Towards the Physics of Calcium Signalling in Plants

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
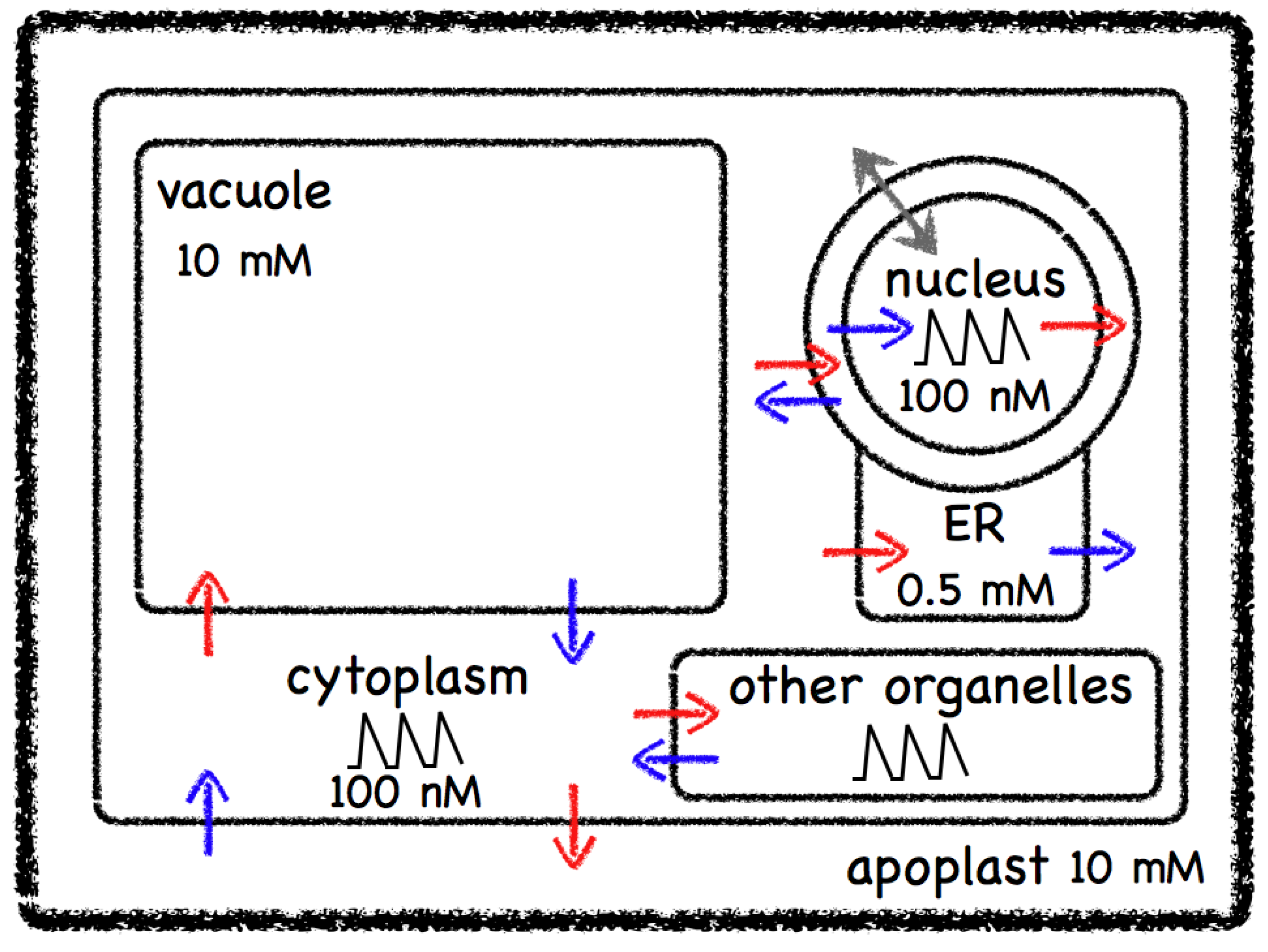
2. Calcium by Numbers
2.1. Calcium Biochemistry

2.2. Calcium Maths

 and
and  represent the sum of all fluxes out of and into the compartment respectively. Describing the changes in different compartments, results in a system of such equations. In some cases it is useful to model explicitly the time lapse before changes occur, such as in gene translation, and this can be accounted for using a delay parameter, leading to a variant of the above approach known as delay differential equations (DDEs). Spatial models are based on partial differential equations (PDEs) and include the additional term,
represent the sum of all fluxes out of and into the compartment respectively. Describing the changes in different compartments, results in a system of such equations. In some cases it is useful to model explicitly the time lapse before changes occur, such as in gene translation, and this can be accounted for using a delay parameter, leading to a variant of the above approach known as delay differential equations (DDEs). Spatial models are based on partial differential equations (PDEs) and include the additional term,  ,
,

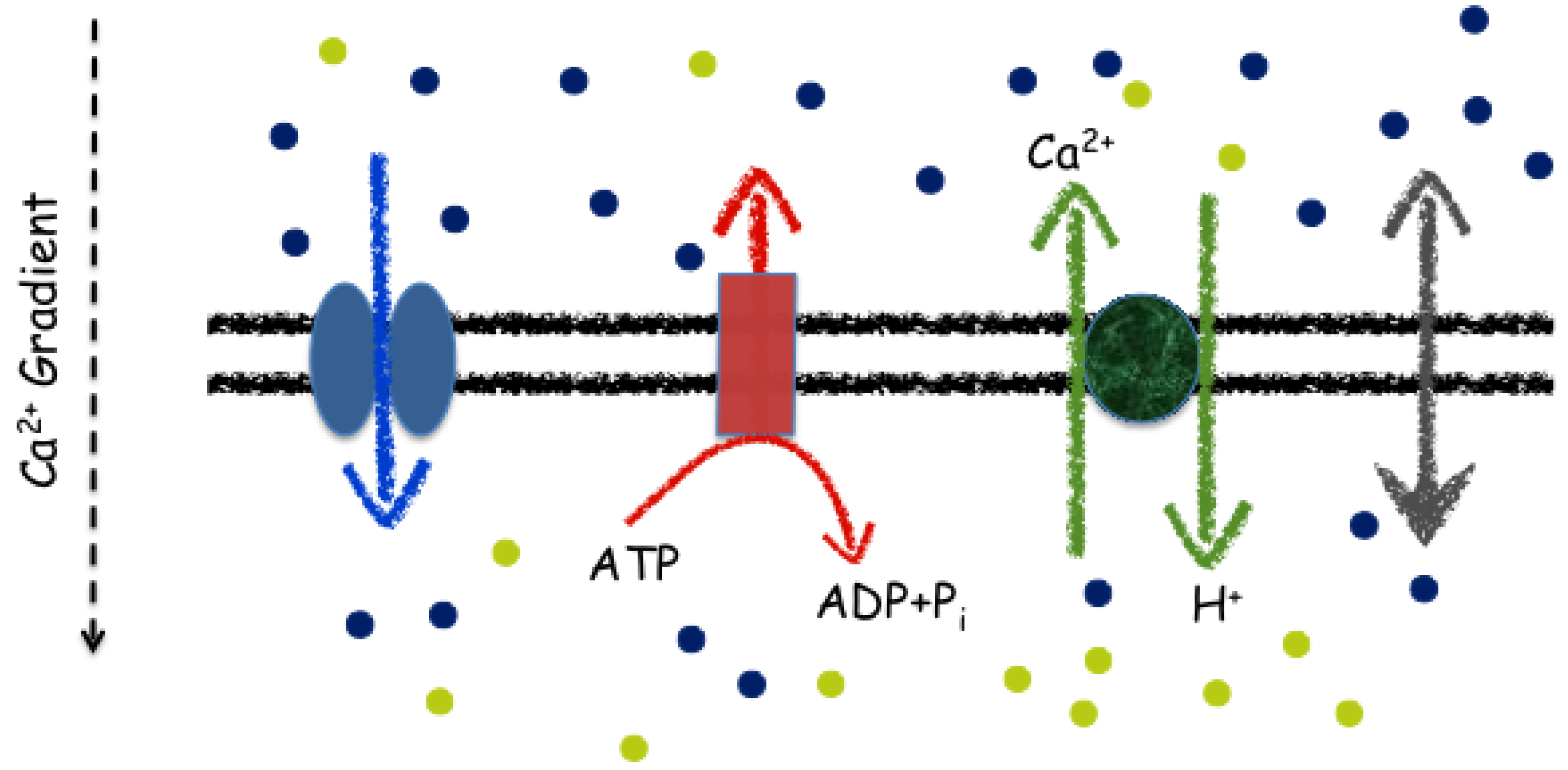
3. Models for the Calcium Machinery





3.1. Fluxes: Ligand- and Voltage-Gated Calcium Release Channels
3.2. Gating in Steps
3.3. The Basic Machinery: Autonomous Regulation of Calcium Levels in Organelles
4. Calcium and Temperature
4.1. The Search for Primary Temperature-Sensing Events
4.2. The Many Elusive Effects of Temperature
5. Calcium and Symbiosis

 . This describes a single compartment
. This describes a single compartment  , denoted l, with two elements, b and c, on its surface (perhaps representing membrane embedded proteins), four atoms of d within the compartment and two atoms of α outside the compartment (representing individual molecules, enzymes or other biologically relevant components within the bulk). An example rule would be
, denoted l, with two elements, b and c, on its surface (perhaps representing membrane embedded proteins), four atoms of d within the compartment and two atoms of α outside the compartment (representing individual molecules, enzymes or other biologically relevant components within the bulk). An example rule would be




6. Calcium and Polar Growth



7. Calcium and Stomata

8. Calcium and the Circadian Clock
9. Calcium and Systemic Responses

10. Calcium and Specificity
10.1. Inferring Calcium Signatures from Calcium Time Series
10.2. Different Stimuli Alter the Functioning of Transporters and Buffers
10.3. The Experimentally Recorded Signature May Be Very Different from the Relevant One
10.4. Finite Oscillatory Signals Are Optimally Decoded by Specific Proteins
11. Conclusions: Calcium and the Experimental-Modelling Cycle—Past, Present and Future
Supplementary Files
Supplementary File 1Acknowledgments
Conflicts of Interest
References
- Morr, S.; Cuartas, E.; Alwattar, B.; Lane, J.M. How much calcium is in your drinking water? A survey of calcium concentrations in bottled and tap water and their significance for medical treatment and drug administration. HSS J. 2006, 2, 130–135. [Google Scholar] [CrossRef]
- Katz, A.K.; Glusker, J.P.; Beebe, S.A.; Bock, C.W. Calcium ion coordination: A comparison with that of beryllium, magnesium, and zinc. J. Am. Chem. Soc. 1996, 118, 5752–5763. [Google Scholar] [CrossRef]
- Williamson, R.E. Cytoplasmic streaming in Chara: A cell model activated by ATP and inhibited by cytochalasin B. J. Cell Sci. 1975, 17, 655–668. [Google Scholar]
- Tazawa, M.; Kikuyama, M.; Shimmen, T. Electrical characteristics and cytoplasmic streaming of Characeae cells lacking tonoplast. Cell Struct. Funct. 1976, 1, 165–176. [Google Scholar] [CrossRef]
- Leinonen, R.; Diez, F.G.; Binns, D.; Fleischmann, W.; Lopez, R.; Apweiler, R. UniProt archive. Bioinformatics 2004, 20, 3236–3237. [Google Scholar] [CrossRef]
- Huala, E.; Dickerman, A.W.; Garcia-Hernandez, M.; Weems, D.; Reiser, L.; LaFond, F.; Hanley, D.; Kiphart, D.; Zhuang, M.; Huang, W.; et al. The Arabidopsis Information Resource (TAIR): A comprehensive database and web-based information retrieval, analysis, and visualization system for a model plant. Nucleic Acids Res. 2001, 29, 102–105. [Google Scholar] [CrossRef]
- Webb, A.A.R.; McAinsh, M.R.; Taylor, J.E.; Hetherington, A.M. Calcium ions as intracellular second messengers in higher plants. Adv. Bot. Res. 1996, 22, 45–96. [Google Scholar] [CrossRef]
- Dodd, A.N.; Kudla, J.; Sanders, D. The language of calcium signaling. Annu. Rev. Plant Biol. 2010, 61, 593–620. [Google Scholar] [CrossRef]
- Sanders, D.; Pelloux, J.; Brownlee, C.; Harper, J.F. Calcium at the crossroads of signaling. Plant Cell 2002, 14, S401–S417. [Google Scholar]
- Berridge, M.J.; Lipp, P.; Bootman, M.D. The versatility and universality of calcium signalling. Nat. Rev. Mol. Cell Biol. 2000, 1, 11–21. [Google Scholar] [CrossRef]
- Clapham, D.E. Calcium signaling. Cell 2007, 131, 1047–1058. [Google Scholar] [CrossRef]
- Kummer, U.; Krajnc, B.; Pahle, J.; Green, A.K.; Dixon, C.J.; Marhl, M. Transition from stochastic to deterministic behavior in calcium oscillations. Biophys. J. 2005, 89, 1603–1611. [Google Scholar] [CrossRef]
- Schuster, S.; Marhl, M.; Hofer, T. Modelling of simple and complex calcium oscillations. From single-cell responses to intercellular signalling. Eur. J. Biochem. 2002, 269, 1333–1355. [Google Scholar] [CrossRef]
- Gillespie, D.T. Exact stochastic simulation of coupled chemical-reactions. J. Phys. Chem. Us 1977, 81, 2340–2361. [Google Scholar] [CrossRef]
- Hodgkin, A.L.; Huxley, A.F. A quantitative description of membrane current and its application to conduction and excitation in nerve. J. Physiol. 1952, 117, 500–544. [Google Scholar]
- Hill, A.V. The possible effects of the aggregation of the molecules of haemoglobin on its dissociation curves. J. Physiol. 1910, 40, iv–vii. [Google Scholar]
- Carafoli, E. The unusual history and unique properties of the calcium signal. New Comp. Biochem. 2007, 41, 3–22. [Google Scholar] [CrossRef]
