



Development of S Haplotype-Specific Markers to Identify Genotypes of Self-Incompatibility in Radish (Raphanus sativus L.)


Abstract
:1. Introduction
2. Results
2.1. Amplification of SRK, SLG Alleles Using Universal Primers
2.2. Identification of S Haplotypes Based on BLAST Search
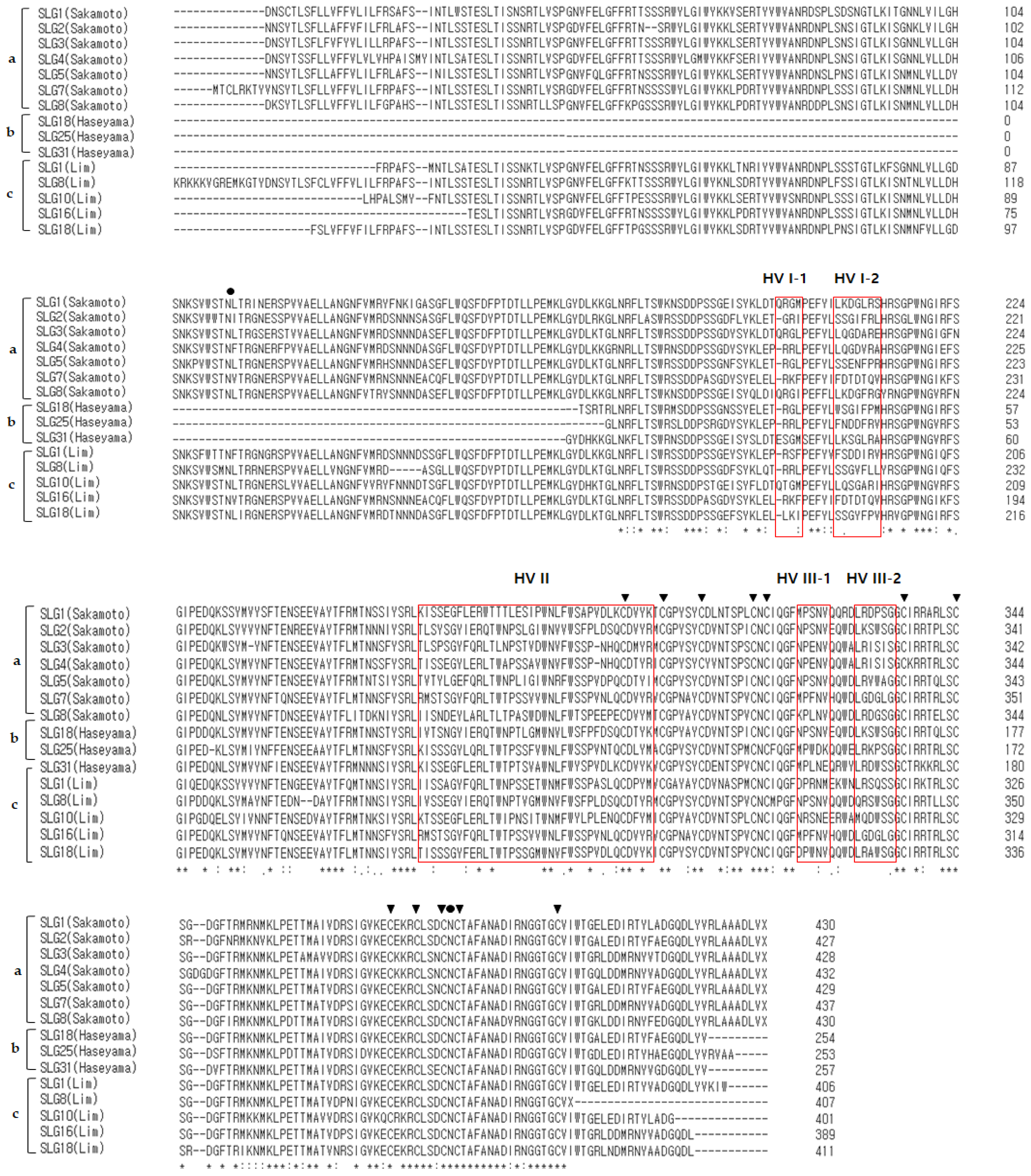
2.3. Multiple Sequence Alignment (MSA) of SRK, SLG Alleles


2.4. Development of S Haplotype-Specific Markers
3. Discussion and Conclusions
4. Materials and Methods
4.1. Plant Materials
4.2. Amplification and Sequencing of SRK, SLG Alleles
4.2.1. Extraction of Genomic DNA and Amplification of SRK, SLG Alleles
4.2.2. Identification of S Haplotypes Based on BLAST Search
4.3. Multiple Sequence Alignment (MSA)
4.4. Development of S Haplotype-Specific Markers
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Manivannan, A.; Kim, J.-H.; Kim, D.-S.; Lee, E.-S.; Lee, H.-E. Deciphering the nutraceutical potential of Raphanus sativus—A comprehensive overview. Nutrients 2019, 11, 402. [Google Scholar] [CrossRef]
- Gamba, M.; Asllanaj, E.; Raguindin, P.F.; Glisic, M.; Franco, O.H.; Minder, B.; Bussler, W.; Metzger, B.; Kern, H.; Muka, T. Nutritional and phytochemical characterization of radish (Raphanus sativus): A systematic review. Trends Food Sci. Technol. 2021, 113, 205–218. [Google Scholar] [CrossRef]
- De Nettancourt, D. Incompatibility in angiosperms. Sex. Plant Reprod. 1997, 10, 185–199. [Google Scholar] [CrossRef]
- Hinata, K.; Watanabe, M.; Toriyama, K.; Isogai, A. A review of recent studies on homomorphic self-incompatibility. Int. Rev. Cytol. 1993, 143, 257–296. [Google Scholar]
- Newbigin, E.; Anderson, M.A.; Clarke, A.E. Gametophytic Self-Incompatibility Systems. Plant Cell 1993, 5, 1315. [Google Scholar] [CrossRef]
- de Nettancourt, D.; de Nettancourt, D. The genetics of self-incompatibility. In Incompatibility and Incongruity in Wild and Cultivated Plants; Springer: Berlin/Heidelberg, Germany, 2001; pp. 25–72. [Google Scholar]
- Stone, S.L.; Goring, D.R. The molecular biology of self-incompatibility systems in flowering plants. Plant Cell Tissue Organ Cult. 2001, 67, 93–114. [Google Scholar] [CrossRef]
- Watanabe, M.; Suwabe, K.; Suzuki, G. Molecular genetics, physiology and biology of self-incompatibility in Brassicaceae. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2012, 88, 519–535. [Google Scholar] [CrossRef] [PubMed]
- Casselman, A.L.; Vrebalov, J.; Conner, J.A.; Singhal, A.; Giovannoni, J.; Nasrallah, M.E.; Nasrallah, J.B. Determining the physical limits of the Brassica S locus by recombinational analysis. Plant Cell 2000, 12, 23–33. [Google Scholar] [CrossRef]
- Kitashiba, H.; Nasrallah, J.B. Self-incompatibility in Brassicaceae crops: Lessons for interspecific incompatibility. Breed. Sci. 2014, 64, 23–37. [Google Scholar] [CrossRef]
