
Linking Seed Traits and Germination Responses in Caribbean Seasonally Dry Tropical Forest Species

Abstract
:1. Introduction
2. Results
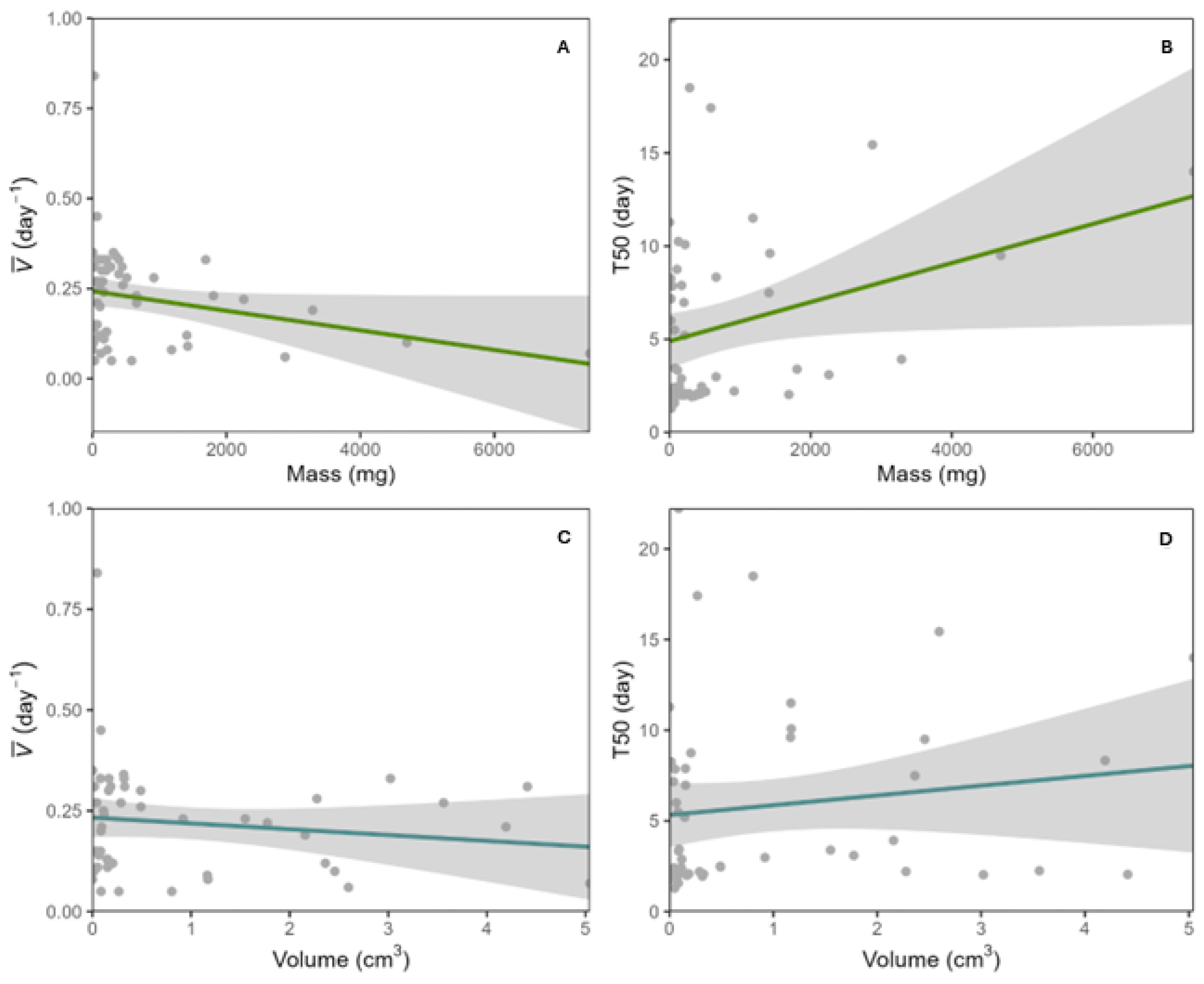
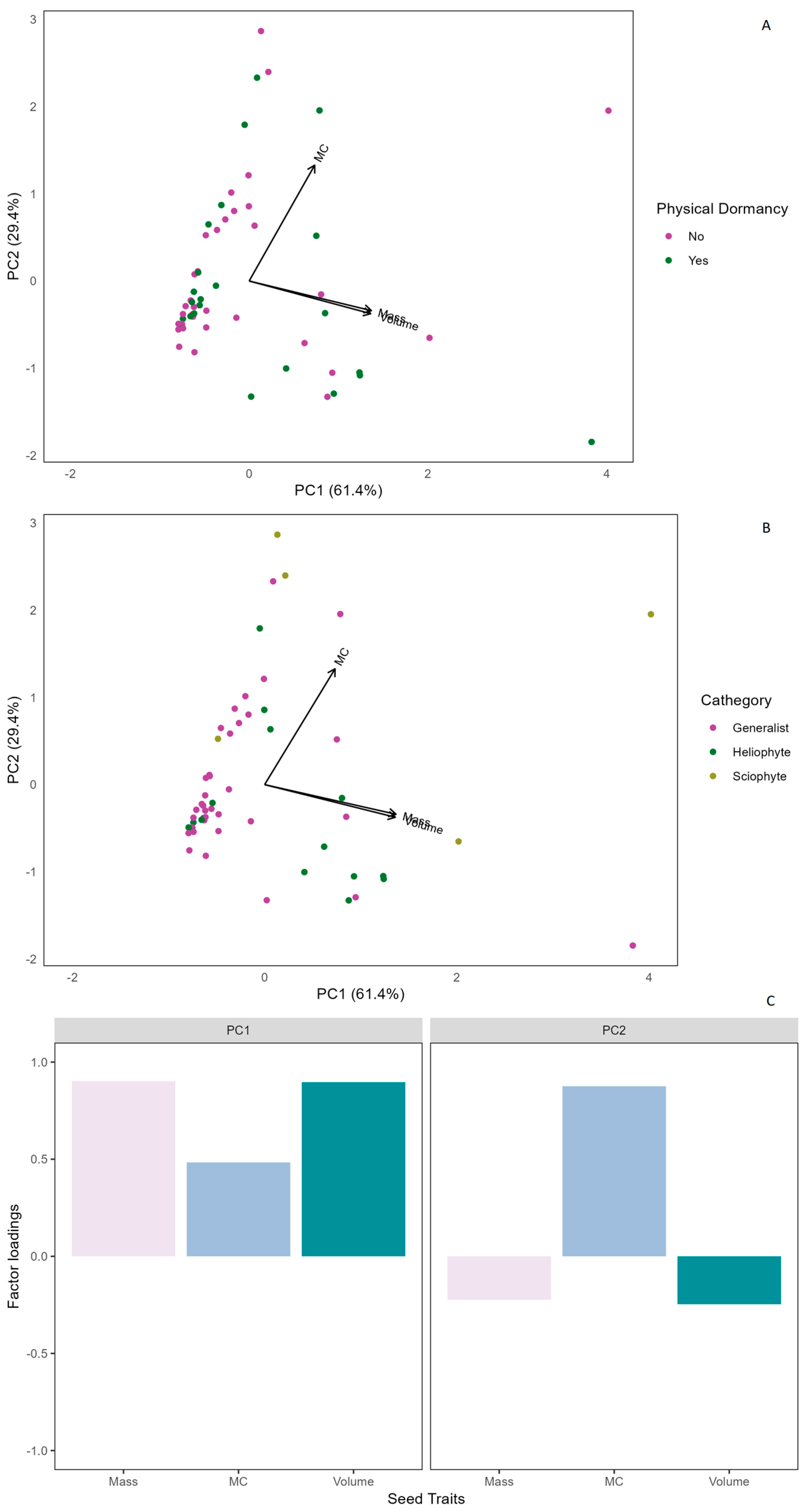
2.1. Seed Traits and Germination Responses in SDTF Species
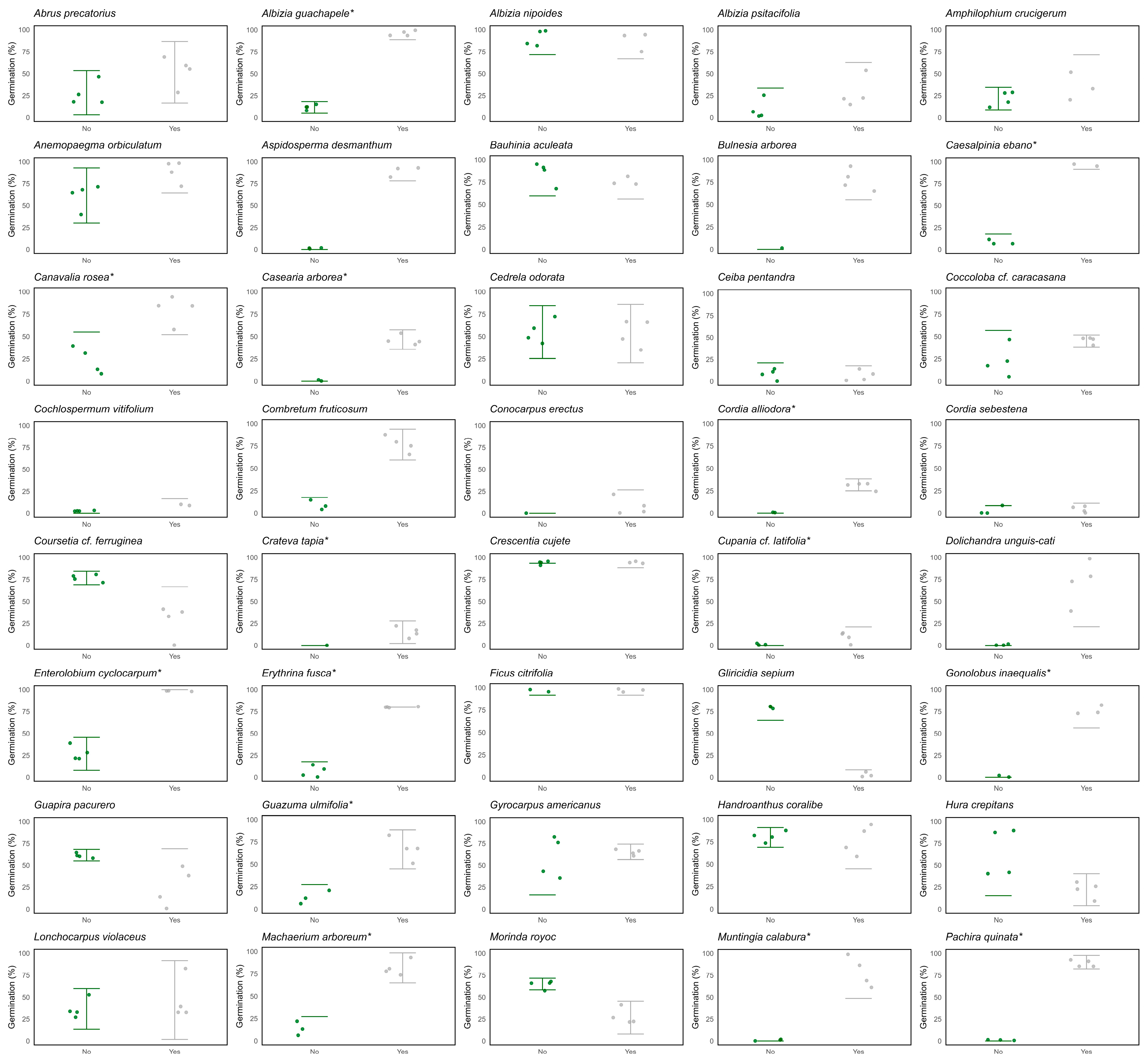
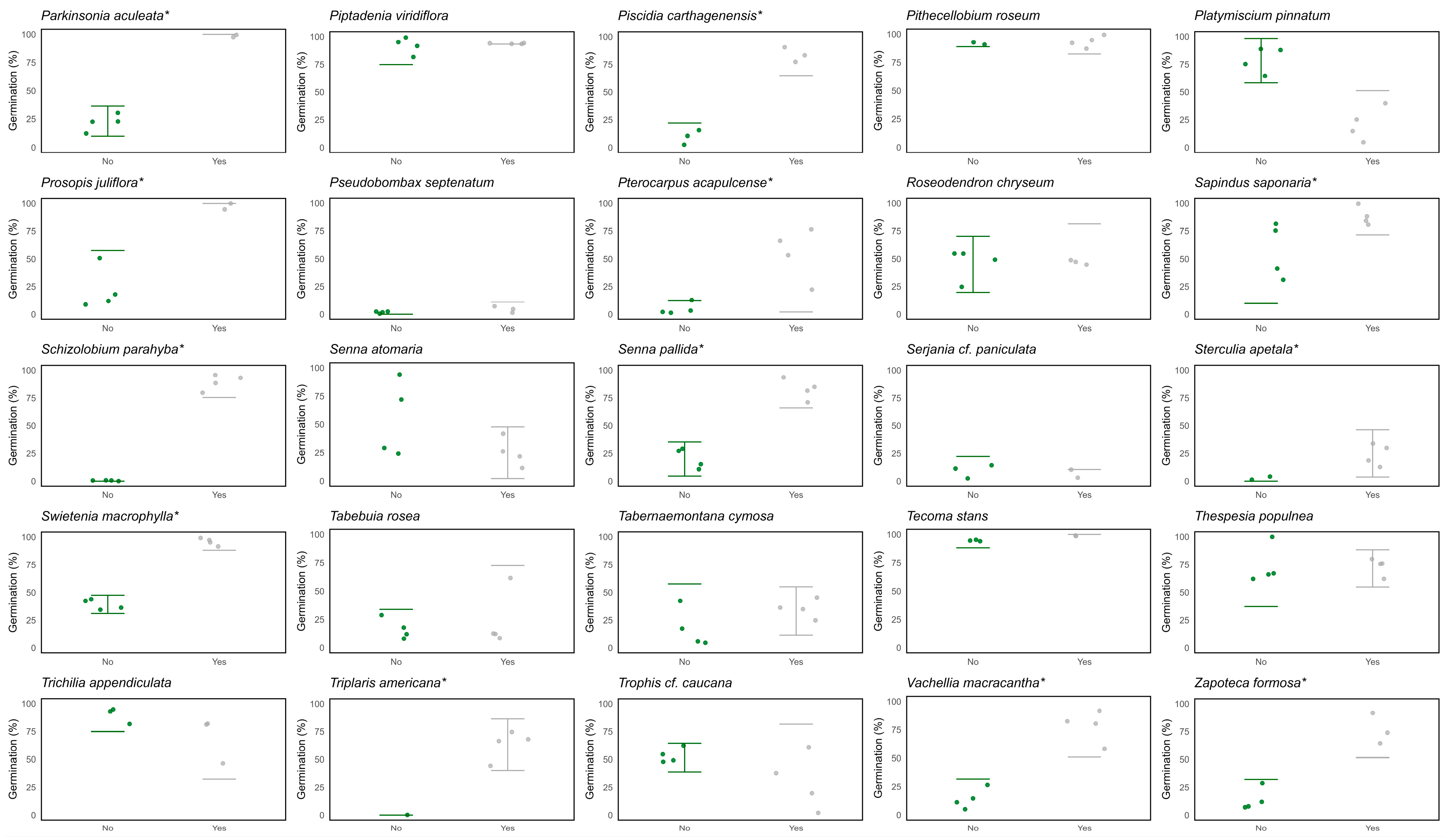
2.2. Seed Physical Dormancy in SDTF Species
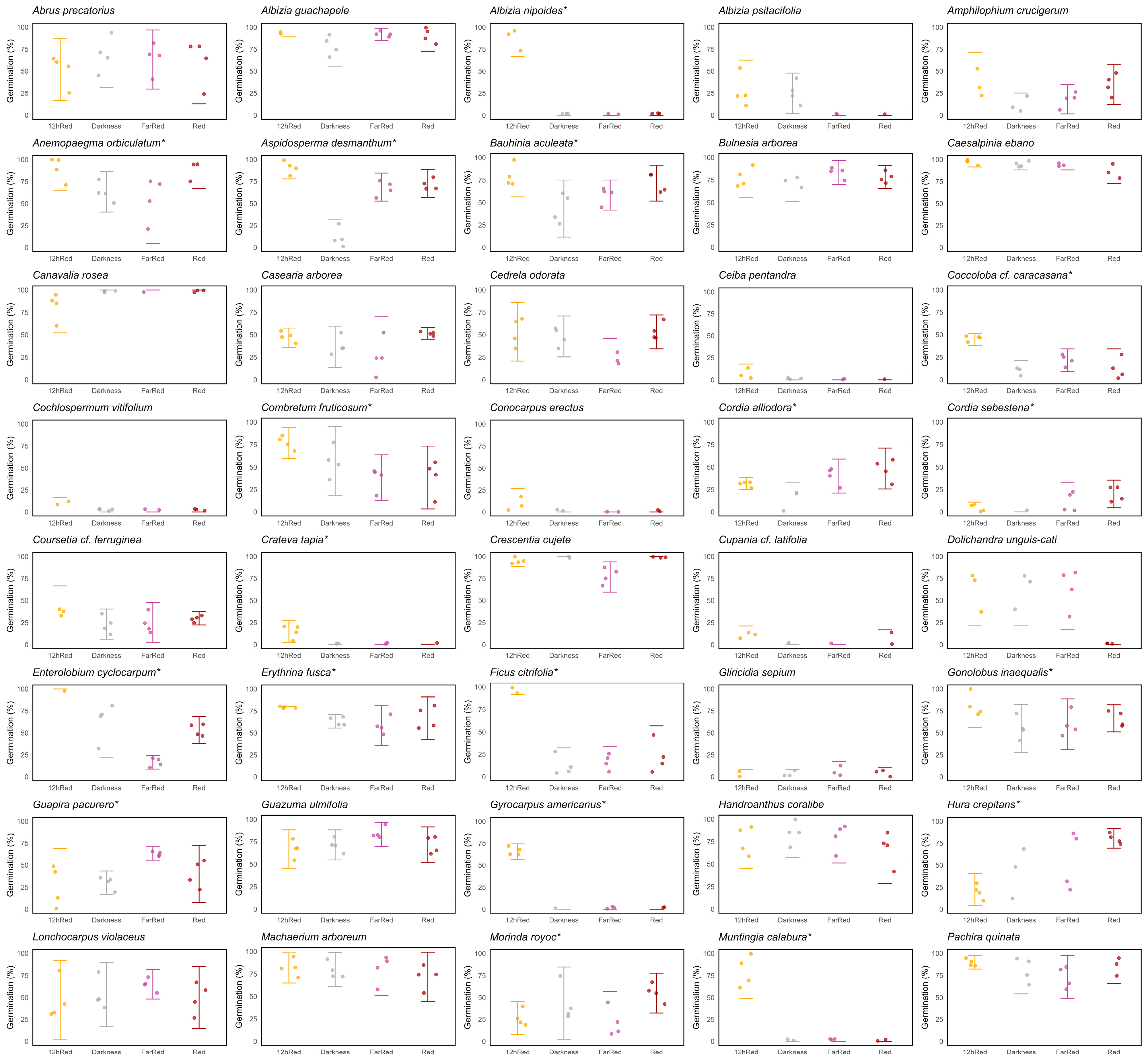
2.3. Germination Responses to Different Light Qualities in SDTF Species
3. Discussion
4. Materials and Methods
4.1. Study Site
4.2. Seed Collection
4.3. Germination Response
4.4. Seed Traits
4.5. Data Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. Physical Dormancy Species-Level Results
References
- Hanbury-Brown, A.R.; Ward, R.E.; Kueppers, L.M. Forest Regeneration within Earth System Models: Current Process Representations and Ways Forward. New Phytol. 2022, 235, 20–40. [Google Scholar] [CrossRef] [PubMed]
- Walck, J.L.; Hidayati, S.N.; Dixon, K.W.; Thompson, K.; Poschlod, P. Climate Change and Plant Regeneration from Seed. Glob. Change Biol. 2011, 17, 2145–2161. [Google Scholar] [CrossRef]
- Donohue, K.; Rubio de Casas, R.; Burghardt, L.; Kovach, K.; Willis, C.G. Germination, Postgermination Adaptation, and Species Ecological Ranges. Annu. Rev. Ecol. Evol. Syst. 2010, 41, 293–319. [Google Scholar] [CrossRef]
- Grubb, P.J. The Maintenance of Species-Richness in Plant Communities: The Importance of the Regeneration Niche. Biol. Rev. 1977, 52, 107–145. [Google Scholar] [CrossRef]
- Finch-Savage, W.E.; Leubner-Metzger, G. Seed Dormancy and the Control of Germination. New Phytol. 2006, 171, 501–523. [Google Scholar] [CrossRef] [PubMed]
- Escobar, D.F.E.; Rubio de Casas, R.; Morellato, L.P.C. Many Roads to Success: Different Combinations of Life-History Traits Provide Accurate Germination Timing in Seasonally Dry Environments. Oikos 2021, 130, 1865–1879. [Google Scholar] [CrossRef]
- Miles, L.; Newton, A.C.; DeFries, R.S.; Ravilious, C.; May, I.; Blyth, S.; Kapos, V.; Gordon, J.E. A Global Overview of the Conservation Status of Tropical Dry Forests. J. Biogeogr. 2006, 33, 491–505. [Google Scholar] [CrossRef]
- Portillo-Quintero, C.A.; Sánchez-Azofeifa, G.A. Extent and Conservation of Tropical Dry Forests in the Americas. Biol. Conserv. 2010, 143, 144–155. [Google Scholar] [CrossRef]
- Dimson, M.; Gillespie, T.W. Trends in Active Restoration of Tropical Dry Forest: Methods, Metrics, and Outcomes. For. Ecol. Manag. 2020, 467, 118150. [Google Scholar] [CrossRef]
- Fremout, T.; Gutiérrez-Miranda, C.E.; Briers, S.; Marcelo-Peña, J.L.; Cueva-Ortiz, E.; Linares-Palomino, R.; La Torre-Cuadros, M.d.l.Á.; Chang-Ruíz, J.C.; Villegas-Gómez, T.L.; Acosta-Flota, A.H.; et al. The Value of Local Ecological Knowledge to Guide Tree Species Selection in Tropical Dry Forest Restoration. Restor. Ecol. 2021, 29, e13347. [Google Scholar] [CrossRef]
