Phenotypic and Physiological Evaluation of Two and Six Rows Barley under Different Environmental Conditions

Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Experimental Conditions
2.2. Studied Traits
2.3. Statistical Analysis
3. Results
3.1. Analysis of Variance
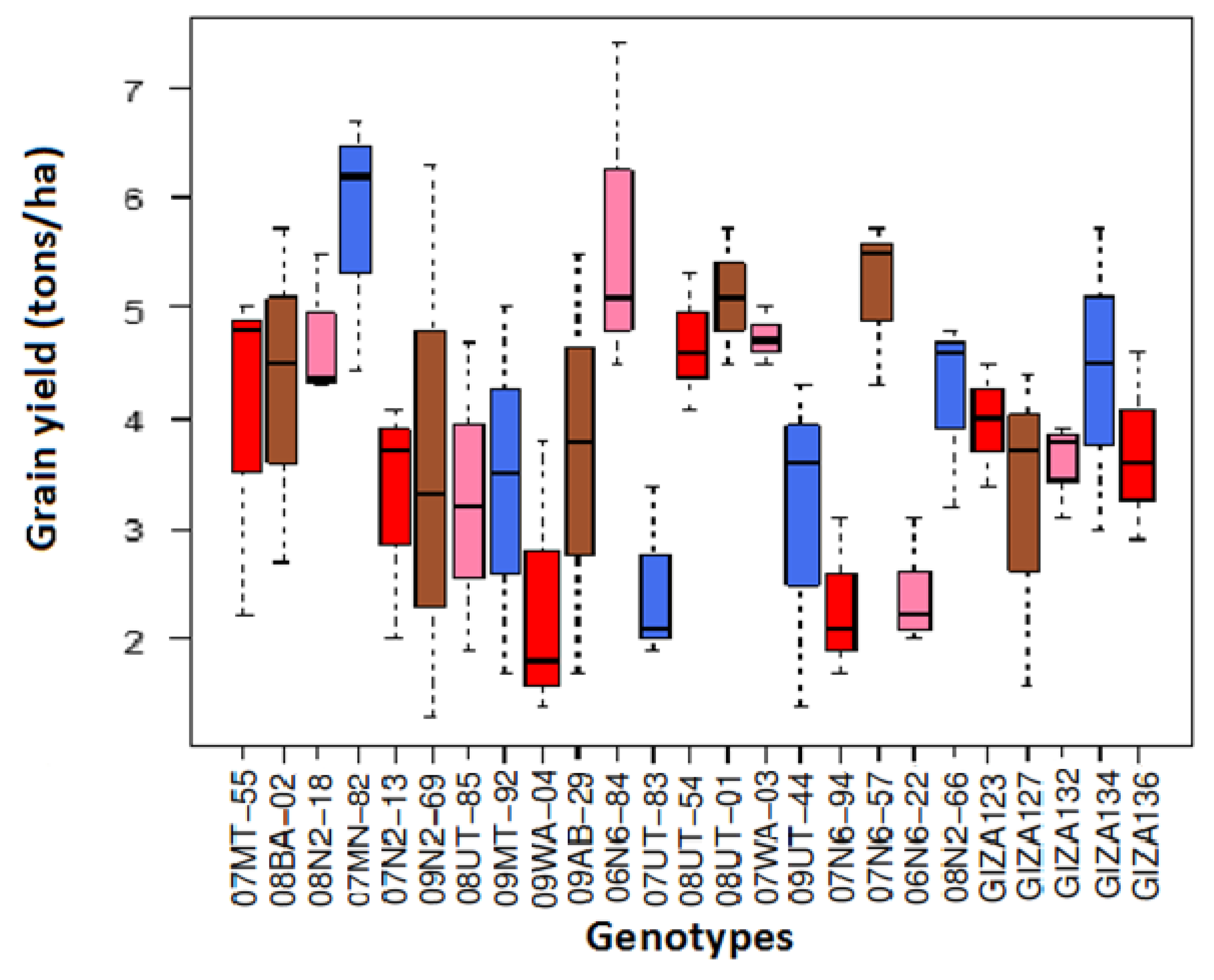
3.2. Traits Mean Across Genotypes and Environments
3.3. Relationships among the Studied Traits
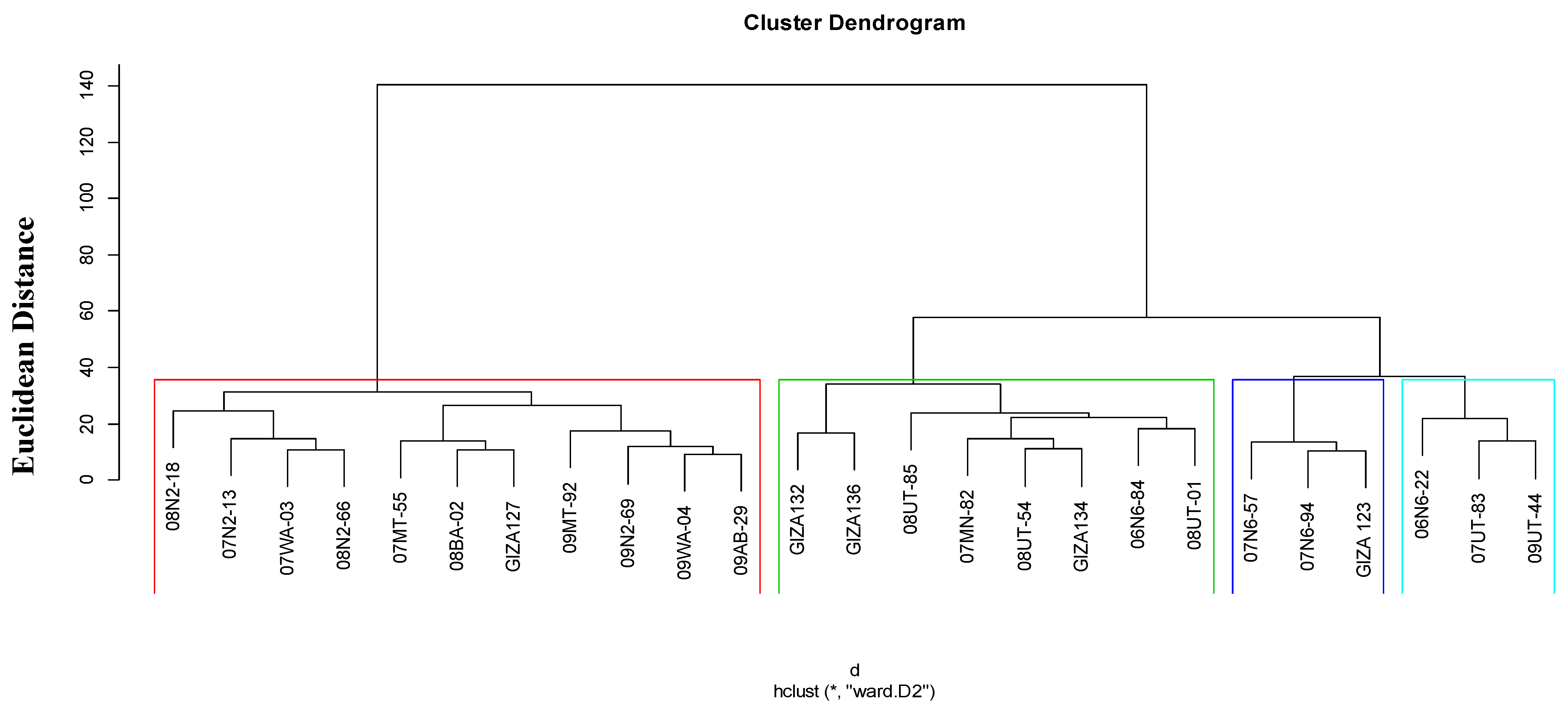
3.4. Relationships among Barley Genotypes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Katja, W.; Weidner, A.; Surabhi, G.K.; Börner, A.; Mock, H.P. Salt Stress-Induced Alterations in the Root Proteome of Barley Genotypes with Contrasting Response towards Salinity. J. Exp. Bot. 2009, 60, 3545–3557. [Google Scholar]
- Reddy, M.; Reddy, P.; Prasad, B.N.; Munilakshmi, U. Grain and Milling Quality of Barley and Their Suitability for Preparation of Traditional South Indian Products. Int. Organ. Sci. Res. J. Pharm. 2014, 4, 23–27. [Google Scholar]