- Bauer, C.S.; Plieth, C.; Bethmann, B.; Popescu, O.; Hansen, U.P.; Simonis, W.; Schonknecht, G. Strontium-induced repetitive calcium spikes in a unicellular green alga. Plant Physiol. 1998, 117, 545–557. [Google Scholar] [CrossRef]
- Schönknecht, G.; Bauer, C. Modelling Ca2+ Oscillations in Plants. In Rhythms in Plants: Phenomenology, Mechanisms, and Adaptive Significance; Mancuso, S., Shabala, S., Eds.; Springer: Wien, Austria, New York, NY, USA, 2007; pp. 295–311. [Google Scholar]
- Tidow, H.; Poulsen, L.R.; Andreeva, A.; Knudsen, M.; Hein, K.L.; Wiuf, C.; Palmgren, M.G.; Nissen, P. A bimodular mechanism of calcium control in eukaryotes. Nature 2012, 491, 468–472. [Google Scholar] [CrossRef]
- Charpentier, M.; Vaz Martins, T.; Granqvist, E.; Oldroyd, G.E.; Morris, R.J. The role of DMI1 in establishing Ca2+ oscillations in legume symbioses. Plant Signal. Behav. 2013, 8. [Google Scholar]
- Granqvist, E.; Wysham, D.; Hazledine, S.; Kozlowski, W.; Sun, J.; Charpentier, M.; Martins, T.V.; Haleux, P.; Tsaneva-Atanasova, K.; Downie, J.A.; et al. Buffering capacity explains signal variation in symbiotic calcium oscillations. Plant Physiol. 2012, 160, 2300–2310. [Google Scholar] [CrossRef]
- Stucki, J.W.; Somogyi, R. A dialog on Ca2+ oscillations—An attempt to understand the essentials of mechanisms leading to hormone-induced intracellular Ca2+ oscillations in various kinds of cell on a theoretical level. Bba Bioenerg. 1994, 1183, 453–472. [Google Scholar] [CrossRef]
- Ane, J.M.; Kiss, G.B.; Riely, B.K.; Penmetsa, R.V.; Oldroyd, G.E.D.; Ayax, C.; Levy, J.; Debelle, F.; Baek, J.M.; Kalo, P.; et al. Medicago truncatula DMI1 required for bacterial and fungal symbioses in legumes. Science 2004, 303, 1364–1367. [Google Scholar] [CrossRef]
- Buschmann, P.; Gradmann, D. Minimal model for oscillations of membrane voltage in plant cells. J. Theor. Biol. 1997, 188, 323–332. [Google Scholar] [CrossRef]
- Glansdorff, P.; Prigogine, I. Thermodynamic Theory of Structure, Stability and Fluctuations; Wiley-Interscience: London, UK, New York, NY, USA, 1971. [Google Scholar]
- Venkateshwaran, M.; Cosme, A.; Han, L.; Banba, M.; Satyshur, K.A.; Schleiff, E.; Parniske, M.; Imaizumi-Anraku, H.; Ane, J.M. The recent evolution of a symbiotic ion channel in the legume family altered ion conductance and improved functionality in calcium signaling. Plant Cell 2012, 24, 2528–2545. [Google Scholar] [CrossRef]
- Rea, P.A.; Britten, C.J.; Jennings, I.R.; Calvert, C.M.; Skiera, L.A.; Leigh, R.A.; Sanders, D. Regulation of vacuolar H+-pyrophosphatase by free calcium—A reaction kinetic-analysis. Plant Physiol. 1992, 100, 1706–1715. [Google Scholar] [CrossRef]
- Gradmann, D.; Johannes, E.; Hansen, U. Kinetic analysis of Ca2+/K+ selectivity of an ion channel by single-binding-site models. J. Membr. Biol. 1997, 159, 169–178. [Google Scholar] [CrossRef]
- Allen, G.J.; Sanders, D.; Gradmann, D. Calcium-potassium selectivity: Kinetic analysis of current-voltage relationships of the open, slowly activating channel in the vacuolar membrane of Vicia faba guard-cells. Planta 1998, 204, 528–541. [Google Scholar] [CrossRef]
- Hansen, U.P.; Keunecke, M.; Blunck, R. Gating and permeation models of plant channels. J. Exp. Bot. 1997, 48, 365–382. [Google Scholar] [CrossRef]
- Plieth, C.; Hansen, U.P. Cytosolic Ca2+ and H+ buffers in green algae: A reply. Protoplasma 1998, 203, 210–213. [Google Scholar] [CrossRef]
- Pottosin, I.I.; Martinez-Estevez, M.; Dobrovinskaya, O.R.; Muniz, J.; Schönknecht, G. Mechanism of luminal Ca2+ and Mg2+ action on the vacuolar slowly activating channels. Planta 2004, 219, 1057–1070. [Google Scholar] [CrossRef]
- Perez, V.; Wherrett, T.; Shabala, S.; Muniz, J.; Dobrovinskaya, O.; Pottosin, I. Homeostatic control of slow vacuolar channels by luminal cations and evaluation of the channel-mediated tonoplast Ca2+ fluxes in situ. J. Exp. Bot. 2008, 59, 3845–3855. [Google Scholar] [CrossRef]
- Peiter, E. The plant vacuole: Emitter and receiver of calcium signals. Cell Calcium. 2011, 50, 120–128. [Google Scholar] [CrossRef]
- Isayenkov, S.; Isner, J.C.; Maathuis, F.J. Vacuolar ion channels: Roles in plant nutrition and signalling. FEBS Lett. 2010, 584, 1982–1988. [Google Scholar] [CrossRef]
- Pottosin, I.I.; Schönknecht, G. Vacuolar calcium channels. J. Exp. Bot. 2007, 58, 1559–1569. [Google Scholar] [CrossRef]
- Dobrovinskaya, O.R.; Muniz, J.; Pottosin, I.I. Asymmetric block of the plant vacuolar Ca(2+)-permeable channel by organic cations. Eur. Biophys. J. 1999, 28, 552–563. [Google Scholar] [CrossRef]
- Kaplan, B.; Sherman, T.; Fromm, H. Cyclic nucleotide-gated channels in plants. FEBS Lett. 2007, 581, 2237–2246. [Google Scholar] [CrossRef]
- Zelman, A.K.; Dawe, A.; Gehring, C.; Berkowitz, G.A. Evolutionary and structural perspectives of plant cyclic nucleotide-gated cation channels. Front. Plant Sci. 2012. [Google Scholar] [CrossRef]
- Craven, K.B.; Zagotta, W.N. CNG and HCN channels: Two peas, one pod. Annu. Rev. Physiol. 2006, 68, 375–401. [Google Scholar] [CrossRef]
- Oldroyd, G.E. Speak, friend, and enter: Signalling systems that promote beneficial symbiotic associations in plants. Nat. Rev. Microbiol. 2013, 11, 252–263. [Google Scholar] [CrossRef]
- Sieberer, B.J.; Chabaud, M.; Fournier, J.; Timmers, A.C.; Barker, D.G. A switch in Ca2+ spiking signature is concomitant with endosymbiotic microbe entry into cortical root cells of Medicago truncatula. Plant J. 2012, 69, 822–830. [Google Scholar] [CrossRef]
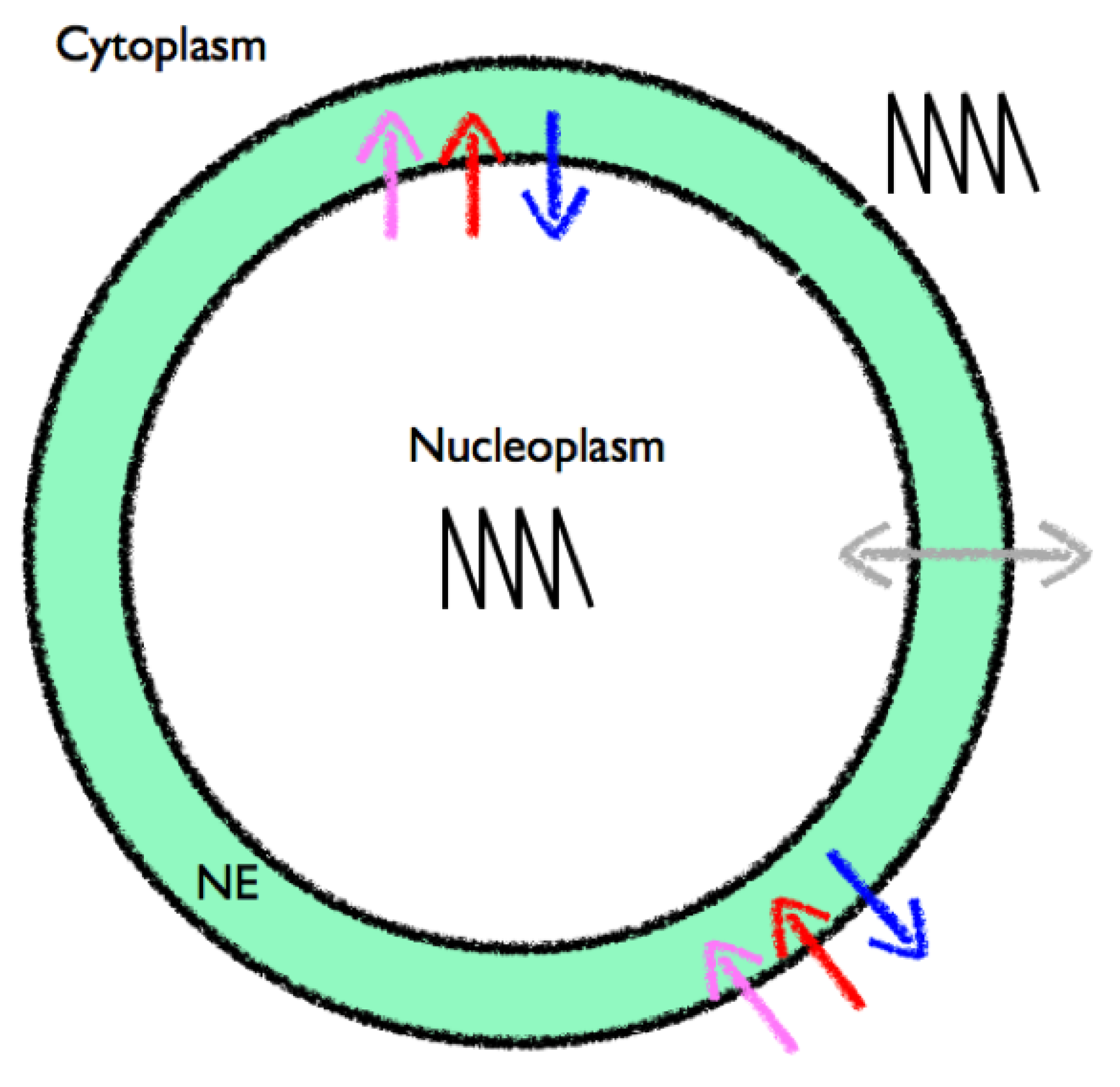
- Bootman, M.D.; Fearnley, C.; Smyrnias, I.; MacDonald, F.; Roderick, H.L. An update on nuclear calcium signalling. J. Cell Sci. 2009, 122, 2337–2350. [Google Scholar]
- Gerasimenko, O.; Gerasimenko, J. New aspects of nuclear calcium signalling. J. Cell Sci. 2004, 117, 3087–3094. [Google Scholar] [CrossRef]
- Mazars, C.; Bourque, S.; Mithofer, A.; Pugin, A.; Ranjeva, R. Calcium homeostasis in plant cell nuclei. New Phytol. 2009, 181, 261–274. [Google Scholar] [CrossRef]
