Identification of Chimeric Repressors that Confer Salt and Osmotic Stress Tolerance in Arabidopsis


Abstract
:1. Introduction
2. Results and Discussion
2.1. Selection of Salt Tolerant CRES-T Lines

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CRES-T lines | Transcription factors converted to chimeric repressors | |||
|---|---|---|---|---|
| Locus | Description | Alias | Family | |
| ADA2b-SRDX | AT4G16420 | DNA binding/transcription coactivator/transcription factor | PRZ1 ADA2B | MYB |
| Msantd-SRDX | AT4G31270 | A transcription factor with Myb/SANT-like DNA-binding domain | Msantd | Trihelix |
| DDF1-SRDX | AT1G21610 | Wound-responsive family protein | DDF1 | Wound-responsive |
| DREB26-SRDX | AT1G21910 | AP2 domain-containing transcription factor family protein | DREB26 | AP2-EREBP |
| AtGeBP-SRDX | AT1G11510 | DNA-binding storekeeper protein-related | AtGeBP | GeBP |
| ATHB23-SRDX | AT1G26960 | DNA binding/transcription factor | ATHB23 | HB |
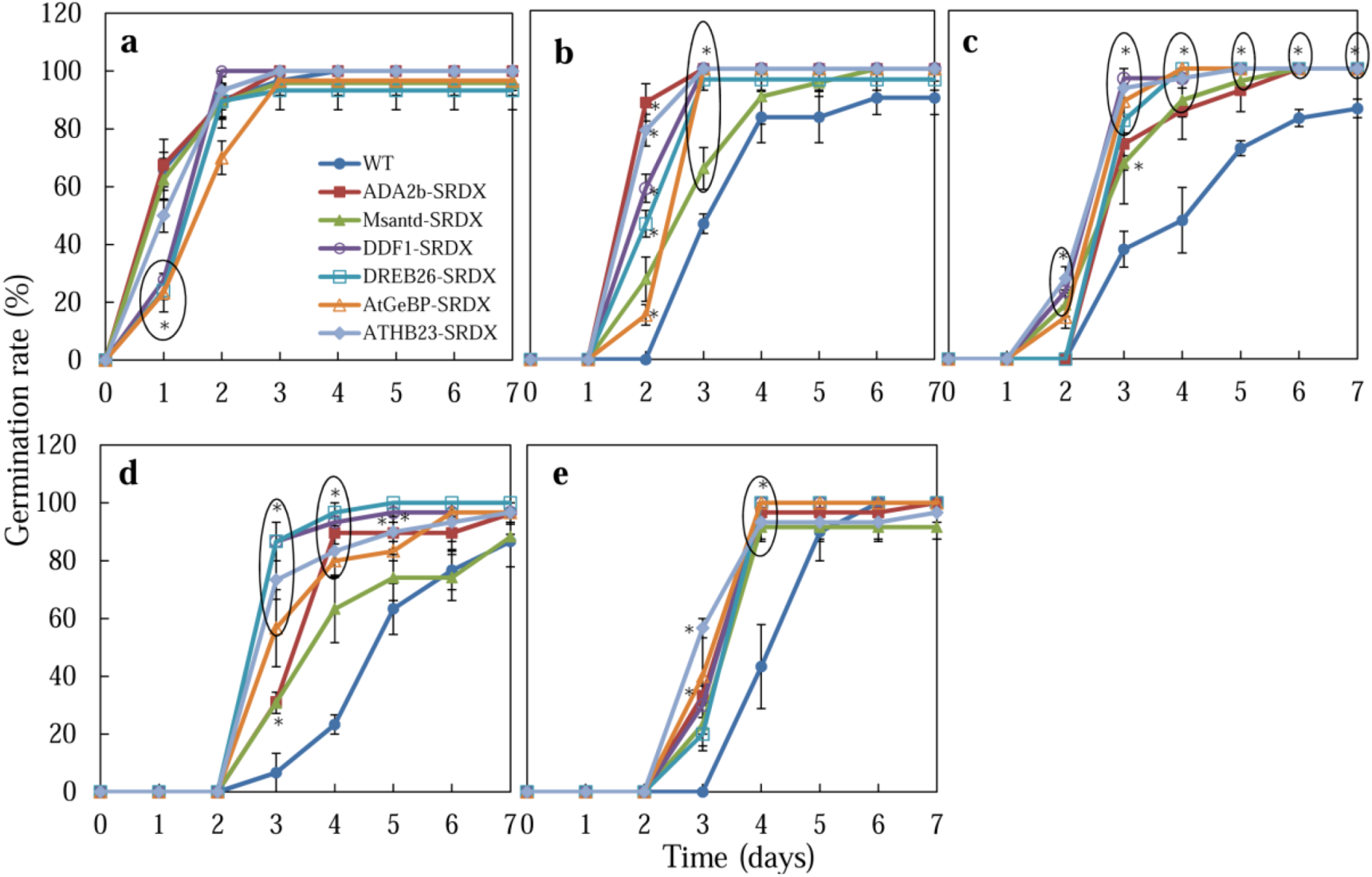
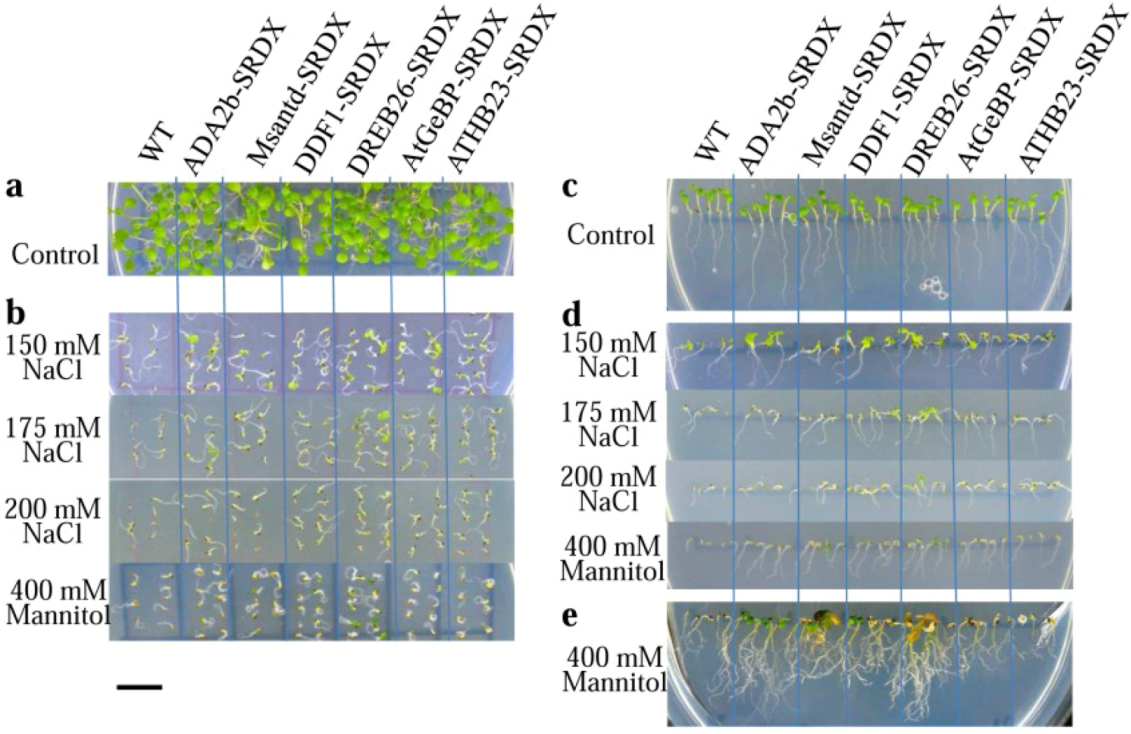
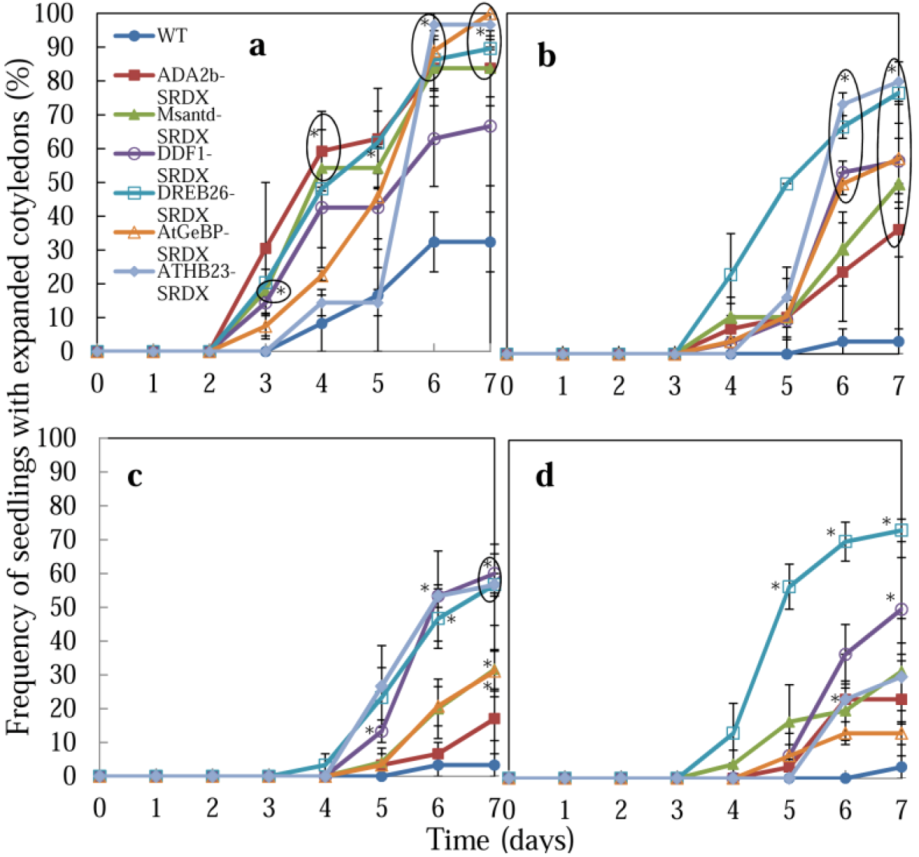
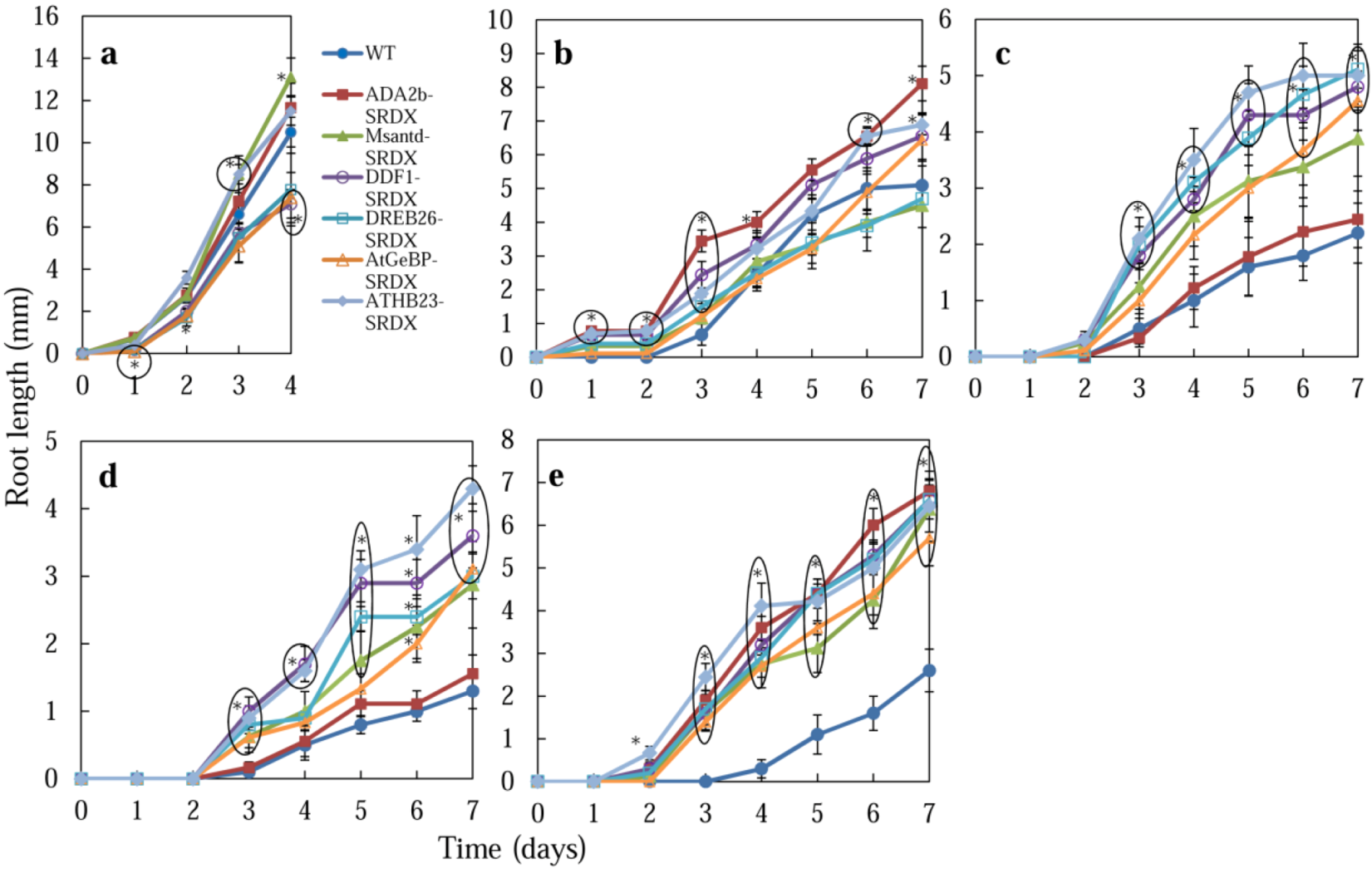
2.2. Germination of CRES-T Lines on Medium Supplemented with Different Concentrations of NaCl and Mannitol
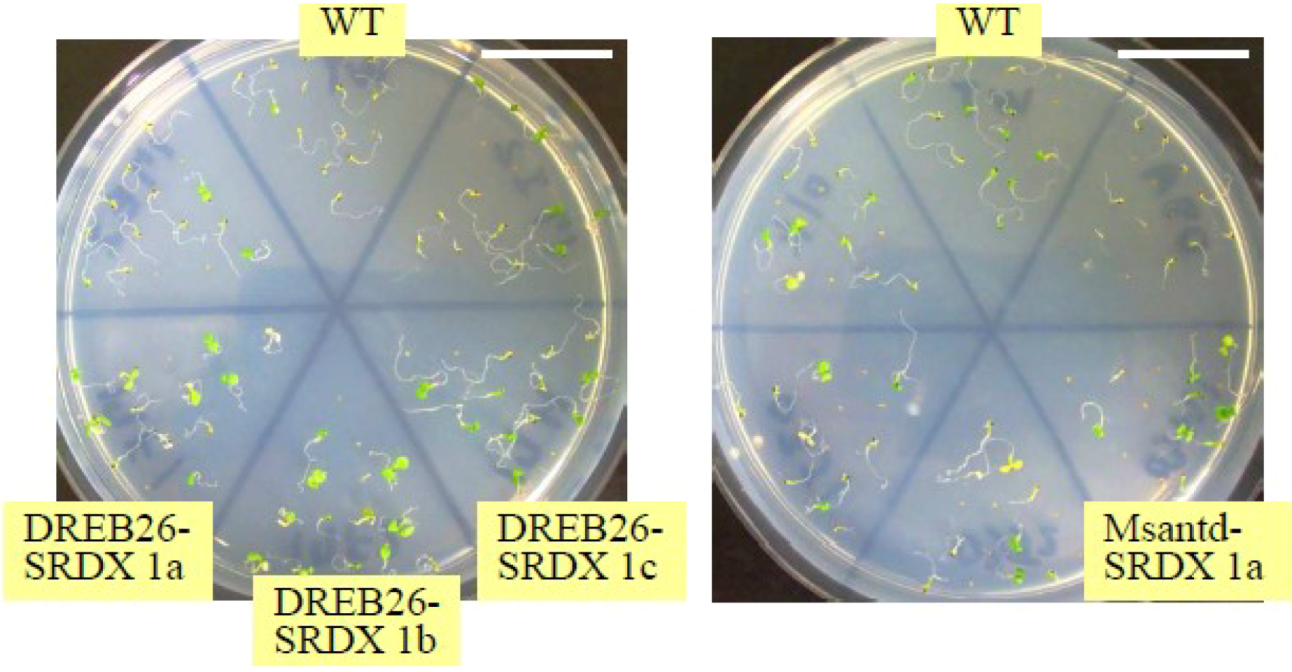
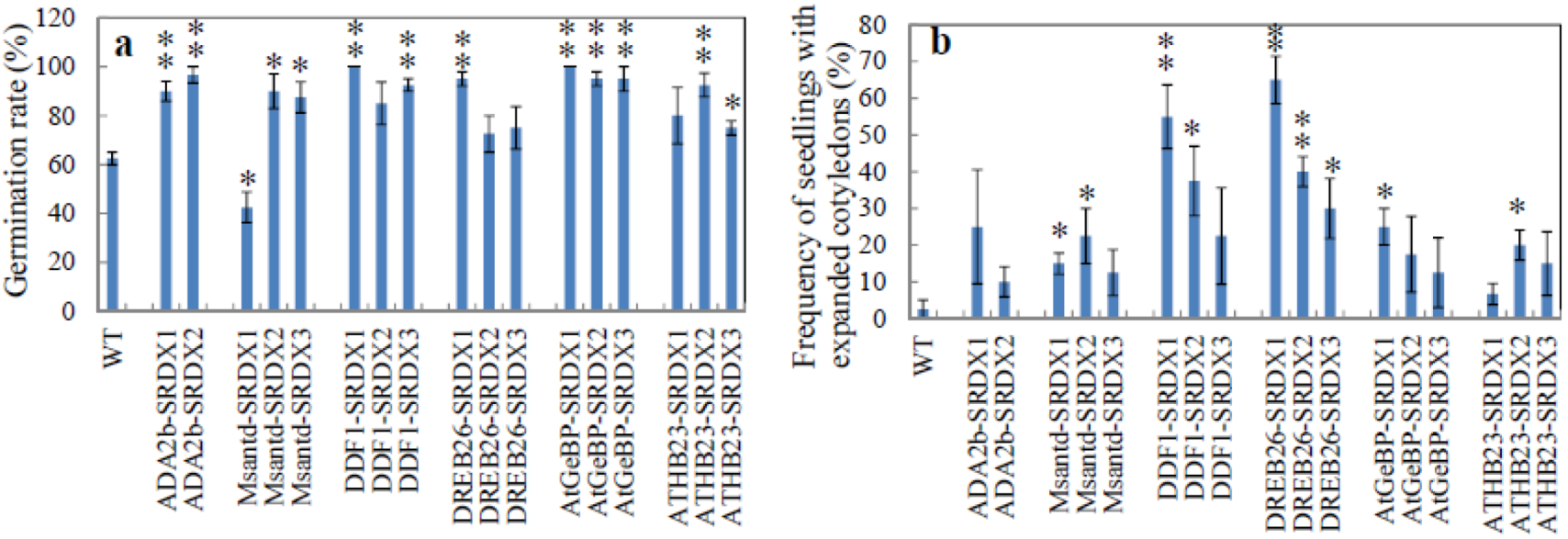
2.2.1. Germination Rate



2.2.2. Rate of Cotyledon Appearance among Seedlings
2.2.3. Root Growth
2.2.4. Possible Roles of Six Transcription Factors in Salt and Osmotic Stress Tolerance

2.3. Salt Tolerance Test in Rosette-Stage Plants
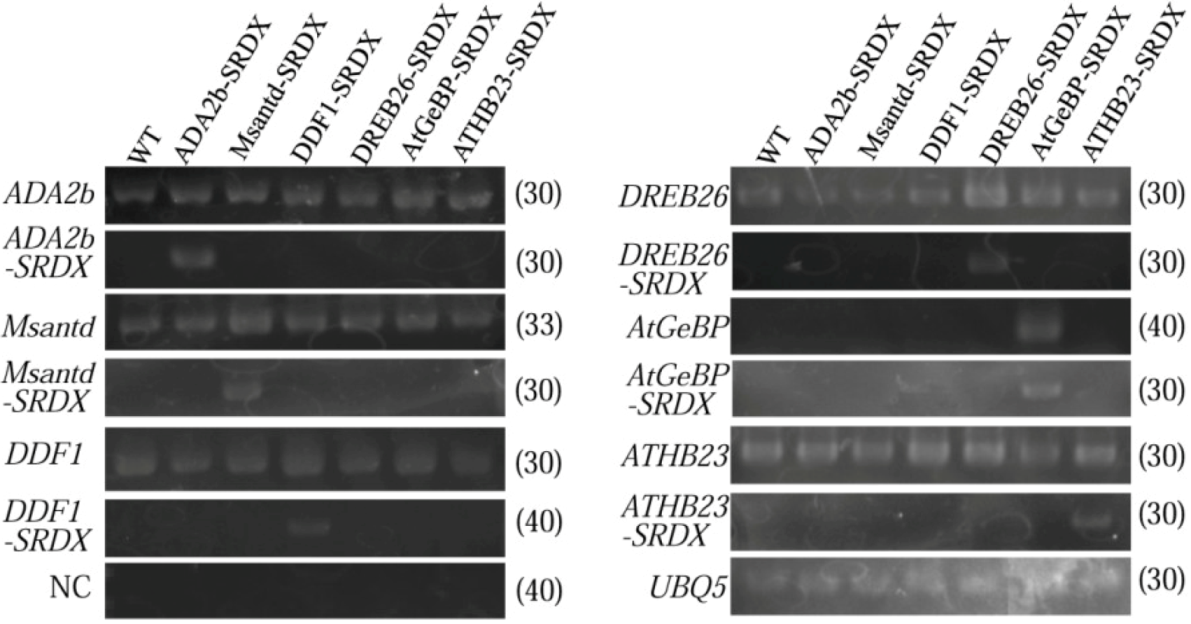
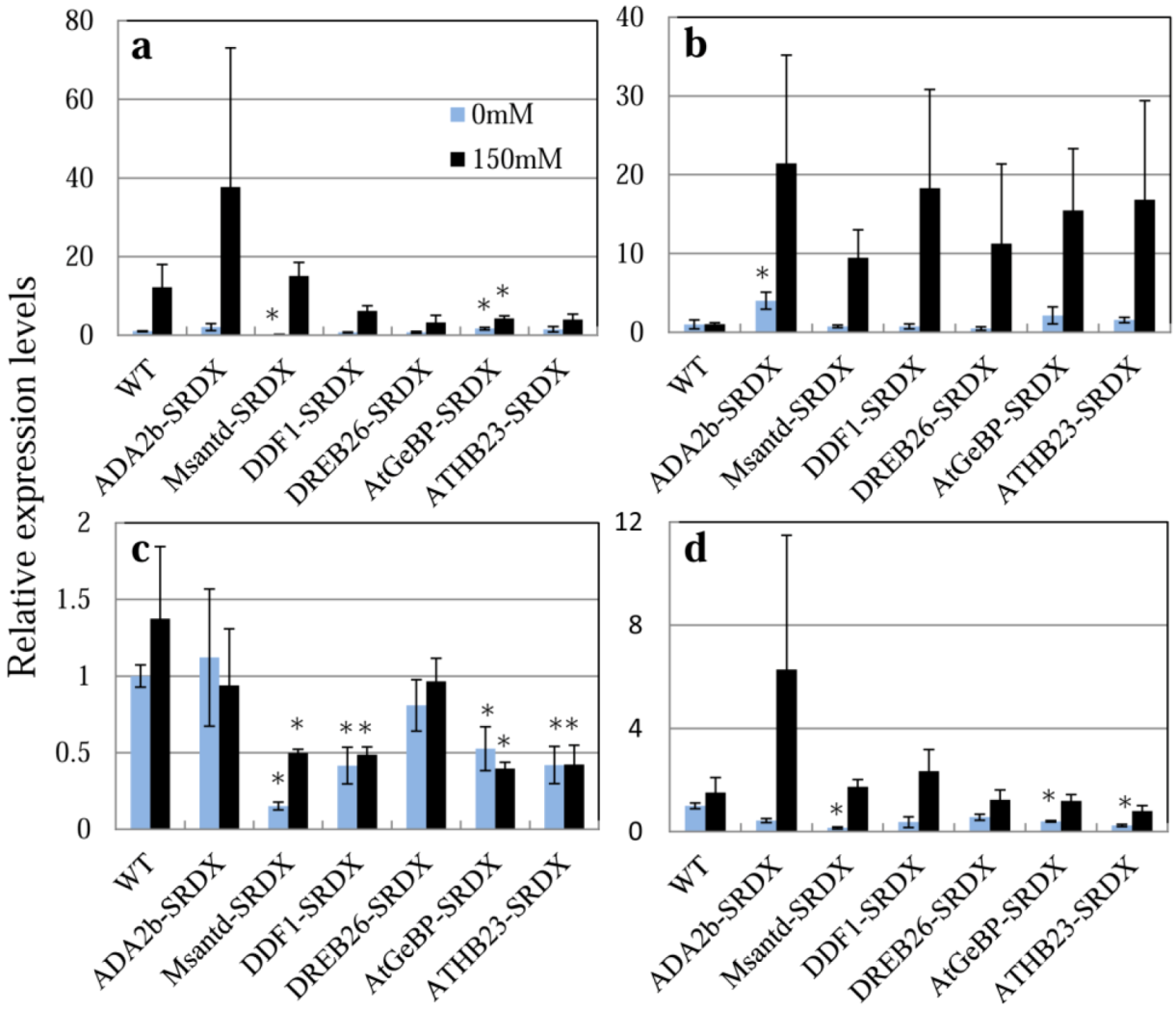