- Wosene, G.A.; Berhane, L.; Bettina, I.G.H.; Karl, J.S. Ethiopian Barley Landraces Show Higher Yield Stability and Comparable Yield to Improved Varieties in Multi-Environment Field Trials. J. Plant Breed. Crop Sci. 2015, 7, 275–291. [Google Scholar] [CrossRef]
- Oscarsson, M.; Andersson, R.; Salomonsson, A.C.; Åman, P. Chemical composition of barley samples focusing on dietary fibre components. J. Cereal Sci. 1996, 24, 161–170. [Google Scholar] [CrossRef]
- Martinez, V.M.; Newman, K.; Newman, C.W. Barley diets with different fat sources have hypocholesterolemic effects in chickens. J. Nutr. 1991, 122, 1070–1076. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.W.; Deakins, D.A.; Floore, T.L.; Smith, B.M.; Whitis, S.E. Dietary fibre and coronary heart disease. Crit. Rev. Food Sci. Nutr. 1990, 29, 95–147. [Google Scholar] [CrossRef] [PubMed]
- Gosain, K. Long-Term Effects of Barley Bread Products on Metabolic Control of Non-Insulin-Dependent Diabetes Mellitus. Master’s Thesis, University of Alberta, Edmonton, AB, Canada, 1996. [Google Scholar]
- Ghassemi, F.; Jakeman, A.J.; Nix, H.A. Salinisation of Land and Water Resources: Human Causes, Extent, Management and Case Studies; UNSW Press: Sydney, Australia; CAB International: Wallingford, UK, 1995. [Google Scholar]
