From Plant Infectivity to Growth Patterns: The Role of Blue-Light Sensing in the Prokaryotic World

Abstract
:1. Introduction
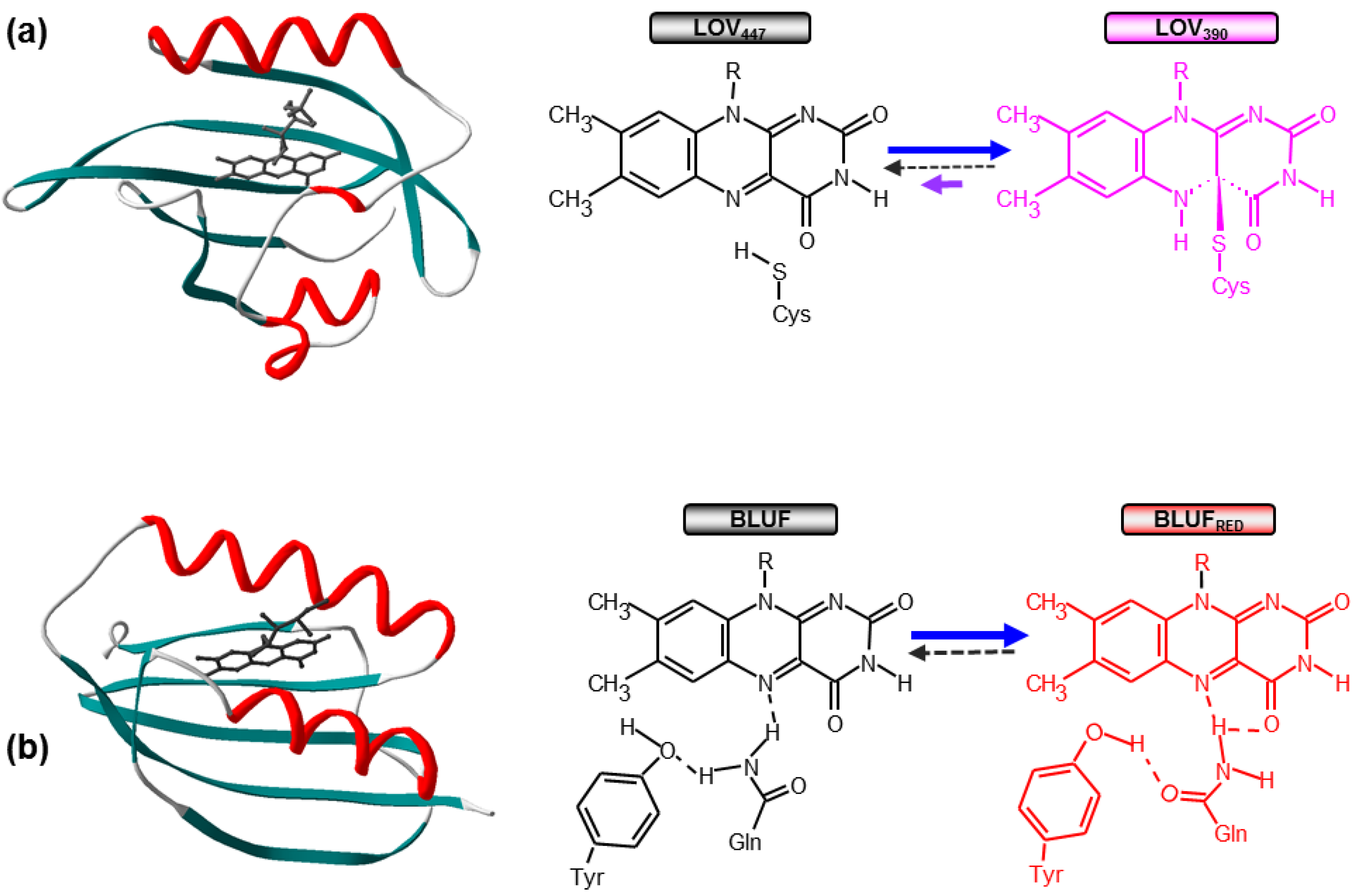
1.1. Photoactivation of LOV and BLUF Domains
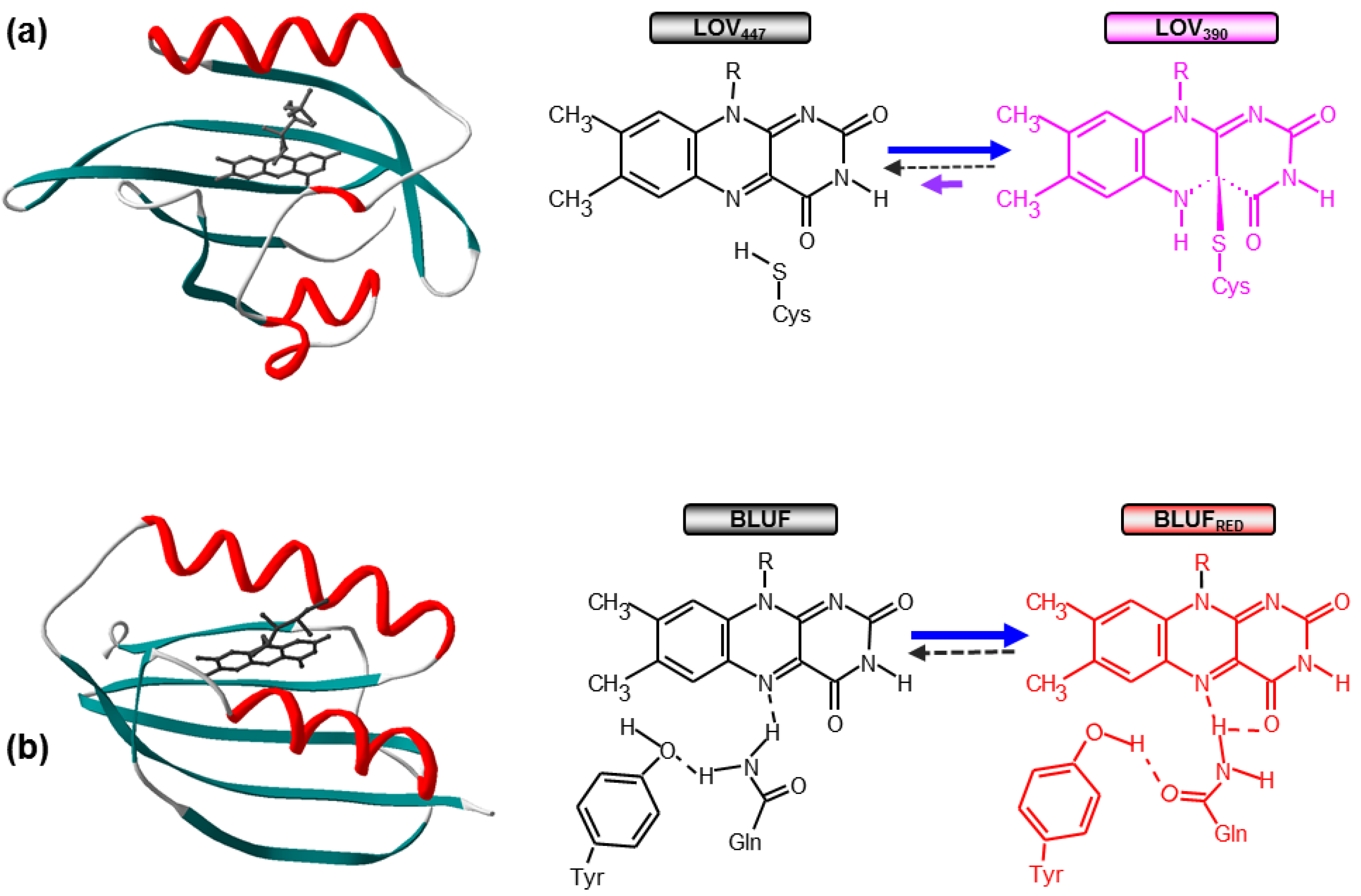
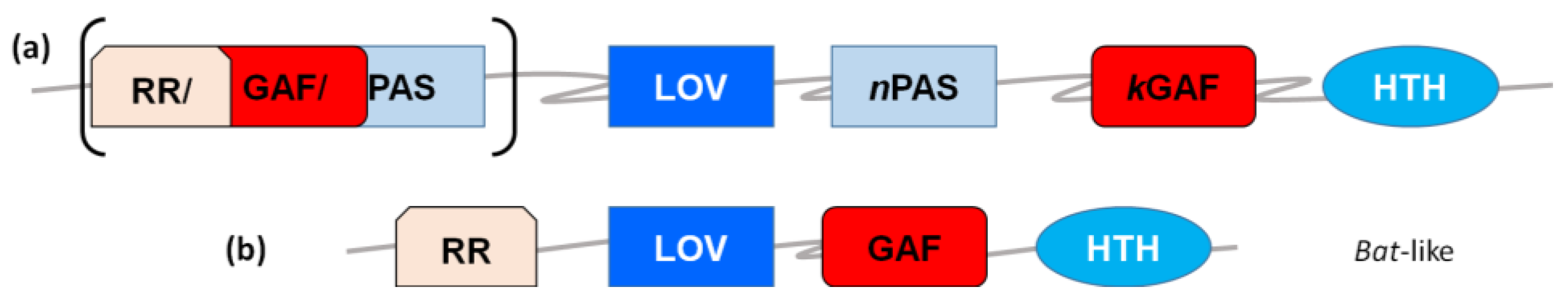
1.2. From Photochemistry to Signaling: Structural Issues and Signal Propagation Mechanisms
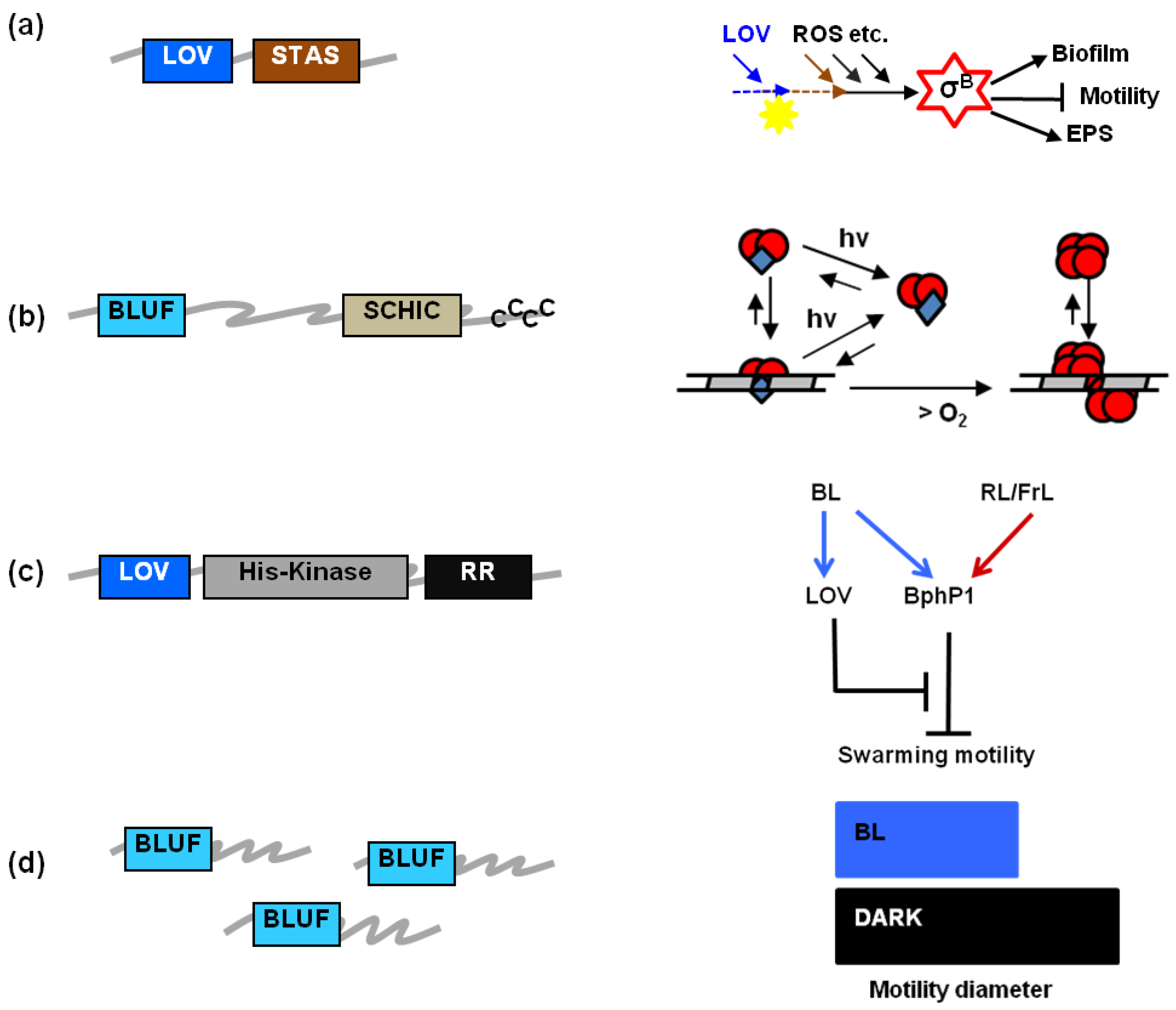
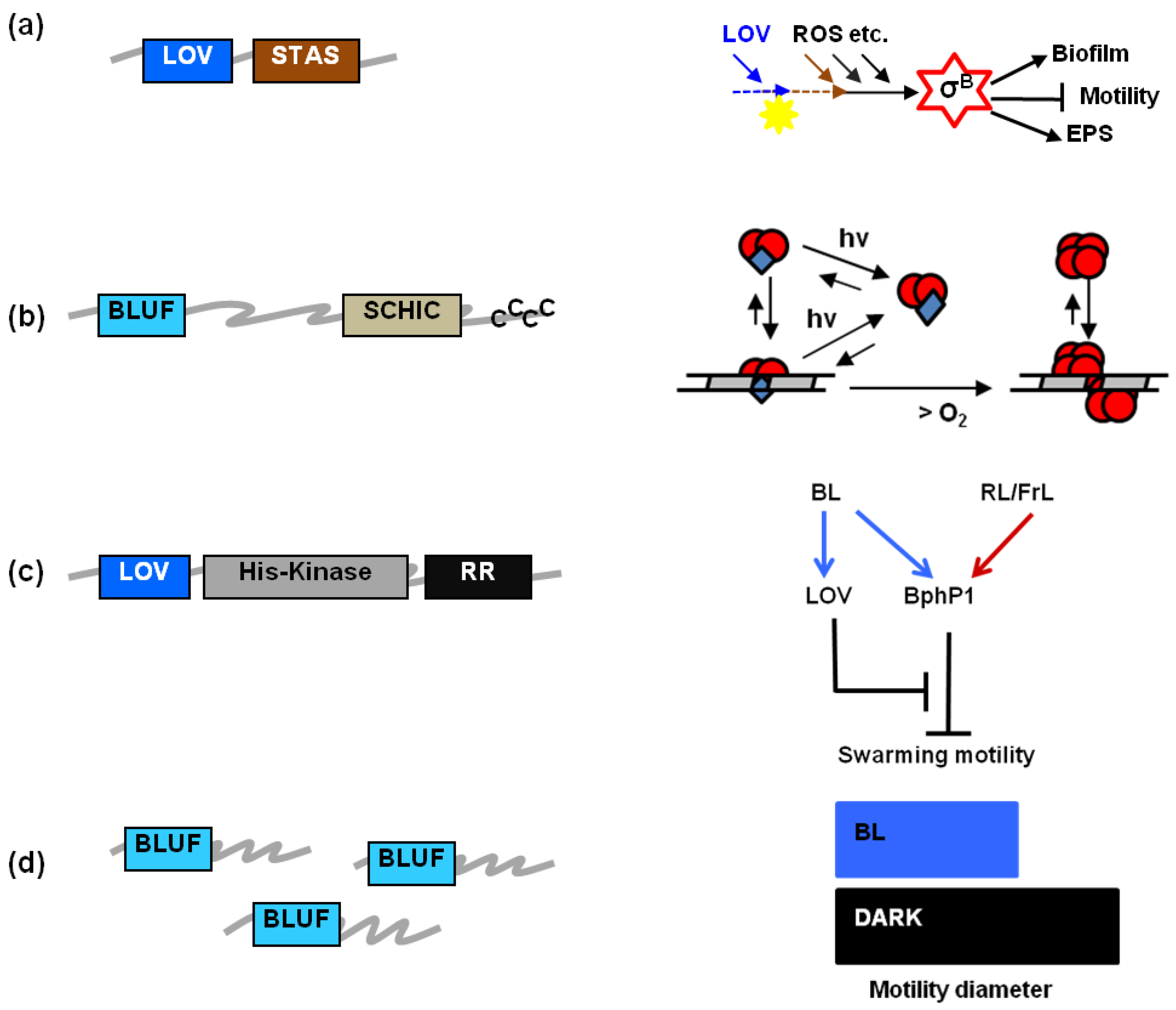
1.3. The Significance of Prokaryotic LOV and BLUF Proteins in Vivo

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| LOV protein | Blue light-regulated phenomena | |
|---|---|---|
| B. subtilis | BsYtvA | Activation of σB stress factor a [11,12,31,56] |
| B. amyloliquefaciens | YtvA-like? | Antifungal lipopeptide synthesis a [59] |
| L. monocytogenes | Lmo0799 (YtvA-like) | σB–mediated invasiveness, swimming motility, salt stress a [61]; colony differentiation [1] |
| R. leguminosarum | LOV-HK | Biofilm; EPS b; flagella; proliferation within roots; competence for N2 fixing [69] |
| C. crescentus | LovK | Cell-cell adhesion [71,72] c |
| E. litoralis | EL222 | Binding to DNA consensus sequences [73] |
| B. abortus | LOV-HK | Infectivity [63] |
| X. axonopodis pv. citri | XacLOV | Adhesion; virulence [65] |
| X. campestris pv. campestris | Growth [66] | |
| P. syringae pv. tomato | PstLOV | Growth; motility (swarming); adhesion; virulence; invasiveness; σ factor gene expression [67,68] |
| P. syringae pv. syringae | PssLOV | Motility (swarming) d [4] |
| R. sphaeroides | RsLOV | Genes for photosynthesis and photo-oxidative stress under σF control [74] e |
| BLUF protein | Blue light-regulated phenomena | |
|---|---|---|
| R. sphaeroides | RsAppA | Photosynthesis gene transcription; [37]; integration of light and redox sensing a [3,75,76] |
| Synechocystis sp. PCC6803 | PixD/Slr1694 | Phototaxis [33] |
| Acinetobacter | BLUF | Surface motility; biofilm [64] |
| A. baumannii | BLUF | Virulence; surface motility; biofilm b [9] |
| A. baylyi ADP1 | BLUF | Surface motility; biofilm b [5,64] |
| R. palustris | PapB | Biofilm [70] |
| E. coli | YcgF | Biofilm b [53,77] |
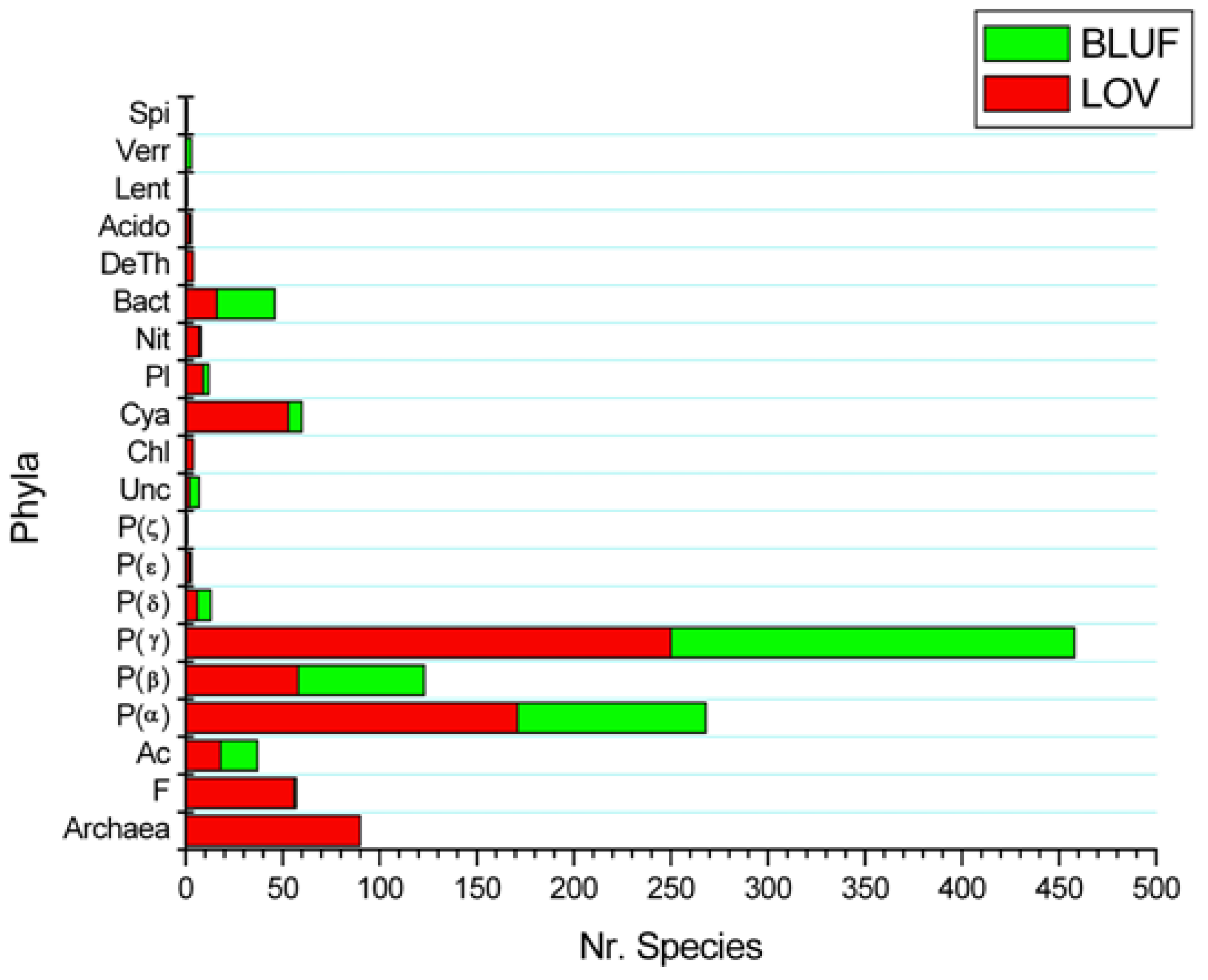
1.4. Distribution and Evolutionary Patterns of Prokaryotic LOV and BLUF Domains
2. Results and Discussion
2.1. The Sequence Logos and Searching Patterns
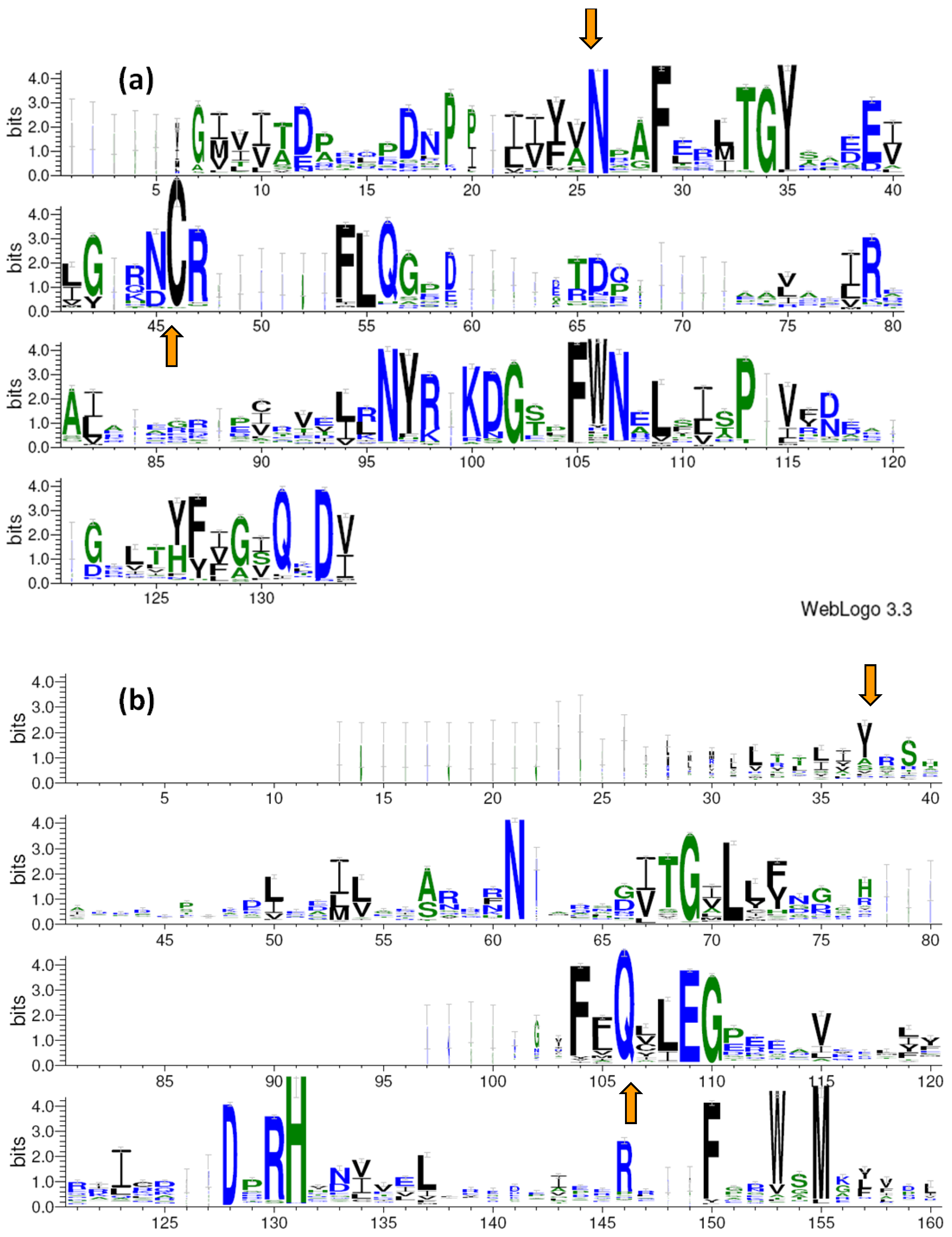
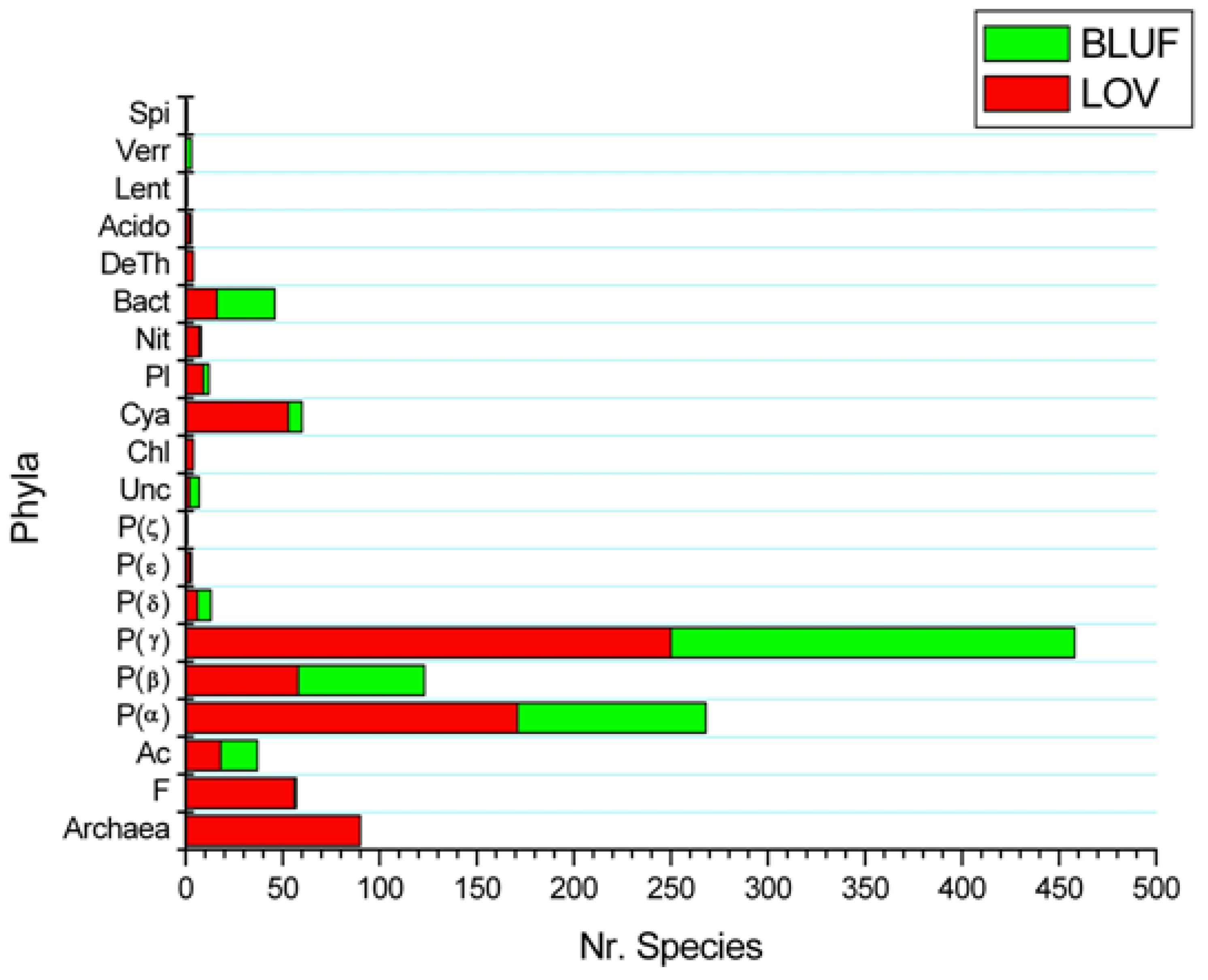
2.2. LOV and BLUF Domains in Bacteria
| Class (phylum)/order | LOV proteins | BLUF proteins | |
|---|---|---|---|
| Aureimonas ureilytica | P(α)/Rhizobiales | 2 × LOV-HK | 2 × short-BLUF |
| Methylobacterium populi a | P(α)/Rhizobiales | 7 × LOV-HK 1 × short LOV | 2 × short-BLUF |
| Sphingobium xenophagum | P(α)/Sphingomonadales | 2 × LOV-RR 1 × LOV-HK | 2 × short-BLUF |
| Herminiimonas arsenicoxydans | P(β)/Burkholderiales | 3 × LOV-GGDEF/EAL | 1 × short-BLUF |
| Methylomicrobium alcaliphilum | P(γ)/Methylococcales | 2 × short LOV 1 × LOV-HK | 3 × short-BLUF 1 × BLUF-GGDEF/EAL |
2.3. LOV in Archaea: DNA-Binding Proteins and Kinases
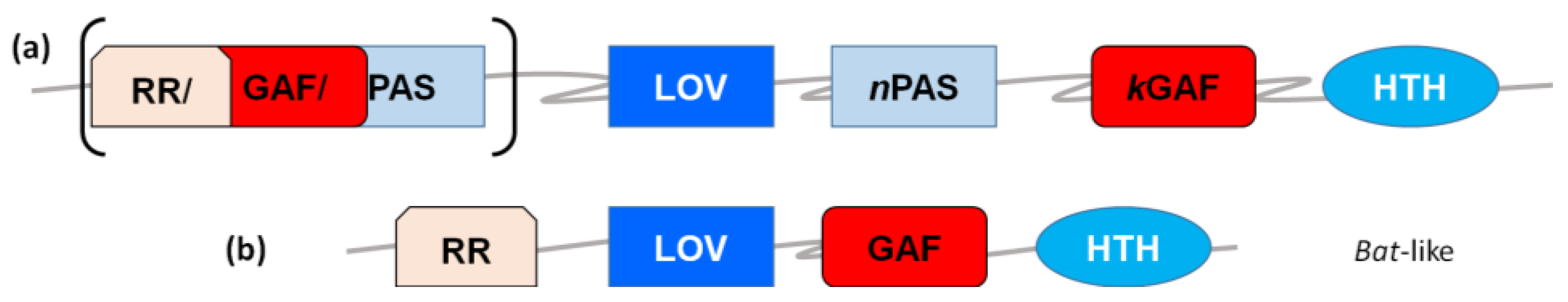

