
Biological Control of Three Major Cucumber and Pepper Pests: Whiteflies, Thrips, and Spider Mites, in High Plastic Tunnels Using Two Local Phytoseiid Mites
 , , , , , , and
, , , , , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Site I (Spring Crop)
2.1.1. Plastic Tunnel Environmental Conditions
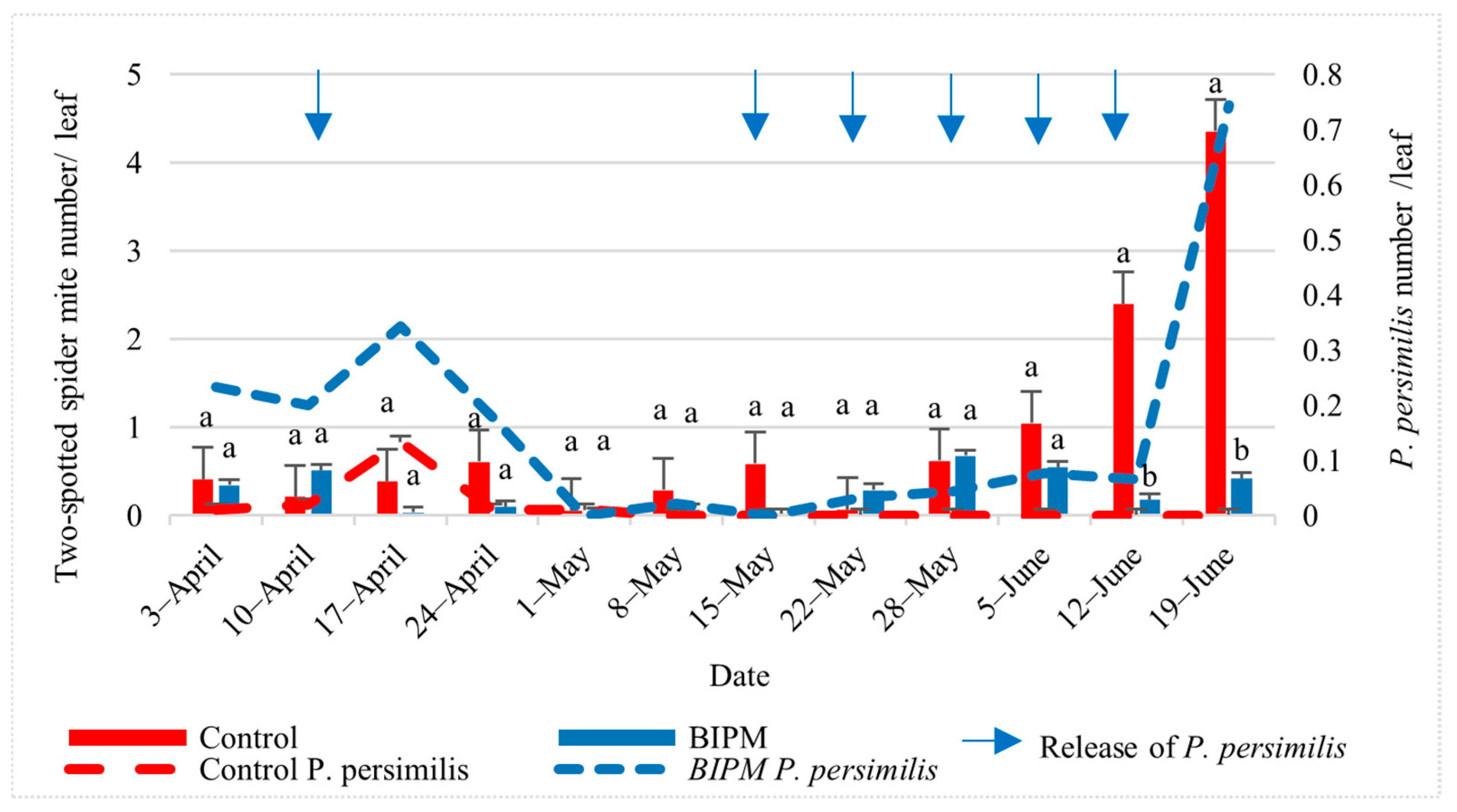
2.1.2. Control of Spider Mites on Cucumbers
2.1.3. Control of Spider Mites on Peppers
2.1.4. Control of Whiteflies on Cucumber
2.1.5. Control of Whiteflies on Pepper
2.1.6. Control of Thrips on Cucumber
2.1.7. Control of Thrips on Pepper
2.2. Site II (Fall Crop)
2.2.1. Plastic Tunnel Environmental Conditions
2.2.2. Control of Spider Mites on Cucumber
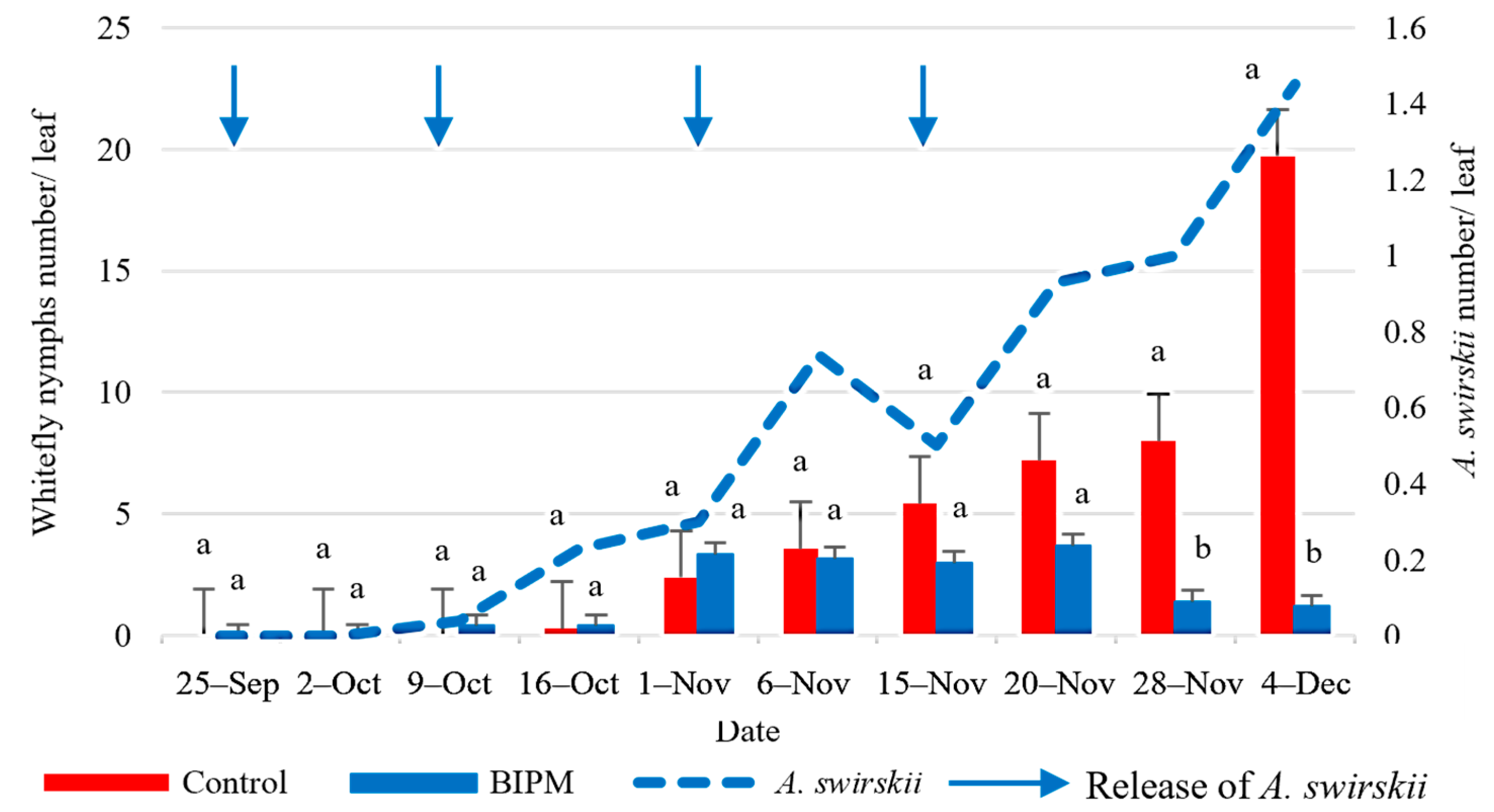
2.2.3. Control of Whitefly on Cucumber
2.2.4. Control of Thrips
3. Discussion
4. Materials and Methods
4.1. Biocontrol Agents
4.2. Site Selection and Greenhouse Structures
4.3. Pre-Transplanting Measures in BIPM Greenhouses
4.4. Post-Transplanting Measures
4.4.1. Insect/Mite Scouting
4.4.2. Control Tunnels
4.4.3. Intrinsic Rate of Increase
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Diouf, J. FAO’s Director-General on How to Feed the world in 2050. Popul. Dev. Rev. 2009, 35, 837–839. [Google Scholar]
- Ray, D.K.; Mueller, N.D.; West, P.; Foley, J.A. Yield trends are insufficient to double global crop production by 2050. PLoS ONE 2013, 8, e66428. [Google Scholar] [CrossRef]
- Talukdar, M.C.; Sarma, B.; Das, S.; Mahanta, S. Evaluation of spray chrysanthemum cultivars under open and polyhouse conditions. In Proceedings of the National Symposium on Recent Advances in Indian Floriculture, Trichur, India, 12–14 November 2003; pp. 12–14. [Google Scholar]
- Pardossi, A.; Tognoni, F.; Incrocci, L. Mediterranean greenhouse technology. Chron. Hortic. 2004, 44, 28–34. [Google Scholar]
- Cuesta Roble Releases 2019 Global Greenhouse Statistics. (7 January 2019). Produce Grower. Available online: https://www.producegrower.com/news/cuesta-roble-2019-global-greenhouse-statistics/ (accessed on 5 February 2024).
