Progress toward Understanding the Molecular Basis of Fruit Response to Hypoxia


Italian National Research Council (CNR), Via Giuseppe Moruzzi 1, 56127 Pisa, Italy
Plants 2018, 7(4), 78; https://doi.org/10.3390/plants7040078
Submission received: 26 August 2018
/
Revised: 19 September 2018
/
Accepted: 20 September 2018
/
Published: 21 September 2018
(This article belongs to the Special Issue Fruit Biology)
Abstract
:Oxygen has shaped life on Earth as we know it today. Molecular oxygen is essential for normal cellular function, i.e., plants need oxygen to maintain cellular respiration and for a wide variety of biochemical reactions. When oxygen levels in the cell are lower than levels needed for respiration, then the cell experiences hypoxia. Plants are known to experience root hypoxia during natural environmental conditions like flooding. Fruit, on the other hand, is known to be hypoxic under normal oxygen conditions. This observation could be explained (at least partially) as a consequence of diffusional barriers, low tissue diffusivity, and high oxygen consumption by respiration. From the physiological point of view, hypoxia is known to have a profound impact on fruit development, since it is well documented that a low oxygen environment can significantly delay ripening and senescence of some fruit. This effect of a low-oxygen environment is readily used for optimizing storage conditions and transport, and for prolonging the shelf life of several fruit commodities. Therefore, further understanding of the complex relationship between oxygen availability within the cell and fruit development could assist postharvest management.
1. Introduction
Fruit is a reproductive organ of flowering plants (angiosperms), which can be found in a myriad of sizes, shapes, textures, colors, flavors, and odors. Through the course of ripening, the physiology of fruit changes developmentally [1,2], making the ripe fruit attractive to animals, which, by eating the fruit, will disperse the seeds and improve the survival chances of the next generation of the plant [3]. From the human point of view, fruits are a delicious dietary product, highly valued for their nutritive properties [4,5]. On the basis of the ripening behavior, fruit can be classified as climacteric or nonclimacteric [6,7]. In climacteric fruit, ripening is accompanied by a peak in respiration and a concomitant burst of ethylene, whereas in nonclimacteric fruit, respiration shows no dramatic change and ethylene production remains at basal level [8]. As early as 1821, Berard recognized that O2 elimination in the storage environment can prevent ripening. Over the course of time, many methods have been developed to efficiently control the ripening of many different fruit commodities. Nowadays, setting the oxygen concentration in storage rooms is one of the most important aspects of postharvest protocols applied to specific fruit, aimed at preserving quality, prolonging shelf life, reducing losses, and obtaining higher market prices [9]. Specifically, controlled atmosphere (CA) storage is based on reducing oxygen and increasing carbon dioxide levels in the environment surrounding the fruit, profoundly affecting general metabolism (respiration in particular), ripening, and senescence [10].
Although oxygen is of paramount importance for normal development, fruits lack an active transport mechanism to distribute oxygen to all cells and a steep oxygen gradient occurs within most fruit tissues, mainly due to respiratory activity [11,12]. Furthermore, different oxygen levels inside the fruit and different tissue diffusivity among varieties could be important factors influencing postharvest quality parameters (such as firmness, shelf life, storage requirements, etc.).
In this paper, first, internal oxygen exchange in the fruit tissue is reviewed. Second, the most important metabolic responses to hypoxia are briefly discussed as well as low-oxygen-based fruit storage. Finally, the oxygen sensing and signaling pathway via N-end rule-mediated degradation of ethylene responsive factor (ERF)-VII proteins is reviewed.
2. Oxygen Exchange in Fruit Tissue
Oxygen is one of the most important elements required to sustain life. In order to supply cells with needed oxygen, fruit tissue mainly relays on oxygen transport by diffusion according to Fick’s law, assuming an effective diffusion process, which is driven by concentration gradients [13,14]. However, fruit skin represents a gas diffusion barrier, due to different layers of tissues (aqueous, cuticular, and waxy layers) [15,16,17,18]. Oxygen diffusion through the skin from openings (stomata and lenticels) is proportional to the difference in concentrations of oxygen across the barrier, total area of the skin, solubility of gas in the skin, solid-state diffusion coefficient, and total hole area available on the skin surface (as contributed by openings of stomata and/or lenticels) [18,19]. Therefore, gaseous oxygen first needs to diffuse through the fruit skin, and then, by the process of diffusion, enters into an intercellular system that forms a lattice and serves as the main pathway of oxygen transport [20,21]. However, the efficiency of oxygen transport within the intercellular system will depend on the porosity of the tissue (gas volume/tissue volume), and highly porous tissue has more enhanced oxygen transport compared to less porous tissue [22]. For example, in apple (Red Delicious) the porosity may reach an average of 25% of the total fruit volume [23], while in peach (Miraflores) it accounts for only about 3% of the tissue volume [24] (Table 1).
From the intracellular space, oxygen permeates through the cellular membrane to the cytoplasm, and finally into the mitochondria [21]. When respiration takes place, the concentration gradients appear because of consumption of O2 and production of CO2 [25,27]. Steep oxygen gradients inside fruit tissues seems to be rather a common phenomenon, especially in large bulky fruit such as pears and apples [28,29]. Oxygen gradients can be observed macroscopically in pears and apples, where the lowest oxygen concentrations are found in the fruit core [27,30]. CO2 diffusivity in fruit tissues can be higher than O2 diffusivity, causing a larger outflow of CO2 than inflow of O2, and a pressure difference between the inside of the fruit and the external atmosphere may develop. This is probably due to the greater solubility of CO2 than O2 in water [27]. Hence, besides gas diffusion driven by concentration gradients, gas exchange in the fruit may occur by permeation due to pressure gradients in the fruit tissues [27,31]. Additionally, when observed microscopically, much more gas diffuses through the pore and cell wall network than through the cytoplasm; therefore, cytoplasm has much lower oxygen concentration than intracellular space [21]. Therefore, hypoxia can occur under normal air conditions in some fruit (especially in the core tissue). However, it is important to note that oxygen gradient can affect fruit physiology profoundly, since hypoxia is known to affect several different biochemical processes.
