
An Insight into the Biology of the Rare and Peculiar Moss Pterygoneurum sibiricum (Pottiaceae): A Conservation Physiology Approach

 , , , , and
, , , , and 
Abstract
:1. Introduction
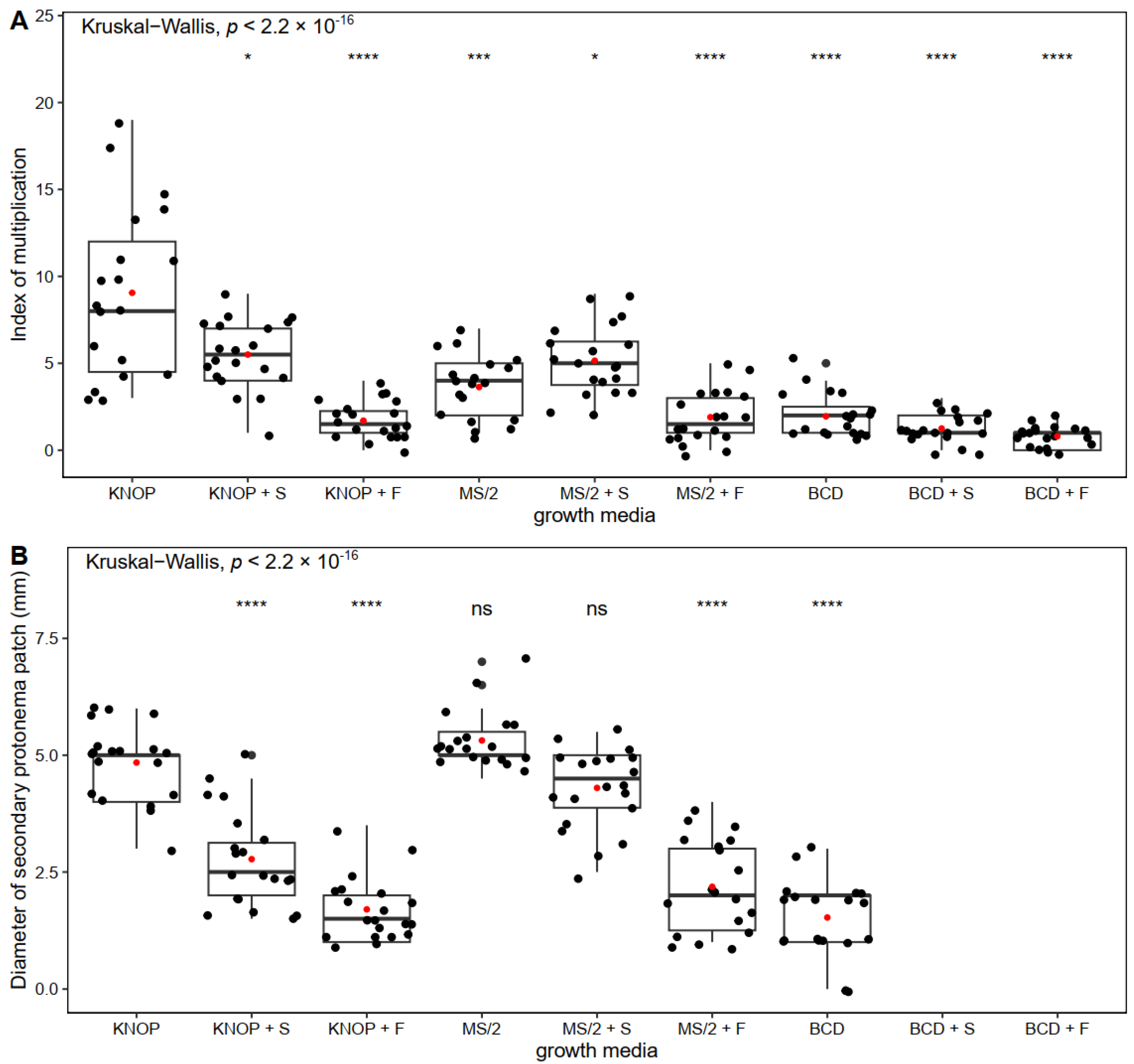
2. Results
2.1. The Influence of Nutrient Medium and Exogenously Added Sugars on Pterygoneurum sibiricum morphogenesis
2.2. The Influence of Growth Regulators on Pterygoneurum sibiricum morphogenesis
2.3. The Influence of NaCl on Pterygoneurum sibiricum morphogenesis
3. Discussion
3.1. The Influence of Nutrient Medium and Exogenously Added Sugars on Pterygoneurum Sibiricum Morphogenesis
3.2. The Influence of Growth Regulators on Pterygoneurum sibiricum morphogenesis
3.3. The Influence of NaCl on Pterygoneurum sibiricum morphogenesis
4. Materials and Methods
4.1. Plant Material
4.2. In Vitro Micropropagation of the Plant Material
4.3. Experimental Design
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Otnyukova, T.N. New cleistocarpous species of the genus Pterygoneurum (Pottiaceae, Bryophyta) from the steppe slopes of Siberia (Russia). Nov. Sist. Nizsh. Rast. 2020, 54, 251–260. [Google Scholar] [CrossRef]
- Pisarenko, O. On the variation and ecology of Pterygoneurum subsessile and P. kozlovii (Pottiaceae). Arctoa 2006, 15, 169–182. [Google Scholar] [CrossRef] [Green Version]
- Kučera, J.; Drevojan, P.; Bradačova, J.; Fialova, L.; Godovičova, L.; Janošik, L.; Kubešova, S.; Manukjanova, A.; Mikulaškova, E.; Skoupa, Z.; et al. Bryophytes recorded during the 2017 spring bryological and lichenological meeting in the Pavlov Hills (Palava). Bryonora 2017, 60, 1–12. [Google Scholar]
- Lazarenko, A.S. Bryological notes. Bot. Zhurn. 1946, 3, 61–63. [Google Scholar]
- Boiko, M.F. Materials for the Red Data Book of Ukraine (Sphagnopsida, Bryopsida). Chorn. Bot. J. 2015, 11, 449–502. [Google Scholar] [CrossRef]
- Šmarda, J. The first supplement to the bryophytes of Slovakia. Act. Mus. Morav. Sci. Biol. 1952, 37, 26–31. [Google Scholar]
- Wallfisch, R. Pterygoneurum kozlovii—New for Romania. Acta Horti Bot. Bucur. 2001, 29, 133–134. [Google Scholar]
- Baisheva, E.; Ignatov, M.; Konstantinova, N.; Maslovsky, O.; Sabovljevic, M.; Ştefănuţ, S. Pterygoneurum kozlovii. IUCN Red List Threat. Species 2019, e.T87562649A87757242. Available online: https://www.iucnredlist.org/species/87562649/87757242 (accessed on 11 January 2023).
- Shafigullina, N.R. New moss records from Tatarstan Republic. 2. In: Sofronova, E.V (ed.). New Bryophyte Records. 1. Arctoa 2012, 21, 275–300. [Google Scholar]