- Dryflor; Banda-R, K.; Delgado-Salinas, A.; Dexter, K.G.; Linares-Palomino, R.; Oliveira-Filho, A.; Prado, D.; Pullan, M.; Quintana, C.; Pennington, R.T.; et al. Plant Diversity Patterns in Neotropical Dry Forests and Their Conservation Implications. Science 2016, 353, 1383–1387. [Google Scholar] [CrossRef] [PubMed]
- Gentry, A.H. Diversity and Floristic Composition of Neotropical Dry Forests. In Seasonally Dry Tropical Forests; Bullock, S.H., Mooney, H.A., Medina, E., Eds.; Cambridge University Press: New York, NY, USA, 1995; pp. 146–194. ISBN 9780511753398. [Google Scholar]
- Murphy, P.G.; Lugo, A.E. Ecology of Tropical Dry Forest. Annu. Rev. Ecol. Syst. 1986, 17, 67–88. [Google Scholar] [CrossRef]
- Balvanera, P.; Lott, E.; Segura, G.; Siebe, C.; Islas, A. Patterns of β-Diversity in a Mexican Tropical Dry Forest. J. Veg. Sci. 2002, 13, 145–158. [Google Scholar] [CrossRef]
- Reich, P.B.; Borchert, R. Water Stress and Tree Phenology in a Tropical Dry Forest in the Lowlands of Costa Rica. J. Ecol. 1984, 72, 61–74. [Google Scholar] [CrossRef]
- Pizano, C.; García, H. El Bosque Seco Tropical En Colombia, 1st ed.; Pizano, C., García, H., Eds.; Instituto de Investigación de Recursos Biológicos Alexander von Humboldt (IAvH): Bogotá, Colombia, 2014; ISBN 978-958-8343-97-6. [Google Scholar]
- Ranal, M.A.; de Santana, D.G. How and Why to Measure the Germination Process? Rev. Bras. De Botânica 2006, 29, 1–11. [Google Scholar] [CrossRef]
- Sales, N.M.; Pérez-García, F.; Silveira, F.A.O. Consistent Variation in Seed Germination across an Environmental Gradient in a Neotropical Savanna. S. Afr. J. Bot. 2013, 87, 129–133. [Google Scholar] [CrossRef]
- Blakesley, D.; Elliott, S.; Kuarak, C.; Navakitbumrung, P.; Zangkum, S.; Anusarnsunthorn, V. Propagating Framework Tree Species to Restore Seasonally Dry Tropical Forest: Implications of Seasonal Seed Dispersal and Dormancy. For. Ecol. Manag. 2002, 164, 31–38. [Google Scholar] [CrossRef]
- Bedoya-Patiño, J.G.; Estévez-Varón, J.V.; Castaño-Villa, G.J. Banco de Semillas Del Suelo y Su Papel En La Recuperación de Los Bosques Tropicales. Boletín Científico Cent. Museos. Mus. Hist. Nat. 2010, 14, 77–91. [Google Scholar]
- Baskin, J.M.; Baskin, C.C. The Great Diversity in Kinds of Seed Dormancy: A Revision of the Nikolaeva-Baskin Classification System for Primary Seed Dormancy. Seed Sci. Res. 2021, 31, 249–277. [Google Scholar] [CrossRef]
- Ibarra-Manríquez, G.; Cortés-Flores, J.; Sánchez-Coronado, M.E.; Soriano, D.; Reyes-Ortega, I.; Orozco-Segovia, A.; Baskin, C.C.; Baskin, J.M. Climate Change and Plant Regeneration from Seeds in Tropical Dry Forests. In Plant Regeneration from Seeds: A Global Warming Perspective; Elsevier: Amsterdam, The Netherlands, 2022; pp. 169–181. ISBN 9780128237311. [Google Scholar]