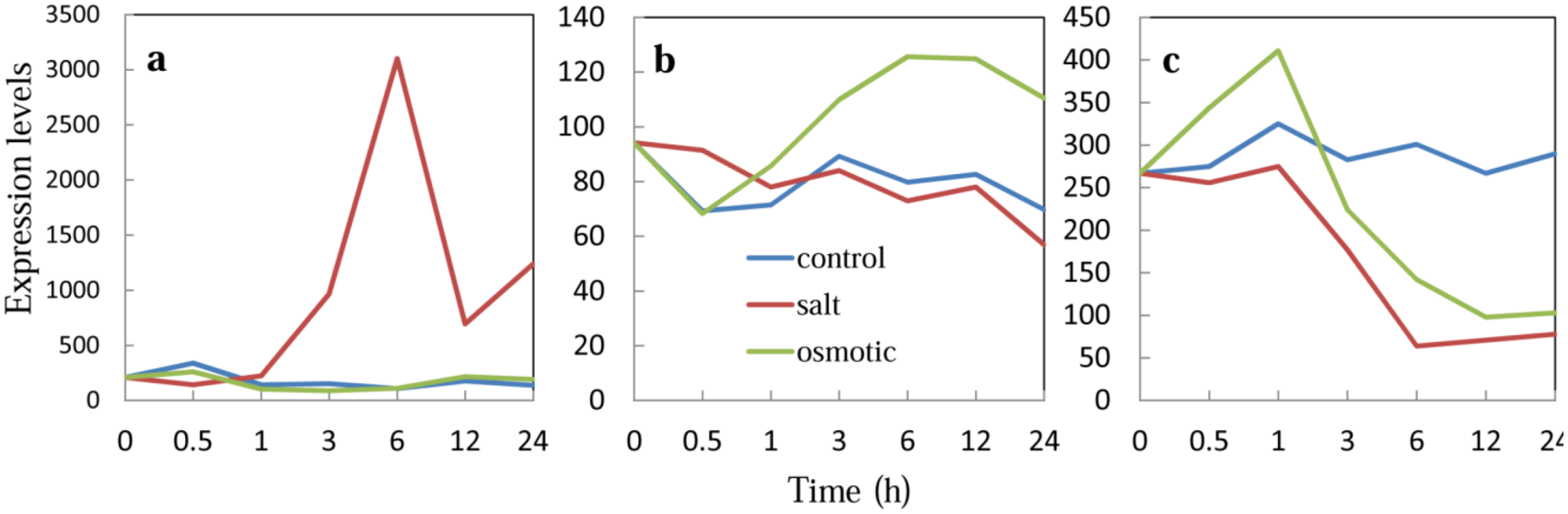
2.4. Expression of Transcription Factor, Chimeric Repressor and Stress-Related Genes in CRES-T Lines


3. Experimental
3.1. Generation and Selection of CRES-T Lines
3.2. Stress Tolerance Assay
3.3. Extraction of Gene Expression Profiles of Transcription Factors
3.4. Quantitative Real-Time RT-PCR and RT-PCR
| Genes | primers | Sequences |
|---|---|---|
| 35S-F | 5'-CGGATTCCCATTGCCCAGCTATCT-3' | |
| NOS-R | 5'-TAATCATCGCAAGACCGGCAACAG-3' | |
| RD29A | Forward | 5'-AAGAGGAAGTGAAAGGAGGAGGAGTCACGC-3' |
| Reverse | 5'-CACCACCAAACCAGCCAGA-3' | |
| RD22 | Forward | 5'-GTCTTCCTCTGATCTGTCTTCTTGG-3' |
| Reverse | 5'-TGGGAATGGGAGTGTTTGG-3' | |
| DREB1A | Forward | 5'-atatgcacgatgaggcgatg-3' |
| Reverse | 5'-tcatcatcgccgtcgacttc-3' | |
| P5CS | Forward | 5'-AGTCGGGGTCGAAGGATTAC-3' |
| Reverse | 5'-gcttggatgggaatgtcctg-3' | |
| UBQ5 | Forward | 5'-tgtgaaggcgaagatccaag-3' |
| Reverse | 5'-gagacggaggacgagatgaag-3' | |
| ADA2b | Forward | 5'-cggctcaaagatctcaaggaagc-3' |
| Reverse | 5'-ggctatatgcatccgacttcttcg-3' | |
| Msantd | Forward | 5'-gccatcaatgcggttgtgatgatcc-3' |
| Reverse | 5'-cactcagacaaccgatgagctcg-3' | |
| DDF1 | Forward | 5'-ggtaccaagaacagactgtactgc-3' |
| Reverse | 5'-tgaggagctgggattacacgtgtc-3' | |
| DREB26 | Forward | 5'-caacagctgaagcagctgctagagc-3' |
| Reverse | 5'-gcggaatgtcagcatcttcgtagaag-3' | |
| AtGeBP | Forward | 5'-cgtcaaacctcatgatcgcaaggc-3' |
| Reverse | 5'-agcctgcatcgctttccactgctcg-3' | |
| ATHB23 | Forward | 5'-agctagctcgtgccttgggattgc-3' |
| Reverse | 5'-tgctggtcaagccatggccagaac-3' | |
| SRDX | SRDX-R | 5'-ttaagcgaaacccaaacggagttctag-3' |
3.5. Statistical Analysis