- Sanzua, L.J.; Saha, H.M.; Mwafaida, J. Status of greenhouse farming in the coastal humid climatic region of Kenya. Univers. J. Agric. Res. 2018, 6, 165–172. [Google Scholar]
- Alsadon, A.A.; Al-Helal, I.; Ibrahim, A.A.; Abdel-Ghany, A.M.; Al-Zaharani, S.M.; Ashour, T. The effects of plastic greenhouse covering on cucumber (Cucumis sativus L.) growth. Ecol. Eng. 2016, 87, 305–312. [Google Scholar] [CrossRef]
- FAO. Global production of vegetables in 2021, by type (in milllion metric tons). In Statista; FAO: Rome, Italy, 2021; Available online: https://www.statista.com/statistics/264065/global-production-of-vegetables-by-type/ (accessed on 4 January 2024).
- Wakil, W.; Brust, G.E.; Perring, T.M. Tomato and management of associated arthropod pests: Past, present, and future. In Sustainable Management of Arthropod Pests of Tomato; Academic Press: Cambridge, MA, USA, 2018; pp. 3–12. [Google Scholar]
- Sato, M.E.; Da Silva, M.Z.; Raga, A.; De Souza Filho, M.F. Abamectin resistance in Tetranychus urticae Koch (Acari: Tetranychidae): Selection, cross-resistance and stability of resistance. Neotrop. Entomol. 2005, 34, 991–998. [Google Scholar] [CrossRef]
- Şovărel, G.; Costache, M.; Cenuşă, A.; Hogea, S. Aspects of simultaneous control of the pathogens and pests on cucumber crops under high plastic tunnels. Curr. Trends Nat. Sci. 2019, 8, 139–143. [Google Scholar]
- Van Lenteren, J.; Bolckmans, K.; Köhl, J.; Ravensberg, W.J.; Urbaneja, A. Biological control using invertebrates and microorganisms: Plenty of new opportunities. BioControl 2017, 63, 39–59. [Google Scholar] [CrossRef]
- Abd-Rabou, S.; Simmons, A.M. Survey of Reproductive Host Plants of Bemisia tabaci (Hemiptera: Aleyrodidae) in Egypt, Including New Host Records. Entomol. News 2010, 121, 456–465. [Google Scholar] [CrossRef]
- Kareh, C.; Nemer, N. Evaluation of Insecticides in the Management of Whiteflies (Bemisia tabaci Gennadius) and their Impacts on Yield of Eggplants. Univers. J. Agric. Res. 2023, 11, 715–722. [Google Scholar] [CrossRef]
- Luan, J.; Li, J.; Varela, N.; Wang, Y.; Li, F.; Bao, Y.; Zhang, C.; Liu, S.; Wang, X. Global analysis of the transcriptional response of whitefly to tomato yellow leaf curl China virus reveals the relationship of coevolved adaptations. J. Virol. 2011, 85, 3330–3340. [Google Scholar] [CrossRef]