- Baskin, C.; Baskin, J. Seeds: Ecology, Biogeography, and Evolution of Dormancy and Germination; Baskin, C., Baskin, J., Eds.; Elsevier: Lexington, KY, USA, 1998; ISBN 9788578110796. [Google Scholar]
- Saatkamp, A.; Cochrane, A.; Commander, L.; Guja, L.K.; Jimenez-Alfaro, B.; Larson, J.; Nicotra, A.; Poschlod, P.; Silveira, F.A.O.; Cross, A.T.; et al. A Research Agenda for Seed-Trait Functional Ecology. New Phytol. 2019, 221, 1764–1775. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Alfaro, B.; Silveira, F.A.O.; Fidelis, A.; Poschlod, P.; Commander, L.E. Seed Germination Traits Can Contribute Better to Plant Community Ecology. J. Veg. Sci. 2016, 27, 637–645. [Google Scholar] [CrossRef]
- Khurana, E.; Singh, J.S. Ecology of Tree Seed and Seedlings: Implications for Tropical Forest Conservation and Restoration. Curr. Sci. 2001, 80, 748–757. [Google Scholar] [CrossRef]
- Buoro, M.; Carlson, S.M. Life-History Syndromes: Integrating Dispersal through Space and Time. Ecol. Lett. 2014, 17, 756–767. [Google Scholar] [CrossRef] [PubMed]
- Moles, A.T.; Ackerly, D.D.; Tweddle, J.C.; Dickie, J.B.; Smith, R.; Leishman, M.R.; Mayfield, M.M.; Pitman, A.; Wood, J.T.; Westoby, M. Global Patterns in Seed Size. Glob. Ecol. Biogeogr. 2006, 16, 109–116. [Google Scholar] [CrossRef]
- Moles, A.T.; Westoby, M. Seed Size and Plant Strategy across the Whole Life Cycle. Oikos 2006, 113, 91–105. [Google Scholar] [CrossRef]
- Norden, N.; Daws, M.I.; Antoine, C.; Gonzalez, M.A.; Garwood, N.C.; Chave, J. The Relationship between Seed Mass and Mean Time to Germination for 1037 Tree Species across Five Tropical Forests. Funct. Ecol. 2009, 23, 203–210. [Google Scholar] [CrossRef]
- Adams, C.A.; Rinne, R.W. Moisture Content as a Controlling Factor in Seed Development and Germination. Int. Rev. Cytol. 1980, 68, 1–8. [Google Scholar] [CrossRef]
- Wendt, A.; Chazdon, R.L.; Vargas Ramirez, O. Successional Trajectories of Seed Dispersal Mode and Seed Size of Canopy Tree Species in Wet Tropical Forests. Front. For. Glob. Change 2022, 5, 946541. [Google Scholar] [CrossRef]
- Pearson, T.R.H.; Burslem, D.F.R.P.; Mullins, C.E.; Dalling, J.W. Germination Ecology of Neotropical Pioneers: Interacting Effects of Environmental Conditions and Seed Size. Ecology 2002, 83, 2798–2807. [Google Scholar] [CrossRef]
- Fenner, M.; Thompson, K. The Ecology of Seeds, 1st ed.; Fenner, M., Thompson, K., Eds.; Cambridge University Press: Cambridge, UK, 2005; ISBN 9780521653114. [Google Scholar]
- Khurana, E.; Singh, J.S. Germination and Seedling Growth of Five Tree Species from Tropical Dry Forest in Relation to Water Stress: Impact of Seed Size. J. Trop. Ecol. 2004, 20, 385–396. [Google Scholar] [CrossRef]
- Augspurger, C.K. Light Requirements of Neotropical Tree Seedlings: A Comparative Study of Growth and Survival. J. Ecol. 1984, 72, 777–795. [Google Scholar] [CrossRef]
