

The Efficiency of CBD Production Using Grafted Cannabis sativa L. Plants Is Highly Dependent on the Type of Rootstock: A Study

Abstract
:1. Introduction
2. Results
2.1. Success of Two-Step Grafted Plant Preparation
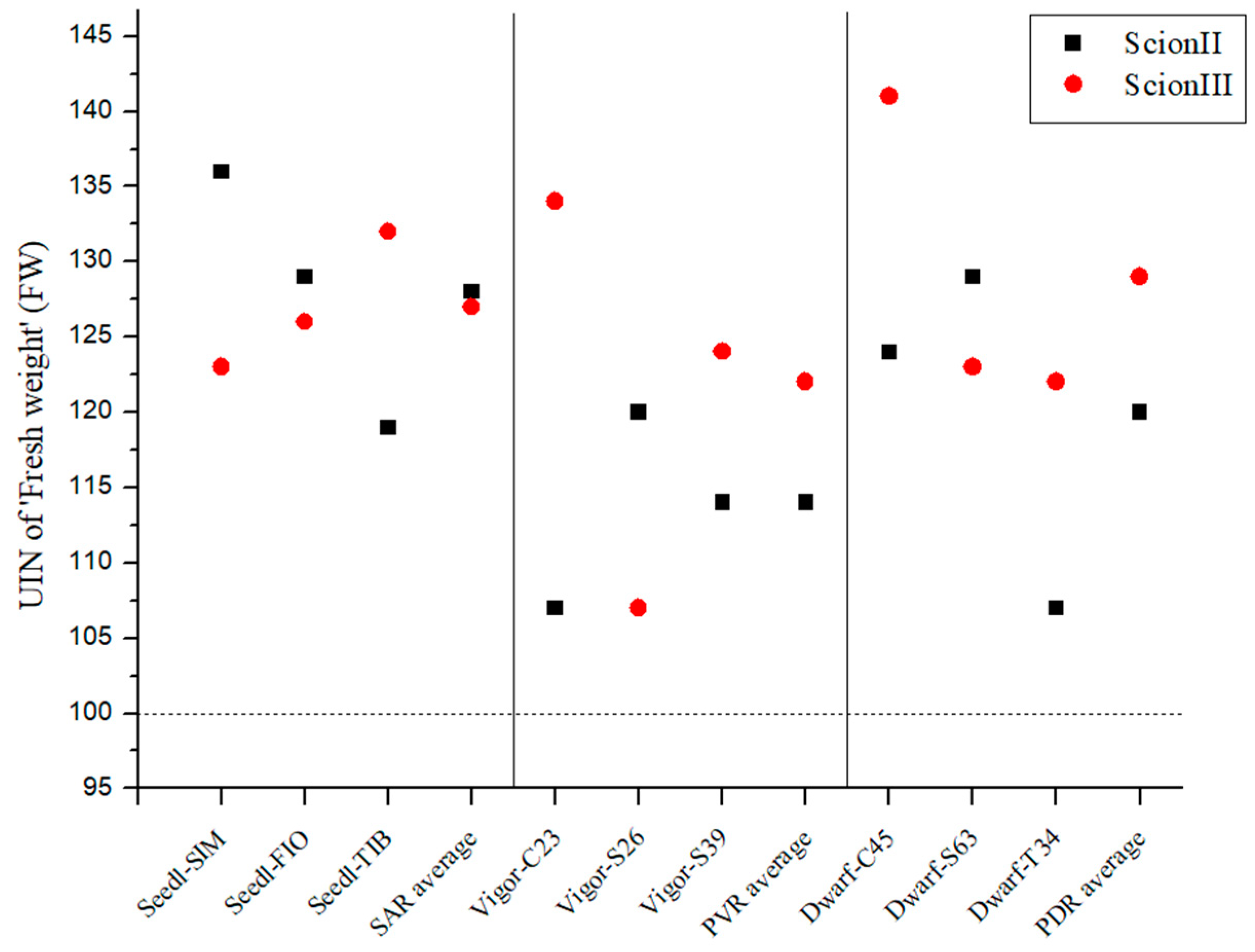
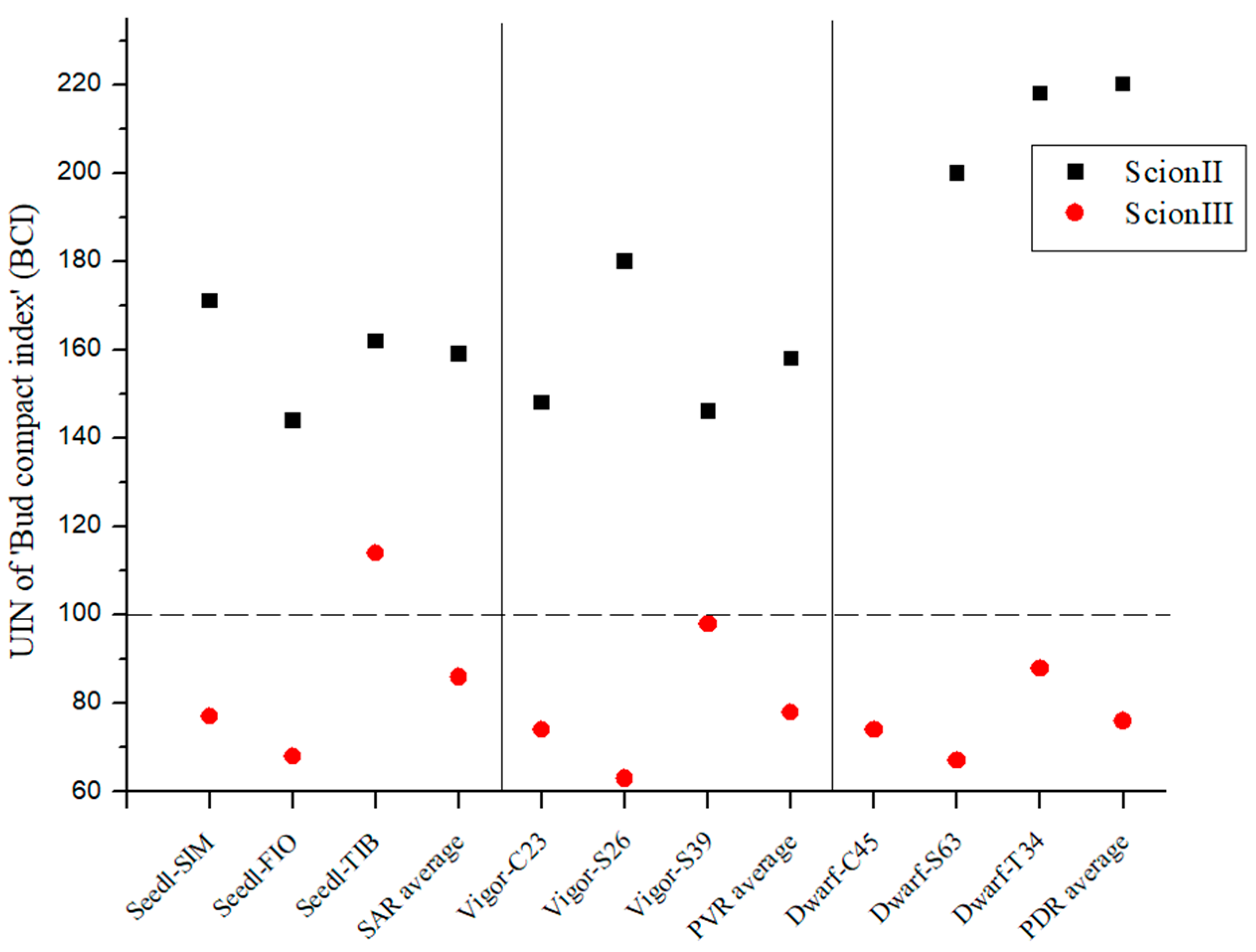
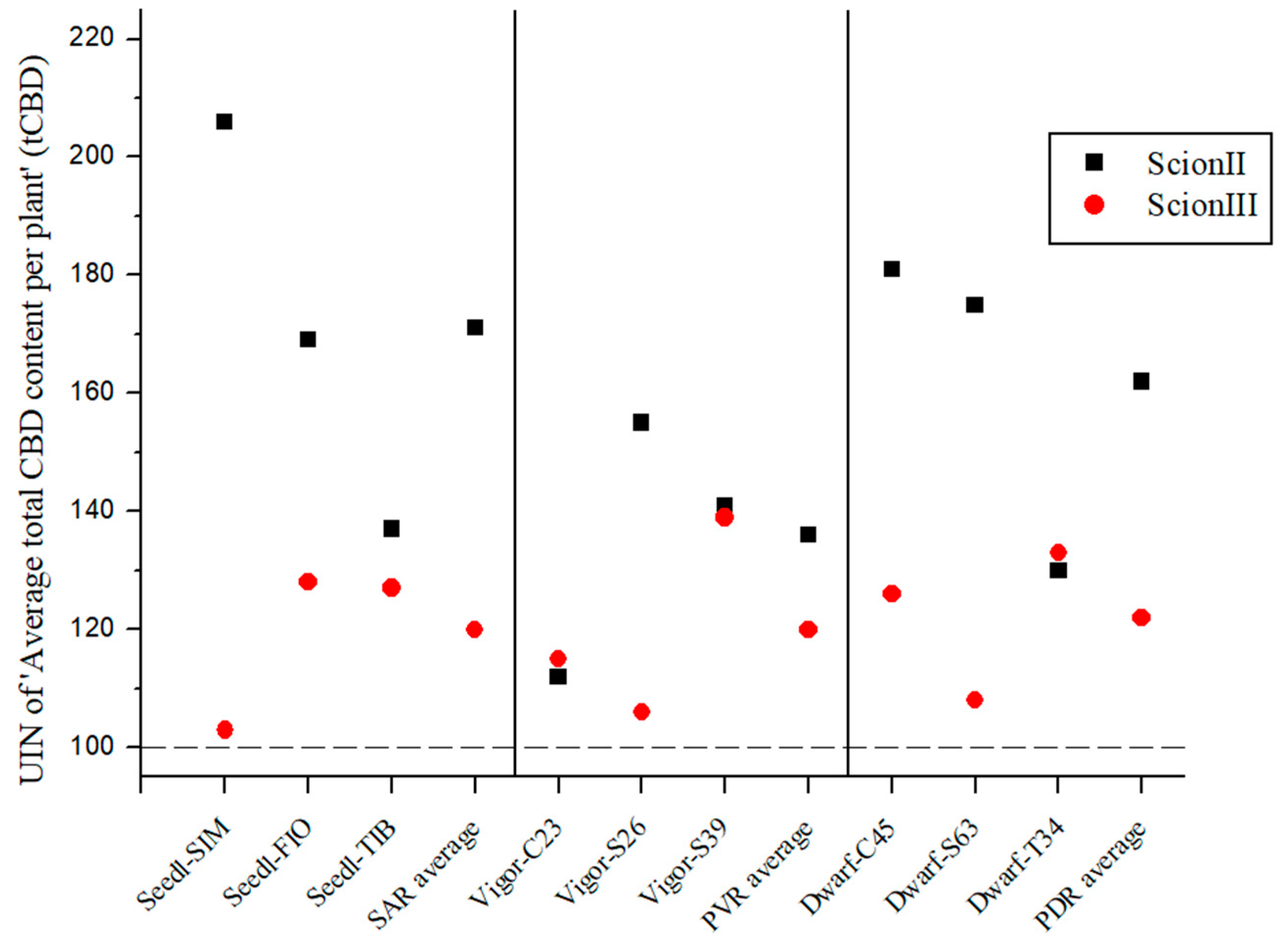
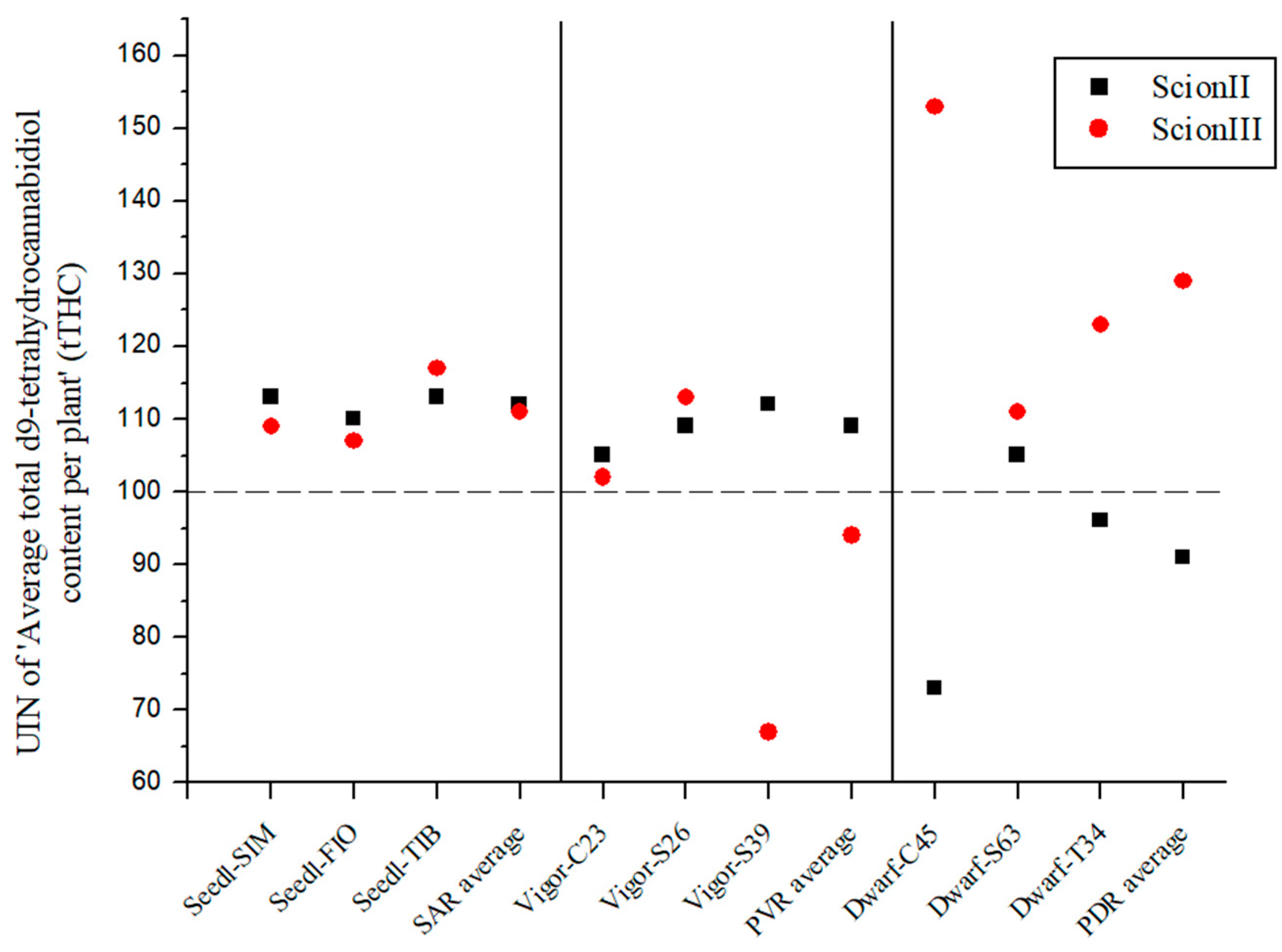
2.2. Influence of Rootstock on Morphological, Biochemical and Yield Parameters of Cannabis Plants
2.2.1. Influence of Rootstock on Morphological Variables of Grafted Cannabis Plants
2.2.2. Influence of Rootstock on Biochemical Variables of Grafted Cannabis Plants
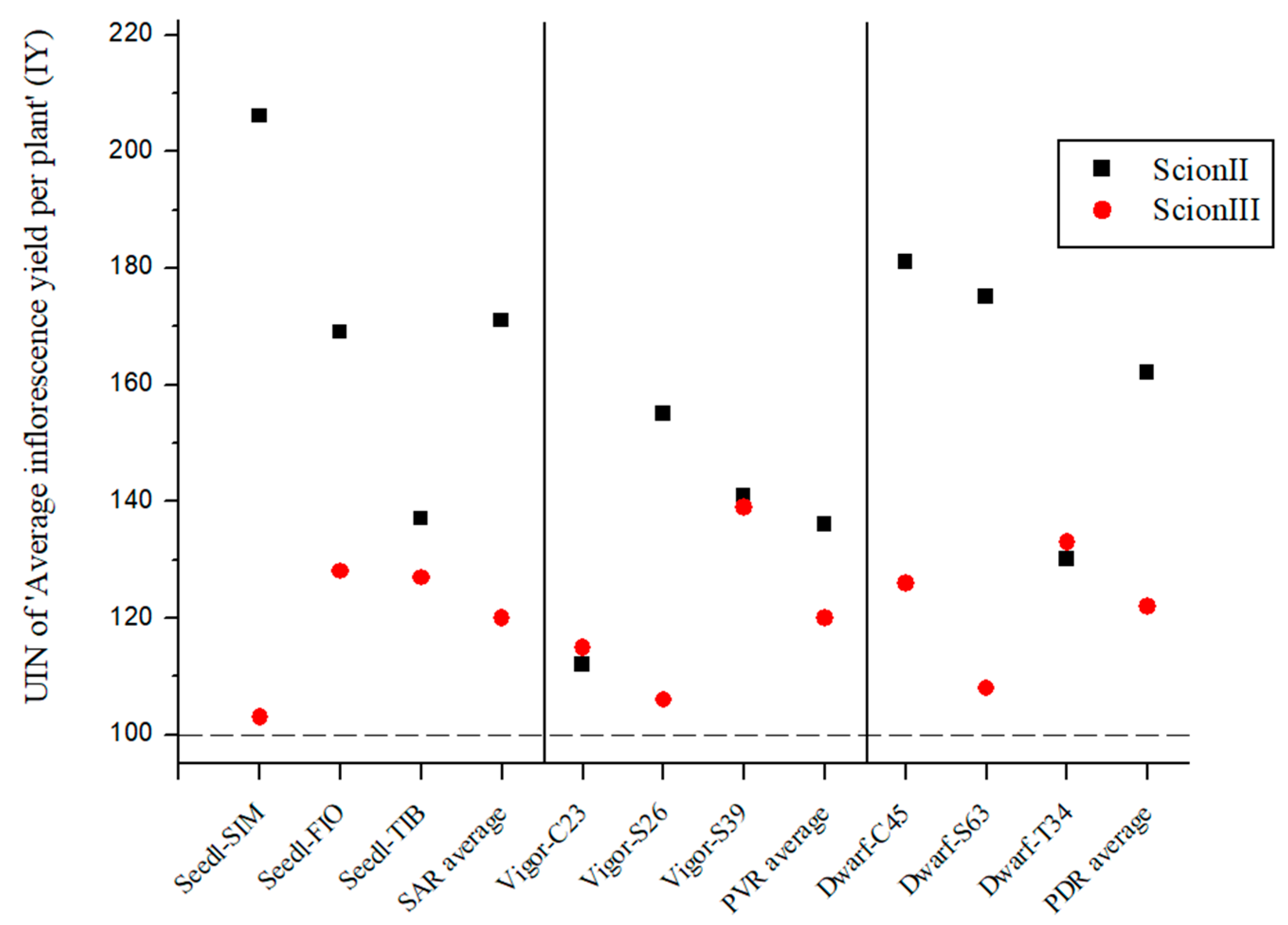
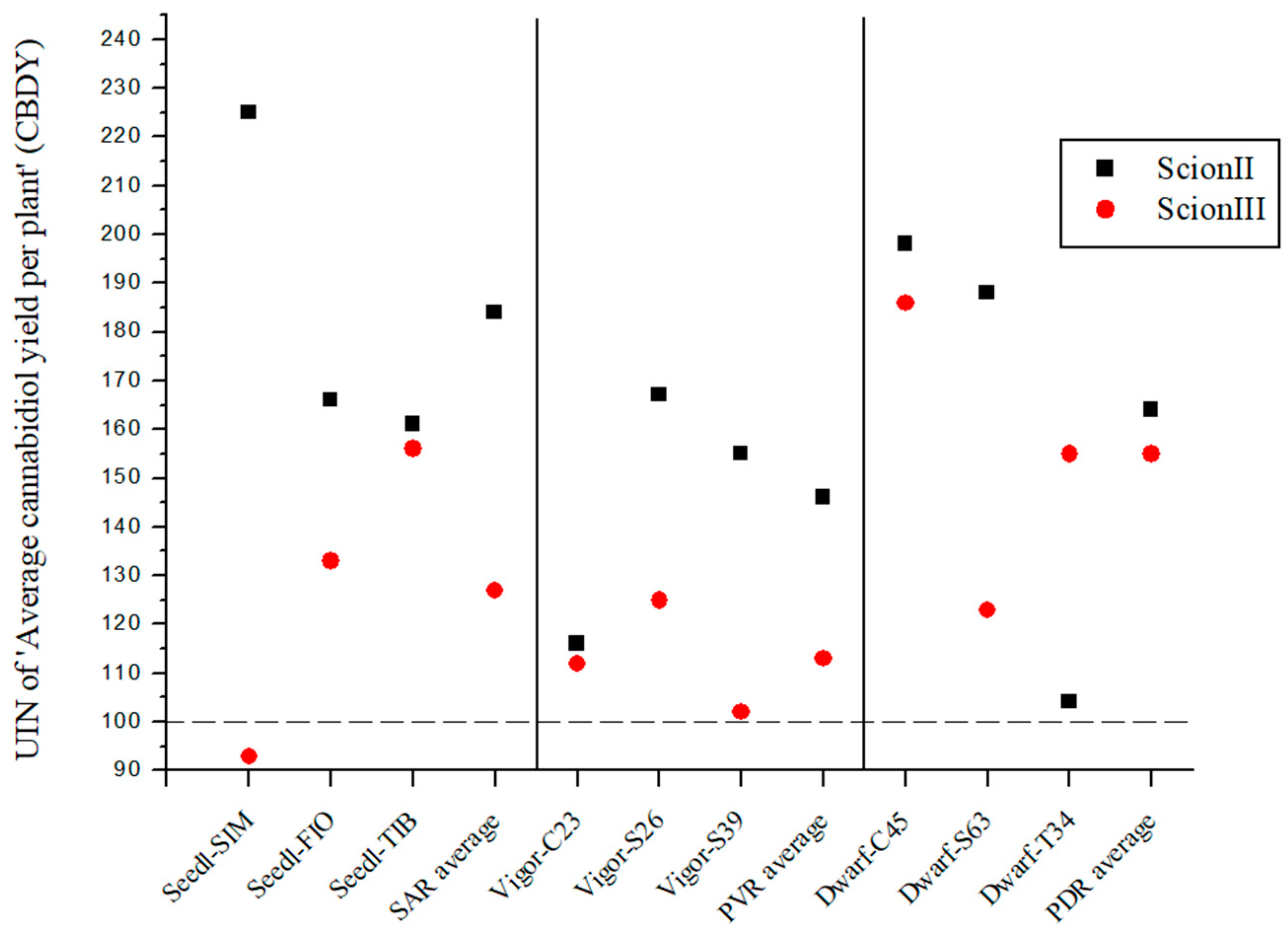
2.2.3. Influence of Rootstock on Yield Parameters of Grafted Cannabis Plants
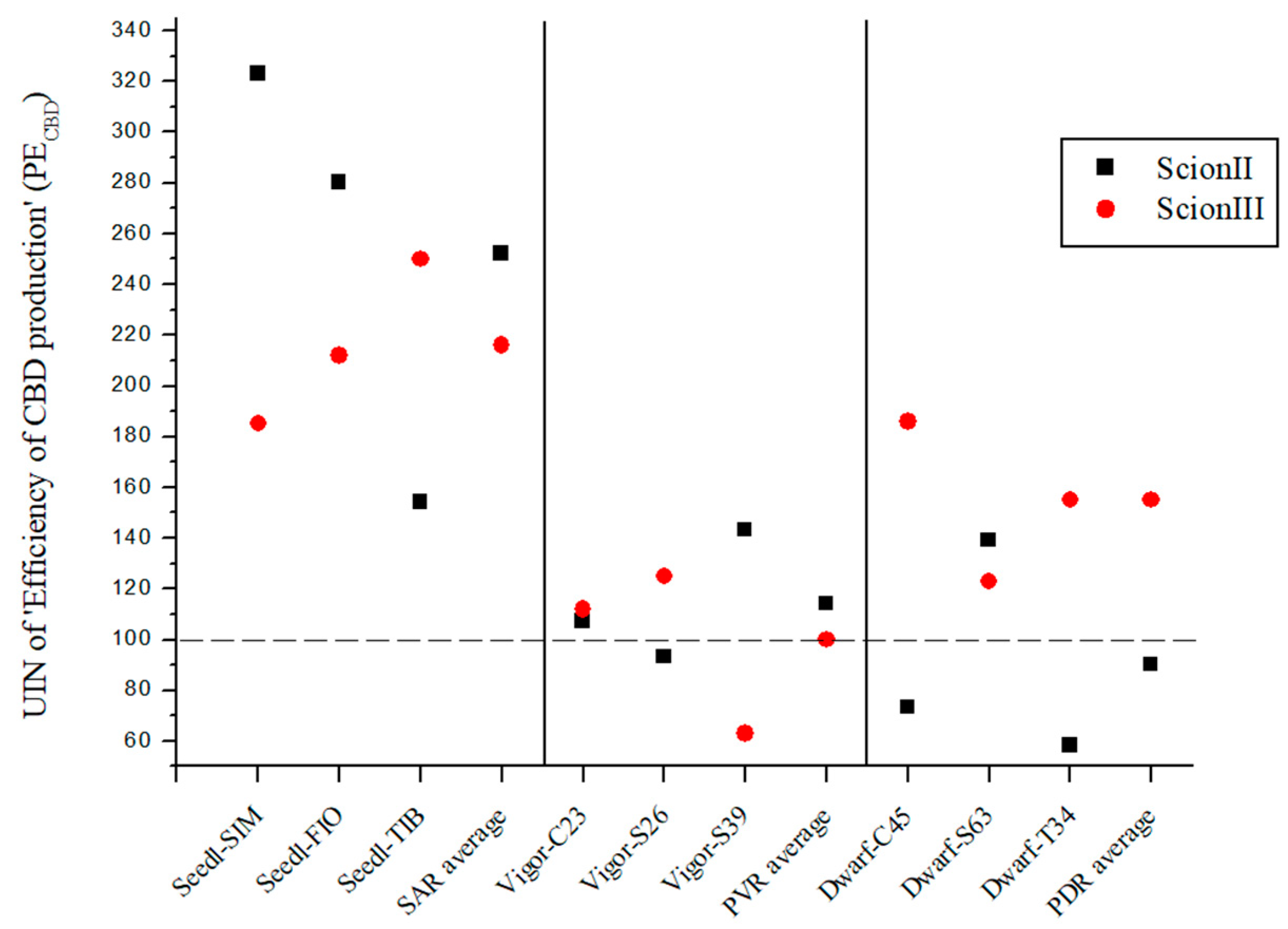
2.3. Efficiency of CBD Production (PECBD) Using Grafted Plants Is Highly Dependent on Rootstock Selection
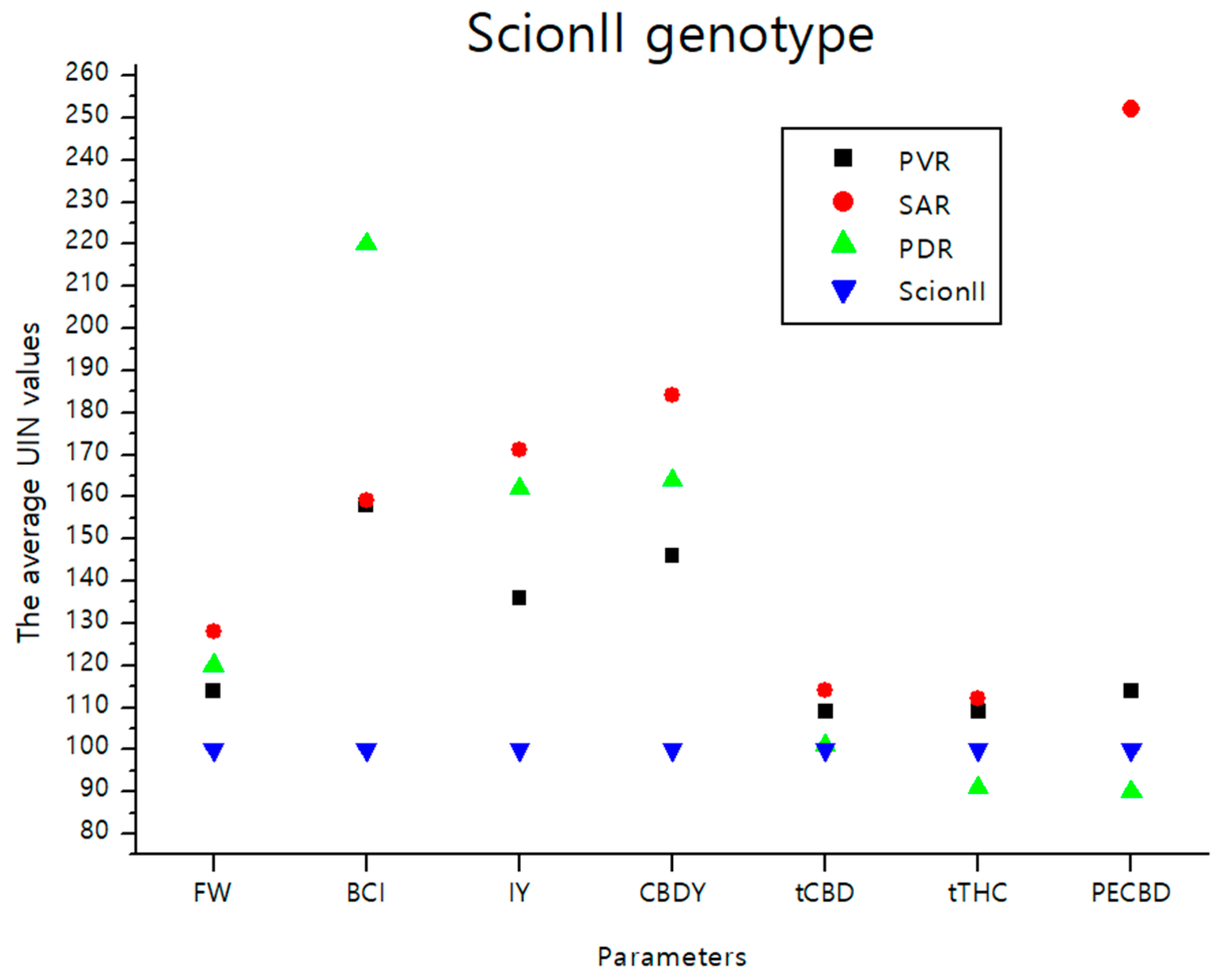
2.4. Comparison of the Influence of Rootstock Type on Measured Variables by Scion Genotype
2.4.1. ‘ScionII’ Genotype
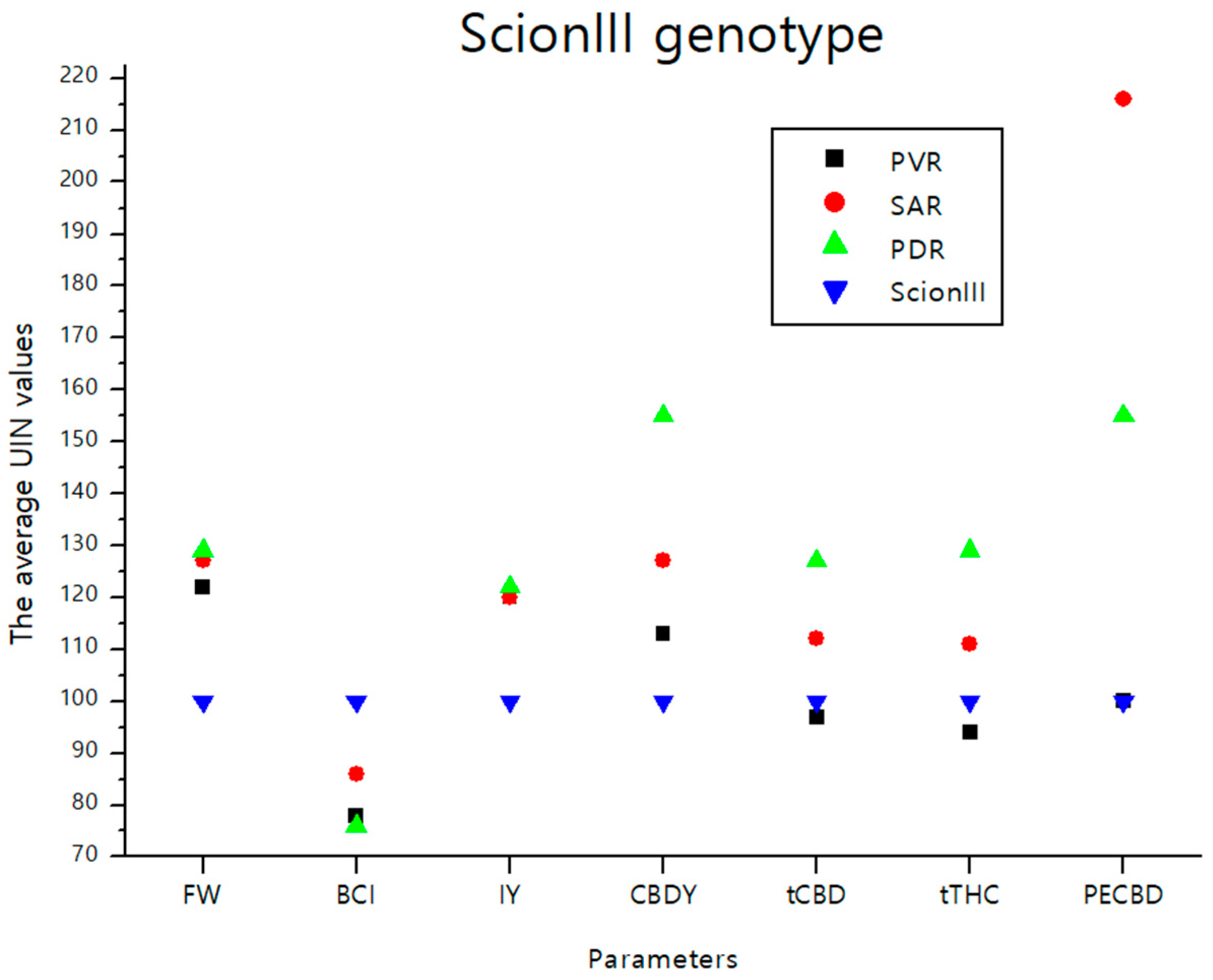
2.4.2. ‘ScionIII’ Genotype
3. Discussion
4. Materials and Methods
4.1. Cannabis Varieties
4.2. Rootstock Types and Scion Genotypes
4.3. Two-Step Grafting Method
4.4. Vegetative and Flowering Growing Conditions
4.5. Harvest Method
4.6. Analysis of Secondary Metabolites
4.6.1. Analysis of Cannabinoids
4.6.2. Terpene Analysis
4.7. Statistical Analysis
4.8. Efficiency of CBD Production
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Andre, C.M.; Hausman, J.F.; Guerriero, G. Cannabis sativa: The Plant of the Thousand and One Molecules. Front. Plant Sci. 2016, 7, 174167. [Google Scholar] [CrossRef] [PubMed]
- Taura, F.; Sirikantaramas, S.; Shoyama, Y.; Yoshikai, K.; Shoyama, Y.; Morimoto, S. Cannabidiolic-acid synthase, the chemotype-determining enzyme in the fiber-type Cannabis sativa. FEBS Lett. 2007, 581, 2929–2934. [Google Scholar] [CrossRef] [PubMed]
- Jin, D.; Dai, K.; Xie, Z. Secondary Metabolites Profiled in Cannabis Inflorescences, Leaves, Stem Barks, and Roots for Medicinal Purposes. Sci. Rep. 2020, 10, 3309. [Google Scholar] [CrossRef]
- Small, E.; Beckstead, H.D. Common Cannabinoid Phenotypes in 350 Stocks of Cannabis. Lloydia 1973, 36, 144–165. [Google Scholar]
- Turner, C.E.; ElSohly, M.A.; Cheng, P.C.; Lewis, G. Constituents of Cannabis sativa L., XIV: Intrinsic Problems in Classifying Cannabis Based on a Single Cannabinoid Analysis. J. Nat. Prod. 1979, 42, 481–490. [Google Scholar] [CrossRef]
