
The Role of Flavonoids in Nodulation Host-Range Specificity: An Update

Abstract
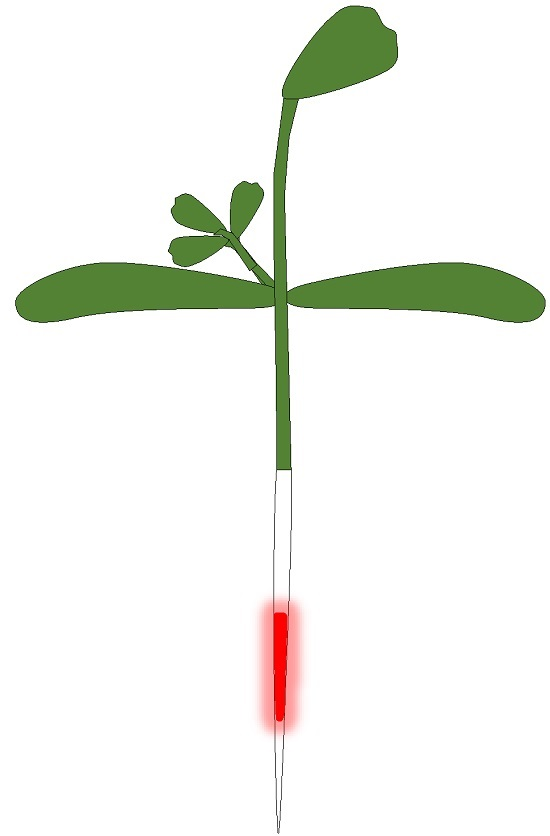
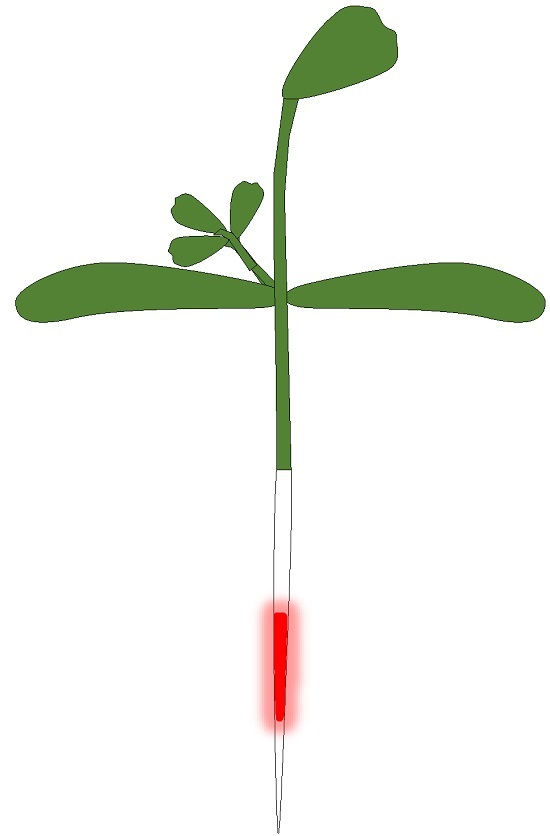
:
1. Introduction
2. Flavonoids as Determinants of Host Range
2.1. Flavonoids as Infection Signals
- “strong” inducers of nod genes in homologous rhizobia
- secreted by roots (i.e., found in root exudate)
- increased biosynthesis in response to rhizobia or Nod factors
- required for rhizobial infection (i.e., genetic evidence)
2.2. Flavonoid Phytoalexins as Determinants of Host Range
2.3. Manipulation of Host Range
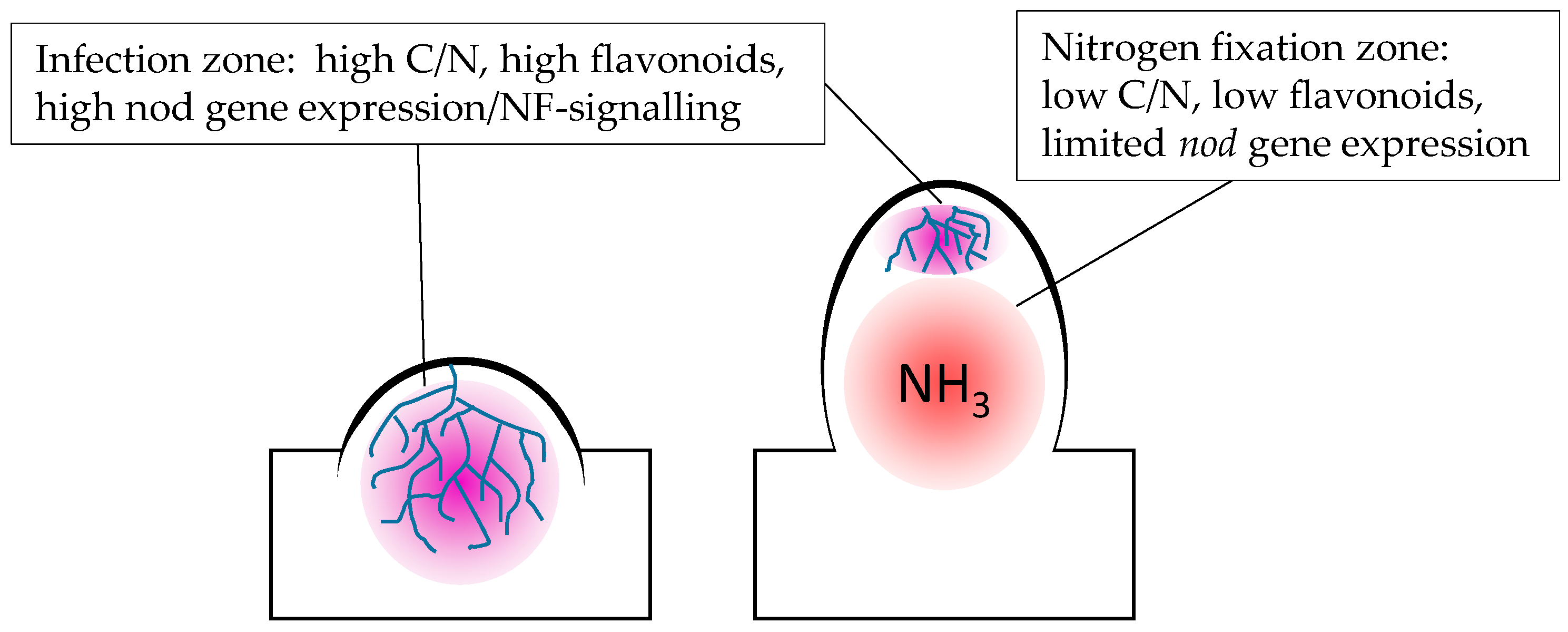
3. C/N Status May Play a Central Role in the Regulation of Flavonoid Levels in Nodules
4. Conclusions and Future Prospects
Acknowledgments
Conflicts of Interest
References
- Spaink, H.P.; Okker, R.J.H.; Wijffelman, C.A.; Tak, T.; Goosenderoo, L.; Pees, E.; Vanbrussel, A.A.N.; Lugtenberg, B.J.J. Symbiotic properties of rhizobia containing a flavonoid-independent hybrid nodD product. J. Bacteriol. 1989, 171, 4045–4053. [Google Scholar] [PubMed]
- Mulligan, J.T.; Long, S.R. Induction of Rhizobium meliloti nodC expression by plant exudate requires nodD. Proc. Natl. Acad. Sci. USA 1985, 82, 6609–6613. [Google Scholar] [CrossRef] [PubMed]
- Oldroyd, G.E.D.; Murray, J.D.; Poole, P.S.; Downie, J.A. The rules of engagement in the legume-rhizobial Symbiosis. Annu. Rev. Genet. 2011, 45, 119–144. [Google Scholar] [CrossRef] [PubMed]
- Lerouge, P.; Roche, P.; Faucher, C.; Maillet, F.; Truchet, G.; Prome, J.C.; Denarie, J. Symbiotic host-specificity of Rhizobium-meliloti is determined by a sulfated and acylated glucosamine oligosaccharide signal. Nature 1990, 344, 781–784. [Google Scholar] [CrossRef] [PubMed]
- Spaink, H.P.; Sheeley, D.M.; Vanbrussel, A.A.N.; Glushka, J.; York, W.S.; Tak, T.; Geiger, O.; Kennedy, E.P.; Reinhold, V.N.; Lugtenberg, B.J.J. A novel highly unsaturated fatty-acid moiety of lipo-oligosaccharide signals determines host specificity of rhizobium. Nature 1991, 354, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Spaink, H.P.; Wijffelman, C.A.; Pees, E.; Okker, R.J.H.; Lugtenberg, B.J.J. Rhizobium nodulation gene nodD as a determinant of host specificity. Nature 1987, 328, 337–340. [Google Scholar] [CrossRef]
- Radutoiu, S.; Madsen, L.H.; Madsen, E.B.; Felle, H.H.; Umehara, Y.; Gronlund, M.; Sato, S.; Nakamura, Y.; Tabata, S.; Sandal, N.; et al. Plant recognition of symbiotic bacteria requires two LysM receptor-like kinases. Nature 2003, 425, 585–592. [Google Scholar] [CrossRef] [PubMed]
- Madsen, E.B.; Madsen, L.H.; Radutoiu, S.; Olbryt, M.; Rakwalska, M.; Szczyglowski, K.; Sato, S.; Kaneko, T.; Tabata, S.; Sandal, N.; et al. A receptor kinase gene of the LysM type is involved in legume perception of rhizobial signals. Nature 2003, 425, 637–640. [Google Scholar] [CrossRef] [PubMed]
- Wasson, A.P.; Pellerone, F.I.; Mathesius, U. Silencing the flavonoid pathway in Medicago truncatula inhibits root nodule formation and prevents auxin transport regulation by rhizobia. Plant Cell 2006, 18, 1617–1629. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, S.; Stacey, G.; Yu, O. Endogenous isoflavones are essential for the establishment of symbiosis between soybean and Bradyrhizobium japonicum. Plant J. 2006, 48, 261–273. [Google Scholar] [CrossRef] [PubMed]
- Auguy, F.; Abdel-Lateif, K.; Doumas, P.; Badin, P.; Guerin, V.; Bogusz, D.; Hocher, V. Activation of the isoflavonoid pathway in actinorhizal symbioses. Funct. Plant Biol. 2011, 38, 690–696. [Google Scholar] [CrossRef]