- Takasaki, T.; Hatakeyama, K.; Suzuki, G.; Watanabe, M.; Isogai, A.; Hinata, K. The S receptor kinase determines self-incompatibility in Brassica stigma. Nature 2000, 403, 913–916. [Google Scholar] [CrossRef]
- Watanabe, M.; Hinata, K. 5 Self-incompatibility. In Developments in Plant Genetics and Breeding; Gómez-Campo, C., Ed.; Elsevier: Amsterdam, The Netherlands, 1999; Volume 4, pp. 149–183. [Google Scholar]
- Takayama, S.; Isogai, A. Self-Incompatibility IN Plants. Annu. Rev. Plant Biol. 2005, 56, 467–489. [Google Scholar] [CrossRef]
- Dixit, R.; Nasrallah Me Fau-Nasrallah, J.B.; Nasrallah, J.B. Post-transcriptional maturation of the S receptor kinase of Brassica correlates with co-expression of the S-locus glycoprotein in the stigmas of two Brassica strains and in transgenic tobacco plants. Plant Physiol. 2000, 124, 297–312. [Google Scholar] [CrossRef]
- Nasrallah, J.B.; Nasrallah, M.E. Pollen—Stigma Signaling in the Sporophytic Self-Incompatibility Response. Plant Cell 1993, 5, 1325–1335. [Google Scholar] [CrossRef] [PubMed]
- Takayama, S.; Isogai, A. Molecular mechanism of self-recognition in Brassica self-incompatibility. J. Exp. Bot. 2003, 54, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Jung, J.; Choi, Y.-O.; Kim, S. Development of a system for S locus haplotyping based on the polymorphic SLL2 gene tightly linked to the locus determining self-incompatibility in radish (Raphanus sativus L.). Euphytica 2016, 209, 525–535. [Google Scholar] [CrossRef]
- Stein, J.C.; Howlett, B.; Boyes, D.C.; Nasrallah, M.E.; Nasrallah, J.B. Molecular cloning of a putative receptor protein kinase gene encoded at the self-incompatibility locus of Brassica oleracea. Proc. Natl. Acad. Sci. USA 1991, 88, 8816–8820. [Google Scholar] [CrossRef]
- Watanabe, M.; Takasaki, T.; Toriyama, K.; Yamakawa, S.; Isogai, A.; Suzuki, A.; Hinata, K. A high degree of homology exists between the protein encoded by SLG and the S receptor domain encoded by SRK in self-incompatible Brassica campestris L. Plant Cell Physiol. 1994, 35, 1221–1229. [Google Scholar] [CrossRef]
- Hatakeyama, K.; Watanabe, M.; Takasaki, T.; Ojima, K.; Hinata, K. Dominance relationships between S-alleles in self-incompatible Brassica campestris L. Heredity 1998, 80, 241–247. [Google Scholar] [CrossRef]
- Sato, K.; Nishio, T.; Kimura, R.; Kusaba, M.; Suzuki, T.; Hatakeyama, K.; Ockendon, D.J.; Satta, Y. Coevolution of the S-locus genes SRK, SLG and SP11/SCR in Brassica oleracea and B. rapa. Genetics 2002, 162, 931–940. [Google Scholar] [CrossRef]
- Fujii, S.; Takayama, S. Multilayered dominance hierarchy in plant self-incompatibility. Plant Reprod. 2018, 31, 15–19. [Google Scholar] [CrossRef]