4. Conclusions
Conflicts of Interest
References
- Apse, M.P.; Aharon, G.S.; Snedden, W.A.; Blumwald, E. Salt tolerance conferred by overexpression of a vacuolar Na+/H+ antiport in Arabidopsis. Science 1999, 285, 1256–1258. [Google Scholar] [CrossRef]
- Xu, D.; Duan, X.; Wang, B.; Hong, B.; Ho, T.; Wu, R. Expression of a late embryogenesis abundant protein gene, HVA1, from barley confers tolerance to water deficit and salt stress in transgenic rice. Plant Physiol. 1996, 110, 249–257. [Google Scholar]
- Gaxiola, R.A.; Li, J.; Undurraga, S.; Dang, L.M.; Allen, G.J.; Alper, S.L.; Fink, G.R. Drought- and salt-tolerant plants result from overexpression of the AVP1 H+-pump. Proc. Natl. Acad. Sci. USA 2001, 98, 11444–11449. [Google Scholar] [CrossRef]
- Shi, H.; Lee, B.H.; Wu, S.J.; Zhu, J.K. Overexpression of a plasma membrane Na+/H+ antiporter gene improves salt tolerance in Arabidopsis thaliana. Nat. Biotechnol. 2003, 21, 81–85. [Google Scholar]
- Agarwal, P.K.; Shukla, P.S.; Gupta, K.; Jha, B. Bioengineering for salinity tolerance in plants: State of the art. Mol. Biotechnol. 2012, 54, 102–123. [Google Scholar] [CrossRef]
- Kasuga, M.; Liu, Q.; Miura, S.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Improving plant drought, salt, and freezing tolerance by gene transfer of a single stress-inducible transcription factor. Nat. Biotechnol. 1999, 17, 287–291. [Google Scholar] [CrossRef]
- Huang, J.; Yang, X.; Wang, M.M.; Tang, H.J.; Ding, L.Y.; Shen, Y.; Zhang, H.S. A novel rice C2H2-type zinc finger protein lacking DLN-box/EAR-motif plays a role in salt tolerance. Biochim. Biophys. Acta 2007, 1769, 220–227. [Google Scholar] [CrossRef]
