Which Traits Make Weeds More Successful in Maize Crops? Insights from a Three-Decade Monitoring in France


Abstract
1. Introduction
2. Results
2.1. Changes in Weed Species Status
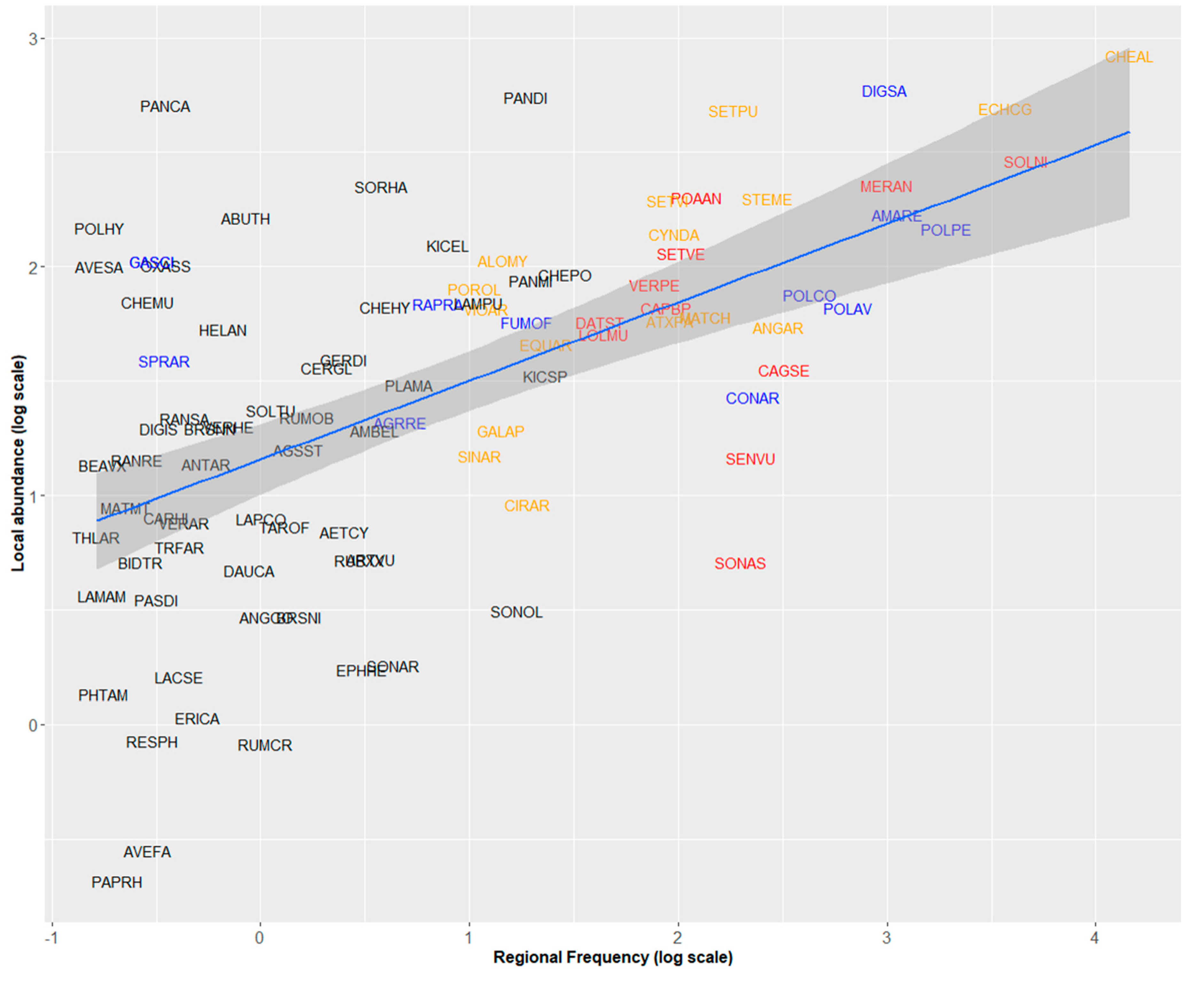
2.2. Regional Frequency, Local Abundance, and Specificity to Maize Crops
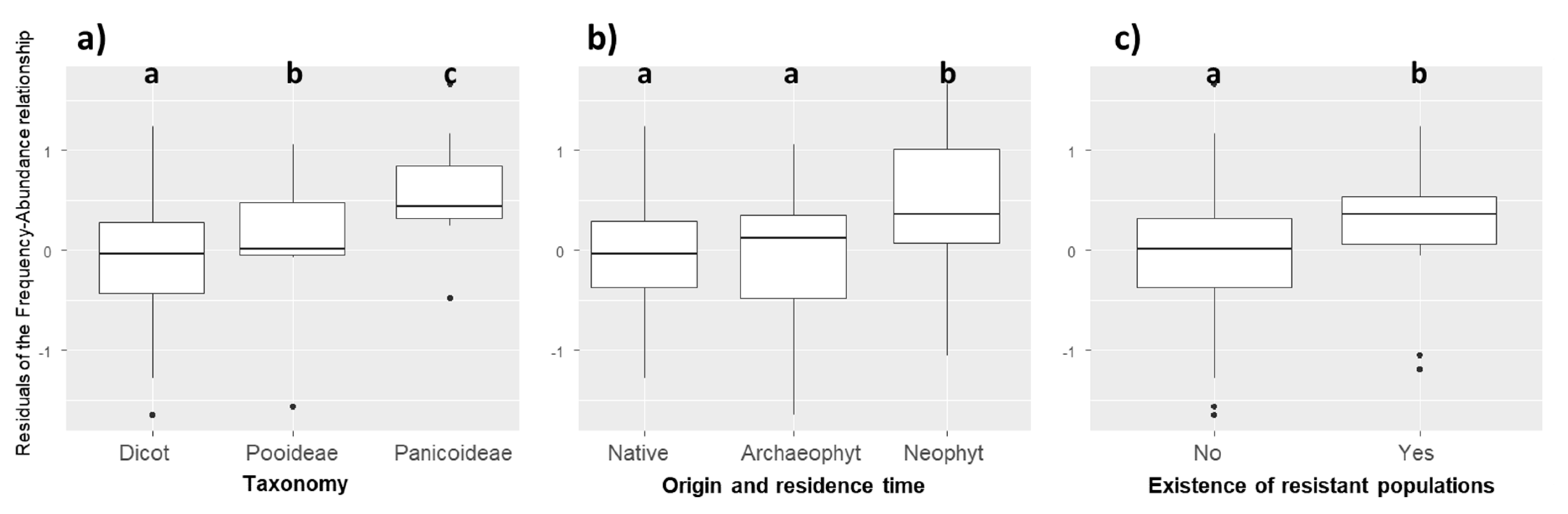
2.3. Traits Related to Success in Maize Crops
2.4. Trait Syndromes Associated with Success in Maize
3. Discussion
3.1. Trends Since the 2000s
3.2. Weediness Traits
3.3. Filtering of Crop Mimicking Traits
3.4. Significance of Frequency–Abundance Relationships for Weed Science
4. Materials and Methods
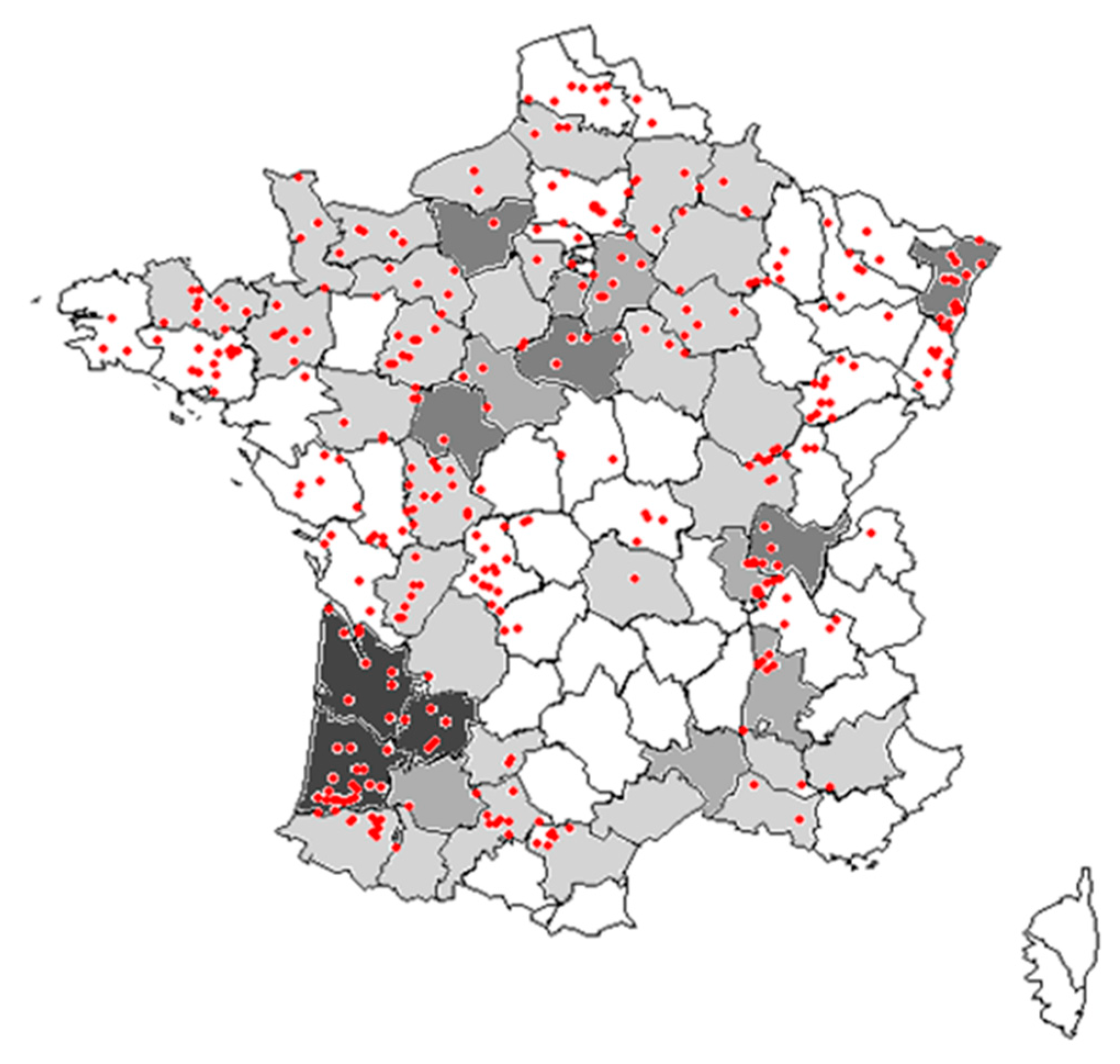
4.1. Weed Surveys
4.2. Measures of Regional Frequency, Mean Local Abundance, Specialization, and Status Changes
4.3. Weed Traits and Phylogeny
4.4. Data Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chenopodium album L., 1753 |
| Solanum nigrum L., 1753 |
| Echinochloa crus-galli (L.) P.Beauv., 1812 |
| Persicaria maculosa Gray, 1821 + Persicaria lapathifolia (L.) Delarbre, 1800 |
| Amaranthus retroflexus L., 1753 |
| Mercurialis annua L., 1753 |
| Digitaria sanguinalis (L.) Scop., 1771 |
| Polygonum aviculare L., 1753 |
| Fallopia convolvulus (L.) Á.Löve, 1970 |
| Calystegia sepium (L.) R.Br., 1810 |
| Lysimachia arvensis (L.) U.Manns & Anderb., 2009 |
| Stellaria media (L.) Vill., 1789 |
| Convolvulus arvensis L., 1753 |
| Senecio vulgaris L., 1753 |
| Sonchus asper (L.) Hill, 1769 |
| Setaria pumila (Poir.) Roem. & Schult., 1817 |
| Matricaria chamomilla L., 1753 + Tripleurospermum inodorum (L.) Sch.Bip., 1844 |
| Poa annua L., 1753 |
| Setaria verticillata (L.) P.Beauv., 1812 |
| Cynodon dactylon (L.) Pers., 1805 |
| Atriplex patula L., 1753 |
| Setaria italica subsp. viridis (L.) Thell., 1912 |
| Capsella bursa-pastoris (L.) Medik., 1792 |
| Veronica persica Poir., 1808 |
| Lolium multiflorum Lam., 1779 |
| Datura stramonium L., 1753 |
| Lipandra polysperma (L.) S.Fuentes, Uotila & Borsch, 2012 |
| Equisetum arvense L., 1753 |
| Kickxia spuria (L.) Dumort., 1827 |
| Panicum miliaceum L., 1753 |
| Cirsium arvense (L.) Scop., 1772 |
| Fumaria officinalis L., 1753 |
| Panicum dichotomiflorum Michx., 1803 |
| Sonchus oleraceus L., 1753 |
| Alopecurus myosuroides Huds., 1762 |
| Galium aparine L., 1753 |
| Viola arvensis Murray, 1770 |
| Sinapis arvensis L., 1753 |
| Lamium purpureum L., 1753 |
| Portulaca oleracea L., 1753 |
| Kickxia elatine (L.) Dumort., 1827 |
| Raphanus raphanistrum L., 1753 |
| Plantago major L., 1753 |
| Elytrigia repens (L.) Desv. ex Nevski, 1934 |
| Sonchus arvensis L., 1753 |
| Chenopodiastrum hybridum (L.) S.Fuentes, Uotila & Borsch, 2012 |
| Sorghum halepense (L.) Pers., 1805 |
| Ambrosia artemisiifolia L., 1753 |
| Artemisia vulgaris L., 1753 |
| Euphorbia helioscopia L., 1753 |
| Rubus fruticosus L., 1753 |
| Aethusa cynapium L., 1753 |
| Geranium dissectum L., 1755 |
| Cerastium glomeratum Thuill., 1799 |
| Rumex obtusifolius L., 1753 |
| Brassica nigra (L.) W.D.J.Koch, 1833 |
| Agrostis stolonifera L., 1753 |