3. Experimental
4. Conclusions
Supplementary Files
Supplementary File 1Acknowledgments
Conflicts of Interest
References
- Tiensuu, T.; Andersson, C.; Rydén, P.; Johansson, J. Cycles of light and dark coordinate reversible colony differentiation in Listeria monocytogenes. Mol. Microbiol. 2013, 87, 909–924. [Google Scholar] [CrossRef]
- Yin, L.; Dragnea, V.; Feldman, G.; Hammad, L.A.; Karty, J.A.; Dann, C.E.; Bauer, C.E. Redox and light control the heme-sensing activity of AppA. MBio 2013, 4. [Google Scholar] [CrossRef]
- Winkler, A.; Heintz, U.; Lindner, R.; Reinstein, J.; Shoeman, R.L.; Schlichting, I. A ternary AppA-PpsR-DNA complex mediates light regulation of photosynthesis-related gene expression. Nat. Struct. Mol. Biol. 2013, 20, 859–867. [Google Scholar] [CrossRef]
- Wu, L.; McGrane, R.S.; Beattie, G.A. Light regulation of swarming motility in Pseudomonas syringae integrates signaling pathways mediated by a bacteriophytochrome and a LOV protein. MBio 2013, 4, e00334-13. [Google Scholar]
- Bitrian, M.; González, R.M.; Paris, G.; Hellingwerf, K.J.; Nudel, C.B. Blue-light-dependent inhibition of twitching motility in Acinetobacter baylyi ADP1: Additive involvement of three BLUF-domain-containing proteins. Microbiology 2013, 159, 1828–1841. [Google Scholar] [CrossRef] [Green Version]
- Mandalari, C.; Losi, A.; Gärtner, W. Distance-tree analysis, distribution and co-presence of bilin- and flavin-binding prokaryotic photoreceptors for visible light. Photochem. Photobiol. Sci. 2013, 12, 1144–1157. [Google Scholar] [CrossRef]
- Kennis, J.T.M.; Mathes, T. Molecular eyes: Proteins that transform light into biological information. Interface Focus 2013, 3, 20130005. [Google Scholar] [CrossRef]
- Nakasone, Y.; Ono, T.; Ishii, A.; Masuda, S.; Terazima, M. Temperature-sensitive reaction of a photosensor protein YcgF: Possibility of a role of temperature sensor. Biochemistry 2010, 49, 2288–2296. [Google Scholar] [CrossRef]
- Mussi, M.A.; Gaddy, J.A.; Cabruja, M.; Arivett, B.A.; Viale, A.M.; Rasia, R.; Actis, L.A. The opportunistic human pathogen Acinetobacter baumannii senses and responds to light. J. Bacteriol. 2010, 192, 6336–6345. [Google Scholar] [CrossRef]
- Purcell, E.B.; McDonald, C.A.; Palfey, B.A.; Crosson, S. An analysis of the solution structure and signaling mechanism of LovK, a sensor histidine kinase integrating light and redox signals. Biochemistry 2010, 49, 6761–6770. [Google Scholar] [CrossRef]
- Avila-Perez, M.; Hellingwerf, K.J.; Kort, R. Blue light activates the sigmaB-dependent stress response of Bacillus subtilis via YtvA. J. Bacteriol. 2006, 188, 6411–6414. [Google Scholar] [CrossRef]
- Avila-Perez, M.; Vreede, J.; Tang, Y.; Bende, O.; Losi, A.; Gaertner, W.; Hellingwerf, K. In vivo mutational analysis of YTVA from Bacillus subtilis: Mechanism of light-activation of the general stress response. J. Biol. Chem. 2009, 284, 24958–24964. [Google Scholar] [CrossRef]
- Losi, A.; Gärtner, W. The evolution of flavin-binding photoreceptors: An ancient chromophore serving trendy blue-light sensors. Annu. Rev. Plant Biol. 2012, 63, 49–72. [Google Scholar] [CrossRef]