- Charpentier, M.; Oldroyd, G.E. Nuclear calcium signaling in plants. Plant Physiol. 2013. [Google Scholar] [CrossRef]
- Xiong, T.C.; Jauneau, A.; Ranjeva, R.; Mazars, C. Isolated plant nuclei as mechanical and thermal sensors involved in calcium signalling. Plant J. 2004, 40, 12–21. [Google Scholar] [CrossRef]
- Briere, C.; Xiong, T.C.; Mazars, C.; Ranjeva, R. Autonomous regulation of free Ca2+ concentrations in isolated plant cell nuclei: A mathematical analysis. Cell Calcium. 2006, 39, 293–303. [Google Scholar] [CrossRef]
- Boruc, J.; Zhou, X.; Meier, I. Dynamics of the plant nuclear envelope and nuclear pore. Plant Physiol. 2012, 158, 78–86. [Google Scholar] [CrossRef]
- Capoen, W.; Sun, J.; Wysham, D.; Otegui, M.S.; Venkateshwaran, M.; Hirsch, S.; Miwa, H.; Downie, J.A.; Morris, R.J.; Ane, J.M.; et al. Nuclear membranes control symbiotic calcium signaling of legumes. Proc. Natl. Acad. Sci. USA 2011, 108, 14348–14353. [Google Scholar] [CrossRef]
- Danker, T.; Schillers, H.; Storck, J.; Shahin, V.; Kramer, B.; Wilhelmi, M.; Oberleithner, H. Nuclear hourglass technique: An approach that detects electrically open nuclear pores in Xenopus laevis oocyte. Proc. Natl. Acad. Sci. USA 1999, 96, 13530–13535. [Google Scholar] [CrossRef]
- Al-Mohanna, F.A.; Caddy, K.W.; Bolsover, S.R. The nucleus is insulated from large cytosolic calcium ion changes. Nature 1994, 367, 745–750. [Google Scholar] [CrossRef]
- Erickson, E.S.; Mooren, O.L.; Moore, D.; Krogmeier, J.R.; Dunn, R.C. The role of nuclear envelope calcium in modifying nuclear pore complex structure. Can. J. Physiol. Pharmacol. 2006, 84, 309–318. [Google Scholar] [CrossRef]
- Perez-Terzic, C.; Pyle, J.; Jaconi, M.; Stehno-Bittel, L.; Clapham, D.E. Conformational states of the nuclear pore complex induced by depletion of nuclear Ca2+ stores. Science 1996, 273, 1875–1877. [Google Scholar] [CrossRef]
- Eder, A.; Bading, H. Calcium signals can freely cross the nuclear envelope in hippocampal neurons: Somatic calcium increases generate nuclear calcium transients. BMC Neurosci. 2007. [Google Scholar] [CrossRef]
- Keizer, J.; Smith, G.D.; Ponce-Dawson, S.; Pearson, J.E. Saltatory propagation of Ca2+ waves by Ca2+ sparks. Biophys. J. 1998, 75, 595–600. [Google Scholar] [CrossRef]
- Dawson, S.P.; Keizer, J.; Pearson, J.E. Fire-diffuse-fire model of dynamics of intracellular calcium waves. Proc. Natl. Acad. Sci. USA 1999, 96, 6060–6063. [Google Scholar] [CrossRef]
- Coombes, S. The effect of ion pumps on the speed of travelling waves in the fire-diffuse-fire model of Ca2+ release. Bull. Math. Biol. 2001, 63, 1–20. [Google Scholar] [CrossRef]
- Sieberer, B.J.; Chabaud, M.; Timmers, A.C.; Monin, A.; Fournier, J.; Barker, D.G. A nuclear-targeted cameleon demonstrates intranuclear Ca2+ spiking in Medicago truncatula root hairs in response to rhizobial nodulation factors. Plant Physiol. 2009, 151, 1197–1206. [Google Scholar] [CrossRef]
- Mazel, T.; Raymond, R.; Raymond-Stintz, M.; Jett, S.; Wilson, B.S. Stochastic modeling of calcium in 3D geometry. Biophys. J. 2009, 96, 1691–1706. [Google Scholar]
- Means, S.; Smith, A.J.; Shepherd, J.; Shadid, J.; Fowler, J.; Wojcikiewicz, R.J.; Mazel, T.; Smith, G.D.; Wilson, B.S. Reaction diffusion modeling of calcium dynamics with realistic ER geometry. Biophys. J. 2006, 91, 537–557. [Google Scholar] [CrossRef]
- Queisser, G.; Wiegert, S.; Bading, H. Structural dynamics of the cell nucleus: Basis for morphology modulation of nuclear calcium signaling and gene transcription. Nucleus 2011, 2, 98–104. [Google Scholar] [CrossRef]
- Klann, M.; Koeppl, H. Spatial simulations in systems biology: From molecules to cells. Int. J. Mol. Sci. 2012, 13, 7798–7827. [Google Scholar] [CrossRef]
- Lenton, T.M.; Crouch, M.; Johnson, M.; Pires, N.; Dolan, L. First plants cooled the Ordovician. Nat. Geosci. 2012, 5, 86–89. [Google Scholar] [CrossRef]
- Thuiller, W.; Lavorel, S.; Araujo, M.B.; Sykes, M.T.; Prentice, I.C. Climate change threats to plant diversity in Europe. Proc. Natl. Acad. Sci. USA 2005, 102, 8245–8250. [Google Scholar]
- Walbot, V. How plants cope with temperature stress. Bmc. Biol. 2011. [Google Scholar] [CrossRef]
- Knight, M.R.; Campbell, A.K.; Smith, S.M.; Trewavas, A.J. Transgenic plant aequorin reports the effects of touch and cold-shock and elicitors on cytoplasmic calcium. Nature 1991, 352, 524–526. [Google Scholar] [CrossRef]
- Minorsky, P.V. Temperature sensing by plants—A review and hypothesis. Plant Cell Environ. 1989, 12, 119–135. [Google Scholar] [CrossRef]
- Minorsky, P.V.; Spanswick, R.M. Electrophysiological evidence for a role for calcium in temperature sensing by roots of cucumber seedlings. Plant Cell Environ. 1989, 12, 137–143. [Google Scholar] [CrossRef]
- Ruelland, E.; Zachowski, A. How plants sense temperature. Environ. Exp. Bot. 2010, 69, 225–232. [Google Scholar] [CrossRef]
- McClung, C.R.; Davis, S.J. Ambient thermometers in plants: From physiological outputs towards mechanisms of thermal sensing. Curr. Biol. 2010, 20, R1086–R1092. [Google Scholar] [CrossRef]
- Browse, J.; Xin, Z. Temperature sensing and cold acclimation. Curr. Opin. Plant. Biol. 2001, 4, 241–246. [Google Scholar] [CrossRef]
- Plieth, C.; Hansen, U.P.; Knight, H.; Knight, M.R. Temperature sensing by plants: The primary characteristics of signal perception and calcium response. Plant J. 1999, 18, 491–497. [Google Scholar] [CrossRef]
- Plieth, C. Temperature sensing by plants: Calcium-permeable channels as primary sensors—A model. J. Membr. Biol. 1999, 172, 121–127. [Google Scholar] [CrossRef]
- Caldwell, C.R.; Haug, A. Temperature-dependence of the barley root plasma membrane-bound Ca2+-dependent and Mg2+-dependent atpase. Physiol. Plant. 1981, 53, 117–124. [Google Scholar] [CrossRef]
- White, P.J.; Ridout, M.S. An energy-barrier model for the permeation of monovalent and divalent cations through the maxi cation channel in the plasma membrane of rye roots. J. Membr. Biol. 1999, 168, 63–75. [Google Scholar] [CrossRef]
- Eyring, H. The activated complex in chemical reactions. J. Chem. Phys. 1953, 3, 107:1–107:9. [Google Scholar]
- Hanggi, P.; Talkner, P.; Borkovec, M. Reaction-rate theory—50 years after kramers. Rev. Mod. Phys. 1990, 62, 251–341. [Google Scholar] [CrossRef]