3. Effects of Oxygen Availability on Metabolism and Development
Hypoxia is a condition in which a deficiency of cellular oxygen limits mitochondrial respiration, whereas anoxia is a condition in which the fluxes of respiratory gases are restricted and adenosine triphosphate (ATP) produced by mitochondria is negligible relative to ATP produced by glycolysis [32], which eventually will lead to cell death [33]. However, as mentioned previously, fruit tissue (especially the cortex of bulky fruit) can be hypoxic at normal air oxygen concentrations [12,27]. Respiration by the fruit tissues and barriers of diffusion/exchange of gases as posed by anatomical, morphological, physical, and/or biochemical components (present either on the surface or inside the fruit) are the factors responsible for the gradual lowering of the O2-to-CO2 ratio from outside to inside the fruit [34]. This tissue-specific hypoxia under normal air oxygen concentrations was observed also in other plant organs (roots, rhizomes, seeds, coleoptiles) and was shown to lead to the development and growth of an anaerobic core in roots [35,36].
Fruit response to hypoxia is mainly reflected in a reduction of cellular respiration followed by induction of anaerobic respiration, which leads to the fast breakdown of sugars [37,38]. Low oxygen is known to affect the transport chain [39,40], glycolysis [41], ethanolic fermentation [42], tricarboxylic acid (TCA) cycle [43], Yang cycle [44], and amino acid metabolism [45]. Recently, cell death was correlated with steep O2 gradients across the skin and toward the middle of the mesocarp in grape berries [46].
Several comprehensive transcriptomic studies of hypoxic fruit tissue resulted in the identification of the whole set of differentially expressed genes, providing a descriptive overview of the fruit-specific transcriptomic adjustments to oxygen deprivation [47,48,49]. For example, gene expression of alcohol dehydrogenase (ADH) is correlated with oxygen availability [50], and enzymatic activity of ADH and pyruvate decarboxylase (PDC) increases under hypoxia [39,51,52].
Hypoxia also decreases ethylene production and tissue sensitivity to ethylene [53]. Specifically, oxygen is a substrate for the reaction catalyzed by 1-aminocyclopropane-1-carboxylic acid (ACC) ACC-oxidase, an enzyme involved in ethylene biosynthesis. Therefore, oxygen is required for the synthesis as well as the action of ethylene in fruits [13,51,54]. Another important effect of hypoxia on fruit physiology is delayed ripening and senescence. This effect of hypoxia is adapted in agricultural storage methods, therefore it comes as no surprise that most of our understanding of fruit response to hypoxia comes from the research of fruit response to low-oxygen storage.
Physiological Effects of Low-Oxygen-Based Storage
As mentioned previously, one of the most important effects of hypoxia is delayed ripening and senescence of some fruit, especially considering its application in commercial fruit storage, transport, and packaging. This effect of hypoxia is adapted in the so-called controlled atmosphere (CA) storage method, in which the concentrations of oxygen, carbon dioxide, and nitrogen are strictly controlled along with the temperature and humidity of the storage room [10]. On the other hand, modified atmosphere (MA) is the practice of modifying the composition of the internal atmosphere of a package of fruit (fresh or cut) in order to improve the shelf life [9].
The most common CA method is used on apples and pears, in which the combination of altered atmospheric conditions and reduced temperature allows prolonged storage. Storage can reach up to one year for some apple varieties (Granny Smith) when applying a CA-based storage protocol [10]. Apart from delaying ripening, CA-based storage has been shown to have several other important positive effects in terms of postharvest parameters. In apple fruit, for example, CA storage is known to enhance several desirable attributes, such as firmness and taste retention, higher titratable acidity, maintenance of soluble solid content, and better skin color [9,10]. Furthermore, low-oxygen storage may also be effective in controlling superficial scald, a symptom of chilling injury that commonly occurs, for example, in Granny Smith apples [55]. However, if not carefully managed, CA storage can result in negative organoleptic/sensorial parameters, such as the production of off-flavors due to ethanol and acetaldehyde accumulation [9,10], underlining the importance of understanding the physiologic response to low oxygen in the storage room.
Physiologically, a low-oxygen storage effect on climacteric fruit ripening is reflected through O2 influence on respiration, largely through its inhibitory effect on ethylene action [56]. On the molecular level, the action of low oxygen on fruit ripening involves suppression of the de novo synthesized ripening proteins and mRNA transcripts, induction of new proteins and mRNA species, and constitutive expression of preexisting proteins or mRNA species [50,51]. Furthermore, hypoxia appears to modulate not only gene expression but also posttranscriptional and posttranslational processes [47]. Additionally, low-oxygen storage is known to affect production of volatiles, cell wall–related metabolism, and overall energy-related metabolism [51,57,58], and it can reduce cytosolic pH and energy charge [39,40].
These studies indicate that low-oxygen-based storage affects ripening at different metabolic and molecular levels, pointing out the importance of such research efforts. Additionally, understanding metabolomic changes and identifying metabolic markers during low-oxygen-storage could be useful in optimizing protocols.
4. Activation of the Anaerobic Response by the N-End Rule Pathway
In several plant species (Arabidopsis, rice, and poplar), a set of 49 genes were identified as ubiquitously induced upon hypoxia and/or submergence, therefore, were named core hypoxia-responsive genes (HRGs) [59,60,61]. HRGs are encoding enzymes involved in sucrose catabolism, anaerobic fermentation (alcohol dehydrogenase and pyruvate decarboxylase), reactive oxygen species regulation, gene transcription, and proteins of unknown function [60]. Major transcriptional control over HRGs is exerted by Group VII ethylene response factors (ERFs), as these transcriptional factors are known to upregulate around 50% of A. thaliana HRGs [62,63,64]. Furthermore, an evolutionarily conserved 12 bp cis-regulatory motif, named hypoxia-responsive promoter element (HRPE), is enriched in promoters of hypoxia-responsive genes in multiple species. In Arabidopsis, 39 of the 49 core HRGs were identified as putative direct targets of the ERF-VIIs that are stabilized in oxygen-deficient cells, based on detectable HRPE [65].
A. thaliana encodes five ERF-VIIs (Table 2), two of which, HYPOXIA RESPONSIVE ERF1 (HRE1) and HRE2, are HRGs [60,65,66], whereas ERF-VII transcripts RELATED TO APETALA2.12 (RAP2.12), RAP2.2, and RAP2.3 accumulate constitutively under normoxic conditions [65,66,67,68,69]. RAP 2.3 and RAP 2.12 can bind to acyl-CoA-binding protein (ACBP) [63,70,71], which prevents their movement into the nucleus under aerobic conditions and protects them against proteasomal degradation in air [63]. Upon hypoxia, ERF-VII transcriptional factors (TFs) move into the nucleus, where they activate anaerobic gene expression. RAP2.2 and RAP2.12 are necessary to activate the transcription of anaerobic genes, whereas HREs sustain their transcription after activation by RAPs [63,65,72]. Upon reoxygenation, RAP2.12 is rapidly degraded via the N-end rule pathway and proteasome-mediated proteolysis to downregulate the hypoxic response [62,63].
