton, E.; Medina, C.; Nguyen, L.; Chiao, E. Distinct DNA methylation patterns characterize differentiated human embryonic stem cells and developing human fetal liver. Genome Res. 2009, 19, 1044–1056. [Google Scholar] [CrossRef]
- Bednarek, P.T.; Orłowska, R.; Niedziela, A. A relative quantitative methylation-sensitive amplified polymorphism (MSAP) method for the analysis of abiotic stress. BMC Plant Biol. 2017, 17, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Bulunuz Palaz, E.; Ugur, R.; Yaman, M. Micropropagation Protocols of New Prunus Hybrids with Significant Rootstock Potential in Fruit Breeding and Cultivation. Erwerbs-Obstbau 2023, 65, 1359–1364. [Google Scholar] [CrossRef]
- Türkoğlu, A.; Haliloğlu, K.; Tosun, M.; Szulc, P.; Demirel, F.; Eren, B.; Bujak, H.; Karagöz, H.; Selwet, M.; Özkan, G. Sodium Azide as a Chemical Mutagen in Wheat (Triticum aestivum L.): Patterns of the Genetic and Epigenetic Effects with iPBS and CRED-iPBS Techniques. Agriculture 2023, 13, 1242. [Google Scholar] [CrossRef]
- Türkoğlu, A.; Tosun, M.; Haliloğlu, K. Evaluation of ethyl methanesulfonate-induced in vitro mutagenesis, polymorphism, and genomic instability in wheat (Triticum aestivum L.). J. Crop Sci.Biotechnol. 2023, 26, 199–213. [Google Scholar] [CrossRef]
- Aasim, M.; Akin, F.; Ali, S.A.; Taskin, M.B.; Colak, M.S.; Khawar, K.M. Artificial neural network modeling for deciphering the in vitro induced salt stress tolerance in chickpea (Cicer arietinum L). Physiol. Mol. Biol. Plants 2023, 29, 289–304. [Google Scholar] [CrossRef] [PubMed]
- Aasim, M.; Ali, S.A.; Altaf, M.T.; Ali, A.; Nadeem, M.A.; Baloch, F.S. Artificial neural network and decision tree facilitated prediction and validation of cytokinin-auxin induced in vitro organogenesis of sorghum (Sorghum bicolor L.). Plant Cell Tissue Organ Cult. (PCTOC) 2023, 153, 611–624. [Google Scholar] [CrossRef]
- Aasim, M.; Ali, S.A.; Bekiş, P.; Nadeem, M.A. Light-emitting diodes induced in vitro regeneration of Alternanthera reineckii mini and validation via machine learning algorithms. In Vitro Cell. Dev. Biol. 2022, 58, 816–825. [Google Scholar] [CrossRef]
- Aasim, M.; Katırcı, R.; Akgur, O.; Yildirim, B.; Mustafa, Z.; Nadeem, M.A.; Baloch, F.S.; Karakoy, T.; Yılmaz, G. Machine learning (ML) algorithms and artificial neural network for optimizing in vitro germination and growth indices of industrial hemp (Cannabis sativa L.). Ind. Crops Prod. 2022, 181, 114801. [Google Scholar] [CrossRef]
- Hesami, M.; Jones, A.M.P. Application of artificial intelligence models and optimization algorithms in plant cell and tissue culture. Appl. Microbiol. Biotechnol. 2020, 104, 9449–9485. [Google Scholar] [CrossRef]
- Niazian, M.; Niedbała, G. Machine learning for plant breeding and biotechnology. Agriculture 2020, 10, 436. [Google Scholar] [CrossRef]
- Gökmen, F.; Uygur, V.; Sukuşu, E. Extreme Gradient Boosting Regression Model for Soil Available Boron. Eurasian Soil Sci. 2023, 56, 738–746. [Google Scholar] [CrossRef]
- Iqbal, F.; Raziq, A.; Tirink, C.; Fatih, A.; Yaqoob, M. Using the artificial bee colony technique to optimize machine learning algorithms in estimating the mature weight of camels. Trop. Anim. Health Prod. 2023, 55, 86. [Google Scholar] [CrossRef] [PubMed]