- Pathak, G.P.; Vrana, J.D.; Tucker, C.L. Optogenetic control of cell function using engineered photoreceptors. Biol. Cell 2013, 105, 59–72. [Google Scholar] [CrossRef]
- Drepper, T.; Gensch, T.; Pohl, M. Advanced in vivo applications of blue light photoreceptors as alternative fluorescent proteins. Photochem. Photobiol. Sci. 2013, 12, 1125–1134. [Google Scholar] [CrossRef]
- Sancar, A. Photolyase and cryptochrome blue-light photoreceptors. Adv. Protein Chem. 2004, 69, 73–100. [Google Scholar] [CrossRef]
- Chaves, I.; Pokorny, R.; Byrdin, M.; Hoang, N.; Ritz, T.; Brettel, K.; Essen, L.O.; van der Horst, G.T.J.; Batschauer, A.; Ahmad, M. The cryptochromes: Blue light photoreceptors in plants and animals. Annu. Rev. Plant Biol. 2011, 62, 335–364. [Google Scholar] [CrossRef]
- Weber, S. Light-driven enzymatic catalysis of DNA repair: A review of recent biophysical studies on photolyase. Biochim. Biophys. Acta 2005, 1707, 1–23. [Google Scholar]
- Geisselbrecht, Y.; Fruehwirth, S.W.; Schroeder, C.; Pierik, A.; Klug, G.; Essen, L.-O. CryB from Rhodobacter sphaeroides: A unique class of cryptochromes with new cofactors. EMBO Rep. 2012, 13, 223–229. [Google Scholar] [CrossRef]
- Biskup, T. Time-resolved EPR of radical pair intermediates in cryptochromes. Mol. Phys. 2013, 111, 3698–3703. [Google Scholar] [CrossRef]
- Liu, Z.; Zhang, M.; Guo, X.; Tan, C.; Li, J.; Wang, L.; Sancar, A.; Zhong, D. Dynamic determination of the functional state in photolyase and the implication for cryptochrome. Proc. Natl. Acad. Sci. USA 2013, 110, 12972–12977. [Google Scholar] [CrossRef]
- Kritsky, M.S.; Telegina, T.A.; Vechtomova, Y.L.; Kolesnikov, M.P.; Lyudnikova, T.A.; Golub, O.A. Excited flavin and pterin coenzyme molecules in evolution. Biochemistry (Mosc.) 2010, 75, 1200–1216. [Google Scholar]
- Bauer, C.; Rabl, C.R.; Heberle, J.; Kottke, T. Indication for a radical intermediate preceding the signaling state in the LOV domain photocycle. Photochem. Photobiol. 2011, 87, 548–553. [Google Scholar] [CrossRef]
- Song, S.H.; Freddolino, P.L.; Nash, A.I.; Carroll, E.C.; Schulten, K.; Gardner, K.H.; Larsen, D.S. Modulating LOV domain photodynamics with a residue alteration outside the chromophore binding site. Biochemistry 2011, 50, 2411–2423. [Google Scholar] [CrossRef]
- Raffelberg, S.; Mansurova, M.; Gärtner, W.; Losi, A. Modulation of the photocycle of a LOV domain photoreceptor by the hydrogen bonding network. J. Am. Chem. Soc. 2011, 133, 5346–5356. [Google Scholar] [CrossRef]
- Raffelberg, S.; Gutt, A.; Gärtner, W.; Mandalari, C.; Abbruzzetti, S.; Viappiani, C.; Losi, A. The amino acids surrounding the flavin 7a-methyl group determine the UVA spectral features of a LOV protein. Biol. Chem. 2013, 394, 1517–1528. [Google Scholar]
- Losi, A. Flavin-based blue-light Photosensors: A photobiophysics update. Photochem. Photobiol. 2007, 83, 1283–1300. [Google Scholar] [CrossRef]
- Rani, R.; Jentzsch, K.; Lecher, J.; Hartmann, R.; Willbold, D.; Jaeger, K.E.; Krauss, U. Conservation of dark recovery kinetic parameters and structural features in the pseudomonadaceae “short” Light, Oxygen, Voltage (LOV) protein family: Implications for the design of LOV-based optogenetic tools. Biochemistry 2013, 52, 4460–4473. [Google Scholar] [CrossRef]