- Navas-Castillo, J.; Fiallo-Olivé, E.; Sánchez-Campos, S. Emerging virus diseases transmitted by whiteflies. Annu. Rev. Phytopathol. 2011, 49, 219–248. [Google Scholar] [CrossRef] [PubMed]
- Calvo, F.; Bolckmans, K.; Belda, J.E. Control of Bemisia tabaci and Frankliniella occidentalis in cucumber by Amblyseius swirskii. BioControl 2010, 56, 185–192. [Google Scholar] [CrossRef]
- Shipp, J.L.; Wang, K.; Binns, M. Economic injury levels for western flower thrips (Thysanoptera: Thripidae) on greenhouse cucumber. J. Econ. Entomol. 2000, 93, 1732–1740. [Google Scholar] [CrossRef] [PubMed]
- Steiner, M.Y. Determining Population Characteristics and Sampling Procedures for the Western Flower Thrips (Thysanoptera: Thripidae) and the Predatory Mite Amblyseius cucumeris (Acari: Phytoseiidae) on Greenhouse Cucumber. Environ. Entomol. 1990, 19, 1605–1613. [Google Scholar] [CrossRef]
- Tatineni, S.; Hein, G.L. Plant viruses of agricultural importance: Current and future perspectives of virus disease management strategies. Phytopathology® 2023, 113, 117–141. [Google Scholar] [CrossRef] [PubMed]
- Bielza, P.; Moreno, I.; Belando, A.; Grávalos, C.; Izquierdo, J.; Nauen, R. Spiromesifen and spirotetramat resistance in field populations of Bemisia tabaci Gennadius in Spain. Pest Manag. Sci. 2018, 75, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Jeschke, P. Status and outlook for acaricide and insecticide discovery. Pest Manag. Sci. 2021, 77, 64–76. [Google Scholar] [CrossRef] [PubMed]
- Feyereisen, R.; Dermauw, W.; Van Leeuwen, T. Genotype to phenotype, the molecular and physiological dimensions of resistance in arthropods. Pestic. Biochem. Physiol. 2015, 121, 61–77. [Google Scholar] [CrossRef]
- Desneux, N.; Wajnberg, E.; Wyckhuys, K.A.; Burgio, G.; Arpaia, S.; Narváez-Vasquez, C.A.; González-Cabrera, J.; Catalán Ruescas, D.; Tabone, E.; Frandon, J.; et al. Biological invasion of European tomato crops by Tuta absoluta: Ecology, geographic expansion and prospects for biological control. J. Pest Sci. 2010, 83, 197–215. [Google Scholar] [CrossRef]