- Kirtis, A.; Aasim, M.; Katırcı, R. Application of artificial neural network and machine learning algorithms for modeling the in vitro regeneration of chickpea (Cicer arietinum L.). Plant Cell Tissue Organ Cult. (PCTOC) 2022, 150, 141–152. [Google Scholar] [CrossRef]
- Mirza, K.; Aasim, M.; Katırcı, R.; Karataş, M.; Ali, S.A. Machine learning and artificial neural networks-based approach to model and optimize ethyl methanesulfonate and sodium azide induced in vitro regeneration and morphogenic traits of water hyssops (Bacopa monnieri L.). J. Plant Growth Regul. 2023, 42, 3471–3485. [Google Scholar] [CrossRef]
- Khodakovskaya, M.V.; De Silva, K.; Biris, A.S.; Dervishi, E.; Villagarcia, H. Carbon nanotubes induce growth enhancement of tobacco cells. ACS Nano 2012, 6, 2128–2135. [Google Scholar] [CrossRef]
- Malik, W.A.; Mahmood, I.; Razzaq, A.; Afzal, M.; Shah, G.A.; Iqbal, A.; Zain, M.; Ditta, A.; Asad, S.A.; Ahmad, I.; et al. Exploring potential of copper and silver nano particles to establish efficient callogenesis and regeneration system for wheat (Triticum aestivum L.). GM Crops Food 2021, 12, 564–585. [Google Scholar] [CrossRef]
- Salama, H.M. Effects of silver nanoparticles in some crop plants, common bean (Phaseolus vulgaris L.) and corn (Zea mays L.). Int. Res. J. Biotechnol. 2012, 3, 190–197. [Google Scholar]
- Selivanov, N.Y.; Selivanova, O.; Sokolov, O.; Sokolova, M.; Sokolov, A.; Bogatyrev, V.; Dykman, L. Effect of gold and silver nanoparticles on the growth of the Arabidopsis thaliana cell suspension culture. Nanotechnol. Russ. 2017, 12, 116–124. [Google Scholar] [CrossRef]
- Zhang, B.; Zheng, L.P.; Yi Li, W.; Wen Wang, J. Stimulation of artemisinin production in Artemisia annua hairy roots by Ag-SiO2 core-shell nanoparticles. Curr. Nanosci. 2013, 9, 363–370. [Google Scholar] [CrossRef]
- Wu, J.; Song, K.H.; Litzinger, T.; Lee, S.Y.; Santoro, R.; Linevsky, M.; Colket, M.; Liscinsky, D. Reduction of PAH and soot in premixed ethylene–air flames by addition of ethanol. Combust. Flame 2006, 144, 675–687. [Google Scholar] [CrossRef]
- Alva Ticona, S.; Oropeza, M. Effect of culture medium consistence and silver nitrate on micropropagation of two potato (Solanum tuberosum) cultivars. Rev. Colomb. Biotecnol. 2013, 15, 55–62. [Google Scholar] [CrossRef]
- Sharma, A.; Kumar, V.; Giridhar, P.; Ravishankar, G.A. Induction of in vitro flowering in Capsicum frutescens under the influence of silver nitrate and cobalt chloride and pollen transformation. Electron. J. Biotechnol. 2008, 11, 84–89. [Google Scholar] [CrossRef]
- Fei, S.; Read, P.E.; Riordan, T.P. Improvement of embryogenic callus induction and shoot regeneration of buffalograss by silver nitrate. Plant Cell Tissue Organ Cult. (PCTOC) 2000, 60, 197–203. [Google Scholar] [CrossRef]
- Kothari-Chajer, A.; Sharma, M.; Kachhwaha, S.; Kothari, S. Micronutrient optimization results into highly improved in vitro plant regeneration in kodo (Paspalum scrobiculatum L.) and finger (Eleusine coracana (L.) Gaertn.) millets. Plant Cell Tissue Organ Cult. (PCTOC) 2008, 94, 105–112. [Google Scholar] [CrossRef]
- Najafi, S.; Heidari, R.; Jamei, R. Photosynthetic characteristics, membrane lipid levels and protein content in the Phaseolus vulgaris L. (cv. Sadri) exposed to magnetic field and silver nanoparticles. Bull. Environ. Pharmacol. Life Sci. 2014, 3, 72–76. [Google Scholar]