- Popovici, J.; Comte, G.; Bagnarol, E.; Alloisio, N.; Fournier, P.; Bellvert, F.; Bertrand, C.; Fernandez, M.P. Differential effects of rare specific flavonoids on compatible and incompatible strains in the Myrica gale-Frankia actinorhizal symbiosis. Appl. Environ. Microbiol. 2010, 76, 2451–2460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popovici, J.; Walker, V.; Bertrand, C.; Bellvert, F.; Fernandez, M.P.; Comte, G. Strain specificity in the Myricaceae-Frankia symbiosis is correlated to plant root phenolics. Funct. Plant Biol. 2011, 38, 682–689. [Google Scholar] [CrossRef]
- Abdel-Lateif, K.; Vaissayre, V.; Gherbi, H.; Verries, C.; Meudec, E.; Perrine-Walker, F.; Cheynier, V.; Svistoonoff, S.; Franche, C.; Bogusz, D.; et al. Silencing of the chalcone synthase gene in Casuarina glauca highlights the important role of flavonoids during nodulation. New Phytol. 2013, 199, 1012–1021. [Google Scholar] [CrossRef] [PubMed]
- Hassan, S.; Mathesius, U. The role of flavonoids in root-rhizosphere signalling: Opportunities and challenges for improving plant-microbe interactions. J. Exp. Bot. 2012, 63, 3429–3444. [Google Scholar] [CrossRef] [PubMed]
- Weston, L.A.; Mathesius, U. Flavonoids: Their structure, biosynthesis and role in the rhizosphere, including allelopathy. J. Chem. Ecol. 2013, 39, 283–297. [Google Scholar] [CrossRef] [PubMed]
- Caetanoanolles, G.; Cristestes, D.K.; Bauer, W.D. Chemotaxis of Rhizobium-meliloti to the plant flavone luteolin requires functional nodulation genes. J. Bacteriol. 1988, 170, 3164–3169. [Google Scholar]
- Aguilar, J.M.M.; Ashby, A.M.; Richards, A.J.M.; Loake, G.J.; Watson, M.D.; Shaw, C.H. Chemotaxis of Rhizobium-leguminosarum biovar phaseoli towards flavonoid inducers of the symbiotic nodulation genes. J. Gen. Microbiol. 1988, 134, 2741–2746. [Google Scholar]
- Theunis, M.; Kobayashi, H.; Broughton, W.J.; Prinsen, E. Flavonoids, nodD1, nodD2, and nod-box NB15 modulate expression of the y4wEFG locus that is required for indole-3-acetic acid synthesis in Rhizobium sp. strain NGR234. Mol. Plant-Microbe Interact. 2004, 17, 1153–1161. [Google Scholar] [CrossRef] [PubMed]
- Maj, D.; Wielbo, J.; Marek-Kozaczuk, M.; Skorupska, A. Response to flavonoids as a factor influencing competitiveness and symbiotic activity of Rhizobium leguminosarum. Microbiol. Res. 2010, 165, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Cesco, S.; Mimmo, T.; Tonon, G.; Tomasi, N.; Pinton, R.; Terzano, R.; Neumann, G.; Weisskopf, L.; Renella, G.; Landi, L.; et al. Plant-borne flavonoids released into the rhizosphere: Impact on soil bio-activities related to plant nutrition. A review. Biol. Fertil. Soils 2012, 48, 123–149. [Google Scholar] [CrossRef]
- Tomasi, N.; Weisskopf, L.; Renella, G.; Landi, L.; Pinton, R.; Varanini, Z.; Nannipieri, P.; Torrent, J.; Martinoia, E.; Cesco, S. Flavonoids of white lupin roots participate in phosphorus mobilization from soil. Soil Biol. Biochem. 2008, 40, 1971–1974. [Google Scholar] [CrossRef]
- Faucher, C.; Maillet, F.; Vasse, J.; Rosenberg, C.; Vanbrussel, A.A.N.; Truchet, G.; Denarie, J. Rhizobium-meliloti host range nodh-gene determines production of an alfalfa-specific extracellular signal. J. Bacteriol. 1988, 170, 5489–5499. [Google Scholar] [PubMed]
- Debelle, F.; Moulin, L.; Mangin, B.; Denarie, J.; Boivin, C. NoD genes and nod signals and the evolution of the Rhizobium legume symbiosis. Acta Biochim. Pol. 2001, 48, 359–365. [Google Scholar] [PubMed]