In Arabidopsis, oxygen-dependent modification and targeted proteolysis of ERF-VII TFs is determined by their conserved N-terminal motif (Met-Cys-Gly-Gly-Ala-Ile/Leu, MCGGAI/L, termed the MC motif), which is recognized by the enzymes involved in the N-end rule pathway [63]. First, the terminal Met is removed from the protein by methionine aminopeptidase (MAP1/2), leaving the second amino acid of the ERF-VII protein (Cys) exposed [73]. This residue is then oxidized to cysteine sulphenic acid in an oxygen-dependent manner catalyzed by plant cysteine oxidase (PCO1/2) [74], followed by transfer of the Arg of amino-acylated tRNAArg to the N terminus of the oxidized NH2-Cys-ERF-VII, catalyzed by arginyl-tRNA transferase (ATE1/2). This triggers subsequent ubiquitination by the single subunit E3 ubiquitin ligase proteolysis 6 (PRT6) and targets the protein to the proteasome for degradation, which can occur in both the cytosol and the nucleus [62,75].
Oxygen-Dependent Degradation of Fruit-Specific ERF VII
The existence of the same mechanism of anaerobic response activation by the N-end rule pathway was demonstrated in apple fruit during low-oxygen storage [47] and partially in transgenic tomato fruit [76].
The apple genome contains eight genes coding for ERF-VII TFs [77], distinguished by a conserved N-terminal consensus MCGGAI (Figure 1). Putative apple ERF proteins (orthologous to the oxygen sensors AtRAP2.12 and AtRAP2.2) were found to be stabilized under hypoxic conditions (Figure 2), suggesting that the oxygen-sensing mechanism based on the N-end rule pathway and on the posttranslational regulation of group VII ERF protein is also present and active in apple fruit [47].
In tomato, there are five members of ERF subgroup E (AtERF VII orthologous genes), expressed in reproductive tissues (E1 and E2), vegetative tissue (E3), or ubiquitously (E4) [78]. Furthermore, these TFs are regulated by ethylene, and in the case of E1 also by auxin [78]. Lee et al. (2012) demonstrated that ERF6 (ERF.E4) acts as a negative regulator of carotenoid accumulation during tomato fruit ripening [79]. Recently, Liu et al. (2016) demonstrated that expression of ERF.E1, ERF.E2, and ERF.E4 is suppressed in the tomato-ripening mutants, suggesting their important regulatory role in fruit ripening [80]. Additionally, it seems that ERF.E members are the most active ERFs in ethylene- and RIN/NOR-dependent ripening [80]. All members of this tomato protein subgroup are characterized by the presence of an MCGGAI motif at the N-terminus, which qualifies ERF.E proteins as candidate substrates of the N-end rule pathway (Figure 1 and Figure 2). Research conducted on tomato suggested that ERF E genes are involved not only in the activation of anaerobic response by the N-end rule pathway, but also in fruit ripening and development, such as regulation of carotenoids [47,79], indicating the importance of understanding the relationship between hypoxia signaling via ERFs and fruit ripening.
Recently, Meitha et al. identified three ERF-VIIs in grape berries, which are substrates for oxygen-dependent N-end rule proteolysis as in Arabidopsis [81]. Additionally, several hypoxia-responsive ERFs were shown to be directly involved in transcriptional regulation of anaerobic metabolism genes involved in persimmon deastringency [82,83,84]; however, their possible oxygen-dependent regulation via the N-end rule pathway is still unknown. Although it is worth noting that DkERF10, based on an amino acid sequence, could be a candidate substrate of the N-end rule pathway of protein degradation. A recent study by Zhu et al. (2018) demonstrated the existence of a transcriptional cascade involving ERF-VII (DkERF9, 10, 18, 19, 21, 22) and MYB transcriptional factors (DkMYB6 and 10) that led to upregulation of the fermentation genes (DkADH1 and DkPDC2) [85].
Taken together, the results obtained for fruit-specific ERF genes (orthologous Arabidopsis ERF-VII) contribute to further understanding of the molecular basis of fruit response to hypoxia. Additionally, ERF-VII’s role in fruit hypoxia response and its physiological role during ripening highlights the importance of future research aimed at further characterization of its role in fruit and the complex oxygen-dependent regulatory mechanism.
5. Conclusions
In the last decade, increasing efforts of the scientific community have contributed greatly to our understanding of fruit low-oxygen response. Understanding the O2 response of fruit requires knowledge of fruit macrostructure, gas-exchange properties of the tissues, cellular microstructure, and respiration kinetics. To date, the complexity of fruit tissue response to hypoxia has been well explained, especially for bulky fruit during low-oxygen storage. Furthermore, the list of newly identified compounds (metabolites, volatiles, and specific proteins) is growing daily; this not only extends our understanding of complex metabolic response to low oxygen but also contributes greatly to fruit storage and transport optimization. Transcriptional control of around 50% of low-oxygen-responsive genes is regulated by ERFs (subgroup VII). Considering the important role of these ERFs in oxygen-sensing mechanisms, and the fact that fruit tissue is hypoxic under normal oxygen concentrations, it would be interesting to further investigate the possible involvement of ERFs in fruit ripening and development. An understanding of the changes in the metabolome, gene transcription patterns, and regulatory mechanisms and their complex relationship not only contributes to our understanding of molecular responses but can also be used to maximize fruit quality after harvest and during storage and could assist in fruit breeding.
Funding
This research received no external funding.
Conflicts of Interest
The author declares no conflict of interest.