- Ouda, S.A.; Gaballah, M.S. Effect of Using Stabilizing Agents on Increasing Yield and Water Use Efficiency in Barley Grown under Water Stress. Aust. J. Basic Appl. Sci. 2007, 1, 571–577. [Google Scholar]
- El-Banna, M.N. Evaluation of 16 Barley Genotypes under Calcareous Soil Conditions in Egypt. J. Agric. Sci. 2012, 3, 105–121. [Google Scholar]
- FAO (Food and Agriculture Organization). Available online: http://www.fao.org/faostat/en/#data/QC.2016 (accessed on 5 October 2017).
- Davies, M.S.; Brown, A.H.D.; Marshall, D.R.; Frankel, O.H.; Williams, J.T. The Use of Plant Genetic Resources. J. Ecol. 1989, 77, 1175. [Google Scholar] [CrossRef]
- Moore, G. Strategic pre-breeding for wheat improvement. Nat. Plants 2015, 1, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Nass, L.L.; Paterniani, E. Pre-Breeding: A Link between Genetic Resources and Maize Breeding. Sci. Agric. 2000, 57, 581–587. [Google Scholar] [CrossRef]
- The Global Crop Diversity Trust. Available online: http://www.croptrust.org/main/sharingknowled.php?itemid =299 (accessed on 20 January 2012).