- Wais, R.J.; Keating, D.H.; Long, S.R. Structure-function analysis of nod factor-induced root hair calcium spiking in Rhizobium-Legume Symbiosis. Plant Physiol. 2002, 129, 211–224. [Google Scholar] [CrossRef] [PubMed]
- Zavala, K.; Opazo, J.C. Lineage-specific expansion of the chalcone synthase gene family in rosids. PLoS ONE 2015, 10, e0133400. [Google Scholar] [CrossRef] [PubMed]
- Dewick, P.M. Isoflavonoids. In The Flavonoids: Advances in Research Since 1980; Harborne, J.B., Ed.; Chapman and Hall: New York, NY, USA, 1988; pp. 125–210. [Google Scholar]
- Wang, X. Structure, function, and engineering of enzymes in isoflavonoid biosynthesis. Funct. Integr. Genom. 2011, 11, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, S.; Stacey, G.; Yu, O. Distinct, crucial roles of flavonoids during legume nodulation. Trends Plant Sci. 2007, 12, 282–285. [Google Scholar] [CrossRef] [PubMed]
- Chovanec, P.; Novak, K. Visualization of nodulation gene activity on the early stages of Rhizobium leguminosarum bv. Viciae symbiosis. Folia Microbiol. 2005, 50, 323–331. [Google Scholar] [CrossRef]
- Zuanazzi, J.A.S.; Clergeot, P.H.; Quirion, J.C.; Husson, H.P.; Kondorosi, A.; Ratet, P. Production of Sinorhizobium meliloti nod gene activator and repressor flavonoids from Medicago sativa roots. Mol. Plant-Microbe Interact. 1998, 11, 784–794. [Google Scholar] [CrossRef]
- Phillips, D.A.; Dakora, F.D.; Sande, E.; Joseph, C.M.; Zon, J. Synthesis, release, and transmission of alfalfa signals to rhizobial symbionts. Plant Soil 1994, 161, 69–80. [Google Scholar] [CrossRef]
- Kape, R.; Parniske, M.; Brandt, S.; Werner, D. Isoliquiritigenin, a strong nod gene-inducing and glyceollin resistance-inducing flavonoid from soybean root exudate. Appl. Environ. Microbiol. 1992, 58, 1705–1710. [Google Scholar] [PubMed]
- Maxwell, C.A.; Hartwig, U.A.; Joseph, C.M.; Phillips, D.A. A chalcone and two related flavonoids released from alfalfa roots induce nod genes of Rhizobium-meliloti. Plant Physiol. 1989, 91, 842–847. [Google Scholar] [CrossRef] [PubMed]
- Peters, N.K.; Frost, J.W.; Long, S.R. A plant flavone, luteolin, induces expression of Rhizobium-meliloti nodulation genes. Science 1986, 233, 977–980. [Google Scholar] [CrossRef] [PubMed]
- Redmond, J.W.; Batley, M.; Djordjevic, M.A.; Innes, R.W.; Kuempel, P.L.; Rolfe, B.G. Flavones induce expression of nodulation genes in Rhizobium. Nature 1986, 323, 632–635. [Google Scholar] [CrossRef]
- Marie, C.; Barny, M.A.; Downie, J.A. Rhizobium leguminosarum has two glucosamine synthases, glms and nodm, required for nodulation and development of nitrogen-fixing nodules. Mol. Microbiol. 1992, 6, 843–851. [Google Scholar] [CrossRef] [PubMed]
- Den Herder, J.; Vanhee, C.; De Rycke, R.; Corich, V.; Holsters, M.; Goormachtig, S. Nod factor perception during infection thread growth fine-tunes nodulation. Mol. Plant-Microbe Interact. 2007, 20, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Bisby, F. Phytochemical Dictionary of the Leguminosae; Chapman and Hall/CRC: London, UK, 1994; Volume 2. [Google Scholar]
- Zaat, S.A.J.; Wijffelman, C.A.; Spaink, H.P.; Vanbrussel, A.A.N.; Okker, R.J.H.; Lugtenberg, B.J.J. Induction of the nodA promoter of Rhizobium leguminosarum Sym plasmid pRL1JI by plant flavanones and flavones. J. Bacteriol. 1987, 169, 198–204. [Google Scholar] [PubMed]
- Suominen, L.; Luukkainen, R.; Roos, C.; Lindstrom, K. Activation of the nodA promoter by the nodD genes of Rhizobium galegae induced by synthetic flavonoids or Galega orientalis root exudate. FEMS Microbiol. Lett. 2003, 219, 225–232. [Google Scholar] [CrossRef]