- Doroshina, G.Y. Mosses of the Southern Kalmykia. Nov. Sist. Nizsh. Rast. 2011, 45, 292–300. [Google Scholar]
- Fedosov, V.F.; Ignatova, E.A.; Ignatov, M.S.; Maksimov, A.I. Rare species and preliminary list of mosses of the Anabar Plateau (Subarctic Siberia). Arctoa 2011, 20, 153–174. [Google Scholar] [CrossRef] [Green Version]
- Tsegmed, T. Checklist and distribution of mosses in Mongolia. Arctoa 2001, 10, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Tan, B.C.; Zhao, J.C.; Hu, R.L. An updated checklist of mosses of Xinjiang, China. Arctoa 1995, 4, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Zander, R.H. Pterygoneurum. In Flora of North America, Bryophytes, Mosses, Part 1; Flora of North America Editorial Committee, Ed.; Oxford University Press: New York, NY, USA, 2007; Volume 27, pp. 606–609. ISBN 978-0-19-531-823-4 (v. 27). [Google Scholar]
- Pocs, T. Studies on the cryptogamic vegetation of loess cliffs, I. Orographic desert in the Carpathian Basin. Kitaibelia 1999, 4, 143–156. [Google Scholar]
- Papp, B.; Alegro, A.; Erzberger, P.; Szurdoki, E.; Segota, V.; Sabovljevic, M. Bryophytes of saline areas in the Pannonian region of Serbia and Croatia. Stud. Bot. Hung. 2016, 47, 141–150. [Google Scholar] [CrossRef] [Green Version]
- Ellis, L.T.; Alikhadzhiev, M.K.; Erzhapova, R.S.; Blom, H.H.; Bednarek-Ochyra, H.; Burghardt, M.; Cano, M.J.; Czernyadjeva, I.V.; Kuzmina, E.Y.; Potemkin, A.D.; et al. New national and regional bryophyte records, 64. J. Bryol. 2020, 42, 393–412. [Google Scholar] [CrossRef]
- Vanek, R. Pterygoneurum smardeanum Vanek sp. nov. Preslia 1952, 24, 211–215. [Google Scholar]
- Abramova, A.L.; Blagodatskih, L.S.; Czerpanova, L.A. The genus Pterygoneurum Jur. (Musci) in the URSS. Nov. Sist. Nizsh. Rast. 1973, 10, 305–316. [Google Scholar]
- Boros, Á. Bryogeographie und Bryoflora Ungarns; Akadémiai Kiadó: Budapest, Hungary, 1968; p. 466. [Google Scholar]
- Corley, M.F.V.; Crundwell, A.C.; Düll, R.; Hill, M.O.; Smith, A.J.E. Mosses of Europe and the Azores; an annotated list of species, with synonyms from the recent literature. J. Bryol. 1981, 11, 609–689. [Google Scholar] [CrossRef]
- Novotný, I.; Sutorý, K. Lectotypification of bryophytes described by Jan Šmarda and Rudolf Vanik and held by the herbarium of the Moravian Museum (BRNM). Act. Mus. Morav. Sci. Biol. 2019, 104, 75–79. [Google Scholar]
- Bachurina, H.F. Contribution to the bryophyte flora of the south-western part of Odessa province. Ukr. Bot. J. 1960, 17, 83–88. [Google Scholar]
