
Bioactive Compounds in Moringa oleifera: Mechanisms of Action, Focus on Their Anti-Inflammatory Properties
 , ,
, ,  , , , , , , , , , and
, , , , , , , , , and
Abstract
:1. Introduction

2. Description of Moringa oleifera
3. Bioactive Components of M. oleifera and Their Mechanisms of Action in Inflammatory Diseases
3.1. M. oleifera Essential Oil and Fatty Acids
3.2. Bioactive Phytochemical Components
3.3. Anti-Inflammatory Activities of M. oleifera Compounds and Their Mechanisms of Action
3.3.1. Cardiovascular Protection and Anti-Hypertensive Activities of M. oleifera
3.3.2. Type 2 Diabetes: Chronic Inflammatory Disease and M. oleifera
3.3.3. Inflammatory Bowel Disease and M. oleifera
3.3.4. M. oleifera Potential Effects on Non-Alcoholic Fatty Liver Disease (NAFLD)
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Ezzat, S.M.; Jeevanandam, J.; Egbuna, C.; Kumar, S.; Ifemeje, J.C. Phytochemicals as Sources of Drugs. In Phytochemistry: An In-Silico and In-Vitro Update; Kumar, S., Egbuna, C., Eds.; Springer: Singapore, 2019; pp. 3–22. [Google Scholar]
- Kuete, V. Moringa oleifera. In Medicinal Spices and Vegetables from Africa; Kuete, V., Ed.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 485–496. [Google Scholar]
- Noubissi, P.A.; Njilifac, Q.; Tagne, M.A.F.; Nguepi, M.S.D.; Fondjo, A.F.; Emégam, N.K.; Mukam, J.N.; Zintchem, R.; Wambe, H.; Fankem, G.O.; et al. Anxiolytic and anti-colitis effects of Moringa oleifera leaf-aqueous extract on acetic acid-induced colon inflammation in rat. Biomed. Pharmacother. 2022, 154, 113652. [Google Scholar] [CrossRef] [PubMed]
- Foidl, N.; Makkar, H.; Becker, K. The Potential of Moringa oleifera for agricultural and industrial uses. In The Miracle Tree: The Multiple Attributes of Moringa; Fuglie, L.J., Ed.; CTA: Wageningen, The Netherlands; CWS: Dakar, Senegal, 2001; pp. 45–76. [Google Scholar]
- Hodas, F.; Zorzenon, M.R.T.; Milani, P.G. Moringa oleifera potential as a functional food and a natural food additive: A biochemical approach. An. Acad. Bras. Cienc. 2021, 93 (Suppl. 4), e20210571. [Google Scholar] [CrossRef] [PubMed]
- Martín Ortega, A.M.; Segura Campos, M.R. Medicinal Plants and Their Bioactive Metabolites in Cancer Prevention and Treatment. In Bioactive Compounds; Segura Campos, M.R., Ed.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 85–109. [Google Scholar] [CrossRef]
- Imran, M.; Hussain, G.; Hameed, A.; Iftikhar, I.; Ibrahim, M.; Asghar, R.; Nisar, I.; Farooq, T.; Khalid, T.; Rehman, K.; et al. Metabolites of Moringa oleifera Activate Physio-Biochemical Pathways for an Accelerated Functional Recovery after Sciatic Nerve Crush Injury in Mice. Metabolites 2022, 12, 1242. [Google Scholar] [CrossRef] [PubMed]
- Llorent-Martínez, E.J.; Gordo-Moreno, A.I.; Córdova, M.L.F.-D.; Ruiz-Medina, A. Preliminary Phytochemical Screening and Antioxidant Activity of Commercial Moringa oleifera Food Supplements. Antioxidants 2023, 12, 110. [Google Scholar] [CrossRef] [PubMed]
- Ndlovu, S.S.; Chuturgoon, A.A.; Ghazi, T. Moringa oleifera Lam Leaf Extract Stimulates NRF2 and Attenuates ARV-Induced Toxicity in Human Liver Cells (HepG2). Plants 2023, 12, 1541. [Google Scholar] [CrossRef] [PubMed]
- Gomes, S.M.; Leitão, A.; Alves, A.; Santos, L. Incorporation of Moringa oleifera Leaf Extract in Yoghurts to Mitigate Children’s Malnutrition in Developing Countries. Molecules 2023, 28, 2526. [Google Scholar] [CrossRef] [PubMed]
- Manjunath, S.H.; Natarajan, P.; Swamy, V.H.; Sugur, K.; Dey, S.K.; Ranganathan, V.; Daniel, S.; Leihang, Z.; Sharon, V.; Chandrashekharappa, S.; et al. Development of Moringa oleifera as functional food targeting NRF2 signaling: Antioxidant and anti-inflammatory activity in experimental model systems. Food Funct. 2023, 14, 4734–4751. [Google Scholar] [CrossRef]
- Bajwa, M.N.; Khanum, M.; Zaman, G.; Ullah, M.A.; Farooq, U.; Waqas, M.; Ahmad, N.; Hano, C.; Abbasi, B.H. Effect of Wide-Spectrum Monochromatic Lights on Growth, Phytochemistry, Nutraceuticals, and Antioxidant Potential of In Vitro Callus Cultures of Moringa oleifera. Molecules 2023, 28, 1497. [Google Scholar] [CrossRef]
- Dzuvor, C.K.O.; Pan, S.; Amanze, C.; Amuzu, P.; Asakiya, C.; Kubi, F. Bioactive components from Moringa oleifera seeds: Production, functionalities and applications—A critical review. Crit. Rev. Biotechnol. 2021, 42, 271–293. [Google Scholar] [CrossRef]
- Singh, B.N.; Singh, B.R.; Singh, R.L.; Prakash, D.; Dhakarey, R.; Upadhyay, G.; Singh, H.B. Oxidative DNA damage protective activity, antioxidant and anti-quorum sensing potentials of Moringa oleifera. Food Chem. Toxicol. 2009, 47, 1109–1116. [Google Scholar] [CrossRef]
- Singh, R.G.; Negi, P.S.; Radha, C. Phenolic composition, antioxidant and antimicrobial activities of free and bound phenolic extracts of Moringa oleifera seed flour. J. Funct. Foods 2013, 5, 1883–1891. [Google Scholar] [CrossRef]
- Wang, S.; Liu, S.; Hao, G.; Zhao, L.; Lü, X.; Wang, H.; Wang, L.; Zhang, J.; Ge, W. Antimicrobial activity and mechanism of isothiocyanate from Moringa oleifera seeds against Bacillus cereus and Cronobacter sakazakii and its application in goat milk. Food Control 2022, 139, 109067. [Google Scholar] [CrossRef]
- Sodvadiya, M.; Patel, H.; Mishra, A.; Nair, S. Emerging Insights into Anticancer Chemopreventive Activities of Nutraceutical Moringa oleifera: Molecular Mechanisms, Signal Transduction and In Vivo Efficacy. Curr. Pharmacol. Rep. 2020, 6, 38–51. [Google Scholar] [CrossRef]
- Amina, M.; Al Musayeib, N.M.; Alarfaj, N.A.; El-Tohamy, M.F.; Orabi, H.E.; Bukhari, S.I.; Mahmoud, A.Z. Exploiting the Potential of Moringa oleifera Oil/Polyvinyl Chloride Polymeric Bionanocomposite Film Enriched with Silver Nanoparticles for Antimicrobial Activity. Int. J. Polym. Sci. 2019, 2019, 5678149. [Google Scholar] [CrossRef]