- Hu, H.; Dai, M.; Yao, J.; Xiao, B.; Li, X.; Zhang, Q.; Xiong, L. Overexpressing a NAM, ATAF, and CUC (NAC) transcription factor enhances drought resistance and salt tolerance in rice. Proc. Natl. Acad. Sci. USA 2006, 103, 12987–12992. [Google Scholar] [CrossRef]
- Hu, H.; You, J.; Fang, Y.; Zhu, X.; Qi, Z.; Xiong, L. Characterization of transcription factor gene SNAC2 conferring cold and salt tolerance in rice. Plant Mol. Biol. 2008, 67, 169–181. [Google Scholar] [CrossRef]
- Jung, C.; Seo, J.S.; Han, S.W.; Koo, Y.J.; Kim, C.H.; Song, S.I.; Nahm, B.H.; Choi, Y.D.; Cheong, J.J. Overexpression of atMYB44 enhances stomatal closure to confer abiotic stress tolerance in transgenic Arabidopsis. Plant Physiol. 2008, 146, 623–635. [Google Scholar]
- Fujita, M.; Mizukado, S.; Fujita, Y.; Ichikawa, T.; Nakazawa, M.; Seki, M.; Matsui, M.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Identification of stress-tolerance-related transcription-factor genes via mini-scale Full-length cDNA Over-eXpressor (FOX) gene hunting system. Biochem. Biophys. Res. Commun. 2007, 364, 250–257. [Google Scholar] [CrossRef]
- Novillo, F.; Alonso, J.M.; Ecker, J.R.; Salinas, J. CBF2/DREB1C is a negative regulator of CBF1/DREB1B and CBF3/DREB1A expression and plays a central role in stress tolerance in Arabidopsis. Proc. Natl. Acad. Sci. USA 2004, 101, 3985–3990. [Google Scholar] [CrossRef]
- Hiratsu, K.; Matsui, K.; Koyama, T.; Ohme-Takagi, M. Dominant repression of target genes by chimeric repressors that include the EAR motif, a repression domain, in Arabidopsis. Plant J. 2003, 34, 733–739. [Google Scholar] [CrossRef]