- Vannini, C.; Domingo, G.; Onelli, E.; Prinsi, B.; Marsoni, M.; Espen, L.; Bracale, M. Morphological and proteomic responses of Eruca sativa exposed to silver nanoparticles or silver nitrate. PLoS ONE 2013, 8, e68752. [Google Scholar] [CrossRef] [PubMed]
- Yin, L.; Cheng, Y.; Espinasse, B.; Colman, B.P.; Auffan, M.; Wiesner, M.; Rose, J.; Liu, J.; Bernhardt, E.S. More than the ions: The effects of silver nanoparticles on Lolium multiflorum. Environ. Sci. Technol. 2011, 45, 2360–2367. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.M.; Kwak, J.I.; An, Y.J. Effect of silver nanoparticles in crop plants Phaseolus radiatus and Sorghum bicolor: Media effect on phytotoxicity. Chemosphere 2012, 86, 491–499. [Google Scholar] [CrossRef]
- Kaveh, R.; Li, Y.S.; Ranjbar, S.; Tehrani, R.; Brueck, C.L.; Van Aken, B. Changes in Arabidopsis thaliana gene expression in response to silver nanoparticles and silver ions. Environ. Sci. Technol. 2013, 47, 10637–10644. [Google Scholar] [CrossRef]
- Mazumdar, H.; Ahmed, G. Phytotoxicity effect of silver nanoparticles on Oryza sativa. Int. J. Chem. Technol. Res. 2011, 3, 1494–1500. [Google Scholar]
- Aghdaei, M.; Sarmast, M.; Salehi, H. Effects of silver nanoparticles on Tecomella undulata (Roxb.) Seem. micropropagation. Adv. Hort. Sci. 2012, 26, 21–24. [Google Scholar]
- Gruyer, N.; Dorais, M.; Bastien, C.; Dassylva, N.; Triffault-Bouchet, G. Interaction between silver nanoparticles and plant growth. ISHS 2013, 1037, 795–800. [Google Scholar] [CrossRef]
- Sarmast, M.; Niazi, A.; Salehi, H.; Abolimoghadam, A. Silver nanoparticles affect ACS expression in Tecomella undulata in vitro culture. Plant Cell Tissue Organ Cult. (PCTOC) 2015, 121, 227–236. [Google Scholar] [CrossRef]
- Karimi, J.; Mohsenzadeh, S. Physiological effects of silver nanoparticles and silver nitrate toxicity in Triticum aestivum. Iran. J. Sci. Technol. Trans. A Sci. 2017, 41, 111–120. [Google Scholar] [CrossRef]
- Bello-Bello, J.J.; Chavez-Santoscoy, R.A.; Lecona-Guzman, C.A.; Bogdanchikova, N.; Salinas-Ruíz, J.; Gomez-Merino, F.C.; Pestryakov, A. Hormetic response by silver nanoparticles on in vitro multiplication of sugarcane (Saccharum spp. Cv. Mex 69–290) using a temporary immersion system. Dose-Response 2017, 15, 1559325817744945. [Google Scholar] [CrossRef]
- Spinoso-Castillo, J.; Chavez-Santoscoy, R.; Bogdanchikova, N.; Pérez-Sato, J.; Morales-Ramos, V.; Bello-Bello, J. Antimicrobial and hormetic effects of silver nanoparticles on in vitro regeneration of vanilla (Vanilla planifolia Jacks. ex-Andrews) using a temporary immersion system. Plant Cell Tissue Organ Cult. (PCTOC) 2017, 129, 195–207. [Google Scholar] [CrossRef]
- Castro-González, C.G.; Sánchez-Segura, L.; Gómez-Merino, F.C.; Bello-Bello, J.J. Exposure of stevia (Stevia rebaudiana B.) to silver nanoparticles in vitro: Transport and accumulation. Sci. Rep. 2019, 9, 10372. [Google Scholar] [CrossRef]
- Timoteo, C.d.O.; Paiva, R.; Dos Reis, M.V.; Claro, P.I.C.; Ferraz, L.M.; Marconcini, J.M.; de Oliveira, J.E. In vitro growth of Physalis peruviana L. affected by silver nanoparticles. 3 Biotech 2019, 9, 145. [Google Scholar] [CrossRef]