- Boiko, M.F. Lazarenkia (Pottiaceae, Bryopsida)—A new genus of mosses and Lazarenkia kozlovii—A new combination at species rank. Chorn. Bot. J. 2011, 7, 400–402. [Google Scholar] [CrossRef]
- Hodgetts, N.G.; Lockhart, N. Checklist and country status of European bryophytes—Update 2020. In Irish Wildlife Manuals 123; National Parks and Wildlife Service, Department of Culture, Heritage and the Gaeltacht: Dublin, Ireland, 2020. [Google Scholar]
- Hodgetts, N.; Calix, M.; Englefield, E.; Fettes, N.; Garcia Criado, M.; Patin, L.; Nieto, A.; Bergamini, A.; Bisang, I.; Baisheva, E.; et al. A Miniature World in Decline: European Red List of Mosses, Liverworts and Hornworts; IUCN: Brussels, Belgium, 2019; ISBN 9782831719931/9782831719948. [Google Scholar]
- Ştefănuţ, S.; Goia, I. Checklist and red list of bryophytes of Romania. Nova Hedwig. 2012, 95, 59–104. [Google Scholar] [CrossRef]
- Kubinská, A.; Janovicová, L.; Šoltes, R. Červený zoznam machorastov Slovenska. In Červený Zoznam Rastlin a živočichov Slovenska; Bálaž, D., Marhold, K., Urban, P., Eds.; Ochrana Prirody 20 (Supplement); Centrum Ochrany Prirody a Krajiny: Banska Bystrica, Slovakia, 2001; pp. 31–43. [Google Scholar]
- Misikova, K.; Godovičova, K.; Širka, P.; Šoltes, R. Checklist and red list of mosses (Bryophyta) of Slovakia. Biologia 2020, 75, 21–37. [Google Scholar] [CrossRef]
- Didukh, Y. Red Data Book of Ukraine. Vegetable Kingdom; Globalkonsalting: Kyiv, Ukraine, 2009; 912p. [Google Scholar]
- Lazarenko, A.S. Handbook of Mosses of Ukraine; Naukova Dumka: Kyiv, Ukraine, 1955; 468p. [Google Scholar]
- Shlyakhtin, G.V. Red Data Book of Saratov Province; Izd. Tor.-promyshl. pal. Sar.: Saratov, Russia, 2006; 528p. [Google Scholar]
- Sereda, V.A.; Fedyaeva, V.V. Bryophytes. In Bryophytes in Rad Data Book of Rostov Province Vol. 2; Fedyaeva, V.V., Ed.; Plants and Mushrooms Min. Prirody Rostovskoy Oblasti: Rostov na Donu, Russia, 2014; pp. 322–484. [Google Scholar]
- Cosewic (Committee on the Status of Endangered Wildlife in Canada). Cosewic Assessment and Status Report on the Alkaline Wing-Nerved Moss Pterygoneurum kozlovii in Canada; Committee on the Status of Endangered Wildlife in Canada: Ottawa, ON, Canada, 2004; 20p, Available online: https://www.canada.ca/en/environment-climate-change/services/species-risk-public-registry/cosewic-assessments-status-reports/alkaline-wing-nerved-moss.html (accessed on 11 January 2023).