- Losi, A.; Gärtner, W.; Raffelberg, S.; Cella Zanacchi, F.; Bianchini, P.; Diaspro, A.; Mandalari, C.; Abbruzzetti, S.; Viappiani, C. A photochromic bacterial photoreceptor with potential for super-resolution microscopy. Photochem. Photobiol. Sci. 2013, 12, 231–235. [Google Scholar]
- Song, S.H.; Madsen, D.; van der Steen, J.B.; Pullman, R.; Freer, L.H.; Hellingwerf, K.J.; Larsen, D.S. Primary photochemistry of the dark- and light-adapted states of the YtvA protein from Bacillus subtilis. Biochemistry 2013, 52, 7951–7963. [Google Scholar] [CrossRef]
- Van der Steen, J.B.; Nakasone, Y.; Hendriks, J.; Hellingwerf, K.J. Modeling the functioning of YtvA in the general stress response in Bacillus subtilis. Mol. BioSyst. 2013, 9, 2331–2343. [Google Scholar] [CrossRef]
- Losi, A.; Polverini, E.; Quest, B.; Gärtner, W. First evidence for phototropin-related blue-light receptors in prokaryotes. Biophys. J. 2002, 82, 2627–2634. [Google Scholar] [CrossRef]
- Masuda, S. Light detection and signal transduction in the BLUF photoreceptors. Plant Cell Physiol. 2013, 54, 171–179. [Google Scholar] [CrossRef]
- Gauden, M.; van Stokkum, I.H.M.; Key, J.M.; Luhrs, D.C.; van Grondelle, R.; Hegemann, P.; Kennis, J.T.M. Hydrogen-bond switching through a radical pair mechanism in a flavin-binding photoreceptor. Proc. Natl. Acad. Sci. USA 2006, 103, 10895–10900. [Google Scholar] [CrossRef]
- Khrenova, M.G.; Nemukhin, A.V.; Domratcheva, T. Photoinduced electron transfer facilitates tautomerization of the conserved signaling glutamine side chain in BLUF protein light sensors. J. Phys. Chem. B 2013, 117, 2369–2377. [Google Scholar] [CrossRef]
- Arents, J.C.; Perez, M.A.; Hendriks, J.; Hellingwerf, K.J. On the midpoint potential of the FAD chromophore in a BLUF-domain containing photoreceptor protein. FEBS Lett. 2011, 585, 167–172. [Google Scholar] [CrossRef]
- Masuda, S.; Bauer, C.E. AppA is a blue light photoreceptor that antirepresses photosynthesis gene expression in Rhodobacter sphaeroides. Cell 2002, 110, 613–623. [Google Scholar] [CrossRef]
- Herrou, J.; Crosson, S. Function, structure and mechanism of bacterial photosensory LOV proteins. Nat. Rev. Microbiol. 2011, 9, 713–723. [Google Scholar] [CrossRef]
- Hoch, J.A. Two-component and phosphorelay signal transduction. Curr. Opin. Microbiol. 2000, 3, 165–170. [Google Scholar] [CrossRef]
- Sondermann, H.; Shikuma, N.J.; Yildiz, F.H. You’ve come a long way: c-di-GMP signaling. Curr. Opin. Microbiol. 2012, 15, 140–146. [Google Scholar] [CrossRef]
- Krauss, U.; Losi, A.; Gärtner, W.; Jaeger, K.-E.; Eggert, T. Initial characterization of a blue-light sensing, phototropin-related protein from Pseudomonas putida: A paradigm for an extended LOV construct. Phys. Chem. Chem. Phys. 2005, 7, 2229–2236. [Google Scholar] [CrossRef]
- Losi, A.; Gartner, W. Old chromophores, new photoactivation paradigms, trendy applications: Flavins in blue light-sensing photoreceptors. Photochem. Photobiol. 2011, 87, 491–510. [Google Scholar] [CrossRef]