- Jiang, H.S.; Li, M.; Chang, F.Y.; Li, W.; Yin, L.Y. Physiological analysis of silver nanoparticles and AgNO3 toxicity to Spirodela polyrhiza. Environ. Toxicol. Chem. 2012, 31, 1880–1886. [Google Scholar] [CrossRef]
- Shang, H.; Guo, H.; Ma, C.; Li, C.; Chefetz, B.; Polubesova, T.; Xing, B. Maize (Zea mays L.) root exudates modify the surface chemistry of CuO nanoparticles: Altered aggregation, dissolution and toxicity. Sci. Total Environ. 2019, 690, 502–510. [Google Scholar] [CrossRef]
- Guo, X.; Ni, J.; Xue, J.; Wang, X. Extract of bulbus Fritillaria cirrhosa perturbs spindle assembly checkpoint, induces mitotic aberrations and genomic instability in human colon epithelial cell line. Exp. Toxicol. Pathol. 2017, 69, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Grafi, G.; Ben-Meir, H.; Avivi, Y.; Moshe, M.; Dahan, Y.; Zemach, A. Histone methylation controls telomerase-independent telomere lengthening in cells undergoing dedifferentiation. Dev. Biol. 2007, 306, 838–846. [Google Scholar] [CrossRef]
- Han, Z.; Crisp, P.A.; Stelpflug, S.; Kaeppler, S.M.; Li, Q.; Springer, N.M. Heritable epigenomic changes to the maize methylome resulting from tissue culture. Genetics 2018, 209, 983–995. [Google Scholar] [CrossRef] [PubMed]
- Kabita, K.C.; Sharma, S.K.; Sanatombi, K. Analysis of capsaicinoid biosynthesis pathway genes expression in callus cultures of Capsicum chinense Jacq. cv. ‘Umorok’. Plant Cell Tissue Organ Cult. (PCTOC) 2019, 137, 565–573. [Google Scholar] [CrossRef]
- Li, H.; Zhao, X.; Dai, H.; Wu, W.; Mao, W.; Zhang, Z. Tissue culture responsive microRNAs in strawberry. Plant Mol. Biol. Rep. 2012, 30, 1047–1054. [Google Scholar] [CrossRef]
- Abdel-Azeem, E.A.; Elsayed, B.A. Phytotoxicity of silver nanoparticles on Vicia faba seedlings. NY Sci. J. 2013, 6, 148–155. [Google Scholar]
- Ewais, E.A.; Desouky, S.A.; Elshazly, E.H. Evaluation of callus responses of Solanum nigrum L. exposed to biologically synthesized silver nanoparticles. Nanosci. Nanotechnol. Lett. 2015, 5, 45–56. [Google Scholar]
- de la Puente, R.; González, A.I.; Ruiz, M.L.; Polanco, C. Somaclonal variation in rye (Secale cereale L.) analyzed using polymorphic and sequenced AFLP markers. In Vitro Cell. Dev. Biol. 2008, 44, 419–426. [Google Scholar] [CrossRef]
- Bednarek, P.T.; Orłowska, R. Plant tissue culture environment as a switch-key of (epi) genetic changes. Plant Cell Tissue Organ Cult. (PCTOC) 2020, 140, 245–257. [Google Scholar] [CrossRef]
- Bednarek, P.T.; Orłowska, R.; Mańkowski, D.R.; Zimny, J.; Kowalczyk, K.; Nowak, M.; Zebrowski, J. Glutathione and copper ions as critical factors of green plant regeneration efficiency of triticale in vitro anther culture. Front. Plant Sci. 2022, 13, 926305. [Google Scholar] [CrossRef]
- Yang, X.; Wang, L.; Yuan, D.; Lindsey, K.; Zhang, X. Small RNA and degradome sequencing reveal complex miRNA regulation during cotton somatic embryogenesis. J. Exp. Bot. 2013, 64, 1521–1536. [Google Scholar] [CrossRef] [PubMed]
- Becker, C.; Weigel, D. Epigenetic variation: Origin and transgenerational inheritance. Curr. Opin. Plant 2012, 15, 562–567. [Google Scholar] [CrossRef] [PubMed]
- Karaca, M.; Aydin, A.; Ince, A.G. Cytosine methylation polymorphisms in cotton using TD-MS-RAPD-PCR. Mod. Phytomorphol. 2019, 13, 13–19. [Google Scholar]