- Kamboj, D.V.; Bhatia, R.; Pathak, D.V.; Sharma, P.K. Role of nodD gene product and flavonoid interactions in induction of nodulation genes in Mesorhizobium ciceri. Physiol. Mol. Biol. Plants 2010, 16, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Kapulnik, Y.; Joseph, C.M.; Phillips, D.A. Flavone limitations to root nodulation and symbiotic Nitrogen-fixation in alfalfa. Plant Physiol. 1987, 84, 1193–1196. [Google Scholar] [CrossRef] [PubMed]
- Spini, G.; Decorosi, F.; Cerboneschi, M.; Tegli, S.; Mengoni, A.; Viti, C.; Giovannetti, L. Effect of the plant flavonoid luteolin on Ensifer meliloti 3001 phenotypic responses. Plant Soil 2016, 399, 159–178. [Google Scholar] [CrossRef]
- Hartwig, U.A.; Maxwell, C.A.; Joseph, C.M.; Phillips, D.A. Chrysoeriol and luteolin released from alfalfa seeds induce nod genes in Rhizobium meliloti. Plant Physiol. 1990, 92, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Recourt, K.; Schripsema, J.; Kijne, J.W.; Vanbrussel, A.A.N.; Lugtenberg, B.J.J. Inoculation of Vicia-sativa subsp nigra roots with Rhizobium-leguminosarum biovar viciae results in release of nod gene activating flavanones and chalcones. Plant Mol. Biol. 1991, 16, 841–852. [Google Scholar] [CrossRef] [PubMed]
- Maxwell, C.A.; Edwards, R.; Dixon, R.A. Identification, purification, and characterization of s-adenosyl-l-methionine-isoliquiritigenin 2′-O-methyltransferase from alfalfa (Medicago sativa L.). Arch. Biochem. Biophys. 1992, 293, 158–166. [Google Scholar] [CrossRef]
- Maxwell, C.A.; Harrison, M.J.; Dixon, R.A. Molecular characterization and expression of alfalfa isoliquiritigenin 2′-O-methyltransferase, an enzyme specifically involved in the biosynthesis of an inducer of Rhizobium meliloti nodulation genes. Plant J. 1993, 4, 971–981. [Google Scholar] [CrossRef] [PubMed]
- Roux, B.; Rodde, N.; Jardinaud, M.-F.; Timmers, T.; Sauviac, L.; Cottret, L.; Carrere, S.; Sallet, E.; Courcelle, E.; Moreau, S.; et al. An integrated analysis of plant and bacterial gene expression in symbiotic root nodules using laser-capture microdissection coupled to rna sequencing. Plant J. 2014, 77, 817–837. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.-S.; Liu, C.-W.; Roy, S.; Cousins, D.; Stacey, N.; Murray, J.D. Identification of a core set of rhizobial infection genes using data from single cell-types. Front. Plant Sci. 2015, 6, 575. [Google Scholar] [CrossRef] [PubMed]
- Breakspear, A.; Liu, C.; Roy, S.; Stacey, N.; Rogers, C.; Trick, M.; Morieri, G.; Mysore, K.S.; Wen, J.; Oldroyd, G.E.D.; et al. The root hair “infectome” of Medicago truncatula uncovers changes in cell cycle genes and reveals a requirement for auxin signaling in rhizobial infection. Plant Cell 2014, 26, 4680–4701. [Google Scholar] [CrossRef] [PubMed]
- Libault, M.; Farmer, A.; Brechenmacher, L.; Drnevich, J.; Langley, R.J.; Bilgin, D.D.; Radwan, O.; Neece, D.J.; Clough, S.J.; May, G.D.; et al. Complete transcriptome of the soybean root hair cell, a single-cell model, and its alteration in response to Bradyrhizobium japonicum infection. Plant Physiol. 2010, 152, 541–552. [Google Scholar] [CrossRef] [PubMed]
- Recourt, K.; van Tunen, A.J.; Mur, L.A.; van Brussel, A.A.; Lugtenberg, B.J.; Kijne, J.W. Activation of flavonoid biosynthesis in roots of Vicia sativa subsp. Nigra plants by inoculation with Rhizobium leguminosarum biovar viciae. Plant Mol. Biol. 1992, 19, 411–420. [Google Scholar] [CrossRef] [PubMed]