- Alberta Biodiversity Monitoring Institute. Pterygoneurum Kozlovii. 2020. Available online: https://beta.abmi.ca/biobrowser/species-detail.html?tsn=548039 (accessed on 24 February 2023).
- British Columbia Bryophyte Recovery Team. Recovery Strategy for Alkaline Wing-Nerved Moss (Pterygoneurum kozlovii) in British Columbia; B.C. Ministry of Environment: Victoria, BC, Canada, 2009; 17p. [Google Scholar]
- Rowntree, J.K.; Pressel, S.; Ramsay, M.M.; Sabovljević, A.; Sabovljević, M. In vitro conservation of European bryophytes. In Vitro Cell. Develop. Biol. Plant 2011, 47, 55–64. [Google Scholar] [CrossRef]
- Sabovljević, M.; Vujičić, M.; Pantović, J.; Sabovljevic, A. Bryophyte conservation biology: In vitro approach to the ex situ conservation of bryophytes from Europe. Plant Biosyst. 2014, 148, 857–868. [Google Scholar] [CrossRef]
- Sabovljević, M.S.; Ćosić, M.V.; Jadranin, B.Z.; Pantović, J.P.; Giba, Z.S.; Vujičić, M.M.; Sabovljević, A.D. The conservation physiology of bryophytes. Plants 2022, 11, 1282. [Google Scholar] [CrossRef] [PubMed]
- Basile, D.V. A comparison of macronutrient media used to culture bryophytes. Bryologist 1975, 78, 403–413. [Google Scholar] [CrossRef]
- Sabovljević, M.S.; Nikolić, N.; Vujičić, M.; Sinžar-Sekulić, J.; Pantović, J.; Papp, B.; Sabovljević, A. Ecology, distribution, propagation in vitro, ex situ conservation and native population strenghtening of rare and threatened halophyte moss Entosthodon hungaricus in Serbia. Wulfenia 2018, 25, 117–130. [Google Scholar]
- Sabovljević, M.S.; Segarra-Moragues, J.G.; Puche, F.; Vujičić, M.; Cogoni, A.; Sabovljević, A. Eco-physiological and biotechnological approach to conservation of the world-wide rare and endangered aquatic liverwort Riella helicophylla (Bory et Mont.) Mont. Acta Bot. Croat. 2016, 75, 194–198. [Google Scholar] [CrossRef] [Green Version]
- Sabovljević, A.; Sabovljevic, M.; Grubisic, D.; Konjevic, R. The effect of sugars on development of two moss species (Bryum argenteum and Atrichum undulatum) during in vitro culture (Hedw.) P.Beauv.). Belg. J. Bot. 2005, 138, 79–84. [Google Scholar]
- Sabovljević, A.; Sabovljević, M. Bryophytes, a source of bioactive and new compounds. In Phytopharmacology and Therapeutic Values IV; the Series “Recent Progress in Medicinal Plants”; Govil, J.N., Ed.; Studium Press: Houston, TX, USA, 2008; pp. 9–25. ISBN 1933699124. [Google Scholar]
- Vujičić, M.; Sabovljević, A.; Sabovljević, M. Axenically culturing the bryophytes: Establishment and propagation of the moss Hypnum cupressiforme Hedw. (Bryophyta, Hypnaceae) in in vitro conditions. Bot. Serb. 2011, 35, 71–77. [Google Scholar]
- Ćosić, M.V.; Sabovljević, M.S.; Papp, B.; Giba, Z.S.; Šinžar-Sekulić, J.B.; Sabovljević, A.D.; Vujičić, M.M. Micropropagation of rare bryo-halophyte Hennediella heimii. Bot. Serb. 2022, 46, 187–195. [Google Scholar] [CrossRef]
- Ćosić, M.; Vujičić, M.; Sabovljević, M.; Sabovljević, A. Effects of salt on selected bryophyte species tested under controlled conditions. Bot. Serb. 2020, 44, 27–35. [Google Scholar] [CrossRef]
- Ćosić, M.; Vujičić, M.; Sabovljević, M.; Sabovljević, A. Effects of ABA and NaCl on physiological responses in selected bryophyte species. Botany 2020, 98, 639–650. [Google Scholar] [CrossRef]
- Hassid, W.Z.; Putman, E.W. Transformation of sugars in plants. Annu. Rev. Plant Physiol. 1950, 1, 109–124. [Google Scholar] [CrossRef]