- Zoltowski, B.D.; Gardner, K.H. Tripping the light fantastic: Blue-light photoreceptors as examples of environmentally modulated protein-protein interactions. Biochemistry 2011, 50, 4–16. [Google Scholar] [CrossRef]
- Harper, S.M.; Neil, L.C.; Gardner, K.H. Structural basis of a phototropin light switch. Science 2003, 301, 1541–1544. [Google Scholar] [CrossRef]
- Kim, B.; Lin, M.Z. Optobiology: Optical control of biological processes via protein engineering. Biochem. Soc. Trans. 2013, 41, 1183–1188. [Google Scholar] [CrossRef]
- Möglich, A.; Moffat, K. Structural basis for light-dependent signaling in the dimeric LOV domain of the photosensor YtvA. J. Mol. Biol. 2007, 373, 112–126. [Google Scholar] [CrossRef]
- Jurk, M.; Dorn, M.; Kikhney, A.; Svergun, D.; Gärtner, W.; Schmieder, P. The switch that does not flip: The blue-light receptor YtvA from Bacillus subtilis adopts an elongated dimer conformation independent of the activation state as revealed by a combined AUC and SAXS study. J. Mol. Biol. 2010, 403, 78–87. [Google Scholar] [CrossRef]
- Möglich, A.; Moffat, K. Engineered photoreceptors as novel optogenetic tools. Photochem. Photobiol. Sci. 2010, 9, 1286–1300. [Google Scholar] [CrossRef]
- Diensthuber, R.P.; Bommer, M.; Gleichmann, T.; Möglich, A. Full-length structure of a sensor histidine kinase pinpoints coaxial coiled coils as signal transducers and modulators. Structure 2013, 21, 1127–1136. [Google Scholar] [CrossRef]
- Zoltowski, B.D.; Motta-Mena, L.B.; Gardner, K.H. Blue light-induced dimerization of a bacterial LOV-HTH DNA-binding protein. Biochemistry 2013, 52, 6653–6661. [Google Scholar] [CrossRef]
- Nash, A.I.; McNulty, R.; Shillito, M.E.; Swartz, T.E.; Bogomolni, R.A.; Luecke, H.; Gardner, K.H. Structural basis of photosensitivity in a bacterial light-oxygen-voltage/helix-turn-helix (LOV-HTH) DNA-binding protein. Proc. Natl. Acad. Sci. USA 2011, 108, 9449–9454. [Google Scholar]
- Barends, T.R.M.; Hartmann, E.; Griese, J.J.; Beitlich, T.; Kirienko, N.V.; Ryjenkov, D.A.; Reinstein, J.; Shoeman, R.L.; Gomelsky, M.; Schlichting, I. Structure and mechanism of a bacterial light-regulated cyclic nucleotide phosphodiesterase. Nature 2009, 459, 1015–1018. [Google Scholar] [CrossRef]
- Tschowri, N.; Busse, S.; Hengge, R. The BLUF-EAL protein YcgF acts as a direct anti-repressor in a blue-light response of Escherichia coli. Gene Dev. 2009, 23, 522–534. [Google Scholar] [CrossRef]
- Strickland, D.; Yao, X.L.; Gawlak, G.; Rosen, M.K.; Gardner, K.H.; Sosnick, T.R. Rationally improving LOV domain-based photoswitches. Nat. Methods 2010, 7, 623–626. [Google Scholar] [CrossRef]
- Correa, F.; Ko, W.H.; Ocasio, V.; Bogomolni, R.A.; Gardner, K.H. Blue Light regulated two-component systems: Enzymatic and functional analyses of Light-Oxygen-Voltage (LOV)-histidine kinases and downstream response regulators. Biochemistry 2013, 52, 4656–4666. [Google Scholar] [CrossRef]
- Jurk, M.; Schramm, P.; Schmieder, P. The blue-light receptor YtvA from Bacillus subtilis is permanently incorporated into the stressosome independent of the illumination state. Biochem. Biophys. Res. Commun. 2013, 432, 499–503. [Google Scholar] [CrossRef]