- Sultan, R.; Ahmed, A.; Wei, L.; Saeed, H.; Islam, M.; Ishaq, M. The anticancer potential of chemical constituents of Moringa oleifera targeting CDK-2 inhibition in estrogen receptor positive breast cancer using in-silico and in vitro approches. BMC Complement. Med. Ther. 2023, 23, 396. [Google Scholar] [CrossRef] [PubMed]
- Meireles, D.; Gomes, J.; Lopes, L.; Hinzmann, M.; Machado, J. A review of properties, nutritional and pharmaceutical applications of Moringa oleifera: Integrative approach on conventional and traditional Asian medicine. Adv. Trad. Med. (ADTM) 2020, 20, 495–515. [Google Scholar] [CrossRef]
- Kumar, S.; Verma, P.K.; Shukla, A.; Singh, R.K.; Patel, A.K.; Yadav, L.; Kumar, S.; Kumar, N.; Kaushalendra; Acharya, A. Moringa oleifera L. leaf extract induces cell cycle arrest and mitochondrial apoptosis in Dalton’s Lymphoma: An in vitro and in vivo study. J. Ethnopharmacol. 2023, 302 Pt A, 115849. [Google Scholar] [CrossRef]
- Aderinola, T.A.; Alashi, A.M.; Fagbemi, A.N.; Enujiugha, V.N.; Aluko, R.E.; Fagbemi, T.N.; Desk, S. Moringa oleifera flour protein fractions as food ingredients with antioxidant properties. SDRP J. Food Sci. Technol. 2019, 4, 720–728. [Google Scholar] [CrossRef]
- Aderinola, T.A.; Alashi, A.M.; Nwachukwu, I.D.; Fagbemi, T.N.; Enujiugha, V.N.; Aluko, R.E.; Nwachukwu, I.D. Antihypertensive and Antioxidant Properties of Moringa Oleifera Seed Enzymatic Protein Hydrolysate and Ultrafiltration Fractions. Curr. Top. Nutraceutical. Res. 2019, 17, 437–444. [Google Scholar]
- Aderinola, T.A.; Alashi, A.M.; Nwachukwu, I.D.; Fagbemi, T.N.; Enujiugha, V.N.; Aluko, R.E. In vitro digestibility, structural and functional properties of Moringa oleifera seed proteins. Food Hydrocoll. 2020, 101, 105574. [Google Scholar] [CrossRef]
- Aderinola, T.A.; Fagbemi, T.N.; Enujiugha, V.N.; Alashi, A.M.; Aluko, R.E. Amino acid composition and antioxidant properties of Moringa oleifera seed protein isolate and enzymatic hydrolysates. Heliyon 2018, 4, e00877. [Google Scholar] [CrossRef]
- Garza, N.G.G.; Koyoc, J.A.C.; Castillo, J.A.T.; Zambrano, E.A.G.; Ancona, D.B.; Guerrero, L.C.; García, S.R.S. Biofunctional properties of bioactive peptide fractions from protein isolates of moringa seed (Moringa oleifera). J. Food Sci. Technol. 2017, 54, 4268–4276. [Google Scholar] [CrossRef] [PubMed]
- Liang, L.; Wang, C.; Li, S.; Chu, X.; Sun, K. Nutritional compositions of Indian Moringa oleifera seed and antioxidant activity of its polypeptides. Food Sci. Nutr. 2019, 7, 1754–1760. [Google Scholar] [CrossRef] [PubMed]
- Liang, L.-L.; Cai, S.-Y.; Gao, M.; Chu, X.-M.; Pan, X.-Y.; Gong, K.-K.; Xiao, C.-W.; Chen, Y.; Zhao, Y.-Q.; Wang, B.; et al. Purification of antioxidant peptides of Moringa oleifera seeds and their protective effects on H2O2 oxidative damaged Chang liver cells. J. Funct. Foods 2020, 64, 103698. [Google Scholar] [CrossRef]
- Oluduro, O.A.; Aderiye, B.I.; Connolly, J.D.; Akintayo, E.T.; Famurewa, O. Characterization and antimicrobial activity of 4-(β-d-glucopyranosyl-1→4-α-l-rhamnopyranosyloxy)-benzyl thiocarboxamide: A novel bioactive compound from Moringa oleifera seed extract. Folia Microbiol. 2010, 55, 422–426. [Google Scholar] [CrossRef] [PubMed]
- Jeon, S.R.; Lee, K.H.; Shin, D.H.; Kwon, S.S.; Hwang, J.S. Synergistic antimicrobial efficacy of mesoporous ZnO loaded with 4-(α-l-rhamnosyloxy)-benzyl isothiocyanate isolated from the Moringa oleifera seed. J. Gen. Appl. Microbiol. 2014, 60, 251–255. [Google Scholar] [CrossRef] [PubMed]
- Galuppo, M.; De Nicola, G.R.; Iori, R.; Dell’Utri, P.; Bramanti, P.; Mazzon, E. Antibacterial activity of glucomoringin bioactivated with myrosinase against two important pathogens affecting the health of long-term patients in hospitals. Molecules 2013, 18, 14340–14348. [Google Scholar] [CrossRef] [PubMed]
- Galuppo, M.; Giacoppo, S.; De Nicola, G.R.; Iori, R.; Navarra, M.; Lombardo, G.E.; Bramanti, P.; Mazzon, E. Antiinflammatory activity of glucomoringin isothiocyanate in a mouse model of experimental autoimmune encephalomyelitis. Fitoterapia 2014, 95, 160–174. [Google Scholar] [CrossRef]
- Giacoppo, S.; Galuppo, M.; De Nicola, G.R.; Iori, R.; Bramanti, P.; Mazzon, E. 4(α-l-rhamnosyloxy)-benzyl isothiocyanate, a bioactive phytochemical that attenuates secondary damage in an experimental model of spinal cord injury. Bioorganic Med. Chem. 2015, 23, 80–88. [Google Scholar] [CrossRef]
- Jaja-Chimedza, A.; Graf, B.L.; Simmler, C.; Kim, Y.; Kuhn, P.; Pauli, G.F.; Raskin, I. Biochemical characterization and anti-inflammatory properties of an isothiocyanate-enriched moringa (Moringa oleifera) seed extract. PLoS ONE 2017, 12, e0182658. [Google Scholar] [CrossRef]
- Jaja-Chimedza, A.; Zhang, L.; Wolff, K.; Graf, B.L.; Kuhn, P.; Moskal, K.; Carmouche, R.; Newman, S.; Salbaum, J.M.; Raskin, I. A dietary isothiocyanate-enriched moringa (Moringa oleifera) seed extract improves glucose tolerance in a high-fat-diet mouse model and modulates the gut microbiome. J. Funct. Foods 2018, 47, 376–385. [Google Scholar] [CrossRef]
- Dhongade, H.K.J.; Paikra, B.K.; Gidwani, B. Phytochemistry and Pharmacology of Moringa oleifera Lam. J. Pharmacopunct. 2017, 20, 194–200. [Google Scholar] [CrossRef] [PubMed]
- Tan, W.S.; Arulselvan, P.; Karthivashan, G.; Fakurazi, S. Moringa oleifera flower extract suppresses the activation of inflammatory mediators in lipopolysaccharide-stimulated RAW 264.7 macrophages via NF-κB pathway. Mediat. Inflamm. 2015, 2015, 720171. [Google Scholar] [CrossRef] [PubMed]
- Vasanth, K.; Minakshi, G.; Ilango, K.; Kumar, R.; Agrawal, A.; Dubey, G. Moringa oleifera attenuates the release of pro-inflammatory cytokines in lipopolysaccharide stimulated human monocytic cell line. Ind. Crop. Prod. 2015, 77, 44–50. [Google Scholar] [CrossRef]