- Lal, M. The culture of bryophytes including apogamy, apospory, parthenogenesis and protoplasts. In The Experimental Biology of Bryophytes; Dyer, A.F., Duckett, J.G., Eds.; Academic Press: London, UK, 1984; pp. 97–115. [Google Scholar]
- Mitra, G.; Allsopp, A. Effects of kinetin, gibberellic acid and certain auxins on the development of shoot buds on the protonema of Pohlia nutans. Nature 1959, 183, 974–975. [Google Scholar] [CrossRef]
- Chopra, R.N.; Kumra, P.K. Protonemal differentiation and bud formation in mosses. In Biology of Bryophytes; Chopra, R.N., Kumra, P.K., Eds.; Wiley Eastern Limited: New Delhi, India, 1988; pp. 40–46. [Google Scholar]
- Gonzalez, M.L.; Mallon, R.; Reinoso, J.; Rodriguez-Oubina, J. In vitro micropropagation and long-ter conservation of the endangered moss Splachnum ampullaceum. Biol. Plant. 2006, 50, 339–345. [Google Scholar] [CrossRef]
- Vujičić, M.; Sabovljević, A.; Šinžar-Sekulić, J.; Skorić, M.; Sabovljević, M. In vitro development of the rare and endangered moss Molendoa hornschuchiana (Hook.) Lindb. ex Limpr. (Pottiaceae, Bryophyta). HortScience 2012, 47, 84–87. [Google Scholar] [CrossRef] [Green Version]
- Sabovljević, M.; Vujičić, M.; Šinžar-Sekulić, J.; Segarra-Moragues, J.G.; Bapp, B.; Skorić, M.; Dragačević, L.; Sabovljević, A. Reviving, in vitro differentiation, development and micropropagation of the rare and endangered moss Bruchia vogesiaca (Bruchiaceae). HortScience 2012, 47, 1347–1350. [Google Scholar] [CrossRef] [Green Version]
- von Schwartzenberg, K. Hormonal regulation of development by auxin and cytokinin in moss. In The Moss Physcomitrella patens; Knight, C., Perroud, P.-F., Cove, D., Eds.; Wiley Online Library: Hoboken, NJ, USA, 2009; Volume 36, pp. 246–281. [Google Scholar] [CrossRef]
- Drábková, L.Z.; Dobrev, P.I.; Motyka, V. Phytohormone profiling across the bryophytes. PLoS ONE 2015, 10, e0125411. [Google Scholar] [CrossRef] [Green Version]
- Szweykowska, A.; Dornowska, E.; Cybulska, A.; Wasiek, G. The cell division response to cytokinins in isolated cell cultures of the protonema of Funaria hygrometrica and its comparison with the bud induction response. Biochem. Physiol. Pflanz. 1971, 162, 514. [Google Scholar] [CrossRef]
- Bijelović, A.; Sabovljević, M.; Grubišić, D.; Konjević, R. Phytohormone influence on the morphogenesis of two mosses (Bryum argenteum Hedw. and Atrichum undulatum (Hedw.) P. Beauv. Isr. J. Plant Sci. 2004, 52, 31–36. [Google Scholar] [CrossRef]
- Ashton, N.W.; Grimsley, N.H.; Cove, D.J. Analysis of gametophyte development in the moss Physcomitrella patens, using auxin and cytokinin resistant mutants. Planta 1979, 144, 427–435. [Google Scholar] [CrossRef]
- Brandes, H.; Kende, H. Studies on cytokinin-controlled bud formation in moss protonemata. Plant Physilogy 1968, 43, 827–837. [Google Scholar] [CrossRef] [Green Version]
- Erichsen, J.; Knoop, B.; Bopp, M. On the action mechanism of cytokinins in mosses: Caulonema specific proteins. Planta 1977, 135, 161–168. [Google Scholar] [CrossRef]
- Bopp, B. Development of the protonema and bud formation in mosses. Bot. J. Linn. Soc. 1963, 58, 305–330. [Google Scholar] [CrossRef]
- Sabovljević, M.; Bijelović, A.; Dragićević, I. Effective and easy way of establishing in vitro culture of mosses, Bryum argenteum Hedw. and Bryum capilare Hedw. (Bryaceae). Arch. Biol. Sci. 2002, 54, 7P–8P. [Google Scholar] [CrossRef]