- De Meijer, E.P.M.; Bagatta, M.; Carboni, A.; Crucitti, P.; Moliterni, V.M.C.; Ranalli, P.; Mandolino, G. The Inheritance of Chemical Phenotype in Cannabis sativa L. Genetics 2003, 163, 335–346. [Google Scholar] [CrossRef]
- Mandolino, G.; Bagatta, M.; Carboni, A.; Ranalli, P.; de Meijer, E. Qualitative and Quantitative Aspects of the Inheritance of Chemical Phenotype in Cannabis. J. Ind. Hemp 2003, 8, 52–72. [Google Scholar] [CrossRef]
- Fournier, G.; Richez-Dumanois, C.; Duvezin, J.; Mathieu, J.P.; Paris, M. Identification of a New Chemotype in Cannabis sativa: Cannabigerol-Dominant Plants, Biogenetic and Agronomic Prospects. Planta Medica 1987, 53, 277–280. [Google Scholar] [CrossRef]
- Magagnini, G.; Grassi, G.; Kotiranta, S. The Effect of Light Spectrum on the Morphology and Cannabinoid Content of Cannabis sativa L. Med. Cannabis Cannabinoids 2018, 1, 19–27. [Google Scholar] [CrossRef]
- Warren, G.S. Regulating Pot to Save the Polar Bear: Energy and Climate Impacts of the Marijuana Industry. Columbia J. Environ. Law 2015, 40, 385. [Google Scholar]
- Mills, E. The Carbon Footprint of Indoor Cannabis Production. Energy Policy 2012, 46, 58–67. [Google Scholar] [CrossRef]
- Flajšman, M.; Slapnik, M.; Murovec, J. Production of Feminized Seeds of High CBD Cannabis sativa L. by Manipulation of Sex Expression and Its Application to Breeding. Front. Plant Sci. 2021, 12, 718092. [Google Scholar] [CrossRef] [PubMed]
- de Andrés, E.F.; Sánchez, F.J.; Catalán, G.; Tenorio, J.L.; Ayerbe, L. Vegetative Propagation of Colutea istria Mill. from Leafy Stem Cuttings. Agrofor. Syst. 2004, 63, 7–14. [Google Scholar] [CrossRef]
- Hinesley, L.E.; Blazich, F.A. Influence of Post-Severance Combinations on the Rooting Capacity of Fraser Fir Stem Cuttings. Can. J. For. Res. 1981, 11, 316–323. [Google Scholar] [CrossRef]
- Dorrell, D.G. Vegetative Propagation of Flax by Stem Cuttings. Can. J. Plant Sci. 1974, 54, 197–201. [Google Scholar] [CrossRef]
- Munoz-Gutierrez, L.; Vargas-Hernández, J.J.; Lopez-Upton, J.; Soto-Hernandez, M. Effect of Cutting Age and Substrate Temperature on Rooting of Taxus globosa. New For. 2009, 38, 187–196. [Google Scholar] [CrossRef]
- Johnson, E.P.; Preece, J.E.; Aradhya, M.; Gradziel, T. Rooting Response of Prunus Wild Relative Semi-Hardwood Cuttings to Indole-3-Butyric Acid Potassium Salt (KIBA). Sci. Hortic. 2020, 263, 109144. [Google Scholar] [CrossRef]
- Salehi-Mohammadi, R.; Khasi, A.; Lee, S.G.; Huh, Y.C.; Lee, J.M.; Delshad, M. Assessing Survival and Growth Performance of Iranian Melon to Grafting onto Cucurbita Rootstocks. Korean J. Hortic. Sci. Technol. 2009, 27, 1–6. [Google Scholar]