| Taraxacum officinale F.H.Wigg., 1780 |
| Solanum tuberosum L., 1753 |
| Lysimachia foemina (Mill.) U.Manns & Anderb., 2009 |
| Rumex crispus L., 1753 |
| Lapsana communis L., 1753 |
| Daucus carota L., 1753 |
| Abutilon theophrasti Medik., 1787 |
| Veronica hederifolia L., 1753 |
| Helianthus annuus L., 1753 |
| Brassica napus L., 1753 |
| Anthemis arvensis L., 1753 |
| Erigeron canadensis L., 1753 |
| Ranunculus sardous Crantz, 1763 |
| Veronica arvensis L., 1753 |
| Trifolium arvense L., 1753 |
| Lactuca serriola L., 1756 |
| Cardamine hirsuta L., 1753 |
| Panicum capillare L., 1753 |
| Oxalis fontana Bunge, 1835 |
| Spergula arvensis L., 1753 |
| Digitaria ischaemum (Schreb.) Mühl., 1817 |
| Paspalum dilatatum Poir., 1804 |
| Reseda phyteuma L., 1753 |
| Galinsoga quadriradiata Ruiz & Pav., 1798 + Galinsoga parviflora Cav., 1795 |
| Avena fatua L., 1753 |
| Chenopodiastrum murale (L.) S.Fuentes, Uotila & Borsch, 2012 |
| Bidens tripartita L., 1753 |
| Ranunculus repens L., 1753 |
| Matricaria discoidea DC., 1838 |
| Papaver rhoeas L., 1753 |
| Phytolacca americana L., 1753 |
| Beta vulgaris L., 1753 |
| Lamium amplexicaule L., 1753 |
| Avena sativa L., 1753 |
| Persicaria hydropiper (L.) Spach, 1841 |
| Thlaspi arvense L., 1753 |
| Cyanus segetum Hill, 1762 |
| Arrhenatherum elatius subsp. bulbosum (Willd.) Schübl. & G.Martens, 1834 |
| Poa trivialis L., 1753 |
| Glebionis segetum (L.) Fourr., 1869 |
| Picris hieracioides L., 1753 |
| Sherardia arvensis L., 1753 |
| Ranunculus arvensis L., 1753 |
| Malva sylvestris L., 1753 |
| Chaenorrhinum minus (L.) Lange, 1870 |
| Trifolium repens L., 1753 |
| Plantago lanceolata L., 1753 |
| Ammi majus L., 1753 |
| Potentilla reptans L., 1753 |
| Hordeum vulgare L., 1753 |
| Trifolium pratense L., 1753 |
| Sicyos angulata L., 1753 |
| Geranium rotundifolium L., 1753 |
| Reseda luteola L., 1753 |
| Stachys arvensis (L.) L., 1763 |
| Juncus bufonius L., 1753 |
| Heliotropium europaeum L., 1753 |
| Phragmites australis (Cav.) Trin. ex Steud., 1840 |
| Euphorbia segetalis L., 1753 |
| Bromus catharticus Vahl, 1791 |
| Lepidium squamatum Forssk., 1775 |
| Aphanes arvensis L., 1753 |
| Symphytum officinale L., 1753 |
| Urtica urens L., 1753 |
| Carex hirta L., 1753 |
| Triticum aestivum L., 1753 |
| Helminthotheca echioides (L.) Holub, 1973 |
| Anthemis cotula L., 1753 |
| Rumex acetosa L., 1753 |
| Verbena officinalis L., 1753 |
| Myosotis arvensis (L.) Hill, 1764 |
| Erodium cicutarium (L.) L’Hér., 1789 |
| Montia fontana L., 1753 |
| Lotus corniculatus L., 1753 |
| Rumex acetosella L., 1753 |
| Cyperus esculentus L., 1753 |
| Euphorbia peplus L., 1753 |
| Phalaris paradoxa L., 1763 |
| Crepis foetida L., 1753 |
| Stachys annua (L.) L., 1763 |
| Galeopsis tetrahit L., 1753 |
| Sorghum bicolor (L.) Moench, 1794 |
| Vicia hirsuta (L.) Gray, 1821 |
| Silybum marianum (L.) Gaertn., 1791 |
| Sisymbrium officinale (L.) Scop., 1772 |
| Agrostis capillaris L., 1753 |
| Medicago lupulina L., 1753 |
| Teesdalia nudicaulis (L.) R.Br., 1812 |
| Oxalis corniculata L., 1753 |
| Scandix pecten-veneris L., 1753 |