- Shimelis, H.; Laing, M. Timelines in conventional crop improvement: Pre-breeding and breeding procedures. AJCS 2012, 6, 1542–1549. [Google Scholar]
- Sharma, S.; Upadhyaya, H.D.; Varshney, R.K.; Gowda, C.L.L. Pre-breeding for diversification of primary gene pool and genetic enhancement of grain legumes. Front. Plant Sci. 2013. [Google Scholar] [CrossRef] [PubMed]
- Black, C.A. (Ed.) Method of Soil Analysis, Part 2, Chemical and Microbiological Properties; American Society of Agronomy Inc.: Madison, WI, USA, 1965. [Google Scholar]
- Chanda, S.V.; Singh, Y.D. Estimation of Leaf Area in Wheat Using Linear Measurements. Plant Breed. Seed Sci. 2002, 46, 75–79. [Google Scholar]
- Peterson, R.F.; Campbell, A.B.; Hannah, A.E. A diagrammatic scale of estimating rust intensity on leaves and stems of cereals. Can. J. Res. 1948, 26, 496–500. [Google Scholar] [CrossRef]
- Stakman, E.C.; Stewart, D.M.; Loegering, W.Q. Identification of Physiologic Races of Puccinia Graminis var. Tritici; United State Department of Agriculture: Washington, DC, USA, 1962.
- Negassa, M. Possible new genes for resistance to powdery mildew, Septoria glume blotch and leaf rust of wheat. Plant Breed. 1987, 98, 37–46. [Google Scholar] [CrossRef]
- Gomez, K.A.; Gomez, A.A. Statistical Procedures for Agricultural Research; John Wiley & Sons: New York, NY, USA, 1984. [Google Scholar]
- Steel, R.G.; Torrie, T.H. Principles and Procedures of Statistics, 2nd ed.; Mc. Graw-Hill Book Co.: New York, NY, USA, 1980. [Google Scholar]
- Cochran, W.G.; Cox, G.M. Experimental Designs, 2nd ed.; John Wiley& Sons Inc.: New York, NY, USA, 1957. [Google Scholar]
- Murtagh, F.; Legendre, P. Ward’s Hierarchical Clustering Method: Clustering Criterion and Agglomerative Algorithm. arXiv, 2011; arXiv:1111.6285. [Google Scholar]
- R Core Team. The R Project for Statistical Computing CRAN-Mirrors. Available online: http://www.rproject.org/ (accessed on 7 September 2017).
- Husson, F.; Josse, J.; Le, S.; Maintainer, J.M. Facto MineR: An R Package for Multivariate Analysis. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar]
- Gutie´ Rrez-Rodri´Guez, M.; Reynolds, M.P.; Larque-Saavedra, A. Photosynthesis of wheat in a warm, irrigated environment. II. Traits associated with genetic gains in yield. Field Crops Res. 2000, 66, 51–62. [Google Scholar] [CrossRef]
- Labuschagne, M.T.; Verhoeven, R.; Nkouanessi, M. Drought tolerance assessment of African cowpea accessions based on stomatal behaviour and cell membrane stability. J. Agric. Sci. 2008, 146, 689–694. [Google Scholar] [CrossRef]
- Hamam, K.A.; Khaled, A.S.G.A. Stability of Wheat Genotypes under Different Environments and Their Evaluation under Sowing Dates and Nitrogen Fertilizer Levels. Aust. J. Basic Appl. Sci. 2009, 3, 206–217. [Google Scholar]