- Cove, D.J.; Ashton, N.W. The hormonal regulation of gametophytic development in bryophytes. In The Experimental Biology of Bryophytes; Dyer, A.F., Duckett, J.G., Eds.; Academic Press: London, UK, 1984; pp. 177–201. [Google Scholar]
- Johri, B.M.; Mohan Ram, H.Y.; Shah, J.J.; Shah, C.K. Form, Structure, and Function in Plants: Professor, B.M. Johri Commemoration Volume; Sarita Prakashan: Meerut, India, 1975. [Google Scholar]
- Bhatla, S.C.; Kapoor, S.; Khurana, J.P. Involvement of calcium in auxin-induced cell differentiation in the protonema of the wild strain and auxin mutants of the moss Funaria hygrometrica. J. Plant Physiol. 1996, 147, 547–552. [Google Scholar] [CrossRef]
- Sakakibara, K.; Nishiyama, T.; Sumikawa, N.; Kofuji, R.; Murata, T.; Hasebe, M. Involvement of auxin and a homeodomain-leucine zipper I gene in rhizoid development of the moss Physcomitrella patens. Development 2003, 130, 4835–4846. [Google Scholar] [CrossRef] [Green Version]
- Ćosić, M.; Vujičić, M.; Sabovljević, M.; Sabovljević, A. What do we know about salt stress in bryophytes? Plant Biosyst. 2019, 153, 478–489. [Google Scholar] [CrossRef]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flowers, T.J. Improving crop salt tolerance. J. Exp. Bot. 2004, 55, 307–319. [Google Scholar] [CrossRef]
- Bogdanović, M.; Ilić, M.; Živković, S.; Sabovljević, A.; Grubišić, D.; Sabovljević, M. Comparative study on the effects of NaCl on selected moss and fern representatives. Aust. J. Bot. 2011, 59, 734–740. [Google Scholar] [CrossRef]
- Goode, J.A.; Stead, A.D.; Duckett, J.G. Redifferentiation of moss protonemata: An experimental and immunofluorescence study of brood cell formation. Can. J. Bot. 1993, 71, 1510–1519. [Google Scholar] [CrossRef]
- Schnepf, E.; Reinhard, C. Brachycytes in Funaria pro tonemate: Induction by abscisic acid and fine structure. J. Plant Physiol. 1997, 151, 166–175. [Google Scholar] [CrossRef]
- Decker, E.L.; Frank, W.; Sarnighausen, E.; Reski, R. Moss systems biology en route: Phytohormones in Physcomitrella development. Plant Biol. 2006, 8, 397–405. [Google Scholar] [CrossRef] [Green Version]
- Arif, M.A.; Hiss, M.; Tomek, M.; Busch, H.; Meyberg, R.; Tintelnot, S.; Reski, R.; Rensing, S.A.; Frank, W. ABA-induced vegetative diaspore formation in Physcomitrella patens. Front. Plant Sci. 2019, 10, 315. [Google Scholar] [CrossRef] [Green Version]
- Frank, W.; Ratnadewi, D.; Reski, R. Physcomitrella patens is highly tolerant against drought, salt and osmotic stress. Planta 2005, 220, 384–394. [Google Scholar] [CrossRef]
- Benito, B.; Rodrıguez-Navarro, A. Molecular cloning and characterization of a sodium-pump ATPase of the moss Physcomitrella patens. Plant J. 2003, 36, 382–389. [Google Scholar] [CrossRef]
- Sabovljević, A.; Vujičić, M.; Skorić, M.; Bajić-Ljubičić, J.; Sabovljević, M. Axenically culturing the bryophytes: Establishment and propagation of the pleurocarpous moss Thamnobryum alopecurum Nieuwland ex Gangulee (Bryophyta, Neckeraceae) in in vitro conditions. Pak. J. Bot. 2012, 44, 339–344. [Google Scholar]
- Sabovljević, M.; Papp, B.; Sabovljević, A.; Vujičić, M.; Szurdoki, E.; Segarra-Moragues, J.G. In vitro micropropagation of rare and endangered moss Entosthodon hungaricus (Funariaceae). Biosci. J. 2012, 28, 632–640. [Google Scholar]
- Knop, W. Quantitative Untersuchungen ueber die Ernahrungsprozesse der Pflanzen. Landwirtsch. Vers. 1865, 7, 93–107. [Google Scholar]