- White, P.J.; Pineros, M.; Tester, M.; Ridout, M.S. Cation permeability and selectivity of a root plasma membrane calcium channel. J. Membr. Biol. 2000, 174, 71–83. [Google Scholar] [CrossRef]
- White, P.J. Calcium signals in root cells: The roles of plasma membrane calcium channels. Biologia 2004, 59, 77–83. [Google Scholar]
- White, P.J. Depolarization-activated calcium channels shape the calcium signatures induced by low-temperature stress. New Phytol. 2009, 183, 6–8. [Google Scholar] [CrossRef]
- Bose, J.; Pottosin, I.I.; Shabala, S.S.; Palmgren, M.G.; Shabala, S. Calcium efflux systems in stress signaling and adaptation in plants. Front. Plant Sci. 2011. [Google Scholar] [CrossRef]
- Clapham, D.E.; Miller, C. A thermodynamic framework for understanding temperature sensing by transient receptor potential (TRP) channels. Proc. Natl. Acad. Sci. USA 2011, 108, 19492–19497. [Google Scholar] [CrossRef]
- Voets, T.; Droogmans, G.; Wissenbach, U.; Janssens, A.; Flockerzi, V.; Nilius, B. The principle of temperature-dependent gating in cold- and heat-sensitive TRP channels. Nature 2004, 430, 748–754. [Google Scholar] [CrossRef] [Green Version]
- Finka, A.; Cuendet, A.F.; Maathuis, F.J.; Saidi, Y.; Goloubinoff, P. Plasma membrane cyclic nucleotide gated calcium channels control land plant thermal sensing and acquired thermotolerance. Plant Cell 2012, 24, 3333–3348. [Google Scholar] [CrossRef]
- Gao, F.; Han, X.W.; Wu, J.H.; Zheng, S.Z.; Shang, Z.L.; Sun, D.Y.; Zhou, R.G.; Li, B. A heat-activated calcium-permeable channel—Arabidopsis cyclic nucleotide-gated ion channel 6—is involved in heat shock responses. Plant J. 2012, 70, 1056–1069. [Google Scholar] [CrossRef]
- Tunc-Ozdemir, M.; Tang, C.; Ishka, M.R.; Brown, E.; Groves, N.R.; Myers, C.T.; Rato, C.; Poulsen, L.R.; McDowell, S.; Miller, G.; et al. A cyclic nucleotide-gated channel (CNGC16) in pollen is critical for stress tolerance in pollen reproductive development. Plant Physiol. 2013, 161, 1010–1020. [Google Scholar]
- Liu, J.; Knight, H.; Hurst, C.H.; Knight, M.R. Modelling and experimental analysis of the role of interacting cytosolic and vacuolar pools in shaping low temperature calcium signatures in plant cells. Mol. Biosyst. 2012, 8, 2205–2220. [Google Scholar] [CrossRef]
- Jones, C.T.; Banks, P. The influence of the intracellular concentration of sodium on the uptake of L-( 14 C)valine by chopped tissue from cerebral cortex. Biochem. J. 1971, 123, 341–345. [Google Scholar]
- Parent, B.; Turc, O.; Gibon, Y.; Stitt, M.; Tardieu, F. Modelling temperature-compensated physiological rates, based on the co-ordination of responses to temperature of developmental processes. J. Exp. Bot. 2010, 61, 2057–2069. [Google Scholar] [CrossRef]
- Murata, N.; Los, D.A. Membrane fluidity and temperature perception. Plant Physiol. 1997, 115, 875–879. [Google Scholar]
- Finka, A.; Goloubinoff, P. The CNGCb and CNGCd genes from Physcomitrella patens moss encode for thermosensory calcium channels responding to fluidity changes in the plasma membrane. Cell Stress Chaperones 2013. [Google Scholar] [CrossRef]
- Oldroyd, G.E.; Murray, J.D.; Poole, P.S.; Downie, J.A. The rules of engagement in the legume-rhizobial symbiosis. Annu. Rev. Genet. 2011, 45, 119–144. [Google Scholar] [CrossRef]
- Shaw, S.L.; Long, S.R. Nod factor elicits two separable calcium responses in Medicago truncatula root hair cells. Plant Physiol. 2003, 131, 976–984. [Google Scholar] [CrossRef]
- Stracke, S.; Kistner, C.; Yoshida, S.; Mulder, L.; Sato, S.; Kaneko, T.; Tabata, S.; Sandal, N.; Stougaard, J.; Szczyglowski, K.; et al. A plant receptor-like kinase required for both bacterial and fungal symbiosis. Nature 2002, 417, 959–962. [Google Scholar] [CrossRef]
- Mitra, R.M.; Gleason, C.A.; Edwards, A.; Hadfield, J.; Downie, J.A.; Oldroyd, G.E.D.; Long, S.R. A Ca2+/calmodulin-dependent protein kinase required for symbiotic nodule development: Gene identification by transcript-based cloning. Proc. Natl. Acad. Sci. USA 2004, 101, 4701–4705. [Google Scholar]
- Gleason, C.; Chaudhuri, S.; Yang, T.B.; Munoz, A.; Poovaiah, B.W.; Oldroyd, G.E.D. Nodulation independent of rhizobia induced by a calcium-activated kinase lacking autoinhibition. Nature 2006, 441, 1149–1152. [Google Scholar] [CrossRef]
- Engstrom, E.M.; Ehrhardt, D.W.; Mitra, R.M.; Long, S.R. Pharmacological analysis of nod factor-induced calcium spiking in Medicago truncatula. Evidence for the requirement of type IIA calcium pumps and phosphoinositide signaling. Plant Physiol. 2002, 128, 1390–1401. [Google Scholar] [CrossRef]
- Charron, D.; Pingret, J.L.; Chabaud, M.; Journet, E.P.; Barker, D.G. Pharmacological evidence that multiple phospholipid signaling pathways link rhizobium nodulation factor perception in Medicago truncatula root hairs to intracellular responses, including Ca2+ spiking and specific ENOD gene expression. Plant Physiol. 2004, 136, 3582–3593. [Google Scholar] [CrossRef]
- Maillet, F.; Poinsot, V.; Andre, O.; Puech-Pages, V.; Haouy, A.; Gueunier, M.; Cromer, L.; Giraudet, D.; Formey, D.; Niebel, A.; et al. Fungal lipochitooligosaccharide symbiotic signals in arbuscular mycorrhiza. Nature 2011, 469, 58–63. [Google Scholar] [CrossRef]
- Miyawaki, A.; Griesbeck, O.; Heim, R.; Tsien, R.Y. Dynamic and quantitative Ca2+ measurements using improved cameleons. Proc. Natl. Acad. Sci. USA 1999, 96, 2135–2140. [Google Scholar] [CrossRef]
- Deisseroth, K.; Heist, E.K.; Tsien, R.W. Translocation of calmodulin to the nucleus supports CREB phosphorylation in hippocampal neurons. Nature 1998, 392, 198–202. [Google Scholar] [CrossRef]
- Sciacca, E.; Spinella, S.; Genre, A.; Calcagno, C. Analysis of calcium spiking in plant root epidermis through CWC modeling. Electron. Notes Theor. Comput. Sci. 2011, 277, 65–76. [Google Scholar] [CrossRef]
- Coppo, M.; Damiani, F.; Drocco, M.; Grassi, E.; Troina, A. Stochastic calculus of wrapped compartments. In Proceeding of Eighth Workshop on Quantitative Aspects of Programming Languages (QAPL 2010), Paphos, Cyprus, 27–28 March 2010; pp. 82–98.