- Abdul, A.; Altuhaish, K. Field Adaptation of Some Introduced Wheat (Triticum Aestivum L.) Genotypes in Two Altitudes of Tropical Agro-Ecosystem Environment of Indonesia. HAYATI J. Biosci. 2014, 21, 31–38. [Google Scholar]
- Ali, F.; Ahsan, M.; Ali, Q.; Kanwal, N. Phenotypic Stability of Zea Mays Grain Yield and Its Attributing Traits under Drought Stress. Front. Plant Sci. 2017, 8, 1397. [Google Scholar] [CrossRef] [PubMed]
- Mutwali, N.I.A.; Mustafa, A.I.; Gorafi, Y.S.A.; Mohamed Ahmed, I.A. Effect of Environment and Genotypes on the Physicochemical Quality of the Grains of Newly Developed Wheat Inbred Lines. Food Sci. Nutr. 2016, 4, 508–520. [Google Scholar] [CrossRef] [PubMed]
- Creissen, H.E.; Jorgensen, T.H.; Brown, J.K.M. Increased Yield Stability of Field-Grown Winter Barley (Hordeum Vulgare L.) Varietal Mixtures through Ecological Processes. Crop Prot. 2016, 85, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Dimova, D.; Krasteva, L.; Panayotov, N.; Svetleva, D.; Dimitrova, M. Evaluation of the Yield and the Yield Stability of Perspective Lines of Barley. Agroznanje 2012, 13, 55–60. [Google Scholar] [CrossRef]
- Adel, A.G.; Al-Ameiri, N.S.; Karajeh, M.R. Resistance of Barley Landraces and Wild Barley Populations to Powdery Mildew in Jordan. Phytopathol. Mediterr. 2008, 47, 92–97. [Google Scholar]
- Adawy, S.S.; Saker, M.M.; Haggag, W.M.; El-Itriby, H.A. Amplified Fragment Length Polymorphism (ALFP) Based Molecular Analysis of Egyptian Barley Lines and Landraces Differing in Their Resistance and Susceptibility to Leaf Rust and Net Blotch Diseases. Landbauforsch. Volkenrode 2008, 58, 125–134. [Google Scholar]
- Najaphy, A.; Ashrafi Parchin, R.; Farshadfar, E. Comparison of phenotypic and molecular characterization of some important wheat cultivars and advanced breeding lines. Aust. J. Crop Sci. 2012, 6, 326–332. [Google Scholar]
- Ribot, G.G.; Silva, P.; Acevedo, E. Morphological and Physiological Traits of Assistance in the Selection of High Yielding Varieties of Durum Wheat (Triticum Turgidum L. Spp. Durum) for the Rainfed Mediterranean Environments of Central Chile. Am. J. Plant Sci. 2012, 3, 1809–1819. [Google Scholar] [CrossRef]
- Rahman, M.; Barma, N.C.D.; Biswas, B.K.; Khan, A.A.; Rahman, J. Study on morpho-physiological traits in spring wheat (Triticum Aestivum L.) under rainfed condition m. Pakistan. J. Biosci. 2016, 7122, 235–250. [Google Scholar]
- Ajalli, J.; Salehi, M. Evaluation of Drought Stress Indices in Barley (Hordeum Vulgare L.). Ann. Biol. Res. 2012, 3, 5515–5520. [Google Scholar]
- Al-Tabbal, J.A.; Al-Fraihat, H.; Genetic Variation, A. Heritability, Phenotypic and Genotypic Correlation Studies for Yield and Yield Components in Promising Barley Genotypes. J. Agric. Sci. 2011, 4, 193–210. [Google Scholar] [CrossRef]
- Aina, O.O.; Dixon, A.G.O.; Akirinde, E.A. Additive Main Effects and Multiplicative Interaction (AMMI) Analysis for Yield of Cassava in Nigeria. J. Biol. Sci. 2007, 7, 796–800. [Google Scholar]