- Guedes, R.N.C.; Picanço, M.C. The tomato borer Tuta absoluta in South America: Pest status, management and insecticide resistance. EPPO Bull. 2012, 42, 211–216. [Google Scholar] [CrossRef]
- Chen, D.; Huang, H.; Huang, Y.; Yang, W.; Shan, W.; Hao, G.; Wu, J.; Song, B. Toxicity Tests for Chemical Pesticide Registration: Requirement Differences among the United States, the European Union, Japan, and China? J. Agric. Food Chem. 2023, 71, 7192–7200. [Google Scholar] [CrossRef]
- Parsa, S.; Morse, S.; Bonifacio, A.; Chancellor, T.; Condori, B.; Crespo-Pérez, V.; Hobbs, S.L.A.; Kroschel, J.; Bâ, M.; Rebaudo, F.; et al. Obstacles to integrated pest management adoption in developing countries. Proc. Natl. Acad. Sci. USA 2014, 111, 3889–3894. [Google Scholar] [CrossRef]
- Ullah, M.S.; Lim, U.T. Laboratory evaluation of the effect of Beauveria bassiana on the predatory mite Phytoseiulus persimilis (Acari: Phytoseiidae). J. Invertebr. Pathol. 2017, 148, 102–109. [Google Scholar] [CrossRef]
- Omkar. Ecofriendly Pest Management for Food Security; Elsevier eBooks; Elsevier: San Diego, CA, USA, 2016. [Google Scholar]
- Bolckmans, K.; Van Houten, Y.; Hoogerbrugge, H. Biological control of whiteflies and western flower thrips in greenhouse sweet peppers with the phytoseiid predatory mite Amblyseius swirskii Athiashenriot (Acari: Phytoseiidae). In Proceedings of the 2nd International Symposium on Biological Control of Arthropods, Davos, Switzerland, 12–16 September 2005; pp. 555–565. [Google Scholar]
- Ghazy, N.A.; Suzuki, T.; Amano, H.; Ohyama, K. Air temperature optimisation for humidity-controlled cold storage of the predatory mites Neoseiulus californicus and Phytoseiulus persimilis (Acari: Phytoseiidae). Pest Manag. Sci. 2013, 70, 483–487. [Google Scholar] [CrossRef]
- Opit, G.P.; Nechols, J.R.; Margolies, D.C. Biological control of twospotted spider mites, Tetranychus urticae Koch (Acari: Tetranychidae), using Phytoseiulus persimilis Athias-Henriot (Acari: Phytoseidae) on ivy geranium: Assessment of predator release ratios. Biol. Control 2004, 29, 445–452. [Google Scholar] [CrossRef]