- Terese Term Rewriting Systems, Cambridge University Press: Cambridge, UK, 2003.
- Jaynes, E.T. Information theory and statistical mechanics. Phys. Rev. 1957, 106, 620–630. [Google Scholar] [CrossRef]
- Jaynes, E.T. Information theory and statistical mechanics 2. Phys. Rev. 1957, 108, 171–190. [Google Scholar] [CrossRef]
- Calcagno, C.; Coppo, M.; Damiani, F.; Drocco, M.; Sciacca, E.; Spinella, S.; Troina, A. Modelling spatial interactions in the arbuscular mycorrhizal symbiosis using the calculus of wrapped compartments. In Proceeding of EPTCS 2011, Aachen, Germany, 10 September 2011; pp. 3–18.
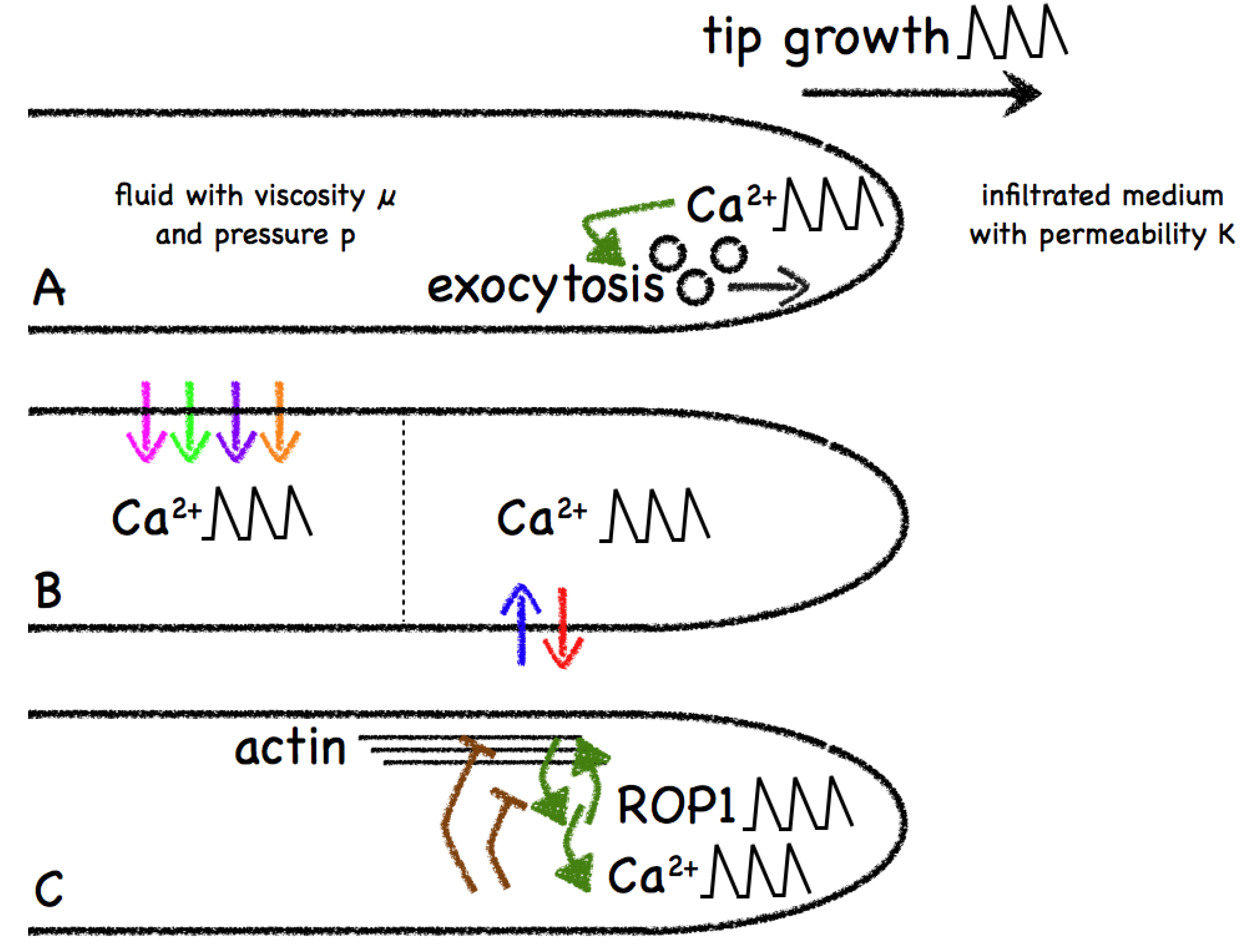
- Kroeger, J.; Geitmann, A. The pollen tube paradigm revisited. Curr. Opin. Plant. Biol. 2012, 15, 618–624. [Google Scholar] [CrossRef]
- Kroeger, J.H.; Geitmann, A. Pollen tube growth: Getting a grip on cell biology through modeling. Mech. Res. Commun. 2012, 42, 32–39. [Google Scholar] [CrossRef]
- Bibikova, T.N.; Zhigilei, A.; Gilroy, S. Root hair growth in Arabidopsis thaliana is directed by calcium and an endogenous polarity. Planta 1997, 203, 495–505. [Google Scholar] [CrossRef]
- Pierson, E.S.; Miller, D.D.; Callaham, D.A.; Shipley, A.M.; Rivers, B.A.; Cresti, M.; Hepler, P.K. Pollen tube growth is coupled to the extracellular calcium ion flux and the intracellular calcium gradient: Effect of BAPTA-type buffers and hypertonic media. Plant Cell 1994, 6, 1815–1828. [Google Scholar]
- Weisenseel, M.H.; Nuccitelli, R.; Jaffe, L.F. Large electrical currents traverse growing pollen tubes. J. Cell Biol. 1975, 66, 556–567. [Google Scholar] [CrossRef]
- Geitmann, A.; Li, Y.Q.; Cresti, M. The role of the cytoskeleton and dictyosome activity in the pulsatory growth of Nicotiana tabacum and Petunia hybrida pollen tubes. Bot. Acta 1996, 109, 102–109. [Google Scholar]
- Messerli, M.A.; Robinson, K.R. Ionic and osmotic disruptions of the lily pollen tube oscillator: Testing proposed models. Planta 2003, 217, 147–157. [Google Scholar]
- Holdaway-Clarke, T.L.; Feijo, J.A.; Hackett, G.R.; Kunkel, J.G.; Hepler, P.K. Pollen tube growth and the intracellular cytosolic calcium gradient oscillate in phase while extracellular calcium influx is delayed. Plant Cell 1997, 9, 1999–2010. [Google Scholar]
- Kroeger, J.H.; Geitmann, A.; Grant, M. Model for calcium dependent oscillatory growth in pollen tubes. J. Theor. Biol. 2008, 253, 363–374. [Google Scholar] [CrossRef]
- Skotheim, J.M.; Mahadevan, L. Dynamics of poroelastic filaments. Proc. R. Soc. Lond. A 2004, 460, 1995–2020. [Google Scholar] [CrossRef]
- Dumais, J.; Shaw, S.L.; Steele, C.R.; Long, S.R.; Ray, P.M. An anisotropic-viscoplastic model of plant cell morphogenesis by tip growth. Int. J. Dev. Biol. 2006, 50, 209–222. [Google Scholar] [CrossRef]
- Holdaway-Clarke, T.L.; Hepler, P.K. Control of pollen tube growth: Role of ion gradients and fluxes. New Phytol. 2003, 159, 539–563. [Google Scholar] [CrossRef]
- Kroeger, J.H.; Zerzour, R.; Geitmann, A. Regulator or driving force? The role of turgor pressure in oscillatory plant cell growth. PLoS One 2011, 6, e18549. [Google Scholar] [CrossRef]
- Lockhart, J.A. An analysis of irreversible plant cell elongation. J. Theor. Biol. 1965, 8, 264–275. [Google Scholar]
- Liu, J.; Piette, B.M.; Deeks, M.J.; Franklin-Tong, V.E.; Hussey, P.J. A compartmental model analysis of integrative and self-regulatory ion dynamics in pollen tube growth. PLoS One 2010, 5, e13157. [Google Scholar]
- Yan, A.; Xu, G.; Yang, Z.B. Calcium participates in feedback regulation of the oscillating ROP1 Rho GTPase in pollen tubes. Proc. Natl. Acad. Sci. USA 2009, 106, 22002–22007. [Google Scholar] [CrossRef]
- Goodwin, B.C.; Trainor, L.E.H. Tip and whorl morphogenesis in acetabularia by calcium-regulated strain fields. J. Theor. Biol. 1985, 117, 79–106. [Google Scholar] [CrossRef]
- Briere, C. Dynamics of the goodwin-trainor mechanochemical model. Acta Biotheor. 1994, 42, 137–146. [Google Scholar] [CrossRef]
- Briere, C.; Goodwin, B. Geometry and dynamics of tip morphogenesis in acetabularia. J. Theor. Biol. 1988, 131, 461–475. [Google Scholar] [CrossRef]
- Briere, C.; Goodwin, B.C. Effects of calcium input output on the stability of a system for calcium-regulated viscoelastic strain fields. J. Math. Biol. 1990, 28, 585–593. [Google Scholar] [CrossRef]
- Kim, T.H.; Bohmer, M.; Hu, H.; Nishimura, N.; Schroeder, J.I. Guard cell signal transduction network: Advances in understanding abscisic acid, CO2, and Ca2+ signaling. Annu. Rev. Plant. Biol. 2010, 61, 561–591. [Google Scholar] [CrossRef]
- Albert, R. Network inference, analysis, and modeling in systems biology. Plant Cell 2007, 19, 3327–3338. [Google Scholar] [CrossRef]
- Li, S.; Assmann, S.M.; Albert, R. Predicting essential components of signal transduction networks: A dynamic model of guard cell abscisic acid signaling. PLoS Biol. 2006, 4, e312. [Google Scholar] [CrossRef]
- Albert, I.; Thakar, J.; Li, S.; Zhang, R.; Albert, R. Boolean network simulations for life scientists. Source Code Biol. Med. 2008. [Google Scholar] [CrossRef]
- Veresov, V.G.; Kabak, A.G.; Volotovsky, I.D. Modeling the calcium signaling in stomatal guard cells under the action of abscisic acid. Russ. J. Plant Physl. 2003, 50, 573–579. [Google Scholar] [CrossRef]
- Allen, G.J.; Muir, S.R.; Sanders, D. Release of Ca2+ from individual plant vacuoles by both insp(3) and cyclic adp-ribose. Science 1995, 268, 735–737. [Google Scholar] [CrossRef]
- Hills, A.; Chen, Z.H.; Amtmann, A.; Blatt, M.R.; Lew, V.L. OnGuard, a computational platform for quantitative kinetic modeling of guard cell physiology. Plant Physiol. 2012, 159, 1026–1042. [Google Scholar] [CrossRef]
- Chen, Z.H.; Hills, A.; Batz, U.; Amtmann, A.; Lew, V.L.; Blatt, M.R. Systems dynamic modeling of the stomatal guard cell predicts emergent behaviors in transport, signaling, and volume control. Plant Physiol. 2012, 159, 1235–1251. [Google Scholar] [CrossRef]
- Dodd, A.N.; Gardner, M.J.; Hotta, C.T.; Hubbard, K.E.; Dalchau, N.; Love, J.; Assie, J.M.; Robertson, F.C.; Jakobsen, M.K.; Goncalves, J.; et al. The Arabidopsis circadian clock incorporates a cADPR-based feedback loop. Science 2007, 318, 1789–1792. [Google Scholar] [CrossRef]
- McAinsh, M.R.; Pittman, J.K. Shaping the calcium signature. New Phytol. 2009, 181, 275–294. [Google Scholar] [CrossRef]