- Lee, J.M.; Oda, M. Grafting of Herbaceous Vegetable and Ornamental Crops. Hortic. Rev. 2002, 28, 61–124. [Google Scholar] [CrossRef]
- Jang, Y.; Yang, E.; Cho, M. Effect of Grafting on Growth and Incidence of Phytophthora Blight and Bacterial Wilt of Pepper (Capsicum annuum L.). Hortic. Environ. Biotechnol. 2012, 53, 9–19. [Google Scholar] [CrossRef]
- Trionfetti Nisini, P.; Buzi, A.; Granati, E.; Chilosi, G.; Crinò, P.; Magro, P. Screening for Resistance to Didymella Bryoniae in Rootstocks of Melon. Bulletin 2008, 30, 231–234. [Google Scholar] [CrossRef]
- Huang, W.; Liao, S.; Lv, H.; Khaldun, A.B.M.; Wang, Y. Characterization of the Growth and Fruit Quality of Tomato Grafted on a Woody Medicinal Plant, Lycium chinense. Sci. Hortic. 2015, 197, 447–453. [Google Scholar] [CrossRef]
- Davis, A.R.; Perkins-Veazie, P.; Hassell, R.; Levi, A.; King, S.R.; Zhang, X. Grafting Effects on Vegetable Quality. HortScience 2008, 43, 1670–1672. [Google Scholar] [CrossRef]
- Kacjan-Marsic, N.; Osvald, J. The Influence of Grafting on Yield of Two Tomato Cultivars (Lycopersicon Esculentum Mill.) Grown in a Plastic House. Acta Agric. Slov. 2004, 83, 243–249. [Google Scholar] [CrossRef]
- Albrecht, U.; Bordas, M.; Lamb, B.; Meyering, B.; Bowman, K.D. Influence of Propagation Method on Root Architecture and Other Traits of Young Citrus Rootstock Plants. HortScience 2017, 52, 1569–1576. [Google Scholar] [CrossRef]
- Anderson, J.M.; Ingram, J.S.I. Tropical Soil Biology and Fertility: A Handbook of Methods. Soil Sci. 1994, 157, 265. [Google Scholar] [CrossRef]
- Lee, J.M.; Kubota, C.; Tsao, S.J.; Bie, Z.; Echevarria, P.H.; Morra, L.; Oda, M. Current Status of Vegetable Grafting: Diffusion, Grafting Techniques, Automation. Sci. Hortic. 2010, 127, 93–105. [Google Scholar] [CrossRef]
- Rivero, R.M.; Ruiz, J.M.; Romero, L. Role of Grafting in Horticultural Plants Under Stress Conditions. Food Agric. Environ. 2003, 1, 70–74. [Google Scholar]
- Bausher, M.G. Grafting Technique to Eliminate Rootstock Suckering of Grafted Tomatoes. HortScience 2011, 46, 596–598. [Google Scholar] [CrossRef]
- Gisbert, C.; Prohens, J.; Nuez, F. Performance of Eggplant Grafted onto Cultivated, Wild, and Hybrid Materials of Eggplant and Tomato. Int. J. Plant Prod. 2011, 5, 367–380. [Google Scholar] [CrossRef]
- Colla, G.; Rouphael, Y.; Cardarelli, M.; Temperini, O.; Rea, E.; Salerno, A.; Pierandrei, F. Influence of Grafting on Yield and Fruit Quality of Pepper (Capsicum annuum L.) Grown Under Greenhouse Conditions. Acta Hortic. 2008, 782, 359–364. [Google Scholar] [CrossRef]