- Escobar Escobar, D.F.; Cardoso, V.J.M. Germinación y Latencia de Semillas de Miconia Chartacea (Melastomataceae), En Respuesta a Luz, Temperatura y Hormonas Vegetales. Rev. De Biol. Trop. 2015, 63, 1169–1184. [Google Scholar] [CrossRef]
- Vargas-Figueroa, J.A.; Duque-Palacio, O.L.; Torres-González, A.M. Germinación de Semillas de Cuatro Especies Arbóreas Del Bosque Seco Tropical Del Valle Del Cauca, Colombia. Rev. Biol. Trop. 2015, 63, 249–261. [Google Scholar] [CrossRef] [PubMed]
- Cortés-Flores, J.; Hernández-Esquivel, K.; González-Rodríguez, A.; Ibarra-Manríquez, G. Flowering Phenology, Growth Forms, and Pollination Syndromes in Tropical Dry Forest Species: Influence of Phylogeny and Abiotic Factors. Am. J. Bot. 2017, 104, 39–49. [Google Scholar] [CrossRef]
- Cortés Flores, J.; Cornejo-Tenorio, G.; Sánchez-Coronado, M.E.; Orozco-Segovia, A.; Ibarra-Manríquez, G. Disentangling the Influence of Ecological and Historical Factors on Seed Germination and Seedling Types in a Neotropical Dry Forest. PLoS ONE 2020, 15, e0231526. [Google Scholar] [CrossRef] [PubMed]
- Bu, H.; Ge, W.; Zhou, X.; Qi, W.; Liu, K.; Xu, D.; Wang, X.; Du, G. The Effect of Light and Seed Mass on Seed Germination of Common Herbaceous Species from the Eastern Qinghai-Tibet Plateau. Plant Species Biol. 2017, 32, 263–269. [Google Scholar] [CrossRef]
- Hammond, D.S.; Brown, V.K. Seed Size of Woody Plants in Relation to Disturbance, Dispersal, Soil Type in Wet Neotropical Forests. Ecology 1995, 76, 2544–2561. [Google Scholar] [CrossRef]
- Pérez-Harguindeguy, N.; Diaz, S.; Garnier, E.; Lavorel, S.; Poorter, H.; Jaureguiberry, P.; Bret-Harte, M.S.S.; Cornwell, W.K.K.; Craine, J.M.M.; Gurvich, D.E.E.; et al. New Handbook for Standardized Measurment of Plant Functional Traits Worldwide. Aust. J. Bot. 2013, 61, 167–234. [Google Scholar] [CrossRef]
- Oliveira, L.M.; Valio, I.F.M. Effects of Moisture Content on Germination of Seeds of Hancornia speciosa Gom. (Apocynaceae). Ann. Bot. 1992, 69, 1–5. [Google Scholar] [CrossRef]
- Jara-Guerrero, A.; Espinosa, C.I.; Méndez, M.; De la Cruz, M.; Escudero, A. Dispersal Syndrome Influences the Match between Seed Rain and Soil Seed Bank of Woody Species in a Neotropical Dry Forest. J. Veg. Sci. 2020, 31, 995–1005. [Google Scholar] [CrossRef]
- Romero-Saritama, J.M.; Pérez Ruiz, C. Seed Morphological Traits and Their Implication in the Ex Situ Conservation of Woody Species in Tumbesian Dry Forests. Ecosistemas 2016, 25, 59–65. [Google Scholar] [CrossRef]
- Daws, M.I.; Garwood, N.C.; Pritchard, H.W. Traits of Recalcitrant Seeds in a Semi-Deciduous Tropical Forest in Panamá: Some Ecological Implications. Funct. Ecol. 2005, 19, 874–885. [Google Scholar] [CrossRef]
- Garwood, N.C. Seed Germination in a Seasonal Tropical Forest in Panama: A Community Study. Ecol. Monogr. 1983, 53, 159–181. [Google Scholar] [CrossRef]
- Hu, X.W.; Wang, Y.R.; Wu, Y.P.; Baskin, C.C. Role of the Lens in Controlling Water Uptake in Seeds of Two Fabaceae (Papilionoideae) Species Treated with Sulphuric Acid and Hot Water. Seed Sci. Res. 2009, 19, 73–80. [Google Scholar] [CrossRef]