- Dodd, A.N.; Jakobsen, M.K.; Baker, A.J.; Telzerow, A.; Hou, S.W.; Laplaze, L.; Barrot, L.; Poethig, R.S.; Haseloff, J.; Webb, A.A. Time of day modulates low-temperature Ca signals in Arabidopsis. Plant J. 2006, 48, 962–973. [Google Scholar] [CrossRef]
- Locke, J.C.; Kozma-Bognar, L.; Gould, P.D.; Feher, B.; Kevei, E.; Nagy, F.; Turner, M.S.; Hall, A.; Millar, A.J. Experimental validation of a predicted feedback loop in the multi-oscillator clock of Arabidopsis thaliana. Mol. Syst. Biol. 2006. [Google Scholar]
- Locke, J.C.; Millar, A.J.; Turner, M.S. Modelling genetic networks with noisy and varied experimental data: The circadian clock in Arabidopsis thaliana. J. Theor. Biol. 2005, 234, 383–393. [Google Scholar] [CrossRef]
- Locke, J.C.; Southern, M.M.; Kozma-Bognar, L.; Hibberd, V.; Brown, P.E.; Turner, M.S.; Millar, A.J. Extension of a genetic network model by iterative experimentation and mathematical analysis. Mol. Syst. Biol. 2005, 1, 2005.0013. [Google Scholar]
- Dalchau, N.; Hubbard, K.E.; Robertson, F.C.; Hotta, C.T.; Briggs, H.M.; Stan, G.B.; Goncalves, J.M.; Webb, A.A.R. Correct biological timing in Arabidopsis requires multiple light-signaling pathways. Proc. Natl. Acad. Sci. USA 2010, 107, 13171–13176. [Google Scholar] [CrossRef]
- Akaike, H. A new look at the statistical model identification. IEEE Trans. Autom. Control 1974, 19, 716–723. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach, 2nd ed.; Springer-Verlag: New York, NY, USA, 2002. [Google Scholar]
- Dalchau, N. Understanding biological timing using mechanistic and black-box models. New Phytol. 2012, 193, 852–858. [Google Scholar] [CrossRef]
- Gutenkunst, R.N.; Waterfall, J.J.; Casey, F.P.; Brown, K.S.; Myers, C.R.; Sethna, J.P. Universally sloppy parameter sensitivities in systems biology models. PLoS Comput. Biol. 2007, 3, 1871–1878. [Google Scholar]
- Rasmussen, C.E.; Williams, C.K.I. Gaussian Processes for Machine Learning; The MIT Press: Cambridge, MA, USA, 2006. [Google Scholar]
- Fromm, J.; Lautner, S. Electrical signals and their physiological significance in plants. Plant Cell Environ. 2007, 30, 249–257. [Google Scholar] [CrossRef]
- Felle, H.H.; Zimmermann, M.R. Systemic signalling in barley through action potentials. Planta 2007, 226, 203–214. [Google Scholar] [CrossRef]
- Cosgrove, D.J.; Hedrich, R. Stretch-activated chloride, potassium, and calcium channels coexisting in plasma membranes of guard cells of Vicia faba L. Planta 1991, 186, 143–153. [Google Scholar]
- Malone, M. Rapid, long-distance signal transmission in higher plants. Adv. Bot. Res. 1996, 22, 163–228. [Google Scholar] [CrossRef]
- Sukhov, V.S.; Vodeneev, V.A. Mathematical Model of Action Potential in Higher Plants. In Proceedings of the XII International Conference “Mathematics. Computer. Education”Section 8: Mathematical methods in biology, ecology and chemistry, Pushchino, Russia, 17–22 January 2005; Riznichenko, G.J., Ed.; Scientific Publishing Centre “Regular and Chaotic Dynamics”: Izhevsk, Russia, 2005; Volume 3, pp. 967–978. (in Russian). [Google Scholar]
- Gradmann, D. Impact of apoplast volume on ionic relations in plant cells. J. Membr. Biol. 2001, 184, 61–69. [Google Scholar] [CrossRef]
- Gradmann, D.; Hoffstadt, J. Electrocoupling of ion transporters in plants: Interaction with internal ion concentrations. J. Membr. Biol. 1998, 166, 51–59. [Google Scholar] [CrossRef]
- Sukhov, V.; Nerush, V.; Orlova, L.; Vodeneev, V. Simulation of action potential propagation in plants. J. Theor. Biol. 2011, 291, 47–55. [Google Scholar] [CrossRef]
- Sibaoka, T. Rapid plant movements triggered by action-potentials. Bot. Mag. Tokyo 1991, 104, 73–95. [Google Scholar] [CrossRef]
- Trebacz, K.; Dziubinska, H.; Krol, E. Electrical signals in long-distance communication in plants. Commun. Plants Neuronal Asp. Plant Life 2006, 2006, 277–290. [Google Scholar]
- Sukhov, V.; Akinchits, E.; Katicheva, L.; Vodeneev, V. Simulation of variation potential in higher plant cells. J. Membr. Biol. 2013, 246, 287–296. [Google Scholar] [CrossRef]
- Stahlberg, R.; Cleland, R.E.; van Volkenburgh, E. Decrement and amplification of slow wave potentials during their propagation in Helianthus annuus L. shoots. Planta 2005, 220, 550–558. [Google Scholar] [CrossRef]
- Sukhov, V.; Vodeneev, V. A mathematical model of action potential in cells of vascular plants. J. Membr. Biol. 2009, 232, 59–67. [Google Scholar] [CrossRef]
- Plieth, C.; Sattelmacher, B.; Hansen, U.P.; Thiel, G. The action potential in Chara: Ca2+ release from internal stores visualized by Mn2+-induced quenching of fura-dextran. Plant J. 1998, 13, 167–175. [Google Scholar] [CrossRef]
- Dolmetsch, R.E.; Xu, K.; Lewis, R.S. Calcium oscillations increase the efficiency and specificity of gene expression. Nature 1998, 392, 933–936. [Google Scholar] [CrossRef]
- Scrase-Field, S.A.; Knight, M.R. Calcium: Just a chemical switch? Curr. Opin. Plant Biol. 2003, 6, 500–506. [Google Scholar] [CrossRef]
- Plieth, C. Calcium: Just another regulator in the machinery of life? Ann. Bot. 2005, 96, 1–8. [Google Scholar] [CrossRef]
- Granqvist, E.; Oldroyd, G.E.; Morris, R.J. Automated Bayesian model development for frequency detection in biological time series. BMC Syst. Biol. 2011. [Google Scholar] [CrossRef]
- Granqvist, E.; Hartley, M.; Morris, R.J. BaSAR-A tool in R for frequency detection. Biosystems 2012, 110, 60–63. [Google Scholar] [CrossRef]
- Sivia, D.S.; Skilling, J. Data Analysis: A Bayesian Tutorial, 2nd ed.; Oxford University Press: Oxford, UK, New York, NY, USA, 2006; pp. xii, 246. [Google Scholar]
- Bretthorst, G.L. Bayesian Spectrum Analysis and Parameter Estimation; Springer-Verlag: New York, NY, USA, 1988; pp. xii, 209. [Google Scholar]
- Skilling, J. Nested sampling for general bayesian computation. Bayesian Anal. 2006, 1, 833–859. [Google Scholar] [CrossRef]
- Casdagli, M. Chaos and deterministic versus stochastic nonlinear modeling. J. R. Stat. Soc. Ser. B Methodol. 1992, 54, 303–328. [Google Scholar]
- Kosuta, S.; Hazledine, S.; Sun, J.; Miwa, H.; Morris, R.J.; Downie, J.A.; Oldroyd, G.E. Differential and chaotic calcium signatures in the symbiosis signaling pathway of legumes. Proc. Natl. Acad. Sci. USA 2008, 105, 9823–9828. [Google Scholar]
- Aparicio, T.; Pozo, E.F.; Saura, D. Detecting determinism using recurrence quantification analysis: Three test procedures. J. Econ. Behav. Organ. 2008, 65, 768–787. [Google Scholar] [CrossRef]