- Hiratsu, K.; Ohta, M.; Matsui, K.; Ohme-Takagi, M. The SUPERMAN protein is an active repressor whose carboxy-terminal repression domain is required for the development of normal flowers. FEBS Lett. 2002, 514, 351–354. [Google Scholar] [CrossRef]
- Mito, T.; Seki, M.; Shinozaki, K.; Ohme-Takagi, M.; Matsui, K. Generation of chimeric repressors that confer salt tolerance in Arabidopsis and rice. Plant Biotechnol. J. 2011, 9, 736–746. [Google Scholar] [CrossRef]
- Hark, A.T.; Vlachonasios, K.E.; Pavangadkar, K.A.; Rao, S.; Gordon, H.; Adamakis, I.D.; Kaldis, A.; Thomashow, M.F.; Triezenberg, S.J. Two Arabidopsis orthologs of the transcriptional coactivator ADA2 have distinct biological functions. Biochim. Biophys. Acta 2009, 1789, 117–124. [Google Scholar] [CrossRef]
- Anzola, J.M.; Sieberer, T.; Ortbauer, M.; Butt, H.; Korbei, B.; Weinhofer, I.; Müllner, A.E.; Luschnig, C. Putative Arabidopsis transcriptional adaptor protein (PROPORZ1) is required to modulate histone acetylation in response to auxin. Proc. Natl. Acad. Sci. USA 2010, 107, 10308–10313. [Google Scholar] [CrossRef]
- Magome, H.; Yamaguchi, S.; Hanada, A.; Kamiya, Y.; Oda, K. Dwarf and delayed-flowering 1, a novel Arabidopsis mutant deficient in gibberellin biosynthesis because of overexpression of a putative AP2 transcription factor. Plant J. 2004, 37, 720–729. [Google Scholar] [CrossRef]
- Magome, H.; Yamaguchi, S.; Hanada, A.; Kamiya, Y.; Oda, K. The DDF1 transcriptional activator upregulates expression of a gibberellin-deactivating gene, GA2ox7, under high-salinity stress in Arabidopsis. Plant J. 2008, 56, 613–626. [Google Scholar] [CrossRef]