- Singh, N. Study of heritability and genetic advance in barley (Hordeum Vulgare L.) on affected soil. Int. J. Food Agric. Vet. Sci. 2012, 2, 35–39. [Google Scholar]
- Leilah, A.A.; Al-Khateeb, S.A. Statistical analysis of wheat yield under drought conditions. J. Arid Environ. 2005, 61, 483–496. [Google Scholar] [CrossRef]
- Ismail, A.A. Identification of selection traits for yield improvement of bread wheat using path analysis. Assiut J. Agric. Sci. 2001, 32, 63–84. [Google Scholar]
- Khokhar, M.I.; Hussain, M.; Zulkiffal, M.; Ahmad, N.; Sabar, W. Correlation and path analysis for yield and yield contributing characters in wheat (Triticum aestivum L.). Afr. J. Plant Sci. 2010, 4, 464–466. [Google Scholar]
- Ashraf, A.; El-Mohsen, A. Correlation and Regression Analysis. Sci. Res. Rev.-J. 2012, 1, 88–100. [Google Scholar]
- Ruzdik, N.M.; Valcheva, D.; Vulchev, D.; Mihajlov, L.; Karov, I.; Ilieva, V. Correlation between grain yield and yield components in winter barley varieties. Agric. Sci. Technol. 2015, 7, 40–44. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NO | Genotype | Synonym | Row Type | Pedigree | Origin |
|---|---|---|---|---|---|
| 1 | MT070107 | 07MT-55 | 2R | MT970148/Haxby | USA |
| 2 | Z155U012V | 08BA-02 | 2R | Z010C020E/Z011L088L | |
| 3 | 2ND27344 | 08N2-18 | 2R | ND24289/ND22996//CONLON | |
| 4 | NEG2-03 | 07MN-82 | 6R | M119/M99-106 NB | |
| 5 | 2ND26318 | 07N2-13 | 2R | ND21957-2/ND23024 | |
| 6 | 2ND27678 | 09N2-69 | 2R | 2ND24175/TR05285 | |
| 7 | 2159-90 | 08UT-85 | 6R | AB 11469 × Aquila | |
| 8 | MT090234 | 09MT-92 | 2R | MT910189/MT910189/Lk644/Eslick BC3F3 7-L | |
| 9 | 07WA-605.17 | 09WA-04 | 2R | WA 9004-99/Merit | |
| 10 | 2AB08-X05M046-21 | 09AB-29 | 2R | 2AB08-X05M046-21 | |
| 11 | ND23366 | 06N6-84 | 6R | ND19656/ND19703 | |
| 12 | 2142-69 | 07UT-83 | 6R | SDB11009/M72395/3/Short2//ID633019/Woodvale/4/Steptoe/M27//Gusto/5/WA11825-95/6//Harrington | |
| 13 | 2155-54 | 08UT-54 | 6R | Goldeneye × UT97B1556-5344 | |
| 14 | 2152-3 | 08UT-01 | 6R | Morex × Goldeneye | |
| 15 | 05WA-319.12 | 07WA-03 | 2R | Bob/Merit//CDC Select | |
| 16 | UT2170-146 | 09UT-44 | 6R | Harrington/3/Kombar/SF8623//Maranna | |
| 17 | ND25736 | 07N6-94 | 6R | ND19474/ND20477 | |
| 18 | ND25697 | 07N6-57 | 6R | STELLAR-ND/ND20603 | |
| 19 | ND23541 | 06N6-22 | 6R | Drummond/ND20414 | |
| 20 | 2ND27392 | 08N2-66 | 2R | ND24519/CONLON | |
| 21 | Giza123 | - | 6R | GIZA117/FAO86 | Egypt |
| 22 | Giza127 | - | 2R | W12291/Bags//Harmal-02 | |
| 23 | Giza132 | - | 6R | Rihane-05//AS 46/Aths*2Athe/Lignee 686 | |
| 24 | Giza134 | - | 6R | - | |
| 25 | Giza136 | - | 6R | - |
| Trait | Source of Variance | df | Mean Squares | ||
|---|---|---|---|---|---|
| E1 | E2 | E3 | |||
| 1000-grain weight | Genotypes | 24 | 326.8 ** | 179.52 ** | 6.99 ** |
| Replicates | 2 | 2.89 | 17.33 | 0.64 | |
| Leaf chlorophyll content | Genotypes | 24 | 249.5 ** | 136.46 ** | 54.61 ** |