- Akbar, S.; Gaidenko, T.A.; Min, K.; O’Reilly, M.; Devine, K.M.; Price, C.W. New family of regulators in the environmental signaling pathway which activates the general stress transcription factor of Bacillus subtilis. J. Bacteriol. 2001, 183, 1329–1338. [Google Scholar] [CrossRef]
- Gaidenko, T.A.; Kim, T.J.; Weigel, A.L.; Brody, M.S.; Price, C.W. The blue-light receptor YtvA acts in the environmental stress signaling pathway of Bacillus subtilis. J. Bacteriol. 2006, 188, 6387–6395. [Google Scholar] [CrossRef]
- Ramkumar, G.; Yu, S.M.; Lee, Y.H. Influence of light qualities on antifungal lipopeptide synthesis in Bacillus amyloliquefaciens JBC36. Eur. J. Plant Pathol. 2013, 137, 243–248. [Google Scholar] [CrossRef]
- Ogata, H.; Cao, Z.; Losi, A.; Gaertner, W. Crystallization and preliminary X-ray analysis of the LOV domain of the blue-light receptor YtvA from Bacillus amyloliquefaciens FZB42. Acta Cryst. 2009, F65, 853–855. [Google Scholar]
- Ondrusch, N.; Kreft, J. Blue and red light modulates SigB-dependent gene transcription, swimming motility and invasiveness in Listeria monocytogenes. PLoS One 2011, 6, e16151. [Google Scholar] [CrossRef]
- Chan, R.H.; Lewis, J.W.; Bogomolni, R.A. Photocycle of the LOV-STAS protein from the pathogen Listeria monocytogenes. Photochem. Photobiol. 2013, 89, 361–369. [Google Scholar] [CrossRef]
- Swartz, T.E.; Tseng, T.S.; Frederickson, M.A.; Paris, G.; Comerci, D.J.; Rajashekara, G.; Kim, J.G.; Mudgett, M.B.; Splitter, G.A.; Ugalde, R.A.; et al. Blue-light-activated histidine kinases: Two-component sensors in bacteria. Science 2007, 317, 1090–1093. [Google Scholar] [CrossRef]
- Golic, A.; Vaneechoutte, M.; Nemec, A.; Viale, A.M.; Actis, L.A.; Mussi, M.A. Staring at the cold sun: Blue light regulation is distributed within the genus Acinetobacter. PLoS One 2013, 8, e55059. [Google Scholar]
- Kraiselburd, I.; Alet, A.I.; Tondo, M.L.; Petrocelli, S.; Daurelio, L.D.; Monzón, J.; Ruiz, O.A.; Losi, A.; Orellano, E.G. A LOV protein modulates the physiological attributes of Xanthomonas axonopodis pv. citri relevant for host plant colonization. PLoS One 2012, 7, e38226. [Google Scholar] [CrossRef]
- Mao, D.; Tao, J.; Li, C.; Luo, C.; Zheng, L.; He, C. Light signalling mediated by Per-ARNT-Sim domain-containing proteins in Xanthomonas campestris pv. campestris. FEMS Microbiol. Lett. 2012, 326, 31–39. [Google Scholar] [CrossRef]
- Moriconi, V.; Sellaro, R.; Ayub, N.; Soto, G.; Rugnone, M.; Shah, R.; Pathak, P.; Gärtner, W.; Casal, J.J. LOV-domain photoreceptor, encoded in a genomic island, attenuates the virulence of Pseudomonas syringae in light-exposed Arabidopsis leaves. Plant J. 2013, 76, 322–331. [Google Scholar]
- Río-Álvarez, I.; Rodríguez-Herva, J.J.; Martínez, P.M.; González-Melendi, P.; García-Casado, G.; Rodríguez-Palenzuela, P.; López-Solanilla, E. Light regulates motility, attachment and virulence in the plant pathogen Pseudomonas syringae pv tomato DC3000. Environ. Microbiol. 2013. [Google Scholar] [CrossRef]