- Adebayo, S.A.; Amoo, S.O. South African botanical resources: A gold mine of natural pro-inflammatory enzyme inhibitors? South Afr. J. Bot. 2019, 123, 214–227. [Google Scholar] [CrossRef]
- Albaayit, S.F.A.; Al-Khafaji, A.S.K.; Alnaimy, H.S. In vitro macrophage nitric oxide and interleukin-1 beta suppression by moringa peregrina seed. Turk. J. Pharm. Sci. 2019, 16, 362–365. [Google Scholar] [CrossRef] [PubMed]
- Cui, C.; Chen, S.; Wang, X.; Yuan, G.; Jiang, F.; Chen, X.; Wang, L. Characterization of Moringa oleifera roots polysaccharide MRP-1 with anti-inflammatory effect. Int. J. Biol. Macromol. 2019, 132, 844–851. [Google Scholar] [CrossRef] [PubMed]
- Luetragoon, T.; Sranujit, R.P.; Noysang, C.; Thongsri, Y.; Potup, P.; Suphrom, N.; Nuengchamnong, N.; Usuwanthim, K. Bioactive compounds in Moringa oleiferaLam. leaves inhibit the pro-inflammatory mediators in lipopolysaccharide-induced human monocyte-derived macrophages. Molecules 2020, 25, 191–207. [Google Scholar] [CrossRef]
- Afonso, A.F.; Pereira, O.R.; Cardoso, S.M. Health-Promoting Effects of Thymus Phenolic-Rich Extracts: Antioxidant, Anti-inflammatory and Antitumoral Properties. Antioxidants 2020, 9, 814. [Google Scholar] [CrossRef]
- Lopez-Rodriguez, N.A.; Gaytán-Martínez, M.; Reyes-Vega, M.d.l.L.; Loarca-Piña, G. Glucosinolates and Isothiocyanates from Moringa oleifera: Chemical and Biological Approaches. Plant Foods Hum. Nutr. 2020, 75, 447–457. [Google Scholar] [CrossRef]
- Fahey, J. Moringa oleifera: A Review of the Medical Evidence for Its Nutritional, Therapeutic, and Prophylactic Properties. Part 1. Trees Life J. 2005, 1, 1–15. [Google Scholar]
- Maurya, S.K.; Singh, A.K. Clinical Efficacy of Moringa oleifera Lam. Stems Bark in Urinary Tract Infections. Int. Sch. Res. Not. 2014, 2014, 906843. [Google Scholar] [CrossRef] [PubMed]
- Rani, N.Z.A.; Husain, K.; Kumolosasi, E. Moringa genus: A review of phytochemistry and pharmacology. Front. Pharmacol. 2018, 9, 108. [Google Scholar] [CrossRef] [PubMed]
- Olson, M.E.; Carlquist, S. Stem and root anatomical correlations with life form diversity, ecology, and systematics in Moringa (Moringaceae). Bot. J. Linnean Soc. 2001, 13i5, 315–348. [Google Scholar] [CrossRef]
- Nadeem, M.; Imran, M. Promising features of Moringa oleifera oil: Recent updates and perspectives. Lipids Heal. Dis. 2016, 15, 212. [Google Scholar] [CrossRef] [PubMed]
- Basuny, A.M.; Al-Marzouq, M.A. Biochemical Studies on Moringa Oleifera Seed Oil. MOJ Food Process. Technol. 2016, 2, 40–46. [Google Scholar] [CrossRef]
- Huang, Y.; Xiao, D.; Burton-Freeman, B.M.; Edirisinghe, I. Chemical Changes of Bioactive Phytochemicals during Thermal Processing. In Reference Module in Food Science; Elsevier: Amsterdam, The Netherlands, 2016. [Google Scholar]
- Prabu, S.L.; Umamaheswari, A.; Puratchikody, A. Phytopharmacological potential of the natural gift Moringa oleifera Lam and its therapeutic application: An overview. Asian Pac. J. Trop. Med. 2019, 12, 485–498. [Google Scholar] [CrossRef]
- Ma, Z.; Ahmad, J.; Zhang, H.; Khan, I.; Muhammad, S. Evaluation of phytochemical and medicinal properties of Moringa (Moringa oleifera) as a potential functional food. South. Afr. J. Bot. 2020, 129, 40–46. [Google Scholar] [CrossRef]
- Kołodziejski, D.; Koss-Mikołajczyk, I.; Abdin, A.Y.; Jacob, C.; Bartoszek, A. Chemical Aspects of Biological Activity of Isothiocyanates and Indoles, the Products of Glucosinolate Decomposition. Curr. Pharm. Des. 2019, 25, 1717–1728. [Google Scholar] [CrossRef]
- Saini, R.K.; Shetty, N.P.; Giridhar, P. Carotenoid content in vegetative and reproductive parts of commercially grown Moringa oleifera Lam. cultivars from India by LC–APCI–MS. Eur. Food Res. Technol. 2014, 238, 971–978. [Google Scholar] [CrossRef]
- Bhatt, T.; Patel, K. Carotenoids: Potent to Prevent Diseases Review. Nat. Prod. Bioprospecting 2020, 10, 109–117. [Google Scholar] [CrossRef]
- Haroen, U.; Syafwan, S.; Kurniawan, K.; Budiansyah, A. Determination of nutrient content, β-carotene, and antioxidant activity of Moringa oleifera extraction using organic solution. J. Adv. Veter-Anim. Res. 2022, 9, 246–254. [Google Scholar] [CrossRef] [PubMed]
- Muteeb, G.; Aatif, M.; Farhan, M.; Alsultan, A.; Alshoaibi, A.; Alam, M.W. Leaves of Moringa oleifera Are Potential Source of Bioactive Compound β-Carotene: Evidence from In Silico and Quantitative Gene Expression Analysis. Molecules 2023, 28, 1578. [Google Scholar] [CrossRef] [PubMed]
- Dey, P.; Kundu, A.; Kumar, A.; Gupta, M.; Lee, B.M.; Bhakta, T.; Dash, S.; Kim, H.S. Analysis of alkaloids (indole alkaloids, isoquinoline alkaloids, tropane alkaloids). In Recent Advances in Natural Products Analysis; Sanches Silva, A., Nabavi, S.F., Saeedi, M., Nabavi, S.M., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 1–9. [Google Scholar] [CrossRef]
- Xie, J.; Luo, F.-X.; Shi, C.-Y.; Jiang, W.-W.; Qian, Y.-Y.; Yang, M.-R.; Song, S.; Dai, T.-Y.; Peng, L.; Gao, X.-Y.; et al. Moringa oleifera Alkaloids Inhibited PC3 Cells Growth and Migration Through the COX-2 Mediated Wnt/β-Catenin Signaling Pathway. Front. Pharmacol. 2020, 11, 523962. [Google Scholar] [CrossRef] [PubMed]
- Adedapo, A.A.; Etim, U.; Falayi, O.O.; Ogunpolu, B.S.; Omobowale, T.O.; Oyagbemi, A.A.; Oguntibeju, O.O. Methanol stem extract of Moringa oleifera mitigates glycerol-induced acute kidney damage in rats through modulation of KIM-1 and NF-kB signaling pathways. Sci. Afr. 2020, 9, e00493. [Google Scholar] [CrossRef]
- Fongang Fotsing, Y.S.; Bankeu Kezetas, J.J. Terpenoids as Important Bioactive Constituents of Essential Oils. In Essential Oils—Bioactive Compounds, New Perspectives and Applications; Santana de Oliveira, M., Almeida da Costa, W., Gomes Silva, S., Eds.; IntechOpen: London, UK, 2020. [Google Scholar] [CrossRef]