| Crepis sancta (L.) Bornm., 1913 |
| Vicia sativa L., 1753 |
| Caucalis platycarpos L., 1753 |
| Hypericum perforatum L., 1753 |
| Lythrum hyssopifolia L., 1753 |
| Gnaphalium uliginosum L., 1753 |
| Apera spica-venti (L.) P.Beauv., 1812 |
| Euphorbia exigua L., 1753 |
| Geranium molle L., 1753 |
| Silene latifolia subsp. alba (Mill.) Greuter & Burdet, 1982 |
| Equisetum telmateia Ehrh., 1783 |
| Torilis arvensis (Huds.) Link, 1821 |
| Geranium robertianum L., 1753 |
| Arenaria serpyllifolia L., 1753 |
| Geranium columbinum L., 1753 |
| x Triticosecale rimpaui (M.Graebn.) Wittm. ex A.W.Hill, 1933 |
| Lycopsis arvensis L., 1753 |
| Dactylis glomerata L., 1753 |
| Misopates orontium (L.) Raf., 1840 |
| Salix L., 1753 |
| Trifolium campestre Schreb., 1804 |
| Arabidopsis thaliana (L.) Heynh., 1842 |
| Holcus mollis L., 1759 |
| Ornithopus compressus L., 1753 |
| Raphanus sativus L., 1753 |
| Coincya monensis subsp. cheiranthos (Vill.) Aedo, Leadlay & Muñoz Garm., 1993 |
| Fagopyrum esculentum Moench, 1794 |
| Veronica serpyllifolia L., 1753 |
| Sambucus nigra L., 1753 |
| Erigeron sumatrensis Retz., 1810 |
Appendix B

References
- Van Kleunen, M.; Weber, E.; Fischer, M. A meta-analysis of trait differences between invasive and non-invasive plant species. Ecol. Lett. 2009, 13, 235–245. [Google Scholar] [CrossRef]
- Daehler, C.C. The taxonomic distribution of invasive angiosperm plants: Ecological insights and comparison to agricultural weeds. Biol. Conserv. 1998, 84, 167–180. [Google Scholar] [CrossRef]
- Baker, H.G. Characteristics and modes of origin of weeds. In Genetics of Colonizing Species; Zohary, D., Ed.; Academic Press: New York, NY, USA, 1965; pp. 147–172. [Google Scholar]
- Baker, H.G. The evolution of weeds. Annu. Rev. Ecol. Syst. 1974, 5, 1–24. [Google Scholar] [CrossRef]
- Lososová, Z.; Chytry, M.; Kuhn, I. Plant attributes determining the regional abundance of weeds on central European arable land. J. Biogeogr. 2008, 35, 177–187. [Google Scholar] [CrossRef]
- Fried, G.; Chauvel, B.; Reboud, X. A functional analysis of large-scale temporal shifts from 1970 to 2000 in weed assemblages of sunflower crops in France. J. Veg. Sci. 2009, 20, 49–58. [Google Scholar] [CrossRef]
- Kuester, A.; Conner, J.K.; Culley, T.; Baucom, R.S. How weeds emerge: A taxonomic and trait-based examination using United States data. New Phytol. 2014, 202, 1055–1068. [Google Scholar] [CrossRef]
- Gaba, S.; Fried, G.; Kazakou, E.; Chauvel, B.; Navas, M.L. Agroecological weed control using a functional approach: A review of cropping systems diversity. Agron. Sustain. Dev. 2014, 34, 103–119. [Google Scholar] [CrossRef]
- Rabinowitz, D. Seven forms of rarity. In The Biological Aspects of Rare Plant Conservation; Synge, H., Ed.; John Wiley: Chichester, UK, 1981; pp. 205–217. [Google Scholar]
- Mahaut, L.; Fried, G.; Gaba, S. Patch dynamics and temporal dispersal partly shape annual plant communities in ephemeral habitat patches. Oikos 2018, 127, 147–159. [Google Scholar] [CrossRef]