- Bonomi, H.R.; Posadas, D.M.; Paris, G.; Carrica, M.D.; Frederickson, M.; Pietrasanta, L.I.; Bogomolni, R.A.; Zorreguieta, A.; Goldbaum, F.A. Light regulates attachment, exopolysaccharide production, and nodulation in Rhizobium leguminosarum through a LOV-histidine kinase photoreceptor. Proc. Natl. Acad. Sci. USA 2012, 109, 12135–12140. [Google Scholar] [CrossRef]
- Kanazawa, T.; Ren, S.; Maekawa, M.; Hasegawa, K.; Arisaka, F.; Hyodo, M.; Hayakawa, Y.; Ohta, H.; Masuda, S. Biochemical and physiological characterization of a BLUF protein-EAL protein complex involved in blue light-dependent degradation of cyclic diguanylate in the purple bacterium Rhodopseudomonas palustris. Biochemistry 2010, 49, 10647–10655. [Google Scholar] [CrossRef]
- Purcell, E.B.; Siegal-Gaskins, D.; Rawling, D.C.; Fiebig, A.; Crosson, S. A photosensory two-component system regulates bacterial cell attachment. Proc. Natl. Acad. Sci. USA 2007, 104, 18241–18246. [Google Scholar] [CrossRef]
- Foreman, R.; Fiebig, A.; Crosson, S. The LovK-LovR two-component system is a regulator of the general stress pathway in Caulobacter crescentus. J. Bacteriol. 2012, 194, 3038–3049. [Google Scholar] [CrossRef]
- Rivera-Cancel, G.; Motta-Mena, L.B.; Gardner, K.H. Identification of natural and artificial DNA substrates for light-activated LOV-HTH transcription factor EL222. Biochemistry 2012, 51, 10024–10034. [Google Scholar] [CrossRef]
- Metz, S.; Jäger, A.; Klug, G. Role of a short light, oxygen, voltage (LOV) domain protein in blue light- and singlet oxygen-dependent gene regulation in Rhodobacter sphaeroides. Microbiology 2012, 158, 368–379. [Google Scholar] [CrossRef]
- Niederman, R.A. Membrane development in purple photosynthetic bacteria in response to alterations in light intensity and oxygen tension. Photosynth. Res. 2013, 116, 338–348. [Google Scholar] [CrossRef]
- Pandey, R.; Flockerzi, D.; Hauser, M.J.B.; Straube, R. An extended model for the repression of photosynthesis genes by the AppA/PpsR system in Rhodobacter sphaeroides. FEBS J. 2012, 279, 3449–3461. [Google Scholar] [CrossRef]
- Tschowri, N.; Lindenberg, S.; Hengge, R. Molecular function and potential evolution of the biofilm-modulating blue light-signalling pathway of Escherichia coli. Mol. Microbiol. 2012, 85, 893–906. [Google Scholar] [CrossRef]
- Metz, S.; Haberzettl, K.; Frühwirth, S.; Teich, K.; Hasewinkel, C.; Klug, G. Interaction of two photoreceptors in the regulation of bacterial photosynthesis genes. Nucleic Acids Res. 2012, 40, 5901–5909. [Google Scholar]
- Metz, S.; Jager, A.; Klug, G. In vivo sensitivity of blue-light-dependent signaling mediated by AppA/PpsR or PrrB/PrrA in Rhodobacter sphaeroides. J. Bacteriol. 2009, 191, 4473–4477. [Google Scholar] [CrossRef]
- Moskvin, O.V.; Gilles-Gonzalez, M.A.; Gomelsky, M. The PpaA/AerR regulators of photosynthesis gene expression from anoxygenic phototrophic proteobacteria contain heme-binding SCHIC domains. J. Bacteriol. 2010, 192, 5253–5256. [Google Scholar] [CrossRef]
- Singh, A.H.; Doerks, T.; Letunic, I.; Raes, J.; BORK, P. Discovering functional novelty in metagenomes: Examples from light-mediated processes. J. Bacteriol. 2009, 191, 32–41. [Google Scholar] [CrossRef]