- Ayabe, S.I.; Kobayashi, M.; Hikichi, M.; Matsumoto, K.; Furuya, T. Studies on plant-tissue cultures. 33. Flavonoids from the cultured-cells of Glycyrrhiza echinata. Phytochemistry 1980, 19, 2179–2183. [Google Scholar] [CrossRef]
- Carlson, R.E.; Dolphin, D.H. Pisum-sativum stress metabolites-2 cinnamylphenols and a 2′-methoxychalcone. Phytochemistry 1982, 21, 1733–1736. [Google Scholar] [CrossRef]
- Kosslak, R.M.; Bookland, R.; Barkei, J.; Paaren, H.E.; Appelbaum, E.R. Induction of Bradyrhizobium japonicum common nod genes by isoflavones isolated from Glycine max. Proc. Natl. Acad. Sci. USA 1987, 84, 7428–7432. [Google Scholar] [CrossRef] [PubMed]
- Pueppke, S.G.; Bolanos-Vasquez, M.C.; Werner, D.; Bec-Ferte, M.P.; Prome, J.C.; Krishnan, H.B. Release of flavonoids by the soybean cultivars mccall and peking and their perception as signals by the nitrogen-fixing symbiont Sinorhizobium fredii. Plant Physiol. 1998, 117, 599–608. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, P.E.; Broughton, W.J.; Werner, D. Nod factors of Bradyrhizobium-japonicum and Rhizobium sp. NGR234 induce flavonoid accumulation in soybean root exudate. Mol. Plant-Microbe Interact. 1994, 7, 384–390. [Google Scholar] [CrossRef]
- Brechenmacher, L.; Lei, Z.; Libault, M.; Findley, S.; Sugawara, M.; Sadowsky, M.J.; Sumner, L.W.; Stacey, G. Soybean metabolites regulated in root hairs in response to the symbiotic bacterium Bradyrhizobium japonicum. Plant Physiol. 2010, 153, 1808–1822. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, T. Flavonoid-responsive NodY-lacZ expression in three phylogenetically different Bradyrhizobium groups. Can. J. Microbiol. 2008, 54, 401–410. [Google Scholar] [CrossRef] [PubMed]
- Lawson, C.G.R.; Rolfe, B.G.; Djordjevic, M.A. Rhizobium inoculation induces condition-dependent changes in the flavonoid composition of root exudates from Trifolium subterraneum. Aust. J. Plant Physiol. 1996, 23, 93–101. [Google Scholar] [CrossRef]
- Vanbrussel, A.A.N.; Recourt, K.; Pees, E.; Spaink, H.P.; Tak, T.; Wijffelman, C.A.; Kijne, J.W.; Lugtenberg, B.J.J. A biovar-specific signal of Rhizobium-leguminosarum bv viciae induces increased nodulation gene-inducing activity in root exudate of Vicia sativa subsp nigra. J. Bacteriol. 1990, 172, 5394–5401. [Google Scholar]
- Dakora, F.D.; Joseph, C.M.; Phillips, D.A. Common bean root exudates contain elevated levels of daidzein and coumestrol in response to Rhizobium inoculation. Mol. Plant-Microbe Interact. 1993, 6, 665–668. [Google Scholar] [CrossRef]
- Janczarek, M.; Rachwal, K.; Marzec, A.; Grzadziel, J.; Palusinska-Szysz, M. Signal molecules and cell-surface components involved in early stages of the legume-Rhizobium interactions. Appl. Soil Ecol. 2015, 85, 94–113. [Google Scholar] [CrossRef]
- Cooper, J.E. Early interactions between legumes and rhizobia: Disclosing complexity in a molecular dialogue. J. Appl. Microbiol. 2007, 103, 1355–1365. [Google Scholar] [CrossRef] [PubMed]
- BolanosVasquez, M.C.; Warner, D. Effects of Rhizobium tropici, R. etli, and R. leguminosarum bv phaseoli on nod gene-inducing flavonoids in root exudates of Phaseolus vulgaris. Mol. Plant-Microbe Interact. 1997, 10, 339–346. [Google Scholar] [CrossRef]
- Dakora, F.D.; Joseph, C.M.; Phillips, D.A. Alfalfa (Medicago sativa l) root exudates contain isoflavonoids in the presence of Rhizobium meliloti. Plant Physiol. 1993, 101, 819–824. [Google Scholar] [PubMed]
- Novak, K.; Lisa, L.; Skrdleta, V. Rhizobial nod gene-inducing activity in pea nodulation mutants: Dissociation of nodulation and flavonoid response. Physiol. Plant 2004, 120, 546–555. [Google Scholar] [CrossRef] [PubMed]