- Mulatya, J.M.; Wilson, J.; Ong, C.K.; Deans, J.D.; Sprent, J.I. Root Architecture of Provenances, Seedlings and Cuttings of Melia Volkensii: Implications for Crop Yield in Dryland Agroforestry. Agrofor. Syst. 2002, 56, 65–72. [Google Scholar] [CrossRef]
- Riedacker, A.; Belgrand, M. Morphogenesis of Root Systems of Seedlings and Cuttings of Quercus robur L. Plant Soil 1983, 71, 131–146. [Google Scholar] [CrossRef]
- Sasse, J.; Sands, R. Configuration and development of root systems of cuttings and seedlings of Eucalyptus globulus. New For. 1997, 14, 85–105. [Google Scholar] [CrossRef]
- Purdy, S.J.; Hewavitharana, A.K.; Azman Halimi, R.; Magner, N.J.; Peterswald, T.J.; Trebilco, A.; Kretzschmar, T.; Hailstones, D.A. One-Step Grafting Methodology Can Adjust Stem Morphology and Increase THCA Yield in Medicinal Cannabis. Agronomy 2022, 12, 852. [Google Scholar] [CrossRef]
- Soltan, M.M.; ElAidy, F.A.; Scheerens, J.C.; Kleinhenz, M.D. Grafting, Scion and Rootstock Effects on Survival Rate, Vegetative Growth, and Fruit Yield of High Tunnel-Grown Grafted Pepper (Capsicum annuum L.) Plants. Adv. Crop Sci. Technol. 2017, 5, 312. [Google Scholar] [CrossRef]
- Condurso, C.; Verzera, A.; Dima, G.; Tripodi, G.; Crinò, P.; Paratore, A.; Romano, D. Effects of Different Rootstocks on Aroma Volatile Compounds and Carotenoid Content of Melon Fruits. Sci. Hortic. 2012, 148, 9–16. [Google Scholar] [CrossRef]
- Krumbein, A.; Schwarz, D. Grafting: A Possibility to Enhance Health-Promoting and Flavour Compounds in Tomato Fruits of Shaded Plants? Sci. Hortic. 2013, 149, 97–107. [Google Scholar] [CrossRef]
- Deng, W.W.; Han, J.; Fan, Y.; Tai, Y.; Zhu, B.; Lu, M.; Wang, R.; Wan, X.; Zhang, Z.Z. Uncovering Tea-Specific Secondary Metabolism Using Transcriptomic and Metabolomic Analyses in Grafts of Camellia sinensis and C. oleifera. Tree Genet. Genomes 2018, 14, 23. [Google Scholar] [CrossRef]
- Gisbert-Mullor, R.; Ceccanti, C.; Gara Padilla, Y.; López-Galarza, S.; Calatayud, Á.; Conte, G.; Guidi, L. Effect of Grafting on the Production, Physico-Chemical Characteristics, and Nutritional Quality of Fruit from Pepper Landraces. Antioxidants 2020, 9, 501. [Google Scholar] [CrossRef]
- Rogers, W. Regression standard errors in clustered samples. Stata Tech. Bull. 1994, 3, 19–23. [Google Scholar]
- Altamura, M. Root Histogenesis in Herbaceous and Woody Explants Cultured in Vitro: A Critical Review. Agronomie 1996, 16, 589–602. [Google Scholar] [CrossRef]
- Husen, A.; Pal, M. Variation in Shoot Anatomy and Rooting Behaviour of Stem Cuttings in Relation to Age of Donor Plants in Teak (Tectona Grandis Linn. F.). New For. 2006, 31, 57–73. [Google Scholar] [CrossRef]