- Metcalfe, H.; Hassall, K.L.; Boinot, S.; Storkey, J. The contribution of spatial mass effects to plant diversity in arable fields. J. Appl. Ecol. 2019, 56, 1560–1574. [Google Scholar] [CrossRef]
- Weber, M.M.; Stevens, R.D.; Diniz-Filho, J.A.F.; Grelle, C.E.V. Is there a correlation between abundance and environmental suitability derived from ecological niche modelling? A meta-analysis. Ecography 2017, 40, 817–828. [Google Scholar] [CrossRef]
- Violle, C.; Navas, M.L.; Vile, D.; Kazakou, E.; Fortunel, C.; Hummel, I.; Garnier, E. Let the concept of trait be functional! Oikos 2007, 116, 882–892. [Google Scholar] [CrossRef]
- Booth, B.D.; Swanton, C.J. Assembly theory applied to weed communities. Weed Sci. 2002, 50, 2–13. [Google Scholar] [CrossRef]
- Fried, G.; Petit, S.; Reboud, X. A specialist-generalist classification of the arable flora and its response to changes in agricultural practices. BMC Ecol. 2010, 10, 20. [Google Scholar] [CrossRef] [PubMed]
- Julliard, R.; Clavel, J.; Devictor, V.; Jiguet, F.; Couvet, D. Spatial segregation of specialists and generalists in bird communities. Ecol. Lett. 2006, 9, 1237–1244. [Google Scholar] [CrossRef]
- Fried, G.; Chauvel, B.; Reboud, X. Weed flora shifts and specialisation in winter oilseed rape in France. Weed Res. 2015, 55, 514–524. [Google Scholar] [CrossRef]
- Johnson, N.C.; Rowland, D.L.; Corkidi, L.; Allen, E.B. Plant winners and losers during grassland n-eutrophication differ in biomass allocation and mycorrhizas. Ecology 2008, 89, 2868–2878. [Google Scholar] [CrossRef] [PubMed]
- Wiegmann, S.M.; Waller, D.M. Fifty years of change in northern upland forest understories: Identity and traits of “winner” and “loser” plant species. Biol. Conserv. 2006, 129, 109–123. [Google Scholar] [CrossRef]
- Von Redwitz, C.; Gerowitt, B. Maize-dominated crop sequences in northern Germany: Reaction of the weed species communities. Appl. Veg. Sci. 2018, 21, 431–441. [Google Scholar] [CrossRef]
- Grime, J.P. Vegetation classification by reference to strategies. Nature 1974, 250, 26–31. [Google Scholar] [CrossRef]
- Petit, S.; Gaba, S.; Grison, A.-L.; Meiss, H.; Simmoneau, B.; Munier-Jolain, N.; Bretagnolle, V. Landscape scale management affects weed richness but not weed abundance in winter wheat fields. Agric. Ecosyst. Environ. 2016, 223, 41–47. [Google Scholar] [CrossRef]
- Juárez-Escario, A.; Solé-Senan, X.; Recasens, J.; Taberner, A.; Conesa, J. Long-term compositional and functional changes in alien and native weed communities in annual and perennial irrigated crops. Ann. Appl. Biol. 2018, 173, 42–54. [Google Scholar] [CrossRef]
- Fanfarillo, E.; Kasperski, A.; Giuliani, A.; Abbate, G. Shifts of arable plant communities after agricultural intensification: A floristic and ecological diachronic analysis in maize fields of Latium (central Italy). Bot. Lett. 2019, 166, 356–365. [Google Scholar] [CrossRef]
- Fagúndez, J.; Olea, P.P.; Tejedo, P.; Mateo-Tomás, P.; Gómez, D. Irrigation and maize cultivation erode plant diversity within crops in Mediterranean dry cereal agro-ecosystems. Environ. Manag. 2016, 58, 164–174. [Google Scholar] [CrossRef] [PubMed]