References
- Barry, C.S.; Giovannoni, J.J. Ethylene and fruit ripening. J. Plant Growth Regul. 2007, 26, 143–159. [Google Scholar] [CrossRef]
- Bouzayen, M.; Latche, A.; Pech, J.C.; Nath, P. Mechanism of fruit ripening. In Plant Developmental Biology-Biotechnological Perspective; Pua, E.C., Davey, M.R., Eds.; Springer: Heidelberg/Berlin, Germany, 2010; Volume 1, pp. 319–339. [Google Scholar]
- Karlova, R.; Chapman, N.; David, K.; Angenent, G.C.; Seymour, G.B.; De Maagd, R.A. Transcriptional control of fleshy fruit development and ripening. J. Exp. Bot. 2014, 65, 4527–4541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Stacewiez-Sapuntzakis, M.; Duncan, C.; Sharifi, R.; Ghosh, L.; Breemen, R.; van Ashton, D.; Bowen, P.E. Oxidative DNA damage in prostate cancer patients consuming tomato saucebased entrees as a whole-food intervention. J. Natl. Cancer Inst. 2001, 93, 1872–1879. [Google Scholar] [CrossRef] [PubMed]
- Mares-Perlman, J.A.; Millen, A.E.; Ficek, T.L.; Hankinson, S.E. The body of evidence to support a protective role for lutein and zeaxanthin in delaying chronic disease. J. Nutr. 2002, 132, 5185–5245. [Google Scholar] [CrossRef] [PubMed]
- Biale, J.B.; Young, R.E. Respiration and ripening in fruits retrospective and prospect. In Recent Advances in the Biochemistry of Fruits and Vegetables; Friend, J., Rhodes, M.J.C., Eds.; Academic Press: New York, NY, USA, 1981; pp. 1–39. [Google Scholar]
- Lelievre, J.M.; Latche, A.; Jones, B.; Bouzayen, M.; Peach, J.C. Ethylene and fruit ripening. Physiol. Plant 1997, 101, 727–739. [Google Scholar] [CrossRef]
- Alexander, L.; Grierson, D. Ethylene biosynthesis and action in tomato: A model for climacteric fruit ripening. J. Exp. Bot. 2002, 53, 2039–2055. [Google Scholar] [CrossRef] [PubMed]
- Yahia, E.M. Modified and Controlled Atmospheres for the Storage, Transportation, and Packaging of Horticultural Commodities, 1st ed.; CRC Press: Boca Raton, FL, USA, 2009; pp. 1–10. [Google Scholar]
- Thompson, K.A. Controlled Atmosphere Storage of Fruits and Vegetables, 2nd ed.; CABI: Cambridge, UK, 2010; pp. 26–43. [Google Scholar]
- Verboven, P.; Kerckhofs, G.; Mebatsion, H.K.; Ho, Q.T.; Temst, K.; Wevers, M.; Cloetens, P.; Nicolai, B.M. Three-Dimensional Gas Exchange Pathways in Pome Fruit Characterized by Synchrotron X-Ray Computed Tomography. Plant Physiol. 2008, 147, 518–527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, Q.T.; Verboven, P.; Verlinden, B.E.; Nicolai, B.M. A model for gas transport in pear fruit at multiple scales. J. Exp. Bot. 2010, 61, 2071–2081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burg, S.P.; Burg, E.A. Gas exchange in fruits. Plant Physiol. 1965, 18, 870–874. [Google Scholar] [CrossRef]
- Cameron, A.C.; Yang, S.F. A simple method for the determination of resistance to gas diffusion in plant organs. Plant Physiol. 1982, 70, 21–23. [Google Scholar] [CrossRef] [PubMed]
- Solomos, T. Principles of gas exchange in bulky plant tissue. HortScience 1987, 22, 766–771. [Google Scholar]
- Schotsmans, W.; Verlinden, B.E.; Lammertyn, J.; Nicolaı, B.M. Simultaneous measurement of oxygen and carbon dioxide diffusivity in pear fruit tissue. Postharvest Biol. Technol. 2003, 29, 155–166. [Google Scholar] [CrossRef]
- Nobel, P.S. Physicochemical and Environmental Plant Physiology; Academic Press Inc.: London, UK, 1991; pp. 1–46. [Google Scholar]
- Hagenmaier, R. Method for measuring internal gases of citrus fruit and determining peel permeance. Proc. Fla. State Hortic. Soc. 2004, 116, 418–423. [Google Scholar]
- Hagenmaier, R. A comparison of ethane, ethylene and CO2 peel permeance for fruits with different coatings. Postharvest Biol. Technol. 2005, 37, 56–64. [Google Scholar] [CrossRef]
- Ben-Yehoshua, S. Gas exchange, transpiration and the commercial deterioration of stored orange fruit. J. Am. Soc. Hortic. Sci. 1969, 94, 524–526. [Google Scholar]
- Ho, Q.T.; Verboven, P.; Mebatsion, H.K.; Verlinden, B.E.; Vandewalle, S.; Nicolaï, B.M. Microscale mechanisms of gas exchange in fruit tissue. New Phytol. 2009, 182, 163–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armstrong, W. Aeration in higher plants. Adv. Bot. Res. 1979, 7, 225–332. [Google Scholar]
- Marcellin, P. Mesure de la diffusion des gas dans Ies organes vegetaux. Bull. Soc. Fr. Physiol. Veg. 1963, 9, 29. [Google Scholar]
- Salvatori, D.; Andrés, A.; Chiralt, A.; Fito, P. The response of some properties of fruit to vacuum impregnation. J. Food Process Eng. 1998, 21, 59–73. [Google Scholar] [CrossRef]
- Rajapakse, N.C.; Banks, N.H.; Hewett, E.W.; Cleland, D.J. Development of oxygen concentration gradients in flesh tissues of bulky plant organs. J. Am. Soc. Hortic. Sci. 1990, 115, 793–797. [Google Scholar]
- Sharifi, M.; Rafiee, S.; Keyhani, A.; Jafari, A.; Mobli, H.; Rajabipour, A.; Akram, A. Some physical properties of orange (var. Tompson). Int. Agrophys. 2007, 21, 391–397. [Google Scholar]
- Ho, Q.T.; Verboven, P.; Verlinden, B.E.; Lammertyn, J.; Vandewalle, S.; Nicolaï, B.M. A Continuum Model for Metabolic Gas Exchange in Pear Fruit. PLoS Comput. Biol. 2008, 4, e1000023. [Google Scholar] [CrossRef] [PubMed]
- Ho, Q.T.; Verboven, P.; Verlinden, B.E.; Herremans, E.; Wevers, M.; Carmeliet, J.; Nicolaï, B.M. A three-dimensional multiscale model for gas exchange in fruit. Plant Physiol. 2011, 155, 1158–1168. [Google Scholar] [CrossRef] [PubMed]