- Boncan, D.A.T.; Tsang, S.S.; Li, C.; Lee, I.H.; Lam, H.-M.; Chan, T.-F.; Hui, J.H. Terpenes and terpenoids in plants: Interactions with environment and insects. Int. J. Mol. Sci. 2020, 21, 7382. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.K.; Rana, H.K.; Tshabalala, T.; Kumar, R.; Gupta, A.; Ndhlala, A.R.; Pandey, A.K. Phytochemical, nutraceutical and pharmacological attributes of a functional crop Moringa oleifera Lam: An overview. South. Afr. J. Bot. 2020, 129, 209–220. [Google Scholar] [CrossRef]
- Korniluk, A.; Koper, O.; Kemona, H.; Dymicka-Piekarska, V. From inflammation to cancer. Ir. J. Med. Sci. 2017, 186, 57–62. [Google Scholar] [CrossRef]
- Germolec, D.R.; Shipkowski, K.A.; Frawley, R.P.; Evans, E. Markers of Inflammation. In Immunotoxicity Testing: Methods and Protocols, Methods in Molecular Biology; Humana Press: New York, NY, USA, 2018; pp. 57–79. [Google Scholar] [CrossRef]
- Rajan, T.S.; Giacoppo, S.; Iori, R.; De Nicola, G.R.; Grassi, G.; Pollastro, F.; Bramanti, P.; Mazzon, E. Anti-inflammatory and antioxidant effects of a combination of cannabidiol and moringin in LPS-stimulated macrophages. Fitoterapia 2016, 112, 104–115. [Google Scholar] [CrossRef]
- Ribaudo, G.; Povolo, C.; Zagotto, G. Moringa oleifera Lam.: A Rich Source of Phytoactives for the Health of Human Being. In Studies in Natural Products Chemistry, 1st ed.; Rahman, A., Ed.; Elsevier: Amsterdam, The Netherlands, 2019. [Google Scholar] [CrossRef]
- Lazou, A.; Ikonomidis, I.; Bartekova, M.; Benedek, T.; Makavos, G.; Palioura, D.; Fuentes, H.C.; Andreadou, I. Chronic inflammatory diseases, myocardial function and cardioprotection. Br. J. Pharmacol. 2020, 177, 5357–5374. [Google Scholar] [CrossRef]
- Mason, J.C.; Libby, P. Cardiovascular disease in patients with chronic inflammation: Mechanisms underlying premature cardiovascular events in rheumatologic conditions. Eur. Hear. J. 2015, 36, 482–489. [Google Scholar] [CrossRef]
- Raaby, L.; Ahlehoff, O.; de Thurah, A. Psoriasis and cardiovascular events: Updating the evidence. Arch. Dermatol. Res. 2017, 309, 225–228. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.-H.; Tian, F. Inflammatory bowel disease and cardiovascular disease incidence and mortality: A meta-analysis. Eur. J. Prev. Cardiol. 2018, 25, 1623–1631. [Google Scholar] [CrossRef] [PubMed]
- Grundy, S.M.; Stone, N.J.; Bailey, A.L.; Beam, C.; Birtcher, K.K.; Blumenthal, R.S.; Braun, L.T.; de Ferranti, S.; Faiella-Tommasino, J.; Forman, D.E.; et al. 2018 AHA/ACC/AACVPR/AAPA/ABC/ACPM/ADA/AGS/APhA/ASPC/NLA/PCNA Guideline on the Management of Blood Cholesterol: A Report of the American College of Cardiology/American Heart Association Task Force on Clinical Practice Guidelines. Circulation 2019, 139, E1082–E1143. [Google Scholar] [CrossRef]
- Aksentijevich, M.; Lateef, S.S.; Anzenberg, P.; Dey, A.K.; Mehta, N.N. Chronic inflammation, cardiometabolic diseases and effects of treatment: Psoriasis as a human model. Trends Cardiovasc. Med. 2020, 30, 472–478. [Google Scholar] [CrossRef] [PubMed]
- Mehta, N.N.; Dey, A.K.; Maddineni, R.; Kraus, W.E.; Huffman, K.M. GlycA measured by NMR spectroscopy is associated with disease activity and cardiovascular disease risk in chronic inflammatory diseases. Am. J. Prev. Cardiol. 2020, 4, 100120. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Li, G.; Chang, D.; Su, X. YKL-40 as a novel biomarker in cardio-metabolic disorders and inflammatory diseases. Clin. Chim. Acta 2020, 511, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Adefegha, S.A.; Oboh, G.; Iyoha, A.E.; Oyagbemi, A.A. Comparative effects of horseradish (Moringa oleifera) leaves and seeds on blood pressure and crucial enzymes relevant to hypertension in rat. PharmaNutrition 2019, 9, 100152. [Google Scholar] [CrossRef]
- Nelin, L.D.; Stenger, M.R.; Malleske, D.T.; Chicoine, L.G. Vascular Arginase and Hypertension. Curr. Hypertens. Rev. 2007, 3, 242–249. [Google Scholar] [CrossRef]
- Cheenpracha, S.; Park, E.-J.; Yoshida, W.Y.; Barit, C.; Wall, M.; Pezzuto, J.M.; Chang, L.C. Potential anti-inflammatory phenolic glycosides from the medicinal plant Moringa oleifera fruits. Bioorganic Med. Chem. 2010, 18, 6598–6602. [Google Scholar] [CrossRef]
- Gilani, A.H.; Aftab, K.; Suria, A.; Siddiqui, S.; Salem, R.; Siddiqui, B.S.; Faizi, S. Pharmacological Studies on Hypotensive and Spasmolytic Activities of Pure Compounds from Moringa oleifera. Phytother. Res. 1994, 8, 87–91. [Google Scholar] [CrossRef]
- Zibrila, A.I.; Li, Y.; Wang, Z.; Zhao, G.; Liu, H.; Leng, J.; Ali, A.; Osei, J.A.; Kang, Y.-M.; Liu, J. Acetylcholinesterase inhibition with pyridostigmine attenuates hypertension and neuroinflammation in the paraventricular nucleus in rat model for Preeclampsia. Int. Immunopharmacol. 2021, 101, 108365. [Google Scholar] [CrossRef] [PubMed]
- Nebbioso, M.; Pascarella, A.; Cavallotti, C.; Pescosolido, N. Monoamine oxidase enzymes and oxidative stress in the rat optic nerve: Age-related changes. Int. J. Exp. Pathol. 2012, 93, 401–405. [Google Scholar] [CrossRef] [PubMed]
- Elhwuegi, A. The Wonders of Phosphodiesterase-5 Inhibitors: A Majestic History. Ann. Med. Heal. Sci. Res. 2016, 6, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Oboh, G.; Oyeleye, S.I.; Akintemi, O.A.; Olasehinde, T.A. Moringa oleifera supplemented diet modulates nootropic-related biomolecules in the brain of STZ-induced diabetic rats treated with acarbose. Metab. Brain Dis. 2018, 33, 457–466. [Google Scholar] [CrossRef] [PubMed]
- Clarke, N.E.; Turner, A.J. Angiotensin-Converting Enzyme 2: The First Decade. Int. J. Hypertens. 2012, 2012, 307315. [Google Scholar] [CrossRef] [PubMed]