- Jaganathan, G.K.; Wu, G.R.; Han, Y.Y.; Liu, B.L. Role of the Lens in Controlling Physical Dormancy Break and Germination of Delonix regia (Fabaceae: Caesalpinioideae). Plant Biol. 2017, 19, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Baskin, J.M.; Baskin, C.C.; Li, X. Taxonomy, Anatomy and Evolution of Physical Dormancy in Seeds. Plant Species Biol. 2000, 15, 139–152. [Google Scholar] [CrossRef]
- Dechaine, J.M.; Gardner, G.; Weinig, C. Phytochromes Differentially Regulate Seed Germination Responses to Light Quality and Temperature Cues during Seed Maturation. Plant Cell Environ. 2009, 32, 1297–1309. [Google Scholar] [CrossRef] [PubMed]
- Vieira, D.L.M.; Lima, V.V.d.; Sevilha, A.C.; Scariot, A. Consequences of Dry-Season Seed Dispersal on Seedling Establishment of Dry Forest Trees: Should We Store Seeds until the Rains? For. Ecol. Manag. 2008, 256, 471–481. [Google Scholar] [CrossRef]
- Milberg, P.; Andersson, L.; Thompson, K. Large-Seeded Species Are Less Dependent on Light for Germination than Small-Seeded Ones. Seed Sci. Res. 2000, 10, 99–104. [Google Scholar] [CrossRef]
- Jankowska-Blaszczuk, M.; Daws, M.I. Impact of Red: Far Red Ratios on Germination of Temperate Forest Herbs in Relation to Shade Tolerance, Seed Mass and Persistence in the Soil. Funct. Ecol. 2007, 21, 1055–1062. [Google Scholar] [CrossRef]
- Agudelo-Ramirez, C.P. Estructura de Los Bosques de Manglar Del Departamento de Bolivar y Su Relación Con Algunos Parámetros Abióticos; Tesis Pregrado, Universidad Jorge Tadeo Lozano: Bogotá, Colombia, 2000. [Google Scholar]
- Di Sacco, A.; Way, M.; León Lobos, P.; Suárez Ballesteros, C.I.; Rodriguez Díaz, J.V. Manual de Recolección, Procesamiento, y Conservación de Semillas de Plantas Silvestres; Royal Botanic Gardens Kew: Richmond, UK; Instituto de Investigación de Recursos Biológicos Alexander von Humboldt: Bogotá, Colombia, 2020; ISBN 9789585418936. [Google Scholar]
- Coolbear, P.; Francis, A.; Grierson, D. The Effect of Low Temperature Pre-Sowing Treatment on the Germination Performance and Membrane Integrity of Artificially Aged Tomato Seeds. J. Exp. Bot. 1984, 35, 1609–1617. [Google Scholar] [CrossRef]
- ISTA Changes to the ISTA Rules for 2017. Int. Rules Seed Test. Zur. Switz. 2017, 12, 345. [CrossRef]
- Society for Ecological Restoration (SER). Seed Information Database (SID). Available online: https://ser-sid.org/ (accessed on 1 July 2023).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2023. [Google Scholar]