- Cooper, J.E.; Rao, J.R. Localized changes in flavonoid biosynthesis in roots of Lotus pedunculatus after infection by Rhizobium loti. Plant Physiol. 1992, 100, 444–450. [Google Scholar] [CrossRef] [PubMed]
- Steele, H.L.; Werner, D.; Cooper, J.E. Flavonoids in seed and root exudates of Lotus pedunculatus and their biotransformation by Mesorhizobium loti. Physiol. Plant 1999, 107, 251–258. [Google Scholar] [CrossRef]
- Parniske, M.; Zimmermann, C.; Cregan, P.B.; Werner, D. Hypersensitive reaction of nodule cells in the Glycine sp./Bradyrhizobium japonicum symbiosis occurs at the genotype-specific level. Bot. Acta 1990, 103, 143–148. [Google Scholar] [CrossRef] [Green Version]
- Novak, K.; Kropacova, M.; Havlicek, V.; Skrdleta, V. Isoflavonoid phytoalexin pisatin is not recognized by the flavonoid receptor nodD of Rhizobium leguminosarum bv viciae. Folia Microbiol. 1995, 40, 535–540. [Google Scholar] [CrossRef]
- Haraguchi, H.; Tanimoto, K.; Tamura, Y.; Mizutani, K.; Kinoshita, T. Mode of antibacterial action of retrochalcones from Glycyrrhiza inflata. Phytochemistry 1998, 48, 125–129. [Google Scholar] [CrossRef]
- Akiyama, K.; Kawazu, K.; Kobayashi, A. Partially n-deacetylated chitin elicitor induces antimicrobial flavonoids in pea epicotyls. Z. Fur Naturforschung C 1994, 49, 811–818. [Google Scholar]
- Ulanowska, K.; Tkaczyk, A.; Konopa, G.; Wegrzyn, G. Differential antibacterial activity of genistein arising from global inhibition of DNA, rna and protein synthesis in some bacterial strains. Arch. Microbiol. 2006, 184, 271–278. [Google Scholar] [CrossRef] [PubMed]
- Weidenborner, M.; Hindorf, H.; Jha, H.C.; Tsotsonos, P.; Egge, H. Antifungal activity of isoflavonoids in different reduced stages on Rhizoctonia-solani and Sclerotium-rolfsii. Phytochemistry 1990, 29, 801–803. [Google Scholar] [CrossRef]
- Paiva, N.L.; Oommen, A.; Harrison, M.J.; Dixon, R.A. Regulation of isoflavonoid metabolism in alfalfa. Plant Cell Tissue Organ Cult. 1994, 38, 213–220. [Google Scholar] [CrossRef]
- Guenoune, D.; Galili, S.; Phillips, D.A.; Volpin, H.; Chet, I.; Okon, Y.; Kapulnik, Y. The defense response elicited by the pathogen Rhizoctonia solani is suppressed by colonization of the am-fungus Glomus intraradices. Plant Sci. 2001, 160, 925–932. [Google Scholar] [CrossRef]
- Benedito, V.A.; Torres-Jerez, I.; Murray, J.D.; Andriankaja, A.; Allen, S.; Kakar, K.; Wandrey, M.; Verdier, J.; Zuber, H.; Ott, T.; et al. A gene expression atlas of the model legume Medicago truncatula. Plant J. 2008, 55, 504–513. [Google Scholar] [CrossRef] [PubMed]
- Uppalapati, S.R.; Marek, S.M.; Lee, H.K.; Nakashima, J.; Tang, Y.; Sledge, M.K.; Dixon, R.A.; Mysore, K.S. Global gene expression profiling during Medicago truncatula-Phymatotrichopsis omnivora interaction reveals a role for jasmonic acid, ethylene, and the flavonoid pathway in disease development. Mol. Plant Microbe Interact. 2009, 22, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Mah, K.M.; Uppalapati, S.R.; Tang, Y.H.; Allen, S.; Shuai, B. Gene expression profiling of Macrophomina phaseolina infected Medicago truncatula roots reveals a role for auxin in plant tolerance against the charcoal rot pathogen. Physiol. Mol. Plant Pathol. 2012, 79, 21–30. [Google Scholar] [CrossRef]
- Dakora, F.D.; Phillips, D.A. Diverse functions of isoflavonoids in legumes transcend anti-microbial definitions of phytoalexins. Physiol. Mol. Plant Pathol. 1996, 49, 1–20. [Google Scholar] [CrossRef]