- Oboh, G.; Ademiluyi, A.O.; Ademosun, A.O.; Olasehinde, T.A.; Oyeleye, S.I.; Boligon, A.A.; Athayde, M.L. Phenolic Extract fromMoringa oleifera Leaves Inhibits Key Enzymes Linked to Erectile Dysfunction and Oxidative Stress in Rats’ Penile Tissues. Biochem. Res. Int. 2015, 2015, 175950. [Google Scholar] [CrossRef] [PubMed]
- Ramamurthy, S.; Varghese, S.; Sudarsan, S.; Muruganandhan, J.; Mushtaq, S.; Patil, P.B.; Raj, A.T.; Zanza, A.; Testarelli, L.; Patil, S. Moringa oleifera: Antioxidant, Anticancer, Anti-inflammatory, and Related Properties of Extracts in Cell Lines: A Review of Medicinal Effects, Phytochemistry, and Applications. J. Contemp. Dent. Pract. 2021, 22, 1483–1492. [Google Scholar] [CrossRef]
- Alia, F.; Putri, M.; Anggraeni, N.; A Syamsunarno, M.R.A. The Potency of Moringa oleifera Lam. as Protective Agent in Cardiac Damage and Vascular Dysfunction. Front. Pharmacol. 2022, 12, 724439. [Google Scholar] [CrossRef]
- Aziz, M.; Yadav, K. Pathogenesis of Atherosclerosis A Review. Med. Clin. Rev. 2016, 2, 22. [Google Scholar] [CrossRef]
- Frąk, W.; Wojtasińska, A.; Lisińska, W.; Młynarska, E.; Franczyk, B.; Rysz, J. Pathophysiology of Cardiovascular Diseases: New Insights into Molecular Mechanisms of Atherosclerosis, Arterial Hypertension, and Coronary Artery Disease. Biomedicines 2022, 10, 1938. [Google Scholar] [CrossRef]
- Eller-Vainicher, C.; Cairoli, E.; Grassi, G.; Grassi, F.; Catalano, A.; Merlotti, D.; Falchetti, A.; Gaudio, A.; Chiodini, I.; Gennari, L. Pathophysiology and Management of Type 2 Diabetes Mellitus Bone Fragility. J. Diabetes Res. 2020, 2020, 7608964. [Google Scholar] [CrossRef] [PubMed]
- Fagninou Nonsito, A.; Ulbad, T.P.; Magloire, N.; Ruffine, F.; Koutinhouin, G.B.; Akadiri, Y. Diabetes mellitus: Classification, epidemiology, physiopathology, immunology, risk factors, prevention and nutrition. Int. J. Adv. Res. 2019, 7, 855–863. [Google Scholar]
- Liu, H.; Cao, M.-M.; Wang, Y.; Li, L.-C.; Zhu, L.-B.; Xie, G.-Y.; Li, Y.-B. Endoplasmic reticulum stress is involved in the connection between inflammation and autophagy in type 2 diabetes. Gen. Comp. Endocrinol. 2015, 210, 124–129. [Google Scholar] [CrossRef] [PubMed]
- Trivedi, P.M.; Fynch, S.; Kennedy, L.M.; Chee, J.; Krishnamurthy, B.; O’reilly, L.A.; Strasser, A.; Kay, T.W.H.; Thomas, H.E. Soluble FAS ligand is not required for pancreatic islet inflammation or beta-cell destruction in non-obese diabetic mice. Cell Death Discov. 2019, 5, 136. [Google Scholar] [CrossRef] [PubMed]
- Adedapo, A.A.; Ogunmiluyi, I.O.; Falayi, O.O.; Ogunpolu, B.S.; Oyagbemi, A.A.; Orishadipe, A.; Omobowale, T.O.; Yakubu, M.A.; Oguntibeju, O.O. The lyophilized aqueous leaf extract of Moringa oleifera blunts streptozocin-induced diabetes in rats through upregulation of GLUT 4 signaling pathway and anti-oxidant effect. Sci. Afr. 2020, 10, e00619. [Google Scholar] [CrossRef]
- Bao, Y.; Xiao, J.; Weng, Z.; Lu, X.; Shen, X.; Wang, F. A phenolic glycoside from Moringa oleifera Lam. improves the carbohydrate and lipid metabolisms through AMPK in db/db mice. Food Chem. 2020, 311, 125948. [Google Scholar] [CrossRef]
- Wu, H.; Deng, X.; Shi, Y.; Su, Y.; Wei, J.; Duan, H. PGC-1α, glucose metabolism and type 2 diabetes mellitus. J. Endocrinol. 2016, 229, R99–R115. [Google Scholar] [CrossRef]
- Gao, T.; Chen, S.; Han, Y.; Zhang, D.; Tan, Y.; He, Y.; Liu, M. Ameliorating Inflammation in Insulin-resistant Rat Adipose Tissue with Abdominal Massage Regulates SIRT1/NF-κB Signaling. Cell Biochem. Biophys. 2022, 80, 579–589. [Google Scholar] [CrossRef]
- Njume, C.; Donkor, O.; McAinch, A.J. Predisposing factors of type 2 diabetes mellitus and the potential protective role of native plants with functional properties. J. Funct. Foods 2019, 53, 115–124. [Google Scholar] [CrossRef]
- Praparatana, R.; Maliyam, P.; Barrows, L.R.; Puttarak, P. Flavonoids and Phenols, the Potential Anti-Diabetic Compounds from Bauhinia strychnifolia Craib. Stem. Molecules 2022, 27, 2393. [Google Scholar] [CrossRef]
- Bharti, S.K.; Krishnan, S.; Kumar, A.; Kumar, A. Antidiabetic phytoconstituents and their mode of action on metabolic pathways. Ther. Adv. Endocrinol. Metab. 2018, 9, 81–100. [Google Scholar] [CrossRef] [PubMed]
- Tarmizi, A.A.A.; Ramli, N.N.N.; Adam, S.H.; Mutalib, M.A.; Mokhtar, M.H.; Tang, S.G.H. Phytofabrication of Selenium Nanoparticles with Moringa oleifera (MO-SeNPs) and Exploring Its Antioxidant and Antidiabetic Potential. Molecules 2023, 28, 5322. [Google Scholar] [CrossRef] [PubMed]
- Shabab, S.; Gholamnezhad, Z.; Mahmoudabady, M. Protective effects of medicinal plant against diabetes induced cardiac disorder: A review. J. Ethnopharmacol. 2021, 265, 113328. [Google Scholar] [CrossRef] [PubMed]
- Xiong, W.; Ma, H.; Zhang, Z.; Jin, M.; Wang, J.; Xu, Y.; Wang, Z. The protective effect of icariin and phosphorylated icariin against LPS-induced intestinal epithelial cells injury. Biomed. Pharmacother. 2019, 118, 109246. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Li, M.; Wang, F.; Yang, Y.; Wang, Q.; Zhao, Y.; Du, F.; Chen, Y.; Shen, J.; Zhao, Q.; et al. The role of intestinal stem cell within gut homeostasis: Focusing on its interplay with gut microbiota and the regulating pathways. Int. J. Biol. Sci. 2022, 18, 5185–5206. [Google Scholar] [CrossRef] [PubMed]
- Passos, M.D.C.F.; Moraes-Filho, J.P. Intestinal microbiota in digestive diseases. Arq. De Gastroenterol. 2017, 54, 255–262. [Google Scholar] [CrossRef] [PubMed]
- Salguero, M.V.; Al-Obaide, M.A.I.; Singh, R.; Siepmann, T.; Vasylyeva, T.L. Dysbiosis of Gram-negative gut microbiota and the associated serum lipopolysaccharide exacerbates inflammation in type 2 diabetic patients with chronic kidney disease. Exp. Ther. Med. 2019, 18, 3461–3469. [Google Scholar] [CrossRef] [PubMed]