- Sabovljević, M.S.; Weidinger, M.L.; Sabovljević, A.D.; Adlassing, W.; Lang, I. Is binding patterns of Zn(II) equal in different bryophytes? Microsc. Microanal. 2018, 24, 69–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tabacco tissue culture. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Sabovljević, A.; Sabovljević, M.; Jocković, N. In vitro culture and secondary metabolite isolation in bryophytes. In Protocols for In Vitro Cultures and Secondary Metabolite Analysis of Aromatic and Medicinal Plants; Methods in Molecular Biology; Jain, S.M., Saxena, P.K., Eds.; Humana Press: Totowa, NJ, USA, 2009; pp. 117–128. [Google Scholar] [CrossRef]
- Krook, J.; Vreugdenhil, D.; van der Plas, L.H.W. 2000. Uptake and phosphorylation of glucose and fructose in Daucus carota cell suspensions are differently regulated. Plant Physiol. Biochem. 2000, 38, 603–612. [Google Scholar] [CrossRef]
- Vujičić, M.; Sabovljević, A.; Milošević, S.; Segarra-Moragues, J.G.; Sabovljevic, M. Effects of abscisic acid (ABA) on the development of selected bryophyte species. Plant Biosyst. 2016, 150, 1023–1029. [Google Scholar] [CrossRef]
- R Core Team, R. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.R-project.org/ (accessed on 25 December 2022).








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experiment type I | KNOP (control): KNOP minimal medium, sugar-free MS/2 (control): MS mineral salts, half strength, sugar-free BCD (control): BCD mineral salts, sugar-free KNOP + S: KNOP minimal medium enriched with 0.05 M sucrose KNOP + F: KNOP minimal medium enriched with 0.1 M fructose MS/2 + S: MS mineral salts, half strength, enriched with 0.05 M sucrose MS/2 + F: MS mineral salts, half strength, enriched with 0.1 M fructose BCD + S: BCD mineral salts enriched with 0.05 M sucrose BCD + F: BCD mineral salts enriched with 0.1 M fructose |
| Experiment type II | KNOP (control): KNOP minimal medium, PGR free KNOP supplemented with 0.1 µm IBA KNOP supplemented with 0.1 µm BAP KNOP supplemented with 0.03 µm IBA and 0.1 µm BAP KNOP supplemented with 0.1 µm IBA and 0.1 µm BAP KNOP supplemented with 0.3 µm IBA and 0.1 µm BAP |
| Experiment type III | KNOP (control): KNOP minimal medium, NaCl free KNOP supplemented with 10 mM NaCl KNOP supplemented with 50 mM NaCl KNOP supplemented with 100 mM NaCl KNOP supplemented with 250 mM NaCl KNOP supplemented with 500 mM NaCl |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jadranin, B.Z.; Ćosić, M.V.; Božović, D.P.; Vujičić, M.M.; Ignatov, M.S.; Ignatova, E.A.; Sabovljević, A.D.; Sabovljević, M.S. An Insight into the Biology of the Rare and Peculiar Moss Pterygoneurum sibiricum (Pottiaceae): A Conservation Physiology Approach. Plants 2023, 12, 1359. https://doi.org/10.3390/plants12061359
Jadranin BZ, Ćosić MV, Božović DP, Vujičić MM, Ignatov MS, Ignatova EA, Sabovljević AD, Sabovljević MS. An Insight into the Biology of the Rare and Peculiar Moss Pterygoneurum sibiricum (Pottiaceae): A Conservation Physiology Approach. Plants. 2023; 12(6):1359. https://doi.org/10.3390/plants12061359
Chicago/Turabian StyleJadranin, Bojana Z., Marija V. Ćosić, Djordje P. Božović, Milorad M. Vujičić, Michael S. Ignatov, Elena A. Ignatova, Aneta D. Sabovljević, and Marko S. Sabovljević. 2023. "An Insight into the Biology of the Rare and Peculiar Moss Pterygoneurum sibiricum (Pottiaceae): A Conservation Physiology Approach" Plants 12, no. 6: 1359. https://doi.org/10.3390/plants12061359