- Renoux, J.; Bibard, V.; Gautier, X.; Hebrard, J. Maïs: Réussir l’après atrazine. Perspect. Agric. 2003, 286, 27–50. [Google Scholar]
- Perry, J.; Firbank, L.; Champion, G.; Clark, S.; Heard, M.; May, M.; Hawes, C.; Squire, G.; Rothery, P.; Woiwod, I. Ban on triazine herbicides likely to reduce but not negate relative benefits of GMHT maize cropping. Nature 2004, 428, 313. [Google Scholar] [CrossRef]
- Bourgeois, B.; Munoz, F.; Fried, G.; Mahaut, L.; Armengot, L.; Denelle, P.; Storkey, J.; Gaba, S.; Violle, C. What makes a weed a weed? A large-scale evaluation of arable weeds through a functional lens. Am. J. Bot. 2019, 106, 90–100. [Google Scholar] [CrossRef]
- Storkey, J.; Moss, S.R.; Cussans, J.W. Using Assembly Theory to Explain Changes in a Weed Flora in Response to Agricultural Intensification. Weed Sci. 2010, 58, 39–46. [Google Scholar] [CrossRef]
- McElroy, J.S. Vavilovian Mimicry: Nikolai Vavilov and His Little-Known Impact on Weed Science. Weed Sci. 2014, 62, 207–216. [Google Scholar] [CrossRef]
- Barrett, S. Crop mimicry in weeds. Econ. Bot. 1983, 37, 255–282. [Google Scholar] [CrossRef]
- Fried, G.; Kazakou, E.; Gaba, S. Trajectories of weed communities explained by traits associated with species’ response to management practices. Agric. Ecosyst. Environ. 2012, 158, 147–155. [Google Scholar] [CrossRef]
- Perronne, R.; Le Corre, V.; Bretagnolle, V.; Gaba, S. Stochastic processes and crop types shape weed community assembly in arable fields. J. Veg. Sci. 2015, 26, 348–359. [Google Scholar] [CrossRef]
- Gaston, K.J.; Blackburn, T.M.; Greenwood, J.J.; Gregory, R.D.; Quinn, R.M.; Lawton, J.H. Abundance–occupancy relationships. J. Appl. Ecol. 2000, 37, 39–59. [Google Scholar] [CrossRef]
- Barralis, G. Répartition et Densité des Principales Mauvaises Herbes en France; Institut National de la Recherche Agronomique, Laboratoire de Malherbologie (INRA-AFPP): Paris, France, 1977; p. 2. [Google Scholar]
- Fried, G.; Norton, L.R.; Reboud, X. Environmental and management factors determining weed species composition and diversity in France. Agric. Ecosyst. Environ. 2008, 128, 68–76. [Google Scholar] [CrossRef]
- Dufrêne, M.; Legendre, P. Species assemblages and indicator species: The need for a flexible asymmetrical approach. Ecol. Monogr. 1997, 67, 345–366. [Google Scholar] [CrossRef]
- Westoby, M. A leaf-height-seed (LHS) plant ecology strategy scheme. Plant Soil 1998, 199, 213–227. [Google Scholar] [CrossRef]
- Storkey, J. Modelling seedling growth rates of 18 temperate arable weed species as a function of the environment and plant traits. Ann. Bot. 2004, 93, 681–689. [Google Scholar] [CrossRef]
- Rao, V.S. Principles of Weed Science, 2nd ed.; Taylor & Francis: London, UK, 2000. [Google Scholar]
- Moles, A.T.; Westoby, M. Seed size and plant strategy across the whole life cycle. Oikos 2006, 113, 91–105. [Google Scholar] [CrossRef]
- Fried, G.; Maillet, J. Diversité et réponses de la flore des champs cultivés à l’évolution des pratiques agricoles en France. In Gestion Durable de la Flore Adventice des Cultures; Chauvel, B., Darmency, H., Munier-Jolain, N., Rodriguez, A., Eds.; Éditions Quæ: Versailles, France, 2018; pp. 39–57. [Google Scholar]