- Lüdecke, D.; Ben-Shachar, M.; Patil, I.; Waggoner, P.; Makowski, D. Performance: An R Package for Assessment, Comparison and Testing of Statistical Models. J. Open Source Softw. 2021, 6, 3139. [Google Scholar] [CrossRef]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Wickham, H.; Averick, M.; Bryan, J.; Chang, W.; McGowan, L.; François, R.; Grolemund, G.; Hayes, A.; Henry, L.; Hester, J.; et al. Welcome to the Tidyverse. J. Open Source Softw. 2019, 4, 1686. [Google Scholar] [CrossRef]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using Lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Habit | Dispersal | Ecological Category | Photoblastic | PY | Mass (mg) | Volume (cm3) | Moisture Content (%) |
|---|---|---|---|---|---|---|---|---|
| APOCYNACEAE | ||||||||
| Aspidosperma desmanthum | Tree | Abiotic | Heliophyte | Yes | No | 1409.3 ± 251.8 | 2.3619 ± 0.6857 | 18.6 ± 14.6 |
| Gonolobus inaequalis | Liana | Abiotic | Heliophyte | Yes | Yes | 69.7 ± 14.7 | 0.085 ± 0.0178 | 7 ± 0.2 |
| Tabernaemontana cymosa | Tree | Biotic | Heliophyte | Yes | No | 284.7 ± 56.6 | 0.805 ± 0.1512 | 23.8 ± 1.2 |
| BIGNONIACEAE | ||||||||
| Amphilophium crucigerum | Liana | Abiotic | Generalist | No | No | 213.6 ± 28.8 | 0.1463 ± 0.0178 | 8.8 ± 0 |
| Anemopaegma orbiculatum | Liana | Abiotic | Heliophyte | Yes | No | 171.7 ± 39.7 | 0.1526 ± 0.0167 | 9 ± 0 |
| Crescentia cujete | Tree | Biotic | Generalist | No | No | 62.2 ± 11.5 | 0.0431 ± 0.0094 | 7.4 ± 0.1 |
| Dolichandra unguis-cati | Liana | Abiotic | Generalist | No | No | 124.9 ± 16.3 | --- | 7 ± 0.1 |
| Handroanthus coralibe | Tree | Abiotic | Generalist | No | No | 24.3 ± 7.2 | 0.0486 ± 0.0077 | 9.5 ± 0.2 |
| Roseodendron chryseum | Tree | Abiotic | Generalist | No | No | 20.9 ± 4.9 | 0.0382 ± 0.0063 | 8.5 ± 0.4 |
| Senna atomaria | Tree | Abiotic | Sciophyte | Yes | No | 63.4 ± 8.2 | 0.0418 ± 0.005 | 18.2 ± 0.4 |
| Tabebuia rosea | Tree | Abiotic | Generalist | No | No | 75.5 ± 14.8 | 0.0928 ± 0.0224 | 7 ± 2.1 |
| Tecoma stans | Tree | Abiotic | Generalist | No | No | 26.5 ± 4 | 0.0213 ± 0.0034 | 6.6 ± 0 |
| BIXACEAE | ||||||||
| Cochlospermum vitifolium | Tree | Abiotic | Generalist | No | No | 93.7 ± 9.3 | 0.0799 ± 0.0082 | 10.4 ± 0 |
| BORAGINACEAE | ||||||||
| Cordia alliodora | Tree | Abiotic | Heliophyte | Yes | Yes | 40.8 ± 8.5 | 0.0524 ± 0.0062 | 8 ± 0.1 |
| Cordia sebestena | Tree | Biotic | Sciophyte | Yes | No | 4912.9 ± 1125.5 | 5.5211 ± 1.2354 | 54.6 ± 5.7 |
| CAPPARACEAE | ||||||||
| Crateva tapia | Tree | Biotic | Heliophyte | Yes | Yes | 95.8 ± 10.8 | 0.2675 ± 0.0609 | 32.2 ± 0 |
| COMBRETACEAE | ||||||||
| Combretum fruticosum | Liana | Abiotic | Heliophyte | Yes | No | 238.5 ± 38.3 | 4.4112 ± 0.8861 | 11.2 ± 0.1 |
| Conocarpus erectus | Tree | Abiotic | Generalist | No | No | 18.5 ± 5.4 | 0.0645 ± 0.0101 | 13.4 ± 0.9 |
| EUPHORBIACEAE | ||||||||
| Hura crepitans | Tree | Abiotic | Sciophyte | Yes | No | 4694.2 ± 646.1 | 2.4583 ± 0.2437 | 19.9 ± 1.3 |
| FABACEAE | ||||||||
| Abrus precatorius | Liana | Abiotic | Generalist | No | No | 313.2 ± 68.5 | --- | 9.5 ± 0 |
| Albizia guachapele | Tree | Abiotic | Generalist | No | Yes | 173.2 ± 30.4 | --- | 10.3 ± 0.5 |