- Ni, M.; Yi, X.; Wang, Q.; Wang, J.; Wang, S.; Liu, L.; Xu, L.; Wang, Y. Classification and Identification of S Haplotypes in Radish Based on SRK Kinase Domain Sequence Analysis. Plants 2022, 11, 2304. [Google Scholar] [CrossRef]
- Wang, Q.; Zheng, P.; Zhang, L.; Snowdon, R. Identification and classification ofShaplotypes in radish (Raphanus sativus). Plant Breed. 2018, 138, 121–130. [Google Scholar] [CrossRef]
- Fang, Z.; Sun, P.; Liu, Y. Problems on the utilization of heterosis in cabbage and the selection of self-incompatibility genotypes. Chung-Kuo Nung Yeh K’o Hsueh = Sci. Agric. Sin. 1983, 3, 51–62. [Google Scholar]
- Pastuglia, M.; Roby, D.; Dumas, C.; Cock, J.M. Rapid induction by wounding and bacterial infection of an S gene family receptor-like kinase gene in Brassica oleracea. Plant Cell 1997, 9, 49–60. [Google Scholar] [PubMed]
- Lim, S.H.; Cho, J.; Lee, J.; Cho, Y.H.; Kim, B.D. Identification and classification of S haplotypes in Raphanus sativus by PCR-RFLP of the S locus glycoprotein (SLG) gene and the S locus receptor kinase (SRK) gene. Theor. Appl. Genet. 2002, 104, 1253–1262. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.T.; Hee, C.; Su, K.; Han Yong, P. Application of SCAR markers to self-incompatibility genotyping in breeding lines of radish (Raphanus sativus L.). Korean J. Breed. Sci. 2009, 41, 397–402. [Google Scholar]
- Tian, L.; Miao, W.; Liu, J.; Fang, Z.; Liu, Y.; Yang, L.; Zhang, Y.; Zhuang, M. Identification of S haplotypes in cabbage inbred lines (Brassica oleracea var. capitata L.). Sci. Hortic. 2013, 164, 400–408. [Google Scholar] [CrossRef]
- Nishio, T.; Sakamoto, K. Polymorphism of Self-Incompatibility Genes. In Radish Genome; Springer: Cham, Switzerland, 2017; pp. 177–188. [Google Scholar]
- Haseyama, Y.; Kitashiba, H.; Okamoto, S.; Tonouchi, E.; Sakamoto, K.; Nishio, T. Nucleotide sequence analysis of S-locus genes to unify S haplotype nomenclature in radish. Mol. Breed. 2018, 38, 116. [Google Scholar] [CrossRef]
- Matsuura, S.N.a.S. Identification of self-incompatibility alleles (S) by PCR-RFLP in radish (Raphanus sativus L.). Euphytica 1998, 102, 379–384. [Google Scholar]
- Sakamoto, K.; Kusaba, M.; Nishio, T. Polymorphism of the S-locus glycoprotein gene (SLG) and the S-locus related gene (SLR1) in Raphanus sativus L. and self-incompatible ornamental plants in the Brassicaceae. Mol. Gen. Genet. 1998, 258, 397–403. [Google Scholar] [CrossRef]
- Okamoto, S.; Sato, Y.; Sakamoto, K.; Nishio, T. Distribution of similar self-incompatibility (S) haplotypes in different genera, Raphanus and Brassica. Sex. Plant Reprod. 2004, 17, 33–39. [Google Scholar] [CrossRef]
- Delorme, V.; Giranton, J.L.; Hatzfeld, Y.; Friry, A.; Heizmann, P.; Ariza, M.J.; Dumas, C.; Gaude, T.; Cock, J.M. Characterization of the S locus genes, SLG and SRK, of the Brassica S3 haplotype: Identification of a membrane-localized protein encoded by the S locus receptor kinase gene. Plant J. 1995, 7, 429–440. [Google Scholar] [CrossRef]