- Pathak, G.; Ehrenreich, A.; Losi, A.; Streit, W.R.; Gaertner, W. Novel blue light-sensitive proteins from a metagenomic approach. Environ. Microbiol. 2009, 11, 2388–2399. [Google Scholar] [CrossRef]
- Pathak, G.; Losi, A.; Gärtner, W. Metagenome-based screening reveals worldwide distribution of LOV-domain proteins. Photochem. Photobiol. 2012, 88, 107–118. [Google Scholar] [CrossRef]
- Inoue, K.; Tsukamoto, T.; Sudo, Y. Molecular and evolutionary aspects of microbial sensory rhodopsins. Biochim. Biophys. Acta 2013. [Google Scholar] [CrossRef]
- Crooks, G.E.; Hon, G.; Chandonia, J.M.; Brenner, S.E. WebLogo: A sequence logo generator. Genome Res. 2004, 14, 1188–1190. [Google Scholar] [CrossRef]
- Schneider, T.D.; Stephens, R.M. Sequence logos: A new way to display consensus sequences. Nucleic Acids Res. 1990, 18, 6097–6100. [Google Scholar] [CrossRef]
- Altschul, S.F.; Madden, T.L.; Schaffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef]
- Hefti, M.H.; Francoijs, K.J.; de Vries, S.C.; Dixon, R.; Vervoort, J. The PAS fold: A redefinition of the PAS domain based upon structural prediction. FEBS J. 2004, 271, 1198–1208. [Google Scholar]
- Ulrich, L.E.; Koonin, E.V.; Zhulin, I.B. One-component systems dominate signal transduction in prokaryotes. Trends Microbiol. 2005, 13, 52–56. [Google Scholar] [CrossRef]
- Sigrist, C.J.; Cerutti, L.; Hulo, N.; Gattiker, A.; Falquet, L.; Pagni, M.; Bairoch, A.; Bucher, P. PROSITE: A documented database using patterns and profiles as motif descriptors. Brief Bioinform. 2002, 3, 265–274. [Google Scholar] [CrossRef]
- Baliga, N.S.; Pan, M.; Goo, Y.A.; Yi, E.C.; Goodlett, D.R.; Dimitrov, K.; Shannon, P.; Aebersold, R.; Ng, W.V.; Hood, L. Coordinate regulation of energy transduction modules in Halobacterium sp. analyzed by a global systems approach. Proc. Natl. Acad. Sci. USA 2002, 99, 14913–14918. [Google Scholar] [CrossRef]
- O’Donovan, C.; Martin, M.J.; Gattiker, A.; Gasteiger, E.; Bairoch, A.; Apweiler, R. High-quality protein knowledge resource: SWISS-PROT and TrEMBL. Brief. Bioinform. 2002, 3, 275–284. [Google Scholar]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef]
- Mulder, N.J.; Apweiler, R.; Attwood, T.K.; Bairoch, A.; Barrel, D.; Bateman, A.; Biswas, M.; Bradley, P.; Bucher, P.; Copley, R.R.; et al. The InterPro Database, 2003 brings increased coverage and new features. Nucleic Acids Res. 2003, 31, 315–318. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Losi, A.; Mandalari, C.; Gärtner, W. From Plant Infectivity to Growth Patterns: The Role of Blue-Light Sensing in the Prokaryotic World. Plants 2014, 3, 70-94. https://doi.org/10.3390/plants3010070
Losi A, Mandalari C, Gärtner W. From Plant Infectivity to Growth Patterns: The Role of Blue-Light Sensing in the Prokaryotic World. Plants. 2014; 3(1):70-94. https://doi.org/10.3390/plants3010070
Chicago/Turabian StyleLosi, Aba, Carmen Mandalari, and Wolfgang Gärtner. 2014. "From Plant Infectivity to Growth Patterns: The Role of Blue-Light Sensing in the Prokaryotic World" Plants 3, no. 1: 70-94. https://doi.org/10.3390/plants3010070