- Ofori, D.A.; Newton, A.C.; Leakey, R.R.B.; Grace, J. Vegetative Propagation of Milicia Excelsa by Leafy Stem Cuttings: Effects of Auxin Concentration, Leaf Area and Rooting Medium. For. Ecol. Manag. 1996, 84, 39–48. [Google Scholar] [CrossRef]
- Zheng, Y. (Ed.) Handbook of Cannabis Production in Controlled Environments, 1st ed.; CRC Press: Boca Raton, FL, USA, 2022; pp. 100–105. [Google Scholar] [CrossRef]
- King, S.R.; Davis, A.R.; Zhang, X.; Crosby, K. Genetics, Breeding and Selection of Rootstocks for Solanaceae and Cucurbitaceae. Sci. Hortic. 2010, 127, 106–111. [Google Scholar] [CrossRef]
- Schwarz, D.; Rouphael, Y.; Colla, G.; Venema, J.H. Grafting as a Tool to Improve Tolerance of Vegetables to Abiotic Stresses: Thermal Stress, Water Stress and Organic Pollutants. Sci. Hortic. 2010, 127, 162–171. [Google Scholar] [CrossRef]
- Gong, T.; Brecht, J.K.; Koch, K.E.; Hutton, S.F.; Zhao, X. A Systematic Assessment of How Rootstock Growth Characteristics Impact Grafted Tomato Plant Biomass, Resource Partitioning, Yield, and Fruit Mineral Composition. Front. Plant Sci. 2022, 13, 948656. [Google Scholar] [CrossRef] [PubMed]
- Ulas, F. Response of Different Rootstocks on Vegetative Growth, Fruit and Seed Yield of Eggplant (Solanum Melongena L.). Genetika 2021, 53, 593–608. [Google Scholar] [CrossRef]
- Babaj, I.; Sallaku, G.; Balliu, A. The Effects of Endogenous Mycorrhiza (Glomus spp.) on Plant Growth and Yield of Grafted Cucumber (Cucumis sativum L) Under Common Commercial Greenhouse Conditions. Albanian J. Agric. Sci. 2014, 13, 24–28. [Google Scholar]
- Bautista, S.A.; Calatayud, A.; Nebauer, S.G.; Pascual, B.; Vicente Maroto, J.; López-Galarza, S. Effects of Simple and Double Grafting Melon Plants on Mineral Absorption, Photosynthesis, Biomass, and Yield. Sci. Hortic. 2011, 130, 575–580. [Google Scholar] [CrossRef]
- Wang, H.; Inukai, Y.; Yamauchi, A. Root Development and Nutrient Uptake. Crit. Rev. Plant Sci. 2006, 25, 279–301. [Google Scholar] [CrossRef]
- De Dorlodot, S.; Forster, B.; Pages, L.; Price, A.; Tuberosa, R.; Draye, X. Root System Architecture: Opportunities and Constraints for Genetic Improvement of Crops. Trends Plant Sci. 2007, 12, 474–481. [Google Scholar] [CrossRef]
- Bailey, P.H.J.; Currey, J.D.; Fitter, A.H. The Role of Root System Architecture and Root Hairs in Promoting Anchorage Against Uprooting Forces in Allium cepa and Root Mutants of Arabidopsis thaliana. J. Exp. Bot. 2002, 53, 333–1340. [Google Scholar] [CrossRef]