- Seo, J.-P.; Park, K.-W.; Kim, H.-J.; Yang, S.-Y.; Nou, I.-S. Identification of S-haplotypes by PCR-RFLP and DNA Sequencing of S-Locus Receptor Kinase (SRK) in Raphanus sativus Inbred Lines. Hortic. Environ. Biotechnol. 2004, 45, 119–126. [Google Scholar]
- Chen, C.-H.; Nasrallah, J.B. A new class of S sequences defined by a pollen recessive self-incompatibility allele of Brassica oleracea. Mol. Gen. Genet. MGG 1990, 222, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Nishio, T.; Kusaba, M.; Watanabe, M.; Hinata, K. Registration of S alleles in Brassica campestris L by the restriction fragment sizes of SLGs. Theor. Appl. Genet. 1996, 92, 388–394. [Google Scholar] [CrossRef] [PubMed]
- Cho, G.; Kim, S. Integration of Korean S haplotypes controlling self-incompatibility into the unified nomenclature in radish (Raphanus sativus L.). J. Hortic. Sci. Biotechnol. 2022, 97, 58–65. [Google Scholar] [CrossRef]
- Cabrillac, D.; Delorme, V.; Garin, J.; Ruffio-Châble, V.; Giranton, J.-L.; Dumas, C.; Gaude, T.; Cock, J.M. The S15 Self-Incompatibility Haplotype in Brassica oleracea Includes Three S Gene Family Members Expressed in Stigmas. Plant Cell 1999, 11, 971–986. [Google Scholar] [CrossRef]
- Takuno, S.; Fujimoto, R.; Sugimura, T.; Sato, K.; Okamoto, S.; Zhang, S.L.; Nishio, T. Effects of recombination on hitchhiking diversity in the Brassica self-incompatibility locus complex. Genetics 2007, 177, 949–958. [Google Scholar] [CrossRef]
- Kusaba, M.; Nishio, T.; Satta, Y.; Hinata, K.; Ockendon, D. Striking sequence similarity in inter- and intra-specific comparisons of class I SLG alleles from Brassica oleracea and Brassica campestris: Implications for the evolution and recognition mechanism. Proc. Natl. Acad. Sci. USA 1997, 94, 7673–7678. [Google Scholar] [CrossRef] [PubMed]
- Kusaba, M.; Nishio, T. Comparative analysis of S haplotypes with very similar SLG alleles in Brassica rapa and Brassica oleracea. Plant J. 1999, 17, 83–91. [Google Scholar] [CrossRef]
- Sato, Y.; Sato, K.; Nishio, T. Interspecific pairs of class II S haplotypes having different recognition specificities between Brassica oleracea and Brassica rapa. Plant Cell Physiol. 2006, 47, 340–345. [Google Scholar] [CrossRef]
- Uyenoyama, M.K. Evolutionary dynamics of self-incompatibility alleles in Brassica. Genetics 2000, 156, 351–359. [Google Scholar] [CrossRef] [PubMed]
- Tantikanjana, T.; Nasrallah, M.E.; Stein, J.C.; Chen, C.H.; Nasrallah, J.B. An alternative transcript of the S locus glycoprotein gene in a class II pollen-recessive self-incompatibility haplotype of Brassica oleracea encodes a membrane-anchored protein. Plant Cell 1993, 5, 657–666. [Google Scholar] [CrossRef] [PubMed]