- Paulitz, T.C.; Bélanger, R.R. Biological control in greenhouse systems. Annu. Rev. Phytopathol. 2001, 39, 103–133. [Google Scholar] [CrossRef]
- Insect Management on Fruiting Vegetables in North Carolina. (n.d.). NC State Extension. Available online: https://entomology.ces.ncsu.edu/insect-management-on-fruiting-vegetables-in-north-carolina (accessed on 10 December 2023).
- Bin-Bin, S.; Ren, S.; Musa, P.H.; Chao, C. A Study on Economic Threshold of Bemisia tabaci. Acta Agric. Univ. Jiangxiensis 2005, 27, 234–237. [Google Scholar]
- Lee, H.; Gillespie, D.R. Life tables and development of Amblyseius swirskii (Acari: Phytoseiidae) at different temperatures. Exp. Appl. Acarol. 2010, 53, 17–27. [Google Scholar] [CrossRef]
- Yanar, D.; Gebologlu, N.; Cakar, T.; Engur, M. The use of predatory mite Phytoseiulus persimilis (Acari: Phytoseiidae) in the control of two spotted sider mite (Tetranychus urticae, Acari, Tetranychidae) at greenhouse cucumber production in Tokat province, Turkey. Appl. Ecol. Environ. Res. 2019, 17, 2033–2041. [Google Scholar] [CrossRef]
- Stavrinides, M.C.; Daane, K.M.; Lampinen, B.; Mills, N.J. Plant water stress, leaf temperature, and spider mite (Acari: Tetranychidae) outbreaks in California vineyards. Environ. Entomol. 2010, 39, 1232–1241. [Google Scholar] [CrossRef]
- Malais, M.; Ravensberg, W.J. Knowing and Recognizing: The Biology of Glasshouse Pests and Their Natural Enemies; Koppert BV: Berkel en Rodenrijs, The Netherlands, 2004. [Google Scholar]
- Messelink, G.; Van Steenpaal, S.; Van Wensveen, W. Typhlodromips swirskii (Athias-Henriot) (Acari: Phytoseiidae): A new predator for thrips control in greenhouse cucumber. IOBC/WPRS Bull. 2005, 28, 183–186. [Google Scholar]
- Calvo, J.; Bolckmans, K.; Belda, J.E. Controlling the tobacco whitefly Bemisia tabaci (Genn.)(Hom.: Aleyrodidae) in horticultural crops with the predatory mite Amblyseius swirskii (Athias-Henriot). J. Insect Sci. 2008, 8. Available online: https://link.gale.com/apps/doc/A197856732/AONE?u=anon~6dd598a0&sid=googleScholar&xid=e2ac09af (accessed on 5 February 2024).