- Demirel, F.; Eren, B.; Yilmaz, A.; Türkoğlu, A.; Haliloğlu, K.; Niedbała, G.; Bujak, H.; Jamshidi, B.; Pour-Aboughadareh, A.; Bocianowski, J. Prediction of Grain Yield in Wheat by CHAID and MARS Algorithms Analyses. Agronomy 2023, 13, 1438. [Google Scholar] [CrossRef]
- Çiçek, S. Cytotoxicity of silver nanoparticles obtained from Eruca vesicaria on rainbow trout gonad cell line-2 (RTG-2). GUFBD 2022, 12, 1093–1101. [Google Scholar]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio-assays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Turkoglu, A. Effects of mammalian sex hormones on regeneration capacity, retrotransposon polymorphism and genomic instability in wheat (Triticum aestivum L.). Plant Cell Tissue Organ Cult. (PCTOC) 2023, 152, 647–659. [Google Scholar] [CrossRef]
- Hossein Pour, A.; Aydin, M.; Haliloglu, K. Plant regeneration system in recalcitrant rye (Secale cereale L.). Biologia 2020, 75, 1017–1028. [Google Scholar] [CrossRef]
- Nalci, O.B.; Nadaroglu, H.; Pour, A.H.; Gungor, A.A.; Haliloglu, K. Effects of ZnO, CuO and γ-Fe 3 O 4 nanoparticles on mature embryo culture of wheat (Triticum aestivum L.). Plant Cell Tissue Organ Cult. (PCTOC) 2019, 136, 269–277. [Google Scholar] [CrossRef]
- Zeinalzadehtabrizi, H.; Hosseinpour, A.; Aydin, M.; Haliloglu, K. A modified genomic DNA extraction method from leaves of sunflower for PCR based analyzes. J. Biodivers. Environ. Sci. 2015, 7, 222–225. [Google Scholar]
- Hosseinpour, A.; Ilhan, E.; Özkan, G.; Öztürk, H.İ.; Haliloglu, K.; Cinisli, K.T. Plant growth-promoting bacteria (PGPBs) and copper (II) oxide (CuO) nanoparticle ameliorates DNA damage and DNA Methylation in wheat (Triticum aestivum L.) exposed to NaCl stress. J. Plant Biochem. Biotechnol. 2022, 31, 751–764. [Google Scholar] [CrossRef]
- Demirel, F.; Türkoğlu, A.; Haliloğlu, K.; Eren, B.; Özkan, G.; Uysal, P.; Pour-Aboughadareh, A.; Leśniewska-Bocianowska, A.; Jamshidi, B.; Bocianowski, J. Mammalian sex hormones as steroid-structured compounds in wheat seedling: Template of the cytosine methylation alteration and retrotransposon polymorphisms with iPBS and CRED-iBPS techniques. Appl. Sci. 2023, 13, 9538. [Google Scholar] [CrossRef]
- Türkoğlu, A.; Haliloğlu, K.; Tosun, M.; Bujak, H.; Eren, B.; Demirel, F.; Szulc, P.; Karagöz, H.; Selwet, M.; Özkan, G. Ethyl methanesulfonate (EMS) mutagen toxicity-induced DNA damage, cytosine methylation alteration, and iPBS-retrotransposon polymorphisms in wheat (Triticum aestivum L.). Agronomy 2023, 13, 1767. [Google Scholar] [CrossRef]
- Noble, W.S. What is a support vector machine? Nat. Biotechnol. 2006, 24, 1565–1567. [Google Scholar] [CrossRef]
- Breiman, L. Random forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Chen, T.; Guestrin, C. Xgboost: A scalable tree boosting system. In Proceedings of the 22nd ACM Sigkdd International Conference on Knowledge Discovery and Data Mining, San Francisco, CA, USA, 13–17 August 2016; pp. 785–794. [Google Scholar]
- Appelhans, T.; Mwangomo, E.; Hardy, D.R.; Hemp, A.; Nauss, T. Evaluating machine learning approaches for the interpolation of monthly air temperature at Mt. Kilimanjaro, Tanzania. Spat. Stat. 2015, 14, 91–113. [Google Scholar] [CrossRef]