- Guazelli, C.F.; Fattori, V.; Ferraz, C.R.; Borghi, S.M.; Casagrande, R.; Baracat, M.M.; Verri, W.A. Antioxidant and anti-inflammatory effects of hesperidin methyl chalcone in experimental ulcerative colitis. Chem. Interactions 2020, 333, 109315. [Google Scholar] [CrossRef]
- Liu, K.; Li, G.; Guo, W.; Zhang, J. The protective effect and mechanism of pedunculoside on DSS (dextran sulfate sodium) induced ulcerative colitis in mice. Int. Immunopharmacol. 2020, 88, 107017. [Google Scholar] [CrossRef]
- Saldanha, E.; Saxena, A.; Kaur, K.; Kalekhan, F.; Venkatesh, P.; Fayad, R.; Rao, S.; George, T.; Baliga, M.S. Polyphenols in the Prevention of Ulcerative Colitis: A Revisit. In Dietary Interventions in Gastrointestinal Diseases; Watson, R.R., Preedy, V.R., Eds.; Academic Press: Cambridge, MA, USA; Elsevier Inc.: London, UK, 2019; Volume 2, pp. 277–287. [Google Scholar] [CrossRef]
- Gholap, P.A.; Nirmal, S.A.; Pattan, S.R.; Pal, S.C.; Mandal, S.C. Potential of Moringa oleifera root and Citrus sinensis fruit rind extracts in the treatment of ulcerative colitis in mice. Pharm. Biol. 2012, 50, 1297–1302. [Google Scholar] [CrossRef]
- Kim, Y.; Wu, A.G.; Jaja-Chimedza, A.; Graf, B.L.; Waterman, C.; Verzi, M.P.; Raskin, I. Isothiocyanate-enriched moringa seed extract alleviates ulcerative colitis symptoms in mice. PLoS ONE 2017, 12, e0184709. [Google Scholar] [CrossRef] [PubMed]
- Nejabati, H.R.; Roshangar, L. Kaempferol: A potential agent in the prevention of colorectal cancer. Physiol. Rep. 2022, 10, e15488. [Google Scholar] [CrossRef] [PubMed]
- Tao, J.; Huang, Z.; Wang, Y.; Liu, Y.; Zhao, T.; Wang, Y.; Tian, L.; Cheng, G. Ethanolic Extract from Pteris wallichiana Alleviates DSS-Induced Intestinal Inflammation and Intestinal Barrier Dysfunction by Inhibiting the TLR4/NF-κB Pathway and Regulating Tight Junction Proteins. Molecules 2022, 27, 3093. [Google Scholar] [CrossRef] [PubMed]
- Park, M.-Y.; Ji, G.E.; Sung, M.-K. Dietary kaempferol suppresses inflammation of dextran sulfate sodium-induced colitis in mice. Dig. Dis. Sci. 2011, 57, 355–363. [Google Scholar] [CrossRef] [PubMed]
- Peng, L.; Gao, X.; Nie, L.; Xie, J.; Dai, T.; Shi, C.; Tao, L.; Wang, Y.; Tian, Y.; Sheng, J. Astragalin Attenuates Dextran Sulfate Sodium (DSS)-Induced Acute Experimental Colitis by Alleviating Gut Microbiota Dysbiosis and Inhibiting NF-κB Activation in Mice. Front. Immunol. 2020, 11, 2058. [Google Scholar] [CrossRef] [PubMed]
- Hong, Z.-S.; Xie, J.; Wang, X.-F.; Dai, J.-J.; Mao, J.-Y.; Bai, Y.-Y.; Sheng, J.; Tian, Y. Moringa oleifera Lam. Peptide Remodels Intestinal Mucosal Barrier by Inhibiting JAK-STAT Activation and Modulating Gut Microbiota in Colitis. Front. Immunol. 2022, 13, 924178. [Google Scholar] [CrossRef] [PubMed]
- Gluvic, Z.; Tomasevic, R.; Bojovic, K.; Obradovic, M.; Isenovic, E.R. Non-alcoholic fatty liver disease: A multidisciplinary clinical practice approach—The institutional adaptation to existing Clinical Practice Guidelines. Emerg. Crit. Care Med. 2021, 2, 12–22. [Google Scholar] [CrossRef]
- Rinella, M.E.; Tacke, F.; Sanyal, A.J.; Anstee, Q.M. Report on the AASLD/EASL joint workshop on clinical trial endpoints in NAFLD. J. Hepatol. 2019, 71, 823–833. [Google Scholar] [CrossRef]
- Wang, X.J.; Malhi, H. Nonalcoholic Fatty Liver Disease. Ann. Intern. Med. 2018, 169, ITC65–ITC80. [Google Scholar] [CrossRef]
- Carr, R.M.; Oranu, A.; Khungar, V. Nonalcoholic Fatty Liver Disease: Pathophysiology and management. Gastroenterol. Clin. North. Am. 2016, 45, 639–652. [Google Scholar] [CrossRef]
- Sweet, P.H.; Khoo, T.; Nguyen, S. Nonalcoholic Fatty Liver Disease. Prim. Care Clin. Off. Primers 2017, 44, 599–607. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, X.-V.K.; Zhang, J.; Chin, K.L.; Bloom, S.; Nicoll, A.J. Is Hepatocellular Carcinoma in Fatty Liver Different to Non-Fatty Liver? Nutrients 2022, 14, 3875. [Google Scholar] [CrossRef] [PubMed]
- Manne, V.; Handa, P.; Kowdley, K.V. Pathophysiology of Nonalcoholic Fatty Liver Disease/Nonalcoholic Steatohepatitis. Clin. Liver Dis. 2018, 22, 23–37. [Google Scholar] [CrossRef]
- Petrelli, F.; Manara, M.; Colombo, S.; De Santi, G.; Ghidini, M.; Mariani, M.; Iaculli, A.; Rausa, E.; Rampulla, V.; Arru, M.; et al. Hepatocellular carcinoma in patients with nonalcoholic fatty liver disease: A systematic review and meta-analysis: HCC and Steatosis or Steatohepatitis. Neoplasia 2022, 30, 100809. [Google Scholar] [CrossRef] [PubMed]
- Püschel, G.P.; Klauder, J.; Henkel, J. Macrophages, Low-Grade Inflammation, Insulin Resistance and Hyperinsulinemia: A Mutual Ambiguous Relationship in the Development of Metabolic Diseases. J. Clin. Med. 2022, 11, 4358. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Wei, Y.; Huang, Q.; Chen, Y.; Zeng, K.; Yang, W.; Chen, J.; Chen, J. Important Hormones Regulating Lipid Metabolism. Molecules 2022, 27, 7052. [Google Scholar] [CrossRef]
- Berndt, N.; Hudert, C.A.; Eckstein, J.; Loddenkemper, C.; Henning, S.; Bufler, P.; Meierhofer, D.; Sack, I.; Wiegand, S.; Wallach, I.; et al. Alterations of Central Liver Metabolism of Pediatric Patients with Non-Alcoholic Fatty Liver Disease. Int. J. Mol. Sci. 2022, 23, 11072. [Google Scholar] [CrossRef]
- Palma, R.; Pronio, A.; Romeo, M.; Scognamiglio, F.; Ventriglia, L.; Ormando, V.M.; Lamazza, A.; Pontone, S.; Federico, A.; Dallio, M. The Role of Insulin Resistance in Fueling NAFLD Pathogenesis: From Molecular Mechanisms to Clinical Implications. J. Clin. Med. 2022, 11, 3649. [Google Scholar] [CrossRef]
- Asgari-Kafrani, A.; Fazilati, M.; Nazem, H. Hepatoprotective and antioxidant activity of aerial parts of Moringa oleifera in prevention of non-alcoholic fatty liver disease in Wistar rats. S. Afr. J. Bot. 2019, 129, 82–90. [Google Scholar] [CrossRef]