- Nasrallah, J.B.; Rundle, S.J.; Nasrallah, M.E. Genetic evidence for the requirement of the Brassica S-locus receptor kinase gene in the self-incompatibility response. Plant J. 1994, 5, 373–384. [Google Scholar] [CrossRef]
- Fujimoto, R.; Sugimura, T.; Nishio, T. Gene conversion from SLG to SRK resulting in self-compatibility in Brassica rapa. FEBS Lett. 2006, 580, 425–430. [Google Scholar] [CrossRef] [PubMed]
- Edh, K.; Widén, B.R.; Ceplitis, A. The Evolution and Diversification of S-Locus Haplotypes in the Brassicaceae Family. Genetics 2009, 181, 977–984. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.-S.; Kim, S. Identification of the S locus core sequences determining self-incompatibility and S multigene family from draft genome sequences of radish (Raphanus sativus L.). Euphytica 2017, 214, 16. [Google Scholar] [CrossRef]
- Watanabe, M.; Hatakeyama, K.; Takada, Y.; Hinata, K. Molecular Aspects of Self-Incompatibility in Brassica Species. Plant Cell Physiol. 2001, 42, 560–565. [Google Scholar] [CrossRef]
- Lee, O.N.; Park, H.Y. Assessment of genetic diversity in cultivated radishes (Raphanus sativus) by agronomic traits and SSR markers. Sci. Hortic. 2017, 223, 19–30. [Google Scholar] [CrossRef]
- Doyle, J.J.; Doyle, J.L. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Acc. | S Haplotype | S Alleles | Tool | Gene | NCBI Acc. | Iden (%) | Query Cov |
|---|---|---|---|---|---|---|---|
| SJ-1~4 | S1 (Lim) | SRK-KD (1063 bp) | BLASTn | SRK1 (Lim) | AY052579 | 100% | 99% |
| SLG (1220 bp) | BLASTn | SLG1 (Lim) | AY052572 | 99% | 99% | ||
| SJ-5~6 | S8 (Lim) | SRK-KD (1133 bp) | BLASTn | SRK8 (Lim) | AY052583 | 100% | 100% |
| SRK19 | KX961713 | 100% | 100% | ||||
| SRK-19 | LC341229 | 100% | 97% | ||||
| SRK-18 | LC341228 | 97% | 97% | ||||
| SLG (1201 bp) | BLASTn | SLG-19 | LC341238 | 99% | 55% | ||
| SJ-7~9 | S10 (Lim) | BLASTn | SRK10 (Lim) | AY052585 | 99.9% | 100% | |
| SRK-KD (1051 bp) | SRK11 (Lim) | AY534533 | 98% | 100% | |||
| SRK16 (Lim) | AY534535 | 98% | 100% | ||||
| SRK20 (Lim) | AY534537 | 98% | 100% | ||||
| SRK29 (Lim) | AY534541 | 99% | 100% | ||||
| SRK-22 | LC341231 | 98% | 95% | ||||
| SRK-23 | LC341232 | 98% | 99% | ||||
| SRK-31 | LC341234 | 99% | 96% | ||||
| SRK7 | KX961701 | 98% | 78% | ||||
| SRK10 | KX961704 | 98% | 78% | ||||
| SRK16 | KX961710 | 99% | 78% | ||||
| SRK17 | KX961711 | 98% | 78% | ||||
| SLG (1202 bp) | BLASTn | SLG10 (Lim) | AY052576 | 99.9% | 100% | ||
| SJ-10~11 | S16 (Lim) | SRK-KD (1071 bp) | BLASTn | SRK16 (Lim) | AY052579 | 99.9% | 100% |
| SRK-22 | LC341231 | 100% | 94% | ||||
| SRK10 (Lim) | AY052585 | 98% | 100% | ||||
| SRK11 (Lim) | AY534533 | 98% | 100% | ||||