- Rasmussen, C.E. Gaussian processes in machine learning. In Summer School on Machine Learning; Springer: Berlin/Heidelberg, Germany, 2003; pp. 63–71. [Google Scholar]
- Sammut, C.; Webb, G.I. Leave-one-out cross-validation. In Encyclopedia of Machine Learning; Sammut, C., Webb, G.I., Eds.; Springer: Boston, MA, USA, 2010; pp. 600–601. [Google Scholar]
- Grzesiak, W.; Zaborski, D. Examples of the use of data mining methods in animal breeding. Data Min. Appl. Eng. Med. 2012, 13, 303–324. [Google Scholar]
- Kuhn, M.; Wing, J.; Weston, S.; Williams, A.; Keefer, C.; Engelhardt, A.; Cooper, T.; Mayer, Z.; Kenkel, B.; Benesty, M. Caret: Classification and Regression Training: R Package; R Core Team: Vienna, Austria, 2019. [Google Scholar]
- Team R Core. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2010. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Concentration (mg L−1) | CI (%) 1 | EC (%) | RE (Number) |
|---|---|---|---|---|
| AgNO3 | 0 | 80.00 abc2 | 65.00 bc | 0.43 bcd |
| AgNO3 | 2 | 82.50 abc | 70.00 abc | 0.20 d |
| AgNO3 | 4 | 60.00 d | 37.50 d | 0.13 d |
| AgNO3 | 6 | 90.00 ab | 80.00 ab | 0.73 ab |
| AgNO3 | 8 | 90.00 ab | 80.00 ab | 0.83 a |
| Means | 80.50 | 66.50 | 0.46 | |
| Ag-NPs | 0 | 80.00 abc | 65.00 bc | 0.43 bcd |
| Ag-NPs | 2 | 95.00 a | 85.00 a | 0.68 ab |
| Ag-NPs | 4 | 87.50 ab | 67.50 abc | 0.55 abc |
| Ag-NPs | 6 | 75.00 bcd | 62.50 bc | 0.65 ab |
| Ag-NPs | 8 | 67.50 cd | 55.00 c | 0.33 cd |
| Means | 81.00 | 67.00 | 0.53 | |
| Mean concentration | 0 | 80.00 | 65.00 a | 0.43 bc |
| 2 | 88.75 | 77.50 a | 0.44 bc | |
| 4 | 73.75 | 52.50 b | 0.34 c | |
| 6 | 82.50 | 71.25 a | 0.69 a | |
| 8 | 78.75 | 67.50 a | 0.58 ab | |
| Mean square of treatment (T) | 2.50 ns | 2.50 ns | 0.04 ns | |
| Mean square of concentration (C) | 241.25 ns | 685.00 ** | 0.15 * | |
| Mean square of T × C | 821.25 *** | 1027.50 *** | 0.32 *** |
| Primers | ± 1 | Control 2 | Experimental Groups | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| AgNO3 | Ag-NPs | |||||||||
| 2 mg L−1 | 4 mg L−1 | 6 mg L−1 | 8 mg L−1 | 2 mg L−1 | 4 mg L−1 | 6 mg L−1 | 8 mg L−1 | |||
| OPA 4 | + | 4 | - | - | - | - | - | - | - | 679; 560; 542; 368 |
| - | 458 | - | - | - | 458 | - | - | - | ||
| OPH 17 | + | 1 | - | - | 375; 216 | 628; 466; 207 | - | - | 588 | 418 |
| - | - | - | 295 | 295 | - | - | 295 | 295 | ||
| OPH 18 | + | 2 | 574 | 267 | 634 | 567 | 629; 226 | - | 604; 571; 285 | 365 |
| - | 467 | 500 | - | 467 | - | - | - | - | ||
| OPW 4 | + | 6 | 549; 412 | 481; 457 | 436 | 481 | 478; 457 | - | 471 | 488 |
| - | - | - | - | - | 297 | 812; 297 | 812 | - | ||
| OPW 6 | + | 7 | 706 | - | 618 | 531 | 502; 367 | 615; 502 | 500 | 413; 331 |
| - | - | - | - | - | - | - | - | - | ||
| OPW 11 | + | 3 | - | - | 511; 469; 400 | - | - | - | 478 | - |
| - | - | - | - | - | - | 418 | - | - | ||
| OPW 17 | + | 2 | - | 629; 312; 208; 156 | - | - | - | - | 613 | 788 |
| - | - | - | 400 | 400 | 400 | 400 | 400 | - | ||
| OPW 20 | + | 4 | - | 500 | 162 | 408 | 412; 236 | 352 | 329 | 527; 111 |
| - | - | - | - | - | - | - | - | |||
| Primers | M*/H* 1 | ± 2 | Control 3 | Experimental Groups | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AgNO3 | Ag-NPs | |||||||||||
| 2 mg L−1 | 4 mg L−1 | 6 mg L−1 | 8 mg L−1 | 2 mg L−1 | 4 mg L−1 | 6 mgL−1 | 8 mg L−1 | |||||