- Turhan, A.; Ozmen, N.; Serbeci, M.S.; Seniz, V. Effects of Grafting on Different Rootstocks on Tomato Fruit Yield and Quality. Hortic. Sci. 2011, 38, 142–149. [Google Scholar] [CrossRef]
- Eissenstat, D.; Achor, D. Anatomical Characteristics of Roots of Citrus Rootstocks that Vary in Specific Root Length. New Phytol. 1999, 141, 309–321. [Google Scholar] [CrossRef]
- Albacete, A.; Martínez-Andújar, C.; Pérez-Alfocea, F. Hormonal and Metabolic Regulation of Source-Sink Relations Under Salinity and Drought: From Plant Survival to Crop Yield Stability. Biotechnol. Adv. 2014, 32, 12–30. [Google Scholar] [CrossRef]
- Doñas-Uclés, F.; Pérez-Madrid, D.; Amate-Llobregat, C.; Rodríguez-García, E.M.; Camacho-Ferre, F. Production of Pepper Cultivar Palermo Grafted onto Serrano de Morelos 2, Jalapeño, and Three Commercial Rootstocks. HortScience 2015, 50, 1018–1022. [Google Scholar] [CrossRef]
- Cantero-Navarro, E.; Romero-Aranda, R.; Fernández-Muñoz, R.; Martínez-Andújar, C.; Pérez-Alfocea, F.; Albacete, A. Improving Agronomic Water Use Efficiency in Tomato by Rootstock-Mediated Hormonal Regulation of Leaf Biomass. Plant Sci. 2016, 251, 90–100. [Google Scholar] [CrossRef]
- López-Marín, J.; Gálvez, A.; del Amor, F.M.; Albacete, A.; Fernández, J.A.; Egea-Gilabert, C.; Pérez-Alfocea, F. Selecting Vegetative/Generative/Dwarfing Rootstocks for Improving Fruit Yield and Quality in Water Stressed Sweet Peppers. Sci. Hortic. 2017, 214, 9–17. [Google Scholar] [CrossRef]
- Lordan, J.; Fazio, G.; Francescatto, P.; Robinson, T. Effects of Apple (Malus × domestica) Rootstocks on Scion Performance and Hormone Concentration. Sci. Hortic. 2017, 225, 96–105. [Google Scholar] [CrossRef]
- Soumelidou, K.; Battey, N.H.; John, P.; Barnett, J.R. The Anatomy of the Developing Bud Union and Its Relationship to Dwarfing in Apple. J. Hortic. Sci. 1994, 74, 605–611. [Google Scholar] [CrossRef]
- Kamboj, J.; Blake, P.S.; Quinlan, J.D. Identification and Quantitation by GC-MS of Zeatin and Zeatin Riboside in Xylem Sap from Rootstock and Scion of Grafted Apple Trees. Plant Growth Regul. 1999, 28, 199–205. [Google Scholar] [CrossRef]
- Soumelidou, K.; Morris, D.A.; Battey, N.H.; Barnett, J.R.; John, P. Auxin Transport Capacity in Relation to the Dwarfing Effect of Apple Rootstocks. J. Hortic. Sci. 1994, 69, 719–725. [Google Scholar] [CrossRef]
- Ruegger, M.; Dewey, E.; Hobbie, L.; Brown, D.; Bernasconi, P.; Turner, J.; Muday, G.; Estelle, M. Reduced Naphthylphthalamic Acid Binding in the tir3 Mutant of Arabidopsis is Associated with a Reduction in Polar Auxin Transport and Diverse Morphological Defects. Plant Cell 1997, 9, 745–757. [Google Scholar] [CrossRef]