- Mellidou, I.; Buts, K.; Hatoum, D.; Ho, Q.T.; Johnston, J.W.; Watkins, C.B.; Schaffer, R.J.; Gapper, N.E.; Giovannoni, J.J.; Rudell, D.R. Transcriptomics events associated with internal browning of apple during postharvest storage. BMC Plant Biol. 2014, 14, 328. [Google Scholar] [CrossRef] [PubMed]
- Ho, Q.T.; Verlinden, B.E.; Verboven, P.; Vandewalle, S.; Nicolaï, B.M. A permeation-diffusion-reaction model of gas transport in cellular tissue of plant materials. J. Exp. Bot. 2006, 57, 4215–4224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, Q.T.; Verlinden, B.E.; Verboven, P.; Nicolaï, B.M. Gas diffusion properties at different positions in the pear. Postharvest Biol. Technol. 2006, 41, 113–120. [Google Scholar] [CrossRef]
- Drew, M.C. Oxygen deficiency and root metabolism: Injury and acclimation under hypoxia and anoxia. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1997, 48, 223–250. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.J.; Chen, K.S.; Ferguson, I.B. Programmed cell death features in apple suspension cells under low oxygen culture. J. Zhejiang Univ. Sci. 2004, 5, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Paul, V.; Pandey, R. Role of internal atmosphere on fruit ripening and storability—A review. Int. J. Food Sci. Technol. 2014, 51, 1223–1250. [Google Scholar] [CrossRef] [PubMed]
- Saglio, P.H.; Rancillac, M.; Bruzau, F.; Pradet, A. Critical oxygen pressure for growth and respiration of excised and intact roots. Plant Physiol. 1984, 76, 151–154. [Google Scholar] [CrossRef] [PubMed]
- Drew, M.C. Soil aeration and plant-root metabolism. Soil Sci. 1992, 154, 259–268. [Google Scholar] [CrossRef]
- Kader, A.A. Biochemical and physiological basis for effects of controlled and modified atmospheres on fruits and vegetables. Food Technol. 1986, 40, 99–104. [Google Scholar]
- Boersig, M.R.; Kader, A.A.; Romani, R.J. Aerobic-anaerobic respiratory transition in pear fruit and cultured pear fruit cells. J. Am. Soc. Hortic. Sci. 1988, 111, 869–873. [Google Scholar]
- Nanos, G.D.; Romani, R.J.; Kader, A.A. Metabolic and other response of ‘Bartlett’ pear fruit and suspension cultured ‘Passe Crassane’ pear fruit cells held in 0.25% O2. J. Am. Soc. Hortic. Sci. 1992, 117, 934–940. [Google Scholar]
- Saquet, A.A.; Streif, J.; Bangerth, F. Changes in ATP, ADP and pyridine nucleotide levels related to the incidence of physiological disorders in ‘Conference’ pears and ‘Jonagold’ apples during controlled atmosphere storage. J. Hortic. Sci. Biotechnol. 2000, 75, 243–249. [Google Scholar] [CrossRef]
- Kerbel, E.L.; Keder, A.A.; Romani, R.S. Effect of elevated CO2 concentrations on glycolysis in intact ‘Bartlet’ pear fruit. Plant Physiol. 1988, 86, 1205–1209. [Google Scholar] [CrossRef] [PubMed]
- Ke, D.; Zhou, L.; Kader, A.A. Mode of oxygen and carbon dioxide action on strawberry ester biosynthesis. J. Am. Soc. Hortic. Sci. 1994, 119, 971–975. [Google Scholar]
- Shipway, M.R.; Bramlage, W.J. Effects of carbon dioxide on activity of apple mitochondria. Plant Physiol. 1973, 51, 1095–1098. [Google Scholar] [CrossRef] [PubMed]
- Bulens, I.; Van de Poel, B.; Hertog, M.L.A.T.M.; Cristescu, S.M.; Harren, F.J.M.; De Proft, M.P.; Geeraerd, A.H.; Nicolai, B.M. Dynamic changes of the ethylene biosynthesis in “Jonagold” apple. Physiol. Plant 2014, 150, 161–173. [Google Scholar] [CrossRef] [PubMed]
- Hansen, M.E.; Sørensen, H.; Cantwell, M. Changes in acetaldehyde, ethanol and amino acid concentrations in broccoli florets during air and controlled atmosphere storage. Postharvest Biol. Technol. 2001, 22, 227–237. [Google Scholar] [CrossRef]
- Xiao, Z.; Rogiers, S.Y.; Sadras, V.O.; Tyerman, S.D. Hypoxia in grape berries: The role of seed respiration and lenticels on the berry pedicel and the possible link to cell death. J. Exp. Bot. 2018, 69, 2071–2083. [Google Scholar] [CrossRef] [PubMed]
- Cukrov, D.; Zermiani, M.; Brizzolara, S.; Cestaro, A.; Licausi, F.; Luchinat, C.; Santucci, C.; Tenori, L.; Van Veen, H.; Zuccolo, A.; et al. Extreme Hypoxic Conditions Induce Selective Molecular Responses and Metabolic Reset in Detached Apple Fruit. Front. Plant Sci. 2016, 7, 146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lara, M.V.; Budde, C.O.; Porrini, L.; Borsani, J.; Murray, R.; Andreo, C.S.; Drincovich, M.F. Peach (Prunus persica) fruit response to anoxia: Reversible ripening delay and biochemical changes. Plant Cell Physiol. 2011, 52, 392–403. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.; Choi, S.C.; Jung, S.; Cho, B.K.; Ahn, G.H.; Ryu, S.B. A Transcriptome Approach Toward Understanding Fruit Softening in Persimmon. Front. Plant Sci. 2017, 8, 1556. [Google Scholar] [CrossRef] [PubMed]
- Loulakakis, K.; Hassan, M.; Gerasopoulos, G.; Kanellis, A.K. Effects of low oxygen on in vitro translation products of poly(A)+ RNA, cellulase and alcohol dehydrogenase expression in preclimacteric and ripening-initiated avocado fruit. Postharvest Biol. Technol. 2016, 39, 29–37. [Google Scholar] [CrossRef]
- Kanellis, A.K.; Solomos, T.; Roubelakis-Angelakis, K.A. Supspecificities of cellulose and polygalacturonase isozymes in avocado fruit mesocarp subjected to low oxygen stress. Plant Physiol. 1991, 96, 269–274. [Google Scholar] [CrossRef] [PubMed]