| OPA 4 | M | + | 9 | - | 647; 585 | 576; 515; 402; 274 | 616; 564; 526 | 668; 600; 547 | 708; 622; 544; 508 | 654; 576; 505 | 691; 641; 564; 528; 512 | |
| - | - | 827 | - | 871; 827 | - | - | - | 827 | ||||
| H | + | 7 | 951; 330 | 907; 751; 600; 498; 433; 406; 289; 256; 227 | 1000; 916; 792; 550; 489; 433; 317; 294; 242 | 990; 541; 498; 472; 323; 160 | 1018; 907; 768; 641; 560; 489; 412; 298 | 907; 852; 792; 708; 553; 538; 519; 474; 416; 360; 330; 307 | - | 900; 871; 725; 491; 472; 319 | ||
| - | - | - | - | - | - | - | - | - | ||||
| OPH 18 | M | + | 15 | - | - | - | - | 1427; 800; 514 | 1427 | - | 1472 | |
| - | 1309; 337; 286 | 1145; 708; 582; 337; 286 | 1145; 1063; 882; 708; 666; 286 | 708; 337; 286 | 286 | 1145; 666; 286 | 708; 337; 286 | 420; 286 | ||||
| H | + | 11 | - | - | 471 | 1018; 832; 492; 359 | 467 | 380; 328 | 1145; 1000 | - | ||
| - | 1336; 1109; 775; 683; 627 | 1336; 891; 775; 683 | 1336; 1181; 891 | 1181 | 775 | 1109 | - | 627 | ||||
| OPW 4 | M | + | 11 | - | 731; 549 | - | - | 600; 434; 306 | 445 | 543; 310 | 800; 434 | |
| - | 362; 259; 205 | 362; 205 | 362; 205 | 205 | 305 | - | 841 | - | ||||
| H | + | 13 | 439 | - | - | 777; 434 | 296 | - | - | - | ||
| - | 900; 600; 327; 149 | 900; 629; 600; 516; 362; 149 | 900; 600 | 900; 629; 600; | 900; 629; 600; | 831; 600; 516 | 900; 600; 516 | 900; 831; 600; 570; 391 | ||||
| OPW 5 | M | + | 6 | 491 | 813;497; 338 | 1318; 864; 654 502; 338; 323 | 826; 494; 354;331; 305 | 852; 747; 578; 494; 406; 313 | 888; 760; 578; 482; 360; 338 | 864; 711; 639;570; 488; 403; 333; 297; 206 | 930; 839; 722; 632; 488 450; 346;320; 292 | |
| - | - | - | - | - | - | - | - | - | ||||
| H | + | 12 | 826 | 900; 760; 679; 600; 524; 510; 264 | 983; 930; 760; 604 | 826; 502 | 800; 711 | 921; 734; 600; 294 | 864 | 888; 773; 513; 292 | ||
| - | 639; 482; 394; 373; 352; 333 | - | 415; 394; 333; 252; 200 | 373 | 394; 373 | 394 | 532; 415; 333; 200 | 415; 394 | ||||
| OPW 6 | M | + | 11 | 829; 296 | 854; 749; 466; 145 | 800; 154 | 966; 866; 715 | 955; 866; 766 | 955; 866; 700; 641; 449; 352; 228;179 | 900; 800; 700; 312 | 900; 800; 715; 191 | |
| - | - | - | - | 485; 429 | - | - | 485 | - | ||||
| H | + | 12 | 605 | 922; 672 252; 150 | 955; 732 | 955; 749; 600; 472; 318; 296; 166 | 456; 432; 324; 179 | 866; 216 | 933; 863 | 429; 220; 154; 100 | ||
| - | 515 | 515 | 515 | 515 | 584; 539; 515 | - | - | 641; 539 | ||||
| OPW 11 | M | + | 9 | - | 646 | 670; 514 | - | - | - | 800; 145 | - | |
| - | - | 238 | 367; 293 | 722; 530; 238 | 722; 400; 293 | 428; 293 | - | - | ||||
| H | + | 6 | 300; 210 | 293 | - | 813; 575; 450; 378 | 455; 312; 232 | 504; 331; 268 | 437; 400 | 679; 495; 437 | ||
| - | - | - | 722; 419 | - | - | - | - | - | ||||
| OPW 17 | M | + | 5 | 788 | 324 | 547 | 180 | 860; 644; 563; 321 | 309 | 900; 810; 419 | 983; 886; 846 | |
| - | 488; 449 | 488 | 362 | - | 221 | 488; 449; 221 | - | 488; 268; 221 | ||||
| H | + | 3 | 860; 737; 609; 514; 465; 435; 375 | - | 724; 582 | 800; 502 | 913; 810; 700; 574; 547; 509 | 567; 524 | 880; 800; 631 | 838; 604; 377 | ||
| - | - | - | 321 | 265 | 321; 265 | 321; 265 | 321; 265 | 265 | ||||
| OPW 20 | M | + | 11 | - | 800; 713 | 710; 655; 512 | 900; 362 | 775; 524; 502; 384 | 818; 652; 561; 507; 306 | 655; 512; 287 | 818; 761; 649; 368; 324 | |