- Libault, M.; Wan, J.; Czechowski, T.; Udvardi, M.; Stacey, G. Identification of 118 Arabidopsis transcription factor and 30 ubiquitin-ligase genes responding to chitin, a plant-defense elicitor. Mol. Plant Microbe Interact. 2007, 20, 900–911. [Google Scholar] [CrossRef]
- Krishnaswamy, S.; Verma, S.; Rahman, M.H.; Kav, N.N. Functional characterization of four APETALA2-family genes (RAP2.6, RAP2.6L, DREB19 and DREB26) in Arabidopsis. Plant Mol. Biol. 2011, 75, 107–127. [Google Scholar] [CrossRef]
- Curaba, J.; Herzog, M.; Vachon, G. GeBP, the first member of a new gene family in Arabidopsis, encodes a nuclear protein with DNA-binding activity and is regulated by KNAT1. Plant J. 2003, 33, 305–317. [Google Scholar] [CrossRef]
- Henriksson, E.; Olsson, A.S.; Johannesson, H.; Johansson, H.; Hanson, J.; Engstrom, P.; Soderman, E. Homeodomain leucine zipper class I genes in Arabidopsis. Expression patterns and phylogenetic relationships. Plant Physiol. 2005, 139, 509–518. [Google Scholar] [CrossRef]
- Tan, Q.K.; Irish, V.F. The Arabidopsis zinc finger-homeodomain genes encode proteins with unique biochemical properties that are coordinately expressed during floral development. Plant Physiol. 2006, 140, 1095–1108. [Google Scholar] [CrossRef]
- Kilian, J.; Whitehead, D.; Horak, J.; Wanke, D.; Weinl, S.; Batistic, O.; D’Angelo, B.-B.E.; Kudla, J.; Harter, K. The AtGenExpress global stress expression data set: Protocols, evaluation and model data analysis of UV-B light, drought and cold stress responses. Plant J. 2007, 50, 347–363. [Google Scholar] [CrossRef]