- Farag, M.A.; Huhman, D.V.; Lei, Z.; Sumner, L.W. Metabolic profiling and systematic identification of flavonoids and isoflavonoids in roots and cell suspension cultures of Medicago truncatula using HPLC-UV-ESI-MS and GC-MS. Phytochemistry 2007, 68, 342–354. [Google Scholar] [CrossRef] [PubMed]
- Hargreaves, J.A.; Mansfield, J.W.; Coxon, D.T. Identification of medicarpin as a phytoalexin in broad bean plant (Vicia-faba-L). Nature 1976, 262, 318–319. [Google Scholar] [CrossRef]
- Guo, L.; Dixon, R.A.; Paiva, N.L. Conversion of vestitone to medicarpin in alfalfa (Medicago sativa L.) is catalyzed by two independent enzymes. Identification, purification, and characterization of vestitone reductase and 7,2′-dihydroxy-4′-methoxyisoflavanol dehydratase. J. Biol. Chem. 1994, 269, 22372–22378. [Google Scholar] [PubMed]
- Guo, L.; Paiva, N.L. Molecular cloning and expression of alfalfa (Medicago sativa L.) vestitone reductase, the penultimate enzyme in medicarpin biosynthesis. Arch. Biochem. Biophys. 1995, 320, 353–360. [Google Scholar] [CrossRef]
- Pankhurst, C.E.; Biggs, D.R. Sensitivity of Rhizobium to selected isoflavonoids. Can. J. Microbiol. 1980, 26, 542–545. [Google Scholar] [CrossRef] [PubMed]
- Parniske, M.; Ahlborn, B.; Werner, D. Isoflavonoid-inducible resistance to the phytoalexin glyceollin in soybean rhizobia. J. Bacteriol. 1991, 173, 3432–3439. [Google Scholar] [PubMed]
- Gonzalez-Pasayo, R.; Martinez-Romero, E. Multiresistance genes of Rhizobium etli CFN42. Mol. Plant-Microbe Interact. 2000, 13, 572–577. [Google Scholar] [CrossRef] [PubMed]
- Lindemann, A.; Koch, M.; Pessi, G.; Muller, A.J.; Balsiger, S.; Hennecke, H.; Fischer, H.M. Host-specific symbiotic requirement of BdeAB, a RegR-controlled RND-type efflux system in Bradyrhizobium japonicum. FEMS Microbiol. Lett. 2010, 312, 184–191. [Google Scholar] [CrossRef] [PubMed]
- Soedarjo, M.; Borthakur, D. Mimosine, a toxin produced by the tree-legume leucaena provides a nodulation competition advantage to mimosine-degrading rhizobium strains. Soil Biol. Biochem. 1998, 30, 1605–1613. [Google Scholar] [CrossRef]
- Broughton, W.J.; Samrey, U.; Stanley, J. Ecological genetics of Rhizobium meliloti-Symbiotic plasmid transfer in the Medicago sativa rhizosphere. FEMS Microbiol. Lett. 1987, 40, 251–255. [Google Scholar] [CrossRef]
- Espuny, M.R.; Ollero, F.J.; Bellogin, R.A.; Ruizsainz, J.E.; Perezsilva, J. Transfer of the Rhizobium-leguminosarum biovar trifolii symbiotic plasmid prtr5a to a strain of rhizobium sp. that nodulates on hedysarum-coronarium. J. Appl. Bacteriol. 1987, 63, 13–20. [Google Scholar] [CrossRef]
- Radutoiu, S.; Madsen, L.H.; Madsen, E.B.; Jurkiewicz, A.; Fukai, E.; Quistgaard, E.M.; Albrektsen, A.S.; James, E.K.; Thirup, S.; Stougaard, J. LysM domains mediate Lipochitin-oligosaccharide recognition and nfr genes extend the symbiotic host range. EMBO J. 2007, 26, 3923–3935. [Google Scholar] [CrossRef] [PubMed]
- Bhagwat, A.A.; Thomas, J. Legume-rhizobium interactions-cowpea root exudate elicits faster nodulation response by rhizobium species. Appl. Environ. Microbiol. 1982, 43, 800–805. [Google Scholar] [PubMed]
- Coronado, C.; Zuanazzi, J.A.S.; Sallaud, C.; Quirion, J.C.; Esnault, R.; Husson, H.P.; Kondorosi, A.; Ratet, P. Alfalfa root flavonoid production is nitrogen regulated. Plant Physiol. 1995, 108, 533–542. [Google Scholar] [PubMed]
- Wan, H.; Zhang, J.; Song, T.; Tian, J.; Yao, Y. Promotion of flavonoid biosynthesis in leaves and calli of ornamental crabapple (Malus sp.) by high carbon to nitrogen ratios. Front. Plant Sci. 2015, 6, 673. [Google Scholar] [CrossRef] [PubMed]