| Albizia nipoides | Tree | Abiotic | Heliophyte | Yes | No | 93.8 ± 12.5 | 3.5611 ± 7 | 12.7 ± 0.2 |
| Albizia psitacifolia | Tree | Abiotic | Generalist | No | No | 395.7 ± 57.7 | --- | 14.4 ± 0.7 |
| Bauhinia aculeata | Liana | Abiotic | Heliophyte | Yes | No | 512.5 ± 114 | 0.491 ± 0.0479 | 11.1 ± 0.6 |
| Caesalpinia ebano | Tree | Abiotic | Generalist | No | Yes | 275.2 ± 56.9 | 0.1855 ± 0.04 | 10.4 ± 0.6 |
| Canavalia rosea | Liana | Abiotic | Generalist | No | Yes | 2258 ± 320.3 | 1.7744 ± 0.2724 | 16.7 ± 1.1 |
| Coursetia cf. ferruginea | Tree | Abiotic | Generalist | No | No | 71.4 ± 9.3 | 0.0657 ± 0.0071 | 7.1 ± 0.1 |
| Enterolobium cyclocarpum | Tree | Abiotic | Heliophyte | Yes | Yes | 3287.9 ± 751.8 | 2.1571 ± 0.6087 | 12.3 ± 0.3 |
| Erythrina fusca | Tree | Abiotic | Heliophyte | Yes | Yes | 1806.7 ± 326.7 | 1.5496 ± 0.4203 | 8.6 ± 0.3 |
| Gliricidia sepium | Tree | Abiotic | Generalist | No | No | 462.7 ± 32.6 | 0.3485 ± 0.0399 | 8.4 ± 0.4 |
| Lonchocarpus violaceus | Tree | Abiotic | Generalist | No | No | 362.8 ± 47.8 | 0.3265 ± 0.0767 | 10.2 ± 0.9 |
| Machaerium arboreum | Tree | Abiotic | Generalist | No | Yes | 917 ± 257.1 | 2.2774 ± 0.7306 | 24.6 ± 22.3 |
| Parkinsonia aculeata | Tree | Abiotic | Generalist | No | Yes | 443.7 ± 78.5 | 0.327 ± 0.0381 | 13.4 ± 0.1 |
| Piptadenia viridiflora | Tree | Abiotic | Generalist | No | No | 213.7 ± 30.8 | 0.1673 ± 0.0229 | 19.4 ± 0.4 |
| Piscidia carthagenensis | Tree | Abiotic | Generalist | No | Yes | 204.2 ± 19.6 | 0.1639 ± 0.0119 | 9.2 ± 0.2 |
| Pithecellobium roseum | Tree | Biotic | Generalist | No | No | 396.3 ± 111.1 | 0.321 ± 0.0898 | 22.4 ± 0.5 |
| Platymiscium pinnatum | Tree | Abiotic | Generalist | No | No | 350.4 ± 71.6 | 0.3171 ± 0.0693 | 5.1 ± 0.1 |
| Prosopis juliflora | Tree | Biotic | Generalist | No | Yes | 115.3 ± 17.1 | 0.0811 ± 0.0103 | 11.5 ± 0.2 |
| Pterocarpus acapulcense | Tree | Abiotic | Generalist | No | Yes | 660.1 ± 206.3 | 4.1939 ± 0.5718 | 9 ± 0.5 |
| Schizolobium parahyba | Tree | Abiotic | Heliophyte | Yes | Yes | 168.3 ± 34.5 | 0.1183 ± 0.0237 | 8.7 ± 0.1 |
| Senna pallida | Tree | Abiotic | Generalist | No | Yes | 95.1 ± 16.6 | 0.0054 ± 0.0013 | 19.5 ± 1.2 |
| Vachellia macracantha | Tree | Abiotic | Generalist | No | Yes | 175.4 ± 43.4 | 0.1385 ± 0.0366 | 22.3 ± 5.5 |
| Zapoteca formosa | Tree | Abiotic | Generalist | No | Yes | 92.9 ± 18.7 | 0.0786 ± 0.0167 | 13.8 ± 0.6 |
| HERNANDIACEAE | ||||||||
| Gyrocarpus americanus | Tree | Abiotic | Heliophyte | Yes | No | 1691 ± 394.3 | 3.0223 ± 0.5012 | 8.1 ± 0.5 |
| MALVACEAE | ||||||||
| Ceiba pentandra | Tree | Abiotic | Generalist | No | No | 105.1 ± 18.3 | 0.2055 ± 0.0427 | 9.9 ± 0 |
| Guazuma ulmifolia | Tree | Biotic | Generalist | No | Yes | 16.9 ± 2.6 | 0.0162 ± 0.0035 | --- |
| Pachira quinata | Tree | Abiotic | Generalist | No | Yes | 107.3 ± 28.2 | 0.1119 ± 0.0174 | 10.3 ± 0 |