- Aloni, B.; Karni, L.; Deventurero, G.; Levin, Z.; Cohen, R.; Katzir, N.; Lotan-Pompan, M.; Edelstein, M.; Aktas, H.; Turhan, E.; et al. Physiological and Biochemical Changes at the Rootstock-Scion Interface in Graft Combinations between Cucurbita Rootstocks and a Melon Scion. J. Hortic. Sci. Biotechnol. 2008, 83, 777–783. [Google Scholar] [CrossRef]
- Tworkoski, T.; Fazio, G. Hormone and Growth Interactions of Scions and Size-Controlling Rootstocks of Young Apple Trees. Plant Growth Regul. 2016, 78, 105–119. [Google Scholar] [CrossRef]
- Spiegelman, Z.; Ham, B.-K.; Zhang, Z.; Toal, T.W.; Brady, S.M.; Zheng, Y.; Fei, Z.; Lucas, W.J.; Wolf, S. A Tomato Phloem-Mobile Protein Regulates the Shoot-to-Root Ratio by Mediating the Auxin Response in Distant Organs. Plant J. 2015, 83, 853–863. [Google Scholar] [CrossRef]
- Wink, M. Plant Breeding: Importance of Plant Secondary Metabolites for Protection Against Pathogens and Herbivores. Theor. Appl. Genet. 1988, 75, 225–233. [Google Scholar] [CrossRef]
- Dong, D.; Shi, Y.-N.; Mou, Z.-M.; Chen, S.-Y.; Zhao, D.-K. Grafting: A Potential Method to Reveal the Differential Accumulation Mechanism of Secondary Metabolites. Hortic. Res. 2022, 9, uhac050. [Google Scholar] [CrossRef]
- Guan, W.J.; Zhao, X.; Hassell, R.; Thies, J. Defense Mechanisms Involved in Disease Resistance of Grafted Vegetables. HortScience 2012, 47, 164–170. [Google Scholar] [CrossRef]
- Dudareva, N.; Klempien, A.; Muhlemann, J.K.; Kaplan, I. Biosynthesis, Function and Metabolic Engineering of Plant Volatile Organic Compounds. New Phytol. 2013, 198, 16–32. [Google Scholar] [CrossRef]
- Frank, A.; Groll, M. The Methylerythritol Phosphate Pathway to Isoprenoids. Chem. Rev. 2017, 117, 5675–5703. [Google Scholar] [CrossRef]
- Sirikantaramas, S.; Taura, F.; Morimoto, S.; Shoyama, Y. Recent Advances in Cannabis sativa Research: Biosynthetic Studies and Its Potential in Biotechnology. Curr. Pharm. Biotechnol. 2007, 8, 237. [Google Scholar] [CrossRef]
- Fraser, P.D.; Bramley, P.M. The Biosynthesis and Nutritional Uses of Carotenoids. Prog. Lipid Res. 2004, 43, 228–265. [Google Scholar] [CrossRef]
- Roell, M.S. Terpenes in Cannabis: Solving the Puzzle of How to Predict Taste and Smell. Plant Physiol. 2020, 184, 8–9. [Google Scholar] [CrossRef]
- Pacifico, D.; Miselli, F.; Carboni, A.; Moschella, A.; Mandolino, G. Time Course of Cannabinoid Accumulation and Chemotype Development during the Growth of Cannabis sativa L. Euphytica 2008, 160, 231–240. [Google Scholar] [CrossRef]
- UNCTAD. Available online: https://unctad.org (accessed on 6 August 2023).