| SRK20 (Lim) | AY534537 | 97% | 100% | ||||
| SRK29 (Lim) | AY534541 | 98% | 100% | ||||
| SRK-23 | LC341232 | 97% | 100% | ||||
| SRK-31 | LC341234 | 98% | 94% | ||||
| SRK7 | KX961701 | 99% | 76% | ||||
| SRK10 | KX961704 | 98% | 76% | ||||
| SRK16 | KX961710 | 99% | 76% | ||||
| SRK17 | KX961711 | 99% | 76% | ||||
| SLG (1168 bp) | BLASTn | SLG-22 | LC341239 | 100% | 64% | ||
| SLG-7 | AB009684 | 99.9% | 100% | ||||
| SJ-12~14 | S18 (Lim) | SRK-KD (1108 bp) | BLASTn | SRK18 (Lim) | AY534536 | 99.9% | 100% |
| SRK-6 | LC341226 | 99.9% | 95% | ||||
| SLG (1235 bp) | BLASTn | SLG18 (Lim) | AY527401 | 100% | 100% | ||
| SLG-6 | AB009682 | 100% | 100% | ||||
| SJ-15~17 | S4 (Lim) | SRK-KD (Not amplified) | - | - | - | - | - |
| SLG (1002 bp) | BLASTn | SLG4 (Lim) | AY052577 | 99% | 100% | ||
| SLG-26 | LC341241 | 99% | 99% | ||||
| SLG-11 | LC341236 | 97% | 87% | ||||
| SJ-18~20 | S5 (Lim) | SRK-KD (1053 bp) | BLASTn | SRK-5 (Wang) | - | 99% | 99% |
| SRK-9 (Wang) | - | 99% | 99% | ||||
| SRK6 | KP117077 | 99% | 94% | ||||
| SLG (1004 bp) | BLASTn | SRK-5 (Wang) | - | 99% | 94% | ||
| SLG5 (Lim) | AY052578 | 97% | 100% | ||||
| SLG21 (Lim) | AY529650 | 95% | 100% | ||||
| SLG24 (Lim) | AY529651 | 96% | 100% | ||||
| SRK1 | KX961695 | 96% | 100% | ||||
| SRK-9 | AB114851 | 96% | 98% | ||||
| SLG-9 | LC341235 | 95% | 90% | ||||
| Rs chr7 | OY743213 | 95,97% | 100% | ||||
| Rs chr8 | LR778317 | 95,97% | 100% | ||||
| SJ-21 | S21 (Lim) | SRK-KD (1015 bp) | BLASTn | SRK-9 (Wang) | - | 99% | 99% |
| SRK-5 (Wang) | - | 99% | 99% | ||||
| SRK6 | KP117077 | 99% | 96% | ||||
| SLG (1055 bp) | BLASTn | SLG21 (Lim) | AY529650 | 100% | 84% | ||
| SLG-9 | LC341235 | 100% | 77% | ||||
| Rs chr7 | OY743213 | 100,94% | 100% | ||||
| Rs chr8 | LR778317 | 100,94% | 100% | ||||
| Rs SLG S13-like | XM_056990347 | 100% | 100% | ||||
| SLG24 (Lim) | AY529651 | 96% | 84% | ||||
| SLG5 (Lim) | AY052578 | 94% | 84% | ||||
| SRK-1 | KX961695 | 94% | 100% | ||||
| SRK-9 | AB114851 | 94% | 100% | ||||
| SJ-22 | S26 (Lim) | SRK-KD (Not amplified) | - | - | - | - | - |
| SLG (802 bp) | BLASTn | SLG26 (Lim) | AY529652 | 100% | 100% | ||
| SLG-29 | LC341242 | 100% | 100% |
| S haplotype | Primer Set | Forward | Reverse | Expected Size |
|---|---|---|---|---|
| S1 (Lim) | SRK1 | KD1-F | KD1-R | 665 bp |
| SLG1 | SLG1-F | SLG1-R | 388 bp | |
| S4 (Lim) | SLG4 | SLG4-F | SLG4-R | 529 bp |
| S5 (Lim) | SLG5 | SLG5-F | SLG5-R | 409 bp |
| S8 (Lim) | SRK8 | KD8-F | KD8-R | 271 bp |
| SLG8 | SLG8-F | SLG8-R | 314 bp | |
| S10 (Lim) | SRK10 | KD10-F | KD10-R | 411 bp |
| SLG10 | SLG10-F | SLG10-R | 369 bp | |
| S16 (Lim) | SLG16 | SLG16-F | SLG16-R | 216 bp |
| S18 (Lim) | SRK18 | KD18-F | KD18-R | 105 bp |
| SLG18 | SLG18-F | SLG18-R | 353 bp | |
| S21 (Lim) | SLG21 | SLG21-F | SLG21-R | 529 bp |