- Noble, I.R.; Gitay, H. A functional classification for predicting the dynamics of landscapes. J. Veg. Sci. 1996, 7, 329–336. [Google Scholar] [CrossRef]
- Gunton, R.M.; Petit, S.; Gaba, S. Functional traits relating arable weed communities to crop characteristics. J. Veg. Sci. 2011, 22, 541–550. [Google Scholar] [CrossRef]
- Zanin, G.; Otto, S.; Riello, L.; Borin, M. Ecological interpretation of weed flora dynamics under different tillage systems. Agric. Ecosyst. Environ. 1997, 66, 177–188. [Google Scholar] [CrossRef]
- Ellenberg, H.; Weber, H.; Düll, R.; Wirth, V.; Werner, W.; Paulissen, D. Indicator values of central European plants. Scr. Geobot. 1992, 18, 1–258. [Google Scholar]
- Haas, H.; Streibig, J.C. Changing Patterns of Weed Distribution as a Result of Herbicide Use and Other Agronomic Factors. In Herbicide Resistance in Plants; LeBaron, H.M., Gressel, J., Eds.; John Wiley & Sons: New York, NY, USA, 1982; pp. 57–79. [Google Scholar]
- Martin, G.; Devictor, V.; Motard, E.; Machon, N.; Porcher, E. Short-term climate-induced change in French plant communities. Biol. Lett. 2019, 15, 20190280. [Google Scholar] [CrossRef] [PubMed]
- Mamarot, J.; Rodriguez, A. Sensibilité des Mauvaises Herbes Aux Herbicides en Grandes Cultures; ACTA: Paris, France, 2003; p. 372. [Google Scholar]
- Kleyer, M.; Bekker, R.M.; Knevel, I.C.; Bakker, J.P.; Thompson, K.; Sonnenschein, M.; Poschlod, P.; Van Groenendael, J.M.; Klimes, L.; Klimesová, J.; et al. The LEDA Traitbase: A database of life-history traits of Northwest European flora. J. Ecol. 2008, 96, 1266–1274. [Google Scholar] [CrossRef]
- Tison, J.-M.; De Foucault, B. Flora Gallica, Flore de France; Biotope Éditions: Mèze, France, 2014; p. 1195. [Google Scholar]
- Kew, R.B.G. Seed Information Database (SID). Available online: http://data.kew.org/sid/ (accessed on 21 March 2019).
- Gaba, S.; Biju-Duval, L.; Strbik, F.; Gaujour, E.; Bretagnolle, F.; Coffin, A.; Cordeau, S.; Dessaint, F.; Fried, G.; Gard, B. Weed-DATA Base de données ‘Traits’ des plantes adventices des agroécosystèmes; Inra: Dijon, France, 2014. [Google Scholar]
- Julve, P. Baseflor. Index Botanique, Écologique et Chorologique de la Flore de France, 4 mars 2012 ed. 1998. Available online: http://philippe.julve.pagesperso-orange.fr/catminat.htm (accessed on 21 March 2019).
- Mamarot, J. Mauvaises Herbes des Cultures; ACTA Editions: Paris, France, 2002; pp. 1–540. [Google Scholar]
- Darmency, H.; Gasquez, J. Résistances aux herbicides chez les mauvaises herbes. Agronomie 1990, 10, 457–472. [Google Scholar] [CrossRef]
- Jauzein, P.; Nawrot, O. Flore d’Île-de-France; Quae: Toulouse, France, 2011; p. 970. [Google Scholar]
- Orme, D.; Freckleton, R.; Thomas, G.; Petzoldt, T.; Fritz, S.; Isaac, N.; Pearse, W. Caper: Comparative Analyses of Phylogenetics and Evolution in R. R Package Version 0.5. 2012, p. 458. Available online: https://cran.r-project.org/web/packages/caper/caper.pdf (accessed on 21 March 2019).