- Abe, H.; Yamaguchi-Shinozaki, K.; Urao, T.; Iwasaki, T.; Hosokawa, D.; Shinozaki, K. Role of Arabidopsis MYC and MYB homologs in drought- and abscisic acid-regulated gene expression. Plant Cell 1997, 9, 1859–1868. [Google Scholar]
- Yamaguchi-Shinozaki, K.; Shinozaki, K. A novel cis-acting element in an Arabidopsis gene is involved in responsiveness to drought, low-temperature, or high-salt stress. Plant Cell 1994, 6, 251–264. [Google Scholar]
- Liu, Q.; Kasuga, M.; Sakuma, Y.; Abe, H.; Miura, S.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Two transcription factors, DREB1 and DREB2, with an EREBP/AP2 DNA binding domain separate two cellular signal transduction pathways in drought- and low-temperature-responsive gene expression, respectively, in Arabidopsis. Plant Cell 1998, 10, 1391–1406. [Google Scholar]
- Delauney, A.; Verma, D.P.S. Proline biosynthesis and osmoregulation in plants. Plant J. 1993, 4, 215–223. [Google Scholar]
- Hare, P.D.; Cress, W.A.; van Staden, J. Proline synthesis and degradation: A model system for elucidating stress-related signal transduction. J. Exp. Bot. 1999, 50, 413–434. [Google Scholar]
- Miyama, M.; Tada, Y. Expression of Bruguiera gymnorhiza BgARP1 enhances salt tolerance in transgenic Arabidopsis plants. Euphytica 2011, 177, 383–392. [Google Scholar] [CrossRef]
- Gilmour, S.J.; Sebolt, A.M.; Salazar, M.P.; Everard, J.D.; Thomashow, M.F. Overexpression of the Arabidopsis CBF3 transcriptional activator mimics multiple biochemical changes associated with cold acclimation. Plant Physiol. 2000, 124, 1854–1865. [Google Scholar] [CrossRef]
- Xiong, Y.; Fei, S.Z. Functional and phylogenetic analysis of a DREB⁄CBF-like gene in perennial ryegrass (Lolium perenne L.). Planta 2006, 224, 878–888. [Google Scholar] [CrossRef]