| - | 485; 469; 203 | 203 | 203 | 677; 203 | 203 | - | - | 540; 485 | ||||
| H | + | 9 | 717; 622 | 818; 619; 600 | 849; 734; 710; 605; 509 | 860; 717; 600; 514; 384 | 800; 734; 448; 386; 306; 219 | 818; 258 | 917; 791; 625;600; 386 | 880; 749; 710; 661; 512; 431; 362 | ||
| - | 258 | 258 | 258 | 258 | 634 | 258 | 258 | 258 | ||||
| Traits | ML Criteria | SVM | RF | XGBoost | KNN | GP | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Train | Test | Train | Test | Train | Test | Train | Test | Train | Test | ||
| CI 1 | R2 | 0.281 | 0.098 | 0.402 | 0.244 | 0.551 | 0.515 | 0.076 | 0.068 | 0.539 | 0.443 |
| MSE | 10.462 | 16.620 | 9.545 | 15.217 | 8.273 | 12.190 | 11.859 | 16.890 | 8.379 | 13.055 | |
| MAPE | 10.438 | 21.635 | 10.267 | 19.753 | 8.607 | 16.248 | 12.025 | 21.691 | 9.028 | 17.425 | |
| MAD | 7.761 | 12.118 | 7.876 | 11.287 | 6.677 | 9.523 | 9.298 | 12.172 | 7.006 | 10.332 | |
| EC | R2 | 0.383 | 0.574 | 0.436 | 0.719 | 0.648 | 0.393 | 0.144 | 0.432 | 0.595 | 0.706 |
| MSE | 13.324 | 9.326 | 12.739 | 7.577 | 10.069 | 11.130 | 15.694 | 10.768 | 10.798 | 7.743 | |
| MAPE | 15.835 | 9.899 | 20.133 | 9.329 | 14.870 | 16.650 | 24.518 | 15.233 | 15.825 | 10.389 | |
| MAD | 8.665 | 6.923 | 10.472 | 6.165 | 8.334 | 9.983 | 12.547 | 9.169 | 8.522 | 6.916 | |
| RE | R2 | 0.526 | 0.505 | 0.502 | 0.422 | 0.671 | 0.461 | 0.145 | 0.236 | 0.659 | 0.525 |
| MSE | 0.185 | 0.173 | 0.190 | 0.186 | 0.155 | 0.180 | 0.249 | 0.214 | 0.157 | 0.169 | |
| MAPE | 37.895 | 55.642 | 56.053 | 59.253 | 34.188 | 54.184 | 72.593 | 69.556 | 36.623 | 48.638 | |
| MAD | 0.131 | 0.154 | 0.161 | 0.167 | 0.121 | 0.159 | 0.201 | 0.191 | 0.126 | 0.138 | |
| Primer Name | Sequence (5′–3′) | Annealing Temperature (°C) |
|---|---|---|
| OPA 4 | AATCGGGCTG | 39.50 |
| OPH 17 | CACTCTCCTC | 35.50 |
| OPH 18 | GAATCGGCCA | 37.50 |
| OPW 4 | CAGAAGCGGA | 39.50 |
| OPW 6 | AGGCCCGATG | 38.50 |
| OPW 11 | CTGATGCGTG | 35.50 |
| OPW 17 | GTCCTGGGTT | 36.50 |
| OPW 20 | TGTGGCAGCA | 44.50 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Türkoğlu, A.; Haliloğlu, K.; Demirel, F.; Aydin, M.; Çiçek, S.; Yiğider, E.; Demirel, S.; Piekutowska, M.; Szulc, P.; Niedbała, G. Machine Learning Analysis of the Impact of Silver Nitrate and Silver Nanoparticles on Wheat (Triticum aestivum L.): Callus Induction, Plant Regeneration, and DNA Methylation. Plants 2023, 12, 4151. https://doi.org/10.3390/plants12244151
Türkoğlu A, Haliloğlu K, Demirel F, Aydin M, Çiçek S, Yiğider E, Demirel S, Piekutowska M, Szulc P, Niedbała G. Machine Learning Analysis of the Impact of Silver Nitrate and Silver Nanoparticles on Wheat (Triticum aestivum L.): Callus Induction, Plant Regeneration, and DNA Methylation. Plants. 2023; 12(24):4151. https://doi.org/10.3390/plants12244151
Chicago/Turabian StyleTürkoğlu, Aras, Kamil Haliloğlu, Fatih Demirel, Murat Aydin, Semra Çiçek, Esma Yiğider, Serap Demirel, Magdalena Piekutowska, Piotr Szulc, and Gniewko Niedbała. 2023. "Machine Learning Analysis of the Impact of Silver Nitrate and Silver Nanoparticles on Wheat (Triticum aestivum L.): Callus Induction, Plant Regeneration, and DNA Methylation" Plants 12, no. 24: 4151. https://doi.org/10.3390/plants12244151