- Chen, Z.; Tian, R.; She, Z.; Cai, J.; Li, H. Role of oxidative stress in the pathogenesis of nonalcoholic fatty liver disease. Free Radic. Biol. Med. 2020, 152, 116–141. [Google Scholar] [CrossRef]
- Ma, Y.; Lee, G.; Heo, S.-Y.; Roh, Y.-S. Oxidative stress is a key modulator in the development of nonalcoholic fatty liver disease. Antioxidants 2021, 11, 91. [Google Scholar] [CrossRef] [PubMed]
- Morgan, C.; Sáez-Briones, P.; Barra, R.; Reyes, A.; Zepeda-Morales, K.; Constandil, L.; Ríos, M.; Ramírez, P.; Burgos, H.; Hernández, A. Prefrontal Cortical Control of Activity in Nucleus Accumbens Core Is Weakened by High-Fat Diet and Prevented by Co-Treatment with N-Acetylcysteine: Implications for the Development of Obesity. Int. J. Mol. Sci. 2022, 23, 10089. [Google Scholar] [CrossRef] [PubMed]
- Cho, A.-S.; Jeon, S.-M.; Kim, M.-J.; Yeo, J.; Seo, K.-I.; Choi, M.-S.; Lee, M.-K. Chlorogenic acid exhibits anti-obesity property and improves lipid metabolism in high-fat diet-induced-obese mice. Food Chem. Toxicol. 2010, 48, 937–943. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Cortegana, C.; García-Galey, A.; Tami, M.; del Pino, P.; Carmona, I.; López, S.; Alba, G.; Sánchez-Margalet, V. Role of leptin in non-alcoholic fatty liver disease. Biomedicines 2021, 9, 762. [Google Scholar] [CrossRef] [PubMed]
- Francisco, V.; Sanz, M.J.; Real, J.T.; Marques, P.; Capuozzo, M.; Eldjoudi, D.A.; Gualillo, O. Adipokines in Non-Alcoholic Fatty Liver Disease: Are We on the Road toward New Biomarkers and Therapeutic Targets? Biology 2022, 11, 1237. [Google Scholar] [CrossRef] [PubMed]
- Almatrafi, M.M.; Vergara-Jimenez, M.; Murillo, A.G.; Norris, G.H.; Blesso, C.N.; Fernandez, M.L. Moringa leaves prevent hepatic lipid accumulation and inflammation in guinea pigs by reducing the expression of genes involved in lipid metabolism. Int. J. Mol. Sci. 2017, 18, 1330. [Google Scholar] [CrossRef]
- Li, X.-J.; Mu, Y.-M.; Li, T.-T.; Yang, Y.-L.; Zhang, M.-T.; Li, Y.-S.; Zhang, W.K.; Tang, H.-B.; Shang, H.-C. Gynura procumbens Reverses Acute and Chronic Ethanol-Induced Liver Steatosis through MAPK/SREBP-1c-Dependent and -Independent Pathways. J. Agric. Food Chem. 2015, 63, 8460–8471. [Google Scholar] [CrossRef]
- Waterman, C.; Cheng, D.M.; Rojas-Silva, P.; Poulev, A.; Dreifus, J.; Lila, M.A.; Raskin, I. Stable, water extractable isothiocyanates from Moringa oleifera leaves attenuate inflammation in vitro. Phytochemistry 2014, 103, 114–122. [Google Scholar] [CrossRef]
- Joung, H.; Kim, B.; Park, H.; Lee, K.; Kim, H.-H.; Sim, H.-C.; Do, H.-J.; Hyun, C.-K.; Do, M.-S. Fermented Moringa oleifera Decreases Hepatic Adiposity and Ameliorates Glucose Intolerance in High-Fat Diet-Induced Obese Mice. J. Med. Food 2017, 20, 439–447. [Google Scholar] [CrossRef]







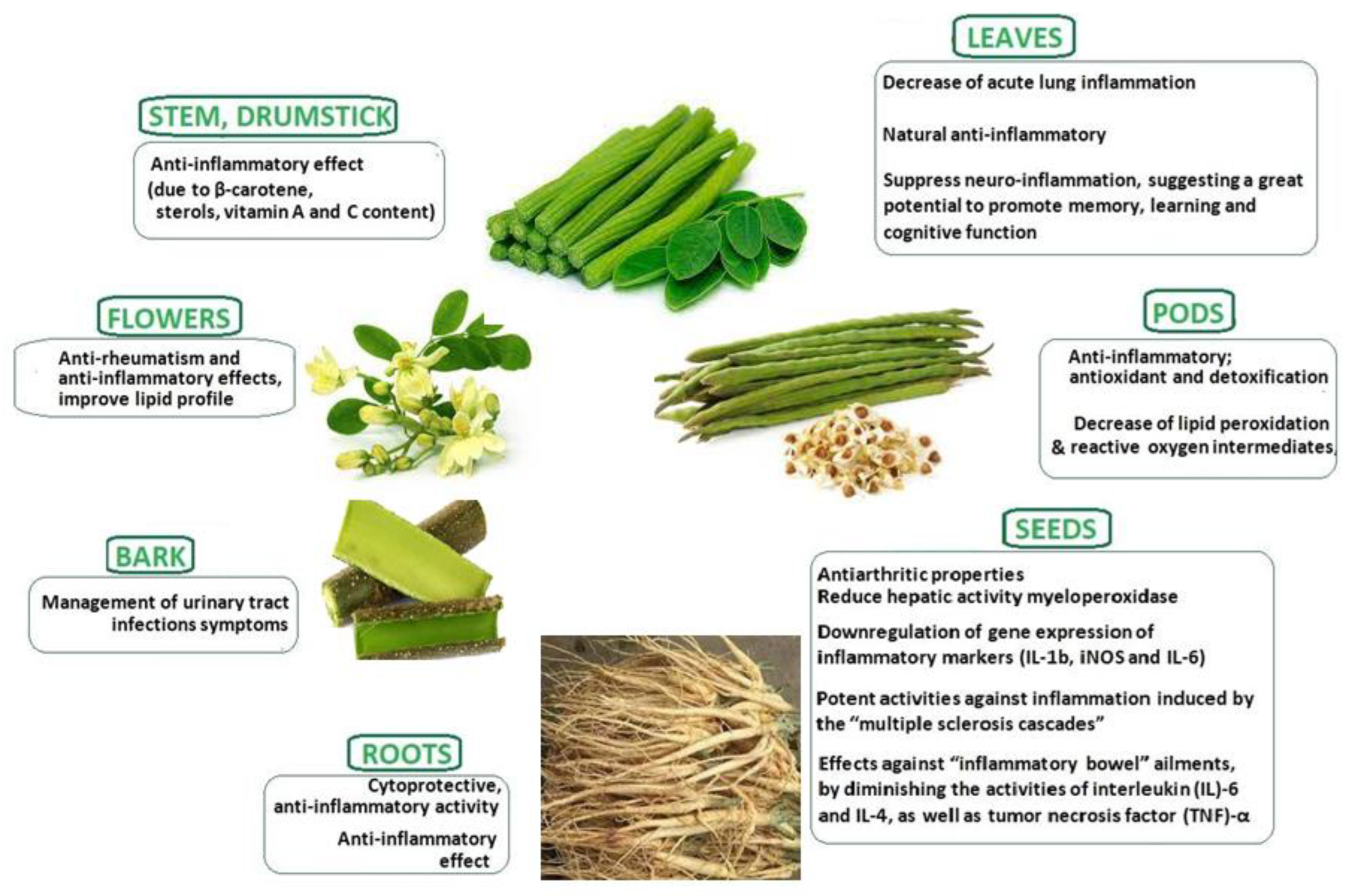{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Part Used/Type of Extract | Experimental Model | Model of Study | Duration of the Treatment | Dosage | Control | Extracts/Secondary Metabolites | Bioactivity/Therapeutic Characteristics | Ref. |
|---|---|---|---|---|---|---|---|---|
| Leaves, fruits, and seeds/aqueous extracts | The experiment investigated the capacity to restrain oxidative DNA damage, antioxidant, and anti-quorum sensing (QS) capabilities | - | 5–10 µg/mL 50 mg/mL | Positive control: furanone and garlic extract | Polyphenolic compounds (aqueous extracts) | Inhibition of proteins and lipid oxidation Antioxidant activities: nitric oxide and superoxide anion scavenging activities | [14] | |
| Leaves | The study screened the functional metabolites of M. oleifera to evaluate their possible role in nerve regeneration after injury | in vivo Male albino mice | 12 days | 2 g/kg bw | Control: standard diet | Leaf extracts in n-Hexane, dichloromethane, ethyl acetate, ethanol, and methanol | Nerve regeneration (restoration of sensory and motor function) Oxidative stress management | [7] |
| Defatted seed flour | The study assessed the antioxidant and antibacterial activities of free and bound phenolic extracts | in vitro B. cereus S. aureus E. coli Y. enterocolitica | - | 0.5–15.0 mg in 150 µL 0.5–1.5 mg/mL | Positive control: ascorbic acid | Phenolic compounds (extracts in ethanol, methanol, acetone, hexane, and chloroform) | Antioxidant, antimicrobial activities | [15] |
| Seeds | The experiments evaluated the antibacterial and antifungal activities of secondary metabolites | in vitro Bacteria: S. aureus S. epidermidis E. coli E. aerogenes K. pneumonia P. aeruginosa B. subtilis Fungal strains: C. albicans T. rubrum E. floccosum | - | 10 mg/mL 1 mg/mL 100 µg/mL 10 µg/mL | Positive control: Ofloxacin and Clotrimazole Negative control: sterile distilled water | Glucosinolates (secondary metabolites extracts in acetone in CH2Cl2) | Antimicrobial activities | [16] |
| Seeds | The study evaluated the antitumoral activity of eight isolate compounds from M. oleifera | in vitro EBV genome carrying lymphoblastoid cell in vivo Specific pathogen-free female ICR mice | - 20 weeks | 100, 10, 1, and 0.1 µg/mL 85 nmol in 0.1 mL acetone | Positive control: n-butyric acid Negative control: 12-O-tetradecanoyl-phorbol-13-acetate (TPA) | Glucosinolates, isothiocyanates, and sterols | Antitumor promoting activities | [17] |
| Seeds | The experiments assessed the antimicrobial activity of a polymeric, naturally extracted M. oleifera oil bionanocomposite film enriched with silver nanoparticles | in vitro S. aureus, E. coli K. pneumoniae S. typhi P. aeruginosa S. flexneri C. albicans | - | 5–10 wt% | Ciprofloxacin and Fluconazole | Fatty acids, sterols, alkanes, and alcohol compounds | Antibacterial activity | [18] |