- Calvo, F.; Lorente, M.J.; Stansly, P.A.; Belda, J.E. Preplant release of Nesidiocoris tenuis and supplementary tactics for control of Tuta absoluta and Bemisa tabaci in greenhouse tomato. Entomol. Exp. Appl. 2012, 143, 111–119. [Google Scholar] [CrossRef]
- Calvo, J.; Belda, J.E. Comparison of biological control strategies for Bemisia tabaci Genn (Hom.: Aleyrodidae) in sweet pepper under semifield conditions. Boletín de Sanidad Vegetal. Plagas 2006, 32, 297–311. [Google Scholar]
- Messelink, G.; Jansen, A. Do whiteflies help controlling thrips. IOBC/WPRS Bull. 2008, 2008, 131–134. [Google Scholar]
- Nomikou, M.; Sabelis, M.W.; Janssen, A. Pollen subsidies promote whitefly control through the numerical response of predatory mites. BioControl 2009, 55, 253–260. [Google Scholar] [CrossRef]
- Albendín, G.; Del Castillo García, M.; Molina, J.M. Multiple natural enemies do not improve two spotted spider mite and flower western thrips control in strawberry tunnels. Chil. J. Agric. Res. 2015, 75, 63–70. [Google Scholar] [CrossRef]
- Fathipour, Y.; Maleknia, B. Mite Predators; Elsevier eBooks; Elsevier: San Diego, CA, USA, 2016; pp. 329–366. [Google Scholar]
- Lanzoni, A.; Martelli, R.; Pezzi, F. Mechanical release of Phytoseiulus persimilis and Amblyseius swirskii on protected crops. Bull. Insectol. 2017, 70, 245–250. [Google Scholar]
- Navajas, M.; Lagnel, J.; Fauvel, G.; De Moraes, G. Sequence variation of ribosomal internal transcribed spacers (ITS) in commercially important Phytoseiidae mites. Exp. Appl. Acarol. 1999, 23, 851–859. [Google Scholar] [CrossRef]
- Elzinga, C. Population Biology: Life Tables & Theoretical Populations; Laboratory Manual for Honors Organismal Biology; Michigan State University: East Lansing, MI, USA, 2014; pp. 37–48. [Google Scholar]












Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abou Jawdah, Y.; Ezzeddine, N.; Fardoun, A.; Kharroubi, S.; Sobh, H.; Atamian, H.S.; Skinner, M.; Parker, B. Biological Control of Three Major Cucumber and Pepper Pests: Whiteflies, Thrips, and Spider Mites, in High Plastic Tunnels Using Two Local Phytoseiid Mites. Plants 2024, 13, 889. https://doi.org/10.3390/plants13060889
Abou Jawdah Y, Ezzeddine N, Fardoun A, Kharroubi S, Sobh H, Atamian HS, Skinner M, Parker B. Biological Control of Three Major Cucumber and Pepper Pests: Whiteflies, Thrips, and Spider Mites, in High Plastic Tunnels Using Two Local Phytoseiid Mites. Plants. 2024; 13(6):889. https://doi.org/10.3390/plants13060889
Chicago/Turabian StyleAbou Jawdah, Yusuf, Nour Ezzeddine, Aya Fardoun, Samer Kharroubi, Hana Sobh, Hagop S. Atamian, Margaret Skinner, and Bruce Parker. 2024. "Biological Control of Three Major Cucumber and Pepper Pests: Whiteflies, Thrips, and Spider Mites, in High Plastic Tunnels Using Two Local Phytoseiid Mites" Plants 13, no. 6: 889. https://doi.org/10.3390/plants13060889