- Harvey, P.H.; Pagel, M.D. The Comparative Method in Evolutionary Biology; citeulike-article-id:768613; Oxford Seies in Ecology and Evolution; Oxford University Press: Oxford, UK, 1991. [Google Scholar]
- Qian, H.; Jin, Y. An updated megaphylogeny of plants, a tool for generating plant phylogenies and an analysis of phylogenetic community structure. J. Plant Ecol. 2016, 9, 233–239. [Google Scholar] [CrossRef]




| Rank | Names 1 | Regional Frequency (%) | Local Abundance (ind./m2) | Trend in the 2000s | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 2000s 2 | 1970s | Status 3 | 2000s 2 | 1970s | Status 3 | Spearman rho | p-Value | Status 3 | ||
| 1 | Chenopodium album | 64.3 [57.7–70.3] | 60.3 | = | 13.9 [11.2–16.6] | 12.4 | = | −0.250 | 0.594 | = |
| 2 | Solanum nigrum | 39.1 [33.1–45.1] | 26.5 | + | 7.5 [5.5–9.6] | 6.9 | = | −0.143 | 0.783 | = |
| 3 | Echinochloa crus-galli | 35.5 [29.7–41.1] | 38.0 | = | 8.4 [6.2–10.6] | 12.9 | − | 0.357 | 0.444 | = |
| 4 | Persicaria maculata + P. lapathifolia | 26.7 [21.1–32.0] | 35.1 | − | 4.9 [3.4–6.6] | 3.1 | + | 0.786 | 0.048 | + |
| 5 | Amaranthus retroflexus | 21.1 (16.0–25.7] | 26.7 | − | 3.6 [2.3–5.2] | 5.8 | − | 0.071 | 0.906 | = |
| 6 | Mercurialis annua | 20.1 [15.5–25.1] | 15.4 | + | 3.3 [2.1–4.7] | 3.9 | = | 0.286 | 0.556 | = |
| 7 | Digitaria sanguinalis | 19.8 [15.4–24.6] | 40.2 | − | 5.2 [3.4–7.1] | 14.5 | − | 0.214 | 0.662 | = |
| 8 | Polygonum aviculare | 16.6 [12.6–21.1] | 26.4 | − | 2.1 [1.4–3.1] | 7.5 | − | −0.357 | 0.444 | = |
| 9 | Fallopia convolvulus | 13.9 [9.7–18.3] | 21.3 | − | 1.8 [1.0–2.6] | 4.1 | − | 0.536 | 0.236 | = |
| 10 | Calystegia sepium | 12.3 [8.0–16.6] | <2.3 | N | 1.7 [1.0–2.6] | ? | N | 0.750 | 0.066 | (+) |
| 11 | Lysimachia arvensis | 12.0 [8.0–16.0] | 13.2 | = | 1.1 [0.6–1.9] | 2.9 | − | 0.571 | 0.200 | = |
| 12 | Stellaria media | 11.3 [7.4–15.4] | 14.3 | = | 2.6 [1.4–4.0] | 3.6 | = | −0.928 | 0.007 | − |
| 13 | Convolvulus arvensis | 10.6 [6.9–14.3] | 15.5 | − | 1.4 [0.8–2.2] | 2.8 | − | 0.143 | 0.783 | = |
| 14 | Senecio vulgaris | 10.5 [6.9–14.3] | <2.3 | N | 1.2 [0.9–1.7] | ? | N | 0.714 | 0.088 | (+) |
| 15 | Sonchus asper | 10.0 [6.8–13.7] | <2.3 | N | 1.2 [0.8–1.7] | ? | N | 0.893 | 0.012 | + |
| 16 | Setaria pumila | 9.6 [6.3–16.1] | 9.8 | = | 2.9 [1.4–4.5] | 3.6 | = | 0.464 | 0.302 | = |
| 17 | Matricaria chamomilla + Tripleurospermum inodorum | 8.4 [5.1–12.0] | 7.9 | = | 1.3 [0.7–2.1] | 2.3 | − | 0.607 | 0.167 | = |
| 18 | Poa annua | 8.1 [4.6–11.4] | <2.3 | N | 1.4 [0.6–2.5] | ? | N | 0.857 | 0.024 | + |
| 19 | Setaria verticillata | 7.5 [4.0–10.9] | <2.3 | N | 0.9 [0.4–1.6] | ? | ? | 0.750 | 0.066 | (+) |
| 20 | Cynodon dactylon | 7.2 [4.0–10.3] | 5.2 | = | 1.5 [0.6–2.6] | 1.3 | = | 0.857 | 0.024 | + |
| 21 | Atriplex patula | 7.1 [4.0–10.3] | 10.3 | = | 1.0 [0.4–1.6] | 2.8 | − | 0.429 | 0.353 | = |
| 22 | Setaria viridis | 7.1 [4.0–10.3] | 10.3 | = | 1.3 [0.5–2.3] | 4.2 | − | 0.286 | 0.556 | = |
| 23 | Capsella bursa-pastoris | 7.0 [4.0–10.3] | <2.3 | N | 1.0 [0.5–1.8] | ? | N | −0.643 | 0.139 | = |
| 24 | Veronica persica | 6.6 [3.4–9.7] | <2.3 | N | 1.0 [0.4–1.8] | ? | ? | 0.250 | 0.595 | = |
| 25 | Lolium multiflorum | 5.2 [2.3–8.0] | <2.3 | N | 0.7 [0.3–1.2] | ? | ? | −0.464 | 0.302 | = |
| 26 | Datura stramonium | 5.1 [2.3–8.0] | <2.3 | N | 1.1 [0.4–1.9] | ? | ? | −0.036 | 0.964 | = |
| 27 | Lipandra polyspermum | 4.3 [1.7–7.4] | <2.3 | ? | 0.5 [0.2–1.0] | ? | ? | −0.071 | 0.906 | = |
| 28 | Equisetum arvense | 3.9 [1.7–6.3] | 6.2 | = | 0.5 [0.2–1.1] | 1.9 | − | 0.071 | 0.906 | = |
| 29 | Kickxia spuria | 3.9 [1.7–6.3] | <2.3 | ? | 0.4 [0.2–0.7] | ? | ? | 0.607 | 0.167 | = |