- Fritz, C.; Palacios-Rojas, N.; Feil, R.; Stitt, M. Regulation of secondary metabolism by the carbon-nitrogen status in tobacco: Nitrate inhibits large sectors of phenylpropanoid metabolism. Plant J. 2006, 46, 533–548. [Google Scholar] [CrossRef] [PubMed]
- Solfanelli, C.; Poggi, A.; Loreti, E.; Alpi, A.; Perata, P. Sucrose-specific induction of the anthocyanin biosynthetic pathway in arabidopsis. Plant Physiol. 2006, 140, 637–646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lea, U.S.; Slimestad, R.; Smedvig, P.; Lillo, C. Nitrogen deficiency enhances expression of specific myb and bhlh transcription factors and accumulation of end products in the flavonoid pathway. Planta 2007, 225, 1245–1253. [Google Scholar] [CrossRef] [PubMed]
- Bonguebartelsman, M.; Phillips, D.A. Nitrogen stress regulates gene-expression of enzymes in the flavonoid biosynthetic-pathway of tomato. Plant Physiol. Biochem. 1995, 33, 539–546. [Google Scholar]
- Dusha, I.; Bakos, A.; Kondorosi, A.; Debruijn, F.J.; Schell, J. The Rhizobium-meliloti early nodulation genes (nodabc) are nitrogen-regulated—Isolation of a mutant strain with efficient nodulation capacity on alfalfa in the presence of ammonium. Mol. Gen. Genet. 1989, 219, 89–96. [Google Scholar] [CrossRef]
- Wang, S.P.; Stacey, G. Ammonia regulation of nod genes in Bradyrhizobium-japonicum. Mol. Gen. Genet. 1990, 223, 329–331. [Google Scholar] [CrossRef] [PubMed]
- Dusha, I. Nitrogen control of bacterial signal production in Rhizobium meliloti—Alfalfa symbiosis. Indian J. Exp. Biol. 2002, 40, 981–988. [Google Scholar] [PubMed]
- Hunter, W.J. Soybean root and nodule nitrate reductase. Physiol. Plant. 1983, 59, 471–475. [Google Scholar] [CrossRef]
- Peters, N.K.; Long, S.R. Alfalfa root exudates and compounds which promote or inhibit induction of Rhizobium-meliloti nodulation genes. Plant Physiol. 1988, 88, 396–400. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Host Species | (iso)Flavonoids | Tissues | Reference |
|---|---|---|---|
| Soybean | isoliquiritigenin 1 | root/ root hair | [59] |
| liquiritigenin 2 | |||
| apigenin | |||
| prunetin | |||
| afrormosin | |||
| amino-flavonoid | |||
| dihydrokaempferol | |||
| genkwanin | |||
| naringenin 3 | |||
| biochanin-A 3 | |||
| daidzein | root exudates | [58] | |
| genistein | |||
| coumestrol | |||
| Phaseolus vulgaris | genistein | root exudates | [66] |
| daidzein | [63] | ||
| coumestrol | |||
| isoliquiritigenin | |||
| naringenin | |||
| liquiritigenin | |||
| Medicago sativa | methoxychalcone | root exudates | [67] |
| formononetin 3 | |||
| medicarpin 2 | |||
| Vicia sativa | methoxychalcone | root exudates | [46] |
| isoliquiritigenin | |||
| liquiritigenin | |||
| hesperitin | |||
| naringenin | |||
| 7,3′-dihydroxy-4′-methoxyflavanone | |||
| 7,4′-dihydroxy-3′-methoxyflavanone | |||
| 5,7,4′-trihydroxy-3′-methoxyflavanone | |||
| Trifolium subterraneum | 4′,7-dihydroxyflavone | root exudates | [61] |
| Pisum sativum | pisatin | root exudate | [68] |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, C.-W.; Murray, J.D. The Role of Flavonoids in Nodulation Host-Range Specificity: An Update. Plants 2016, 5, 33. https://doi.org/10.3390/plants5030033
Liu C-W, Murray JD. The Role of Flavonoids in Nodulation Host-Range Specificity: An Update. Plants. 2016; 5(3):33. https://doi.org/10.3390/plants5030033
Chicago/Turabian StyleLiu, Cheng-Wu, and Jeremy D. Murray. 2016. "The Role of Flavonoids in Nodulation Host-Range Specificity: An Update" Plants 5, no. 3: 33. https://doi.org/10.3390/plants5030033