- Seong, E.S.; Cho, H.S.; Choi, D.; Joung, Y.H.; Lim, C.K.; Hur, J.H.; Wang, M.H. Tomato plants overexpressing caKR1 enhanced tolerance to salt and oxidative stress. Biochem. Biophys. Res. Commun. 2007, 363, 983–988. [Google Scholar] [CrossRef]
- Miyama, M.; Tada, Y. Transcriptional and physiological study of the response of Burma mangrove (Bruguiera gymnorhiza) to salt and osmotic stress. Plant Mol. Biol. 2008, 68, 119–129. [Google Scholar] [CrossRef]
Appendix





© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Kazama, D.; Itakura, M.; Kurusu, T.; Mitsuda, N.; Ohme-Takagi, M.; Tada, Y. Identification of Chimeric Repressors that Confer Salt and Osmotic Stress Tolerance in Arabidopsis. Plants 2013, 2, 769-785. https://doi.org/10.3390/plants2040769
Kazama D, Itakura M, Kurusu T, Mitsuda N, Ohme-Takagi M, Tada Y. Identification of Chimeric Repressors that Confer Salt and Osmotic Stress Tolerance in Arabidopsis. Plants. 2013; 2(4):769-785. https://doi.org/10.3390/plants2040769
Chicago/Turabian StyleKazama, Daisuke, Masateru Itakura, Takamitsu Kurusu, Nobutaka Mitsuda, Masaru Ohme-Takagi, and Yuichi Tada. 2013. "Identification of Chimeric Repressors that Confer Salt and Osmotic Stress Tolerance in Arabidopsis" Plants 2, no. 4: 769-785. https://doi.org/10.3390/plants2040769
APA StyleKazama, D., Itakura, M., Kurusu, T., Mitsuda, N., Ohme-Takagi, M., & Tada, Y. (2013). Identification of Chimeric Repressors that Confer Salt and Osmotic Stress Tolerance in Arabidopsis. Plants, 2(4), 769-785. https://doi.org/10.3390/plants2040769