- Chen, A.R.S.; Chase, T., Jr. Alcohol dehydrogenase and pyruvate decarboxylase induction in ripening and hypoxia tomato fruit. Plant Physiol. Biochem. 1993, 31, 875–885. [Google Scholar]
- Kader, A.A.; Zagory, D.; Kerbel, E.L. Modified atmosphere packaging of fruits and vegetables. Crit. Rev. Food Sci. Nutr. 1989, 28, 1–30. [Google Scholar] [CrossRef] [PubMed]
- Gorny, J.R.; Kader, A.A. Fresh-cut fruit products. In Fresh-Cut Products: Maintaining Quality and Safety; Cantwell, M., Ed.; University of California: San Diego, CA, USA, 1996; Volume 10, pp. 14–59. [Google Scholar]
- Wang, Z.; Dilley, D.R. Control of superficial scald of apples by low-oxygen atmospheres. Hortic. Sci. 1999, 34, 1145–1151. [Google Scholar]
- Lanahan, M.B.; Yen, H.C.; Giovannoni, J.J.; Klee, H.J. The never ripe mutation blocks ethylene perception in tomato. Plant Cell 1994, 6, 521–530. [Google Scholar] [CrossRef] [PubMed]
- Pasentsis, K.; Falara, V.; Pateraki, I.; Gerasopoulos, D.; Kanellis, A.K. Identification and expression profiling of low oxygen regulated genes from Citrus flavedo tissues using RT-PCR differential display. J. Exp. Bot. 2007, 58, 2203–2216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lumpkin, C.; Fellman, J.K.; Rudell, D.R.; Mattheis, J. Scarlett spur Red Delicious apple volatile production accompanying disorder development during low pO2 controlled atmosphere storage. J. Agric. Food Chem. 2014, 62, 1741–1754. [Google Scholar] [CrossRef] [PubMed]
- Mustroph, A.; Zanetti, M.E.; Jang, C.J.H.; Holtan, H.E.; Repetti, P.P.; Galbraith, D.W.; Girke, T.; Bailey-Serres, J. Profiling translatomes of discrete cell populations resolves altered cellular priorities during hypoxia in Arabidopsis. Proc. Natl. Acad. Sci. USA 2009, 106, 18843–18848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mustroph, A.; Lee, S.C.; Oosumi, T.; Zanetti, M.E.; Yang, H.; Ma, K.; Yaghoubi-Masihi, A.; Fukao, T.; Bailey-Serres, J. Cross-kingdom comparis on of transcriptomic adjustments to low-oxygen stress highlights conserved and plant-specific responses. Plant Physiol. 2010, 152, 1484–1500. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.C.; Mustroph, A.; Sasidharan, R.; Vashisht, D.; Pedersen, O.; Oosumi, T.; Voesenek, L.A.C.J.; Bailey-Serres, J. Molecular characterization of the submergence response of the Arabidopsis thaliana ecotype Columbia. New Phytol. 2011, 190, 457–471. [Google Scholar] [CrossRef] [PubMed]
- Gibbs, D.J.; Lee, S.C.; Isa, N.M.; Gramuglia, S.; Fukao, T.; Bassel, G.W.; Correia, C.S.; Corbineau, F.; Theodoulou, F.L.; Bailey-Serres, J.; et al. Homeostatic response to hypoxia is regulated by the N-end rule pathway in plants. Nature 2011, 479, 415–418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Licausi, F.; Kosmacz, M.; Weits, D.A.; Giuntoli, B.; Giorgi, F.M.; Voesenek, L.A.C.J.; Perata, P.; van Dongen, J.T. Oxygen sensing in plants is mediated by an N-end rule pathway for protein destabilization. Nature 2011, 479, 419–422. [Google Scholar] [CrossRef] [PubMed]
- Kosmacz, M.; Parlanti, S.; Schwarzländer, M.; Kragler, F.; Licausi, F.; van Dongen, J.T. The stability and nuclear localization of the transcription factor RAP2.12 are dynamically regulated by oxygen concentration. Plant Cell Environ. 2015, 38, 1094–1103. [Google Scholar] [CrossRef] [PubMed]
- Gasch, P.; Fundinger, M.; Müller, J.T.; Lee, T.; Bailey-Serres, J.; Mustroph, A. Redundant ERF-VII transcription factors bind to an evolutionarily conserved cis-motif to regulate hypoxia-responsive gene expression in Arabidopsis. Plant Cell 2015, 28, 60–80. [Google Scholar] [CrossRef] [PubMed]
- Licausi, F.; van Dongen, J.T.; Giuntoli, B.; Novi, G.; Santaniello, A.; Geigenberger, P.; Perata, P. HRE1 and HRE2, two hypoxia-inducible ethylene response factors, affect anaerobic responses in Arabidopsis thaliana. Plant J. 2010, 62, 302–315. [Google Scholar] [CrossRef] [PubMed]
- Büttner, M.; Singh, K.B. Arabidopsis thaliana ethylene-responsive element binding protein (AtEBP), an ethylene-inducible, GCC box DNA-binding protein interacts with an ocs element binding protein. Proc. Natl. Acad. Sci. USA 1997, 94, 5961–5966. [Google Scholar] [CrossRef] [PubMed]
- Papdi, C.; Abrahám, E.; Joseph, M.P.; Popescu, C.; Koncz, C.; Szabados, L. Functional identification of Arabidopsis stress regulatory genes using the controlled cDNA overexpression system. Plant Physiol. 2008, 147, 528–542. [Google Scholar] [CrossRef] [PubMed]
- Hinz, M.; Wilson, I.W.; Yang, J.; Buerstenbinder, K.; Llewellyn, D.; Dennis, E.S.; Sauter, M.; Dolferus, R. Arabidopsis RAP2.2: An ethylene response transcription factor that is important for hypoxia survival. Plant Physiol. 2010, 153, 757–772. [Google Scholar] [CrossRef] [PubMed]
- Li, H.-Y.; Chye, M.-L. Arabidopsis Acyl-CoA-binding protein ACBP2 interacts with an ethylene-responsive element-binding protein, AtEBP, via its ankyrin repeats. Plant Mol. Biol. 2004, 54, 233–243. [Google Scholar] [CrossRef] [PubMed]