- Kantz, H.; Schreiber, T. Nonlinear Time Series Analysis; Cambridge University Press: Cambridge, UK, New York, NY, USA, 1997; pp. xvi, 304. [Google Scholar]
- Hazledine, S.; Sun, J.; Wysham, D.; Downie, J.A.; Oldroyd, G.E.; Morris, R.J. Nonlinear time series analysis of nodulation factor induced calcium oscillations: Evidence for deterministic chaos? PLoS One 2009, 4, e6637. [Google Scholar]
- Kiegle, E.; Moore, C.A.; Haseloff, J.; Tester, M.A.; Knight, M.R. Cell-type-specific calcium responses to drought, salt and cold in the Arabidopsis root. Plant J. 2000, 23, 267–278. [Google Scholar] [CrossRef]
- Spiller, D.G.; Wood, C.D.; Rand, D.A.; White, M.R. Measurement of single-cell dynamics. Nature 2010, 465, 736–745. [Google Scholar] [CrossRef]
- Allen, G.J.; Chu, S.P.; Schumacher, K.; Shimazaki, C.T.; Vafeados, D.; Kemper, A.; Hawke, S.D.; Tallman, G.; Tsien, R.Y.; Harper, J.F.; et al. Alteration of stimulus-specific guard cell calcium oscillations and stomatal closing in Arabidopsis det3 mutant. Science 2000, 289, 2338–2342. [Google Scholar] [CrossRef]
- Plieth, C. Signal percolation through plants and the shape of the calcium signature. Plant Signal. Behav. 2010, 5, 379–385. [Google Scholar] [CrossRef]
- Ehrhardt, D.W.; Wais, R.; Long, S.R. Calcium spiking in plant root hairs responding to Rhizobium nodulation signals. Cell 1996, 85, 673–681. [Google Scholar] [CrossRef]
- Marhl, M.; Perc, M.; Schuster, S. A minimal model for decoding of time-limited Ca2+ oscillations. Biophys. Chem. 2006, 120, 161–167. [Google Scholar] [CrossRef]
- Miwa, H.; Sun, J.; Oldroyd, G.E.; Downie, J.A. Analysis of calcium spiking using a cameleon calcium sensor reveals that nodulation gene expression is regulated by calcium spike number and the developmental status of the cell. Plant J. 2006, 48, 883–894. [Google Scholar] [CrossRef]
- Li, Y.; Wang, G.X.; Xin, M.; Yang, H.M.; Wu, X.J.; Li, T. The parameters of guard cell calcium oscillation encodes stomatal oscillation and closure in Vicia faba. Plant Sci. 2004, 166, 415–421. [Google Scholar] [CrossRef]
- Yang, H.M.; Zhang, X.Y.; Wang, G.X. Cytosolic calcium oscillation signaling in guard cell. Plant Sci. 2004, 166, 549–556. [Google Scholar] [CrossRef]
- DeFalco, T.A.; Bender, K.W.; Snedden, W.A. Breaking the code: Ca2+ sensors in plant signalling. Biochem. J. 2010, 425, 27–40. [Google Scholar] [CrossRef]
- Schwaller, B. The continuing disappearance of “pure” Ca2+ buffers. Cell. Mol. Life Sci. 2009, 66, 275–300. [Google Scholar] [CrossRef]
- Gilroy, S.; Hughes, W.A.; Trewavas, A.J. The measurement of intracellular calcium levels in protoplasts from higher plant cells. FEBS Lett. 1986, 199, 217–221. [Google Scholar] [CrossRef]
- Miller, A.J.; Sanders, D. Depletion of cytosolic free calcium induced by photosynthesis. Nature 1987, 326, 397–400. [Google Scholar] [CrossRef]
- Felle, H. Auxin causes oscillations of cytosolic free calcium and Ph in zea-mays coleoptiles. Planta 1988, 174, 495–499. [Google Scholar] [CrossRef]
- McAinsh, M.R.; Brownlee, C.; Hetherington, A.M. Abscisic acid-induced elevation of guard-cell cytosolic Ca-2+ precedes stomatal closure. Nature 1990, 343, 186–188. [Google Scholar] [CrossRef]
- Hepler, P.K. Calcium: A central regulator of plant growth and development. Plant Cell 2005, 17, 2142–2155. [Google Scholar] [CrossRef]
- Swanson, S.J.; Choi, W.G.; Chanoca, A.; Gilroy, S. In vivo imaging of Ca2+, pH, and reactive oxygen species using fluorescent probes in plants. Annu. Rev. Plant Biol. 2011, 62, 273–297. [Google Scholar] [CrossRef]
- Fricker, M.; Runions, J.; Moore, I. Quantitative fluorescence microscopy: From art to science. Annu. Rev. Plant Biol. 2006, 57, 79–107. [Google Scholar] [CrossRef]
- McCombs, J.E.; Palmer, A.E. Measuring calcium dynamics in living cells with genetically encodable calcium indicators. Methods 2008, 46, 152–159. [Google Scholar] [CrossRef]
- Iwano, M.; Entani, T.; Shiba, H.; Kakita, M.; Nagai, T.; Mizuno, H.; Miyawaki, A.; Shoji, T.; Kubo, K.; Isogai, A.; et al. Fine-tuning of the cytoplasmic Ca2+ concentration is essential for pollen tube growth. Plant Physiol. 2009, 150, 1322–1334. [Google Scholar] [CrossRef]
- Horikawa, K.; Yamada, Y.; Matsuda, T.; Kobayashi, K.; Hashimoto, M.; Matsu-ura, T.; Miyawaki, A.; Michikawa, T.; Mikoshiba, K.; Nagai, T. Spontaneous network activity visualized by ultrasensitive Ca2+ indicators, yellow Cameleon-Nano. Nat. Methods 2010, 7, U729–U788. [Google Scholar] [CrossRef]
- Tsien, R.Y. Very long-term memories may be stored in the pattern of holes in the perineuronal net. Proc. Natl. Acad. Sci. USA 2013, 110, 12157–12158. [Google Scholar] [CrossRef]
- Magde, D.; Webb, W.W.; Elson, E. Thermodynamic fluctuations in a reacting system—Measurement by fluorescence correlation spectroscopy. Phys. Rev. Lett. 1972, 29, 705–709. [Google Scholar] [CrossRef]
- Eigen, M.; Rigler, R. Sorting single molecules: Application to diagnostics and evolutionary biotechnology. Proc. Natl. Acad. Sci. USA 1994, 91, 5740–5747. [Google Scholar] [CrossRef]
- Digman, M.A.; Dalal, R.; Horwitz, A.F.; Gratton, E. Mapping the number of molecules and brightness in the laser scanning microscope. Biophys. J. 2008, 94, 2320–2332. [Google Scholar] [CrossRef]
- Ries, J.; Schwille, P. Fluorescence correlation spectroscopy. Bioessays 2012, 34, 361–368. [Google Scholar] [CrossRef]
- Elson, E.L. Fluorescence correlation spectroscopy: Past, present, future. Biophys. J. 2011, 101, 2855–2870. [Google Scholar] [CrossRef]
- Sanabria, H.; Digman, M.A.; Gratton, E.; Waxham, M.N. Spatial diffusivity and availability of intracellular calmodulin. Biophys. J. 2008, 95, 6002–6015. [Google Scholar] [CrossRef]
- Hebert, B.; Costantino, S.; Wiseman, P.W. Spatiotemporal image correlation spectroscopy (STICS) theory, verification, and application to protein velocity mapping in living CHO cells. Biophys. J. 2005, 88, 3601–3614. [Google Scholar] [CrossRef]
- Brown, C.M.; Hebert, B.; Kolin, D.L.; Zareno, J.; Whitmore, L.; Horwitz, A.R.; Wiseman, P.W. Probing the integrin-actin linkage using high-resolution protein velocity mapping. J. Cell Sci. 2006, 119, 5204–5214. [Google Scholar] [CrossRef]
- Cardarelli, F.; Gratton, E. In vivo imaging of single-molecule translocation through nuclear pore complexes by pair correlation functions. PLoS One 2010, 5, e10475. [Google Scholar] [CrossRef]