| Pseudobombax septenatum | Tree | Abiotic | Generalist | No | No | 262.7 ± 42 | 0.2347 ± 0.0331 | 24.2 ± 0.4 |
| Sterculia apetala | Tree | Biotic | Generalist | No | Yes | 7425.6 ± 685.3 | 5.0445 ± 0.7927 | 18.1 ± 3.8 |
| Thespesia populnea | Tree | Abiotic | Generalist | No | No | 659.2 ± 122.7 | 0.9187 ± 0.1853 | 11.2 ± 0.7 |
| MELIACEAE | ||||||||
| Cedrela odorata | Tree | Abiotic | Generalist | No | No | 71.4 ± 14.5 | 0.082 ± 0.0114 | 13.9 ± 2.2 |
| Swietenia macrophylla | Tree | Abiotic | Generalist | No | Yes | 1422.9 ± 357.9 | 1.1649 ± 0.1498 | 3.6 ± 0.2 |
| Trichilia appendiculata | Tree | Biotic | Generalist | No | No | 111.8 ± 20 | 0.0859 ± 0.0098 | 4.8 ± 0 |
| MORACEAE | ||||||||
| Ficus citrifolia | Tree | Biotic | Heliophyte | Yes | No | 5.1 ± 0.8 | 0.0011 ± 0.0003 | 7.2 ± 0 |
| Trophis cf. caucana | Tree | Biotic | Generalist | No | No | 585.6 ± 145.7 | 0.2676 ± 0.0543 | 27 ± 1.7 |
| MUNTINGIACEAE | ||||||||
| Muntingia calabura | Tree | Biotic | Heliophyte | Yes | Yes | 0.8 ± 0.1 | 0.0003 ± 0.0001 | --- |
| NYCTAGINACEAE | ||||||||
| Guapira pacurero | Tree | Biotic | Sciophyte | Yes | No | 139.3 ± 23.9 | 0.4885 ± 0.0709 | 39.2 ± 0.2 |
| POLYGONACEAE | ||||||||
| Coccoloba cf. caracasana | Tree | Biotic | Heliophyte | Yes | No | 220.5 ± 52.9 | 1.1716 ± 0.1789 | 22.1 ± 9.2 |
| Triplaris americana | Tree | Abiotic | Heliophyte | Yes | Yes | 153.8 ± 28.1 | 0.2886 ± 0.0533 | 11.1 ± 0.1 |
| RUBIACEAE | ||||||||
| Morinda royoc | Liana | Biotic | Sciophyte | Yes | No | 30.7 ± 6.6 | 0.0873 ± 0.0204 | 43.1 ± 0 |
| SALICACEAE | ||||||||
| Casearia arborea | Tree | Biotic | Generalist | No | Yes | 203.9 ± 19 | 0.1538 ± 0.0265 | 37.9 ± 1.5 |
| SAPINDACEAE | ||||||||
| Cupania cf. latifolia | Tree | Biotic | Generalist | No | Yes | 1181.9 ± 208.6 | 1.1685 ± 0.1767 | 38 ± 0.6 |
| Sapindus saponaria | Tree | Biotic | Heliophyte | Yes | Yes | 2876.3 ± 383.5 | 2.5974 ± 0.3211 | 12.1 ± 0.3 |
| Serjania cf. paniculata | Liana | Biotic | Generalist | No | No | 262.1 ± 74.3 | 0.271 ± 0.1129 | 21 ± 18.5 |
| ZYGOPHYLLACEAE | ||||||||
| Bulnesia arborea | Tree | Abiotic | Generalist | No | No | 454 ± 98 | --- | 13.7 ± 5.4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Londoño-Lemos, V.; Torres-Gonzáles, A.M.; Madriñán, S. Linking Seed Traits and Germination Responses in Caribbean Seasonally Dry Tropical Forest Species. Plants 2024, 13, 1318. https://doi.org/10.3390/plants13101318
Londoño-Lemos V, Torres-Gonzáles AM, Madriñán S. Linking Seed Traits and Germination Responses in Caribbean Seasonally Dry Tropical Forest Species. Plants. 2024; 13(10):1318. https://doi.org/10.3390/plants13101318
Chicago/Turabian StyleLondoño-Lemos, Viviana, Alba Marina Torres-Gonzáles, and Santiago Madriñán. 2024. "Linking Seed Traits and Germination Responses in Caribbean Seasonally Dry Tropical Forest Species" Plants 13, no. 10: 1318. https://doi.org/10.3390/plants13101318