- Li, H.-Y.; Xiao, S.; Chye, M.-L. Ethylene- and pathogen-inducible Arabidopsis acyl-CoA-binding protein 4 interacts with an ethylene-responsive element binding protein. J. Exp. Bot. 2008, 59, 3997–4006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bui, L.T.; Giuntoli, B.; Kosmacz, M.; Parlanti, S.; Licausi, F. Constitutively expressed ERF-VII transcription factors redundantly activate the core anaerobic response in Arabidopsis thaliana. Plant Sci. 2015, 236, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Gibbs, D.J.; Md Isa, N.; Movahedi, M.; Lozano-Juste, J.; Mendiondo, G.M.; Berckhan, S.; Marín-de la Rosa, N.; Vicente Conde, J.; Sousa Correia, C.; Pearce, S.P.; et al. Nitric oxide sensing in plants is mediated by proteolytic control of group VII ERF transcription factors. Mol. Cell 2014, 53, 369–379. [Google Scholar] [CrossRef] [PubMed]
- Weits, D.A.; Giuntoli, B.; Kosmacz, M.; Parlanti, S.; Hubberten, H.M.; Riegler, H.; Hoefgen, R.; Perata, P.; Van Dongen, J.T.; Licausi, F. Plant cysteine oxidases control the oxygen-dependent branch of the N-end-rule pathway. Nat. Commun. 2014, 5, 3425. [Google Scholar] [CrossRef] [PubMed]
- Licausi, F.; Weits, D.A.; Pant, B.D.; Scheible, W.R.; Geigenberger, P.; van Dongen, J.T. Hypoxia responsive gene expression is mediated by various subset of transcription factors and miRNAs that are determined by the actual oxygen availability. New Phytol. 2011, 190, 442–456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cukrov, D. Molecular Characterization of Fruit Responses to Low Oxygen. Doctoral Dissertation, Scuola Superiore Sant’Anna, Institute of Life Sciences, Pisa, Italy, 4 July 2016. [Google Scholar]
- Girardi, C.L.; Rombaldi, C.V.; Dal Cero, J.; Nobile, P.M.; Laurens, F.; Bouzayen, M.; Quecini, V. Genome-wide analysis of the AP2/ERF superfamily in apple and transcriptional evidence of ERF involvement in scab pathogenesis. Sci. Hortic. 2013, 151, 112–121. [Google Scholar] [CrossRef] [Green Version]
- Pirrello, J.; Prasad, N.; Zhang, W.; Chen, K.; Mila, I.; Zouine, M.; Latché, A.; Pech, J.C.; Ohme-Takagi, M.; Regad, F.; et al. Functional analysis and binding affinity of tomato ethylene response factors provide insight on the molecular bases of plant differential responses to ethylene. BMC Plant Biol. 2012, 12, 190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.; Joung, J.G.; McQuinn, R.; Chung, M-Y.; Fei, Z.; Tieman, D.; Klee, H.; Giovannoni, J.J. Combined transcriptome, genetic diversity and metabolite profiling in tomato fruit reveals that the ethylene response factor SlERF6 plays an important role in ripening and carotenoid accumulation. Plant J. 2012, 70, 191–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, M.; Lima Gomes, B.; Mila, I.; Purgatto, E.; Peres, L.E.P.; Frasse, P.; Maza, E.; Zouine, M.; Roustan, J.P.; Bouzayen, M.; et al. Comprehensive profiling of Ethylene Response Factors expression identifies ripening-associated ERF genes and their link to key regulators of fruit ripening in tomato (Solanum lycopersicum). Plant Physiol. 2016, 33, 01859. [Google Scholar] [CrossRef] [PubMed]
- Meitha, K.; Agudelo-Romero, P.; Signorelli, S.; Gibbs, D.J.; Considine, J.A.; Foyer, C.H.; Considine, M.J. Developmental control of hypoxia during bud burst in grapevine. Plant Cell Environ. 2018, 41, 1154–1170. [Google Scholar] [CrossRef] [PubMed]
- Min, T.; Yin, X.R.; Shi, Y.N.; Luo, Z.R.; Yao, Y.C.; Grierson, D.; Ferguson, I.B.; Chen, K.S. Ethylene-responsive transcription factors interact with promoters of ADH and PDC involved in persimmon (Diospyros kaki) fruit de-astringency. J. Exp. Bot. 2012, 63, 6393–6405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Min, T.; Fang, F.; Ge, H.; Shi, Y.N.; Luo, Z.R.; Yao, Y.C.; Grierson, D.; Yin, X.R.; Chen, K.S. Two novel anoxia-induced ethylene response factors that interact with promoters of deastringency-related genes from persimmon. PLoS ONE 2014, 9, e97043. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.M.; Zhu, Q.G.; Deng, C.L.; Luo, Z.R.; Sun, N.J.; Grierson, D.; Yin, X.R.; Chen, K.S. Hypoxia-responsive ERFs involved in postdeastringency softening of persimmon fruit. Plant Biotechnol. J. 2017, 15, 1409–1419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Q.G.; Gong, Z.Y.; Wang, M.M.; Li, X.; Grierson, D.; Yin, X.R.; Chen, K.S. A transcription factor network responsive to high CO2/hypoxia is involved in deastringency in persimmon fruit. J. Exp. Bot. 2018, 69, 2061–2070. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
ERF-VII protein sequence similarity among Arabidopsis, tomato, grapevine, and apple fruit. (A) Between-species phylogenetic tree representing relationships among ERF-VII proteins (represented values are real distance). (B) Multiple sequence alignment of ERF-VII proteins. Conserved MCGGAI amino acid sequence can clearly be seen. This amino acid sequence is recognized by the enzymes involved in the N-end rule pathway. Multiple sequence alignments were performed using Clustal Omega software (EMBL-EBI, September 2018).
Figure 1.
ERF-VII protein sequence similarity among Arabidopsis, tomato, grapevine, and apple fruit. (A) Between-species phylogenetic tree representing relationships among ERF-VII proteins (represented values are real distance). (B) Multiple sequence alignment of ERF-VII proteins. Conserved MCGGAI amino acid sequence can clearly be seen. This amino acid sequence is recognized by the enzymes involved in the N-end rule pathway. Multiple sequence alignments were performed using Clustal Omega software (EMBL-EBI, September 2018).