- Schermelleh, L.; Heintzmann, R.; Leonhardt, H. A guide to super-resolution fluorescence microscopy. J. Cell Biol. 2010, 190, 165–175. [Google Scholar] [CrossRef]
- Baker, S.; Kanade, T. Limits on super-resolution and how to break them. IEEE Trans. Pattern Anal. 2002, 24, 1167–1183. [Google Scholar] [CrossRef]
- Cho, S.; Jang, J.; Song, C.; Lee, H.; Ganesan, P.; Yoon, T.Y.; Kim, M.W.; Choi, M.C.; Ihee, H.; Do Heo, W.; et al. Simple super-resolution live-cell imaging based on diffusion-assisted Forster resonance energy transfer. Sci. Rep. 2013, 3, 1208:1–1208:16. [Google Scholar]
- Fernandez-Suarez, M.; Ting, A.Y. Fluorescent probes for super-resolution imaging in living cells. Nat. Rev. Mol. Cell Biol. 2008, 9, 929–943. [Google Scholar] [CrossRef]
- Henriques, R.; Griffiths, C.; Hesper Rego, E.; Mhlanga, M.M. PALM and STORM: Unlocking live-cell super-resolution. Biopolymers 2011, 95, 322–331. [Google Scholar] [CrossRef]
- Huang, B.; Babcock, H.; Zhuang, X. Breaking the diffraction barrier: Super-resolution imaging of cells. Cell 2010, 143, 1047–1058. [Google Scholar] [CrossRef]
- Lippincott-Schwartz, J.; Manley, S. Putting super-resolution fluorescence microscopy to work. Nat. Methods 2009, 6, 21–23. [Google Scholar] [CrossRef]
- Lu, D.; Liu, Z. Hyperlenses and metalenses for far-field super-resolution imaging. Nat. Commun. 2012, 3, 1205:1–1205:7. [Google Scholar]
- Lubeck, E.; Cai, L. Single-cell systems biology by super-resolution imaging and combinatorial labeling. Nat. Methods 2012, 9, 743–748. [Google Scholar] [CrossRef]
- Tipping, M.E.; Bishop, C.M. Bayesian Image Super-Resolution. In Advances in Neural Information Processing Systems; Thrun, S., Becker, S., Obermayer, K., Eds.; MIT Press: Cambridge, MA, USA, 2003; Volume 15, pp. 1279–1286. [Google Scholar]
- Gutierrez, R.; Grossmann, G.; Frommer, W.B.; Ehrhardt, D.W. Opportunities to explore plant membrane organization with super-resolution microscopy. Plant Physiol. 2010, 154, 463–466. [Google Scholar] [CrossRef]
- Fitzgibbon, J.; Bell, K.; King, E.; Oparka, K. Super-resolution imaging of plasmodesmata using three-dimensional structured illumination microscopy. Plant Physiol. 2010, 153, 1453–1463. [Google Scholar] [CrossRef]
- Sparkes, I.A.; Graumann, K.; Martiniere, A.; Schoberer, J.; Wang, P.; Osterrieder, A. Bleach it, switch it, bounce it, pull it: Using lasers to reveal plant cell dynamics. J. Exp. Bot. 2011, 62, 1–7. [Google Scholar] [CrossRef]
- Da Fonseca, P.C.A.; Morris, S.A.; Nerou, E.P.; Taylor, C.W.; Morris, E.P. Domain organization of the type 1 inositol 1,4,5-trisphosphate receptor as revealed by single-particle analysis. Proc. Natl. Acad. Sci. USA 2003, 100, 3936–3941. [Google Scholar]
- Thuleau, P.; Briere, C.; Mazars, C. Recent advances in plant cell nuclear signaling. Mol. Plant 2012, 5, 968–970. [Google Scholar] [CrossRef]
- Falcke, M. Reading the patterns in living cells—The physics of Ca2+ signaling. Adv. Phys. 2004, 53, 255–440. [Google Scholar] [CrossRef]
- Shuai, J.W.; Jung, P. Optimal ion channel clustering for intracellular calcium signaling. Proc. Natl. Acad. Sci. USA 2003, 100, 506–510. [Google Scholar] [CrossRef]
- Skupin, A.; Falcke, M. From puffs to global Ca2+ signals: How molecular properties shape global signals. Chaos 2009. [Google Scholar] [CrossRef]
- Williams, G.S.; Molinelli, E.J.; Smith, G.D. Modeling local and global intracellular calcium responses mediated by diffusely distributed inositol 1,4,5-trisphosphate receptors. J. Theor. Biol. 2008, 253, 170–188. [Google Scholar] [CrossRef]
- Moenke, G.; Falcke, M.; Thurley, K. Hierarchic stochastic modelling applied to intracellular Ca(2+) signals. PLoS One 2012, 7, e51178. [Google Scholar] [CrossRef]
- Dupont, G.; Croisier, H. Spatiotemporal organization of Ca dynamics: A modeling-based approach. HFSP J. 2010, 4, 43–51. [Google Scholar] [CrossRef]
- Solovey, G.; Dawson, S.P. Intra-cluster percolation of calcium signals. PLoS One 2010, 5, e8997. [Google Scholar] [CrossRef]
- Ullah, G.; Parker, I.; Mak, D.O.; Pearson, J.E. Multi-scale data-driven modeling and observation of calcium puffs. Cell Calcium 2012, 52, 152–160. [Google Scholar] [CrossRef]
- Baddeley, D.; Jayasinghe, I.D.; Lam, L.; Rossberger, S.; Cannell, M.B.; Soeller, C. Optical single-channel resolution imaging of the ryanodine receptor distribution in rat cardiac myocytes. Proc. Natl. Acad. Sci. USA 2009, 106, 22275–22280. [Google Scholar]
- Wiltgen, S.M.; Smith, I.F.; Parker, I. Superresolution localization of single functional IP3R channels utilizing Ca2+ flux as a readout. Biophys. J. 2010, 99, 437–446. [Google Scholar] [CrossRef]
- Parker, I.; Smith, I.F. Recording single-channel activity of inositol trisphosphate receptors in intact cells with a microscope, not a patch clamp. J. Gen. Physiol. 2010, 136, 119–127. [Google Scholar] [CrossRef]
- Smith, I.F.; Parker, I. Imaging the quantal substructure of single IP3R channel activity during Ca2+ puffs in intact mammalian cells. Proc. Natl. Acad. Sci. USA 2009, 106, 6404–6409. [Google Scholar] [CrossRef]
- Vogelstein, J.T.; Watson, B.O.; Packer, A.M.; Yuste, R.; Jedynak, B.; Paninski, L. Spike inference from calcium imaging using sequential monte carlo methods. Biophys. J. 2009, 97, 636–655. [Google Scholar] [CrossRef]
- Bhargava, A.; Lin, X.; Novak, P.; Mehta, K.; Korchev, Y.; Delmar, M.; Gorelik, J. Super-resolution scanning patch clamp reveals clustering of functional ion channels in adult ventricular myocyte. Circ. Res. 2013, 112, 1112–1120. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Martins, T.V.; Evans, M.J.; Woolfenden, H.C.; Morris, R.J. Towards the Physics of Calcium Signalling in Plants. Plants 2013, 2, 541-588. https://doi.org/10.3390/plants2040541
Martins TV, Evans MJ, Woolfenden HC, Morris RJ. Towards the Physics of Calcium Signalling in Plants. Plants. 2013; 2(4):541-588. https://doi.org/10.3390/plants2040541
Chicago/Turabian StyleMartins, Teresa Vaz, Matthew J. Evans, Hugh C. Woolfenden, and Richard J. Morris. 2013. "Towards the Physics of Calcium Signalling in Plants" Plants 2, no. 4: 541-588. https://doi.org/10.3390/plants2040541
APA StyleMartins, T. V., Evans, M. J., Woolfenden, H. C., & Morris, R. J. (2013). Towards the Physics of Calcium Signalling in Plants. Plants, 2(4), 541-588. https://doi.org/10.3390/plants2040541