- Westmoreland, F.M.; Kusuma, P.; Bugbee, B. Cannabis Lighting: Decreasing Blue Photon Fraction Increases Yield but Efficacy Is More Important for Cost-Effective Production of Cannabinoids. PLoS ONE 2021, 16, e0248988. [Google Scholar] [CrossRef]
- Adhikary, D.; Kulkarni, M.; El-Mezawy, A.; Mobini, S.; Elhiti, M.; Gjuric, R.; Ray, A.; Polowick, P.; Slaski, J.J.; Jones, M.P.; et al. Medical Cannabis and Industrial Hemp Tissue Culture: Present Status and Future Potential. Front. Plant Sci. 2021, 12, 627240. [Google Scholar] [CrossRef]
- Kakabouki, I.; Kousta, A.; Folina, A.; Karydogianni, S.; Zisi, C.; Kouneli, C.; Papastylianou, P. Effect of Fertilization with Urea and Inhibitors on Growth, Yield and CBD Concentration of Hemp (Cannabis sativa L.). Sustainability 2021, 13, 2157. [Google Scholar] [CrossRef]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectrometry, 4th ed.; Allured Publications: Carol Stream, IL, USA, 2007; p. 804. [Google Scholar]
- Gallily, R.; Yekhtin, Z.; Hanuš, L.O. The anti-inflammatory properties of terpenoids from cannabis. Cannabis Cannabinoid Res. 2018, 3, 282–290. [Google Scholar] [CrossRef]
- The R Project for Statistical Computing. Available online: https://www.r-project.org/ (accessed on 30 January 2024).
- OriginLab. Available online: https://www.originlab.com/ (accessed on 30 January 2024).











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Material/ Combinations | Number of Rootstocks (NR) | Number of Successfully Rooted Rootstocks (RR) | Number of Survived Plants (SP) |
|---|---|---|---|
| ’Vigor-C23/ScionII’ | 13 | 7 | 5 |
| ’Vigor-S26/ScionII’ | 13 | 12 | 3 |
| ’Vigor-S39/ScionII’ | 13 | 9 | 5 |
| ’Dwarf-C45/ScionII’ | 13 | 6 | 2 |
| ’Dwarf-S63/ScionII’ | 13 | 6 | 4 |
| ’Dwarf-T34/ScionII’ | 13 | 6 | 3 |
| ’Seedl-TIB/ScionII’ | 10 | 10 | 4 |
| ’Seedl-FIO/ScionII’ | 10 | 10 | 7 |
| ’Seedl-SIM/ScionII’ | 10 | 10 | 6 |
| ’Control/ScionII’ | 12 | 5 | 5 |
| Plant Material/ Combinations | Number of Rootstocks (NR) | Successfully Rooted Rootstocks (RR) | Survived Plants (SP) |
|---|---|---|---|
| ’Vigor-C23/ScionIII’ | 12 | 5 | 3 |
| ’Vigor-S26/ScionIII’ | 12 | 11 | 3 |
| ’Vigor-S39/ScionIII’ | 13 | 8 | 2 |
| ’Dwarf-C45/ScionIII’ | 12 | 6 | 3 |
| ’Dwarf-S63/ScionIII’ | 12 | 6 | 3 |
| ’Dwarf-T34/ScionIII’ | 12 | 10 | 3 |
| ’Seedl-TIB/ScionIII’ | 10 | 10 | 4 |
| ’Seedl-FIO/ScionIII’ | 10 | 10 | 4 |
| ’Seedl-SIM/ScionIII’ | 10 | 10 | 5 |
| ’Control/ScionIII’ | 12 | 3 | 3 |
| Plant Material/Combination | BCI (g/cm ± SD) | p ≤ 0.05 1 |
|---|---|---|
| ‘Dwarf-C45/ScionII’ | 0.2960 ± 0.012 | a |
| ‘Dwarf-T34/ScionII’ | 0.2669 ± 0.042 | ab |
| ‘Dwarf-S63/ScionII’ | 0.2445 ± 0.053 | ab |
| ‘Vigor-S26/ScionII’ | 0.2209 ± 0.026 | ab |
| ‘Seedl-SIM/ScionII’ | 0.2092 ± 0.064 | abc |
| ‘Seedl-TIB/ScionII’ | 0.1980 ± 0.088 | bc |
| ‘Vigor-C23/ScionII | 0.1810 ± 0.070 | bc |
| ‘Vigor-S39/ScionII’ | 0.1792 ± 0.069 | bc |
| ‘Seedl-FIO/ScionII’ | 0.1766 ± 0.032 | bc |
| ‘Control/ScionII’ | 0.1225 ± 0.042 | c |
| Rootstock Type | Variety—Genotype | Height | Stem Thickness | Natural Branching |
|---|---|---|---|---|
| Vigorous | Carmagnola—C23 | High | High | Low |
| Simba—S26 | High | High | High | |
| Simba—S39 * | Mid | High | Low | |
| Merlot—MER * | Mid | High | High | |
| Dwarfing | Carmagnola—C45 | Low | Low | Low |
| Simba—S63 | Low | Low | High | |
| Tiborszallasi—T34 | Low | Low | High | |
| Seedlings | ‘Tiborszallasi’ seedlings | Specific characteristics of the variety | ||
| ‘Fiona’ seedlings | ||||
| ‘Simba’ seedlings | ||||
| Potentially Vigorous Rootstock (PVR) | Potentially Dwarfing Rootstock (PDR) | Seedling-as-Rootstock (SAR) | ||||||
|---|---|---|---|---|---|---|---|---|
| Rootstock Genotype | Scion Genotype | Combination Name | Rootstock Genotype | Scion Genotype | Combination Name | Rootstock Genotype | Scion Genotype | Combination Name |
| C23 | ScionII | Vigor-C23/ScionII | C45 | ScionII | Dwarf-C45/ScionII | Tiborszallasi seedlings | ScionII | Seedl-TIB/ScionII |
| S26 | Vigor-S26/ScionII | S63 | Dwarf-S63/ScionII | Fiona seedlings | Seedl-FIO/ScionII | |||
| S39 | Vigor-S39/ScionII | T34 | Dwarf-T34/ScionII | Simba seedlings | Seedl-SIM/ScionII | |||
| C23 | ScionIII | Vigor-C23/ScionIII | C45 | ScionIII | Dwarf-C45/ScionIII | Tiborszallasi seedlings | ScionIII | Seedl-TIB/ScionIII |
| S26 | Vigor-S26/ScionIII | S63 | Dwarf-S63/ScionIII | Fiona seedlings | Seedl-FIO/ScionIII | |||
| MER | Vigor-MER/ScionIII | T34 | Dwarf-T34/ScionIII | Simba seedlings | Seedl-SIM/ScionIII | |||
| Control combinations | ||||||||
| Control/ScionII: non-grafted clones of ScionII scion genotype | ||||||||
| Control/ScionIII: non-grafted clones of ScionIII scion genotype | ||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bitežnik, L.; Štukelj, R.; Flajšman, M. The Efficiency of CBD Production Using Grafted Cannabis sativa L. Plants Is Highly Dependent on the Type of Rootstock: A Study. Plants 2024, 13, 1117. https://doi.org/10.3390/plants13081117
Bitežnik L, Štukelj R, Flajšman M. The Efficiency of CBD Production Using Grafted Cannabis sativa L. Plants Is Highly Dependent on the Type of Rootstock: A Study. Plants. 2024; 13(8):1117. https://doi.org/10.3390/plants13081117
Chicago/Turabian StyleBitežnik, Luka, Roman Štukelj, and Marko Flajšman. 2024. "The Efficiency of CBD Production Using Grafted Cannabis sativa L. Plants Is Highly Dependent on the Type of Rootstock: A Study" Plants 13, no. 8: 1117. https://doi.org/10.3390/plants13081117