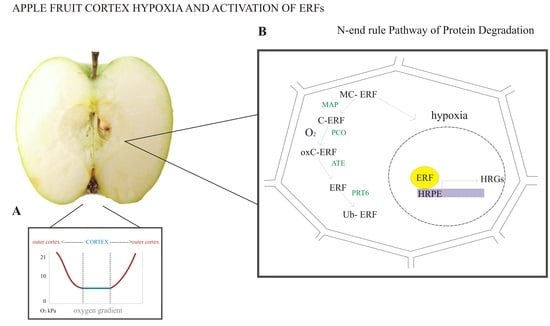
Figure 2.
Fruit tissue hypoxia and oxygen signaling pathway. (A) Bulky fruit, such as apple, can be hypoxic under a normal oxygen environment, where the lowest oxygen concentrations are found in the cortex. The graph illustrates the possible gradient formed inside the fruit in normal oxygen conditions (adapted from Paul and Pandey [34]). (B) The oxygen concentration in the cortex can reach very low values, and the cell can experience hypoxia. Hypoxia triggers the accumulation of ERFs in the nucleus, which can bind the hypoxia-responsive promoter element (HRPE) sequence, which in turn activates transcription of hypoxia-responsive genes (HRGs). On the other hand, under normal oxygen conditions, ERFs are subjected to enzymatic degradation via the N-end rule pathway. Enzymes are shown in green. MAP, methionine aminopeptidase; PCO, plant cysteine oxidase; ATE, arginyl-tRNA transferase; PRT6-E3, ubiquitin ligase proteolysis; MC-ERF, amino acid sequence of the protein; oxC-ERF, oxidation of the cysteine residue; Ub-ERF, ubiquitination of ERF. Although available literature indicates ERF degradation via the N-end rule pathway, genes coding the enzymes involved in the N-end rule pathway of protein degradation have yet to be identified. Therefore, this is an illustration of ERF oxygen signaling based on knowledge obtained in Arabidopsis and apple fruit (adapted from Licausi et al. [64]).
Figure 2.
Fruit tissue hypoxia and oxygen signaling pathway. (A) Bulky fruit, such as apple, can be hypoxic under a normal oxygen environment, where the lowest oxygen concentrations are found in the cortex. The graph illustrates the possible gradient formed inside the fruit in normal oxygen conditions (adapted from Paul and Pandey [34]). (B) The oxygen concentration in the cortex can reach very low values, and the cell can experience hypoxia. Hypoxia triggers the accumulation of ERFs in the nucleus, which can bind the hypoxia-responsive promoter element (HRPE) sequence, which in turn activates transcription of hypoxia-responsive genes (HRGs). On the other hand, under normal oxygen conditions, ERFs are subjected to enzymatic degradation via the N-end rule pathway. Enzymes are shown in green. MAP, methionine aminopeptidase; PCO, plant cysteine oxidase; ATE, arginyl-tRNA transferase; PRT6-E3, ubiquitin ligase proteolysis; MC-ERF, amino acid sequence of the protein; oxC-ERF, oxidation of the cysteine residue; Ub-ERF, ubiquitination of ERF. Although available literature indicates ERF degradation via the N-end rule pathway, genes coding the enzymes involved in the N-end rule pathway of protein degradation have yet to be identified. Therefore, this is an illustration of ERF oxygen signaling based on knowledge obtained in Arabidopsis and apple fruit (adapted from Licausi et al. [64]).


{kind=link}
{kind=link}
{kind=link}
| Fruit | Porosity (%) | Respiration |
|---|---|---|
| Apple (Granny Smith) | 23.8 | Climacteric |
| Strawberry (Chandler) | 6.3 | Nonclimacteric |
| Peach (Miraflores) | 2.6 | Climacteric |
| Mango (Tommy Atkins) | 9.9 | Climacteric |
| Nectarine (Sunglo) | 4.0 | Climacteric |
| Orange (Thomson) | 48.5 | Nonclimacteric |
| Pear (Hosui) | 1.7 | Climacteric |
Table 2.
Gene nomenclature of ethylene response factor (ERF) subgroup VII proteins in Arabidopsis, tomato, apple, and grapevine. Arabidopsis and tomato have five genes in the ERF-VII subfamily, apple has eight, and grapevine has three.
Table 2.
Gene nomenclature of ethylene response factor (ERF) subgroup VII proteins in Arabidopsis, tomato, apple, and grapevine. Arabidopsis and tomato have five genes in the ERF-VII subfamily, apple has eight, and grapevine has three.
| Species | Gene ID | Symbol |
|---|---|---|
| Solanum lycopersicum | Solyc09g075420 | ERF2b (ERF.E.1) * |
| Solyc06g063070 | JERF1 (ERF.E.2) | |
| Solyc03g123500 | JERF3 (ERF.E.3) | |
| Solyc01g65980 | ERF 6 (ERF.E.4) | |
| Soyc12g049560 | ERF3 (ERF.E.5) | |
| Arabidopsis thaliana | AT1G53910.1 | RAP2.12 |
| AT1G72360.2 | HRE1 | |
| AT2G47520.1 | HRE2 | |
| AT3G14230.1 | RAP2.2 | |
| AT3G16770.1 | RAP2.3 | |
| Malus x domestica | MDP0000308922 | Mdo006699 |
| MDP0000403580 | Mdo006712 | |
| MDP0000679280 | - | |
| MDP0000566690 | - | |
| MDP0000413387 | Mdo002990 | |
| MDP0000128979 | Mdo003328 | |
| MDP0000848905 | Mdo002404 | |
| MDP0000288465 | Mdo002401 | |
| Vitis vinifera | VIT_07s0005g00820 | VvERF057 |
| VIT_05s077g01860 | VvERF058 | |
| VIT_09s0002g00470 | VvERF059 |
* Gene nomenclature proposed by Pirrello et al. (2012) [74].
© 2018 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Cukrov, D. Progress toward Understanding the Molecular Basis of Fruit Response to Hypoxia. Plants 2018, 7, 78. https://doi.org/10.3390/plants7040078
AMA Style
Cukrov D. Progress toward Understanding the Molecular Basis of Fruit Response to Hypoxia. Plants. 2018; 7(4):78. https://doi.org/10.3390/plants7040078
Chicago/Turabian StyleCukrov, Dubravka. 2018. "Progress toward Understanding the Molecular Basis of Fruit Response to Hypoxia" Plants 7, no. 4: 78. https://doi.org/10.3390/plants7040078
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.