| Seeds | The study assessed the cytotoxic activity of seed essential oil obtained from M. oleifera | in vitro HeLa, HepG2, MCF-7, CACO-2, and L929 cell lines | 24 h | 0.15 to 1.0 mg/mL | DMSO | Essential oils (extracted from seeds through cold pressing) | Antiproliferative activity | [19] |
| Flowers | PC3 cell lines | 0.01–100 µg/mL | DMSO | Methanol extracts | Anticancer activity | [20] | ||
| Leaves | The study has investigated the anticancer activity of the M. oleifera leaf extract | in vitro murine Non-Hodgkin Lymphoma (NHL) in vivo Balb/c mice | 24 h 16 days | 100 to 450 µg/mL 100 and 200 mg/kg | Healthy mice without Dalton’s lymphoma cell transplantation | Methanol-based leaf extract | Triggers apoptosis and inhibits the growth of Dalton’s lymphoma | [21] |
| Seeds | Experiments assessed the antioxidant, antihypertensive, and potential cardioprotective properties of bioactive peptides | in vivo Wistar rats | - - | - 200 mg/kg | - - | Peptides obtained through enzymatic hydrolysis of M. oleifera seed (shorter peptides (1–3 kDa) and longer peptides (>10 kDa)) | Oxidative stress management; antihypertensive, and cardioprotective properties | [22,23] |
| Seeds | The study compared the structural and functional properties of albumin and globulin in M. oleifera seeds with those of the isoelectric pH-precipitated protein isolate | - | - | 6.25 mg/mL | - | Globulins, Albumins, Iso–electric precipitated isolates | Reduce free radicals Globulin-enhanced metal ion chelation activity Antioxidant properties | [24] |
| Seeds | The experiments compared the antioxidant and angiotensin-converting enzyme (ACE) inhibitory properties of M. oleifera seed protein isolate (ISO) | - | - | - | - | Protein hydrolysate fractions (<1 kDa, 3–5 kDa, and 5–10 kDa) | Antioxidative properties ACE inhibition; | [25] |
| Seeds | The study evaluated the biofunctional properties of total hydrolysates and peptide fractions from protein isolates of moringa seeds | in vitro ACE extracted from rabbit lungs | 5 h | - | - | Protein hydrolysate fractions >10 kDa) | Antioxidant, antihypertensive, and antidiabetic properties; | [26] |
| Seeds | The study evaluated the nutritional composition of Indian M. oleifera seed, the antioxidant activity of its polypeptides, and the protective effects on H2O2 oxidative-damaged Chang liver cells | in vitro Chang liver cell line | 3 h | 100, 300, and 500 µM | PBS | Peptide isolates and hydrolysate Fractions (>3.5 kDa) PFE, GY, YTR, QY, FG, SF, IN, SP, YFE, IY, LY | Oxidative stress management Antioxidant activity Hepatoprotector | [27,28] |
| Seeds | Seed powder was extracted in hexane, petroleum ether, ethyl acetate, or methanol, and the study evaluated the extracts antimicrobial activity | in vitro E. coli P. aeruginosa S. aureus C. cladosporioides P. sclerotigenum | - | - | - | Glucosinolates and isothiocyanates | Flocculating and antimicrobial activities | [29] |
| Seeds | The experiments assessed the antimicrobial activities of isolated compounds from seed extracts of M. oleifera and their synergistic effect through a hybridized complex of organic–inorganic composite materials | in vitro S. aureus E. coli P. aeruginosa C. albicans A. niger | 24 h (37 °C) 48 h (25 °C) 5 days (25 °C) | 5 mg | - | Glucosinolates and isothiocyanates (ethanol, methanol, hexane, acetate ethanol extracts) | Antimicrobial activities | [30] |
| Seeds | The study evaluated the antimicrobial activities of 4-(α-l-rhamnosyloxy) benzyl glucosinolate isolated from M. oleifera seed and its protective effect on an experimental model of spinal cord injury | in vitro S. aureus E. casseliflavus C. albicans in vivo Male adult C57Bl/6 mice | 24 h 8 days | - 10 mg/kg | PBS Gentamicin Chloramphenicol Negative control: naive group (no treatment) Positive control: GMG-ITC (control group) | Glucosinolates and isothiocyanates | Antibiotic activity Nerve regeneration | [31,32,33] |
| Leaves | The study evaluated the anxiolytic and anti-colitis effects of M. oleifera leaf-aqueous extract on acetic acid-induced colon inflammation in rat | in vivo Adult albino Wistar rats | 20 days | 25, 50, and 100 mg/kg | Loperamide | - | Anxiolytic, anti-inflammatory Antioxidant and anti-colitis properties | [3] |
| Seeds | The study evaluated the dietary isothiocyanate-enriched moringa seed extract on glucose tolerance in a high fat diet mouse model and its modulatory activity on the gut microbiome | in vivo Male Sprague–Dawley rats | 12 weeks | 0.54 and 0.73% of moringa seed extract containing moringa isothiocyanate-1 | Vehicle control: 15% sodium carboxymethyl cellulose | Glucosinolates, phenolic glycosides, flavonoids, and carbohydrates | Anti-inflammatory Antioxidant properties Improves glucose tolerance and modulates the gut microbiome | [34,35] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chiș, A.; Noubissi, P.A.; Pop, O.-L.; Mureșan, C.I.; Fokam Tagne, M.A.; Kamgang, R.; Fodor, A.; Sitar-Tăut, A.-V.; Cozma, A.; Orășan, O.H.; et al. Bioactive Compounds in Moringa oleifera: Mechanisms of Action, Focus on Their Anti-Inflammatory Properties. Plants 2024, 13, 20. https://doi.org/10.3390/plants13010020
Chiș A, Noubissi PA, Pop O-L, Mureșan CI, Fokam Tagne MA, Kamgang R, Fodor A, Sitar-Tăut A-V, Cozma A, Orășan OH, et al. Bioactive Compounds in Moringa oleifera: Mechanisms of Action, Focus on Their Anti-Inflammatory Properties. Plants. 2024; 13(1):20. https://doi.org/10.3390/plants13010020
Chicago/Turabian StyleChiș, Adina, Paul Aimé Noubissi, Oana-Lelia Pop, Carmen Ioana Mureșan, Michel Archange Fokam Tagne, René Kamgang, Adriana Fodor, Adela-Viviana Sitar-Tăut, Angela Cozma, Olga Hilda Orășan, and et al. 2024. "Bioactive Compounds in Moringa oleifera: Mechanisms of Action, Focus on Their Anti-Inflammatory Properties" Plants 13, no. 1: 20. https://doi.org/10.3390/plants13010020