| S26 (Lim) | SLG26 | SLG26-F | SLG26-R | 131 bp |
| Accession Number | Type of Fleshy Root | Color of Fleshy Root | Source |
|---|---|---|---|
| SJ-1 | Narrow elliptic | Green and White | South Chinese |
| SJ-2 | Acicular | Green and White | South Chinese |
| SJ-3 | Narrow elliptic | Green and White | South Chinese |
| SJ-4 | Oblong | Green and White | South Chinese |
| SJ-5 | Medium elliptic | Green and White | North Chinese |
| SJ-6 | Medium elliptic | Green and White | North Chinese |
| SJ-7 | Bell shaped | Green and White | North Chinese |
| SJ-8 | Oblong | Green and White | South Chinese |
| SJ-9 | Oblong | Green and White | South Chinese |
| SJ-10 | Oblong | Green and White | South Chinese |
| SJ-11 | Oblong | Green and White | South Chinese |
| SJ-12 | Ovate | Green and White | South Chinese |
| SJ-13 | Medium elliptic | Green and White | North Chinese |
| SJ-14 | Medium elliptic | Green and White | North Chinese |
| SJ-15 | Bell shaped | Green and White | North Chinese |
| SJ-16 | Oblong | White | South Chinese |
| SJ-17 | Oblong | White | South Chinese |
| SJ-18 | Bell shaped | Green and White | North Chinese |
| SJ-19 | Oblong | Green and White | South Chinese |
| SJ-20 | Oblong | Green and White | South Chinese |
| SJ-21 | Bell shaped | Green and White | South Chinese |
| SJ-22 | Narrow elliptic | Green and White | North Chinese |
| Gene | Forward | Reverse | Annealing Temperature/Cycle |
|---|---|---|---|
| Class I SRK-KD | UVSRK-F | UVSRK-R | 54 °C/34X |
| Class I SLG | SLG-I-F | SLG-I-R | 50 °C/39X |
| PS22 | SLG-I-R | 54 °C/35X | |
| Class II SRK-KD | KS2 | KA2 | 61 °C/35X |
| Class II SLG | SLG-II-F | SLG-II-R | 55.5 °C/39X |
| Rs9SLG-F | Rs9SLG-R | 53 °C/35X | |
| UVSLGII-F | UVSLGII-R | 61 °C/30X |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Heo, S.-H.; Kim, S.-Y.; Mo, S.-Y.; Park, H.-Y. Development of S Haplotype-Specific Markers to Identify Genotypes of Self-Incompatibility in Radish (Raphanus sativus L.). Plants 2024, 13, 725. https://doi.org/10.3390/plants13050725
Heo S-H, Kim S-Y, Mo S-Y, Park H-Y. Development of S Haplotype-Specific Markers to Identify Genotypes of Self-Incompatibility in Radish (Raphanus sativus L.). Plants. 2024; 13(5):725. https://doi.org/10.3390/plants13050725
Chicago/Turabian StyleHeo, Seong-Ho, Su-Yeon Kim, Suk-Yeon Mo, and Han-Yong Park. 2024. "Development of S Haplotype-Specific Markers to Identify Genotypes of Self-Incompatibility in Radish (Raphanus sativus L.)" Plants 13, no. 5: 725. https://doi.org/10.3390/plants13050725
APA StyleHeo, S.-H., Kim, S.-Y., Mo, S.-Y., & Park, H.-Y. (2024). Development of S Haplotype-Specific Markers to Identify Genotypes of Self-Incompatibility in Radish (Raphanus sativus L.). Plants, 13(5), 725. https://doi.org/10.3390/plants13050725