| Replicates | 2 | 10.68 | 124.4 | 9.488 | |
| Leaf rust | Genotypes | 24 | 25.60 ** | 20.694 ** | 2321.01 ** |
| Replicates | 2 | 0.093 | 0.28 | 7.413 | |
| Net blotch | Genotypes | 24 | 1 ns | 1 ns | 11.32 ** |
| Replicates | 2 | 1 | 1 | 0.56 | |
| Powdery mildew | Genotypes | 24 | 1 ns | 12.97 ** | 1 ns |
| Replicates | 2 | 1 | 0.77 | 1 | |
| Trait | Source of Variance | df | S.S | MS |
|---|---|---|---|---|
| Number of days to flowering | E | 2 | 1281.7 | 640.87 ** |
| Replicates (E) | 6 | 16.68 | 2.78 | |
| G | 24 | 9618.4 | 400.76 ** | |
| E × G | 48 | 3846 | 80.12 ** | |
| Plant height | E | 2 | 4567.1 | 2283.5 ** |
| Replicates (E) | 6 | 85.55 | 14.26 | |
| G | 24 | 13,262.9 | 552.6 ** | |
| E × G | 48 | 10,347.7 | 215.58 ** | |
| Number of grains/spike | E | 2 | 675 | 337.5 ** |
| Replicates (E) | 6 | 123.2 | 20.5 | |
| G | 24 | 66796 | 2783.2 ** | |
| E × G | 48 | 3764 | 78.42 ** | |
| Grain yield | E | 2 | 86.28 | 43.14 ** |
| Replicates (E) | 6 | 3.01 | 0.50 | |
| G | 24 | 250.43 | 10.43 ** | |
| E × G | 48 | 177.65 | 3.70 ** | |
| Leaf area | E | 2 | 1508.8 | 754.4 ** |
| Replicates (E) | 6 | 91.85 | 15.31 | |
| G | 24 | 8292.6 | 345.5 ** | |
| E × G | 48 | 2335.3 | 48.7 ** |
| CHL | DF | PH | NG/S | TGW | LR | NB | PM | LA | GY | |
|---|---|---|---|---|---|---|---|---|---|---|
| CHL | 1 | |||||||||
| DF | −0.4 * | 1 | ||||||||
| PH | 0.41 * | −0.33 | 1 | |||||||
| NG/S | 0.25 | −0.26 | 0.76 ** | 1 | ||||||
| TGW | 0.23 | −0.42 * | −0.10 | −0.17 | 1 | |||||
| LR | −0.001 | 0.20 | 0.43 * | 0.50 * | −0.33 | 1 | ||||
| NB | 0.12 | −0.15 | −0.33 | −0.22 | −0.07 | −0.14 | 1 | |||
| PM | 0.02 | −0.11 | −0.19 | 0.02 | −0.17 | 0.02 | 0.88 ** | 1 | ||
| LA | 0.43 * | −0.18 | 0.56 * | 0.77 ** | −0.07 | 0.44 * | 0.07 | 0.29 | 1 | |
| GY | 0.27 | −0.55 ** | 0.21 | 0.22 | 0.54 ** | −0.25 | −0.01 | −0.1 | 0.26 | 1 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Naser, M.; Badran, M.; Abouzied, H.; Ali, H.; Elbasyoni, I. Phenotypic and Physiological Evaluation of Two and Six Rows Barley under Different Environmental Conditions. Plants 2018, 7, 39. https://doi.org/10.3390/plants7020039
Naser M, Badran M, Abouzied H, Ali H, Elbasyoni I. Phenotypic and Physiological Evaluation of Two and Six Rows Barley under Different Environmental Conditions. Plants. 2018; 7(2):39. https://doi.org/10.3390/plants7020039
Chicago/Turabian StyleNaser, Mahmoud, Mohamed Badran, Hanaa Abouzied, Heba Ali, and Ibrahim Elbasyoni. 2018. "Phenotypic and Physiological Evaluation of Two and Six Rows Barley under Different Environmental Conditions" Plants 7, no. 2: 39. https://doi.org/10.3390/plants7020039
APA StyleNaser, M., Badran, M., Abouzied, H., Ali, H., & Elbasyoni, I. (2018). Phenotypic and Physiological Evaluation of Two and Six Rows Barley under Different Environmental Conditions. Plants, 7(2), 39. https://doi.org/10.3390/plants7020039