| 30 | Panicum miliaceum | 3.7 [1.7–6.3] | <2.3 | ? | 0.7 [0.2–1.3] | ? | ? | −0.179 | 0.713 | = |
| 31 | Cirsium arvense | 3.6 [1.7–6.3] | 6.3 | = | 0.5 [0.3–0.9] | 1.6 | − | −0.214 | 0.662 | = |
| 32 | Fumaria officinalis | 3.6 [1.7–6.3] | 6.4 | − | 0.8 [0.3–1.4] | 1.4 | = | 0.536 | 0.236 | = |
| 33 | Panicum dichotomiflorum | 3.6 [1.1–6.3] | <2.3 | ? | 0.8 [0.2–1.6] | ? | ? | 0.036 | 0.966 | = |
| 34 | Sonchus oleraceus | 3.4 [1.1–5.7] | <2.3 | ? | 0.5 [0.3–0.9] | ? | ? | 0.214 | 0.662 | = |
| 35 | Alopecurus myosuroides | 3.2 [1.1–5.1] | 2.9 | = | 0.8 [0.2–1.5] | 0.8 | = | −0.821 | 0.034 | − |
| 36 | Galium aparine subsp. aparine | 3.2 [1.1–5.1] | 4.6 | = | 0.4 [0.2–0.8] | 1.1 | − | 0.500 | 0.267 | = |
| 37 | Viola arvensis | 2.9 [1.1–5.1] | 4.6 | = | 0.4 [0.1–1.0] | 1.4 | − | −0.464 | 0.302 | = |
| 38 | Sinapis arvensis | 2.9 [1.1–5.1] | 4.6 | = | 0.4 [0.2–0.8] | 2.0 | − | −0.428 | 0.354 | = |
| 39 | Lamium purpureum | 2.8 [1.1–5.1] | <2.3 | ? | 0.4 [0.1–0.9] | ? | ? | −0.929 | 0.007 | − |
| 40 | Portulacca oleracea | 2.8 [1.1–5.1] | 2.3 | = | 0.8 [0.1–1.6] | 1.8 | − | −0.429 | 0.353 | = |
| 42 | Raphanus raphanistrum | 2.3 [0.6–4.6] | 15.6 | − | 0.4 [0.1–1.1] | 3.0 | − | 0.643 | 0.139 | = |
| 44 | Elytrigia repens | 1.9 [0.6–4.0] | 4.5 | − | 0.3 [0.1–0.8] | 2.1 | − | 0.107 | 0.840 | = |
| 77 | Spergula arvensis | 0.6 [0.0–1.7] | 6.9 | − | 0.1 [0.0–0.2] | 2.9 | − | −0.211 | 0.669 | = |
| 81 | Galinsoga quadriradiata + G. parviflora | 0.6 [0.0–1.7] | 2.3 | − | 0.1 [0.0–0.3] | 0.4 | − | 0.556 | 0.256 | = |
| Regional Frequency | Local Abundance | Specificity | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Estimate | Std. Err. | t-Value | p-Value | Estimate | Std. Err. | t-Value | p-Value | Estimate | Std. Err. | t-Value | p-Value | |
| Hemicryptophytes | −1.247 | 0.523 | −2.384 | 0.019 * | −0.447 | 0.306 | −1.463 | 0.147 | 23.945 | 10.214 | 1.251 | 0.021 * |
| Therophytes | 0.238 | 0.412 | 0.579 | 0.564 | 0.209 | 0.240 | 0.873 | 0.385 | 0.793 | 8.000 | 0.099 | 0.921 |
| C4 | 0.760 | 0.358 | 2.124 | 0.036 * | 0.854 | 0.205 | 4.163 | <0.001 *** | 34.053 | 6.734 | 5.057 | <0.001 *** |
| Plant Height | 0.280 | 0.206 | 1.362 | 0.177 | −0.126 | 0.124 | −1.016 | 0.312 | −2.583 | 4.238 | −0.610 | 0.544 |
| Seed weight | 0.011 | 0.087 | 0.123 | 0.903 | −0.005 | 0.050 | −0.091 | 0.928 | −2.598 | 1.621 | −1.602 | 0.113 |
| SLA | 0.005 | 0.013 | 0.379 | 0.706 | 0.016 | 0.007 | 2.248 | 0.027 * | −0.083 | 0.236 | −0.351 | 0.727 |
| All-year-round | 0.851 | 0.379 | 2.242 | 0.027 * | 0.248 | 0.218 | 1.141 | 0.257 | 9.493 | 7.031 | 1.350 | 0.180 |
| Spring | 0.398 | 0.503 | 0.790 | 0.431 | 0.225 | 0.289 | 0.781 | 0.437 | 22.534 | 9.322 | 2.417 | 0.018 * |
| Spring & Summer | 1.284 | 0.393 | 3.270 | 0.002 ** | 0.654 | 0.225 | 2.903 | 0.005 ** | 26.125 | 7.277 | 3.590 | 0.001 ** |
| Summer | 1.310 | 0.417 | 3.141 | 0.002 ** | 1.097 | 0.239 | 4.581 | <0.001 | 43.553 | 7.730 | 5.634 | <0.001 *** |
| Flow. onset | 0.008 | 0.073 | 0.104 | 0.917 | −0.038 | 0.046 | −0.894 | 0.374 | 4.045 | 1.398 | 2.893 | 0.005 ** |
| Flow. duration | 0.133 | 0.055 | 2.409 | 0.018 * | 0.046 | 0.032 | 1.414 | 0.161 | −0.838 | 1.076 | −0.779 | 0.438 |
| Fecundity | 0.036 | 0.075 | 0.485 | 0.628 | 0.019 | 0.044 | 0.438 | 0.664 | 3.361 | 1.432 | 2.327 | 0.021 * |
| Seed longevity | 0.272 | 0.122 | 2.239 | 0.028 * | 0.094 | 0.075 | 1.265 | 0.209 | 2.819 | 2.479 | 1.137 | 0.258 |
| Gravity | 0.201 | 0.335 | 0.599 | 0.551 | 0.162 | 0.201 | 0.805 | 0.423 | 2.467 | 6.856 | 0.360 | 0.720 |
| Wind-dispersal | 0.366 | 0.327 | 1.117 | 0.267 | 0.148 | 0.200 | 0.743 | 0.460 | 2.008 | 6.843 | 0.294 | 0.770 |
| Ellenberg-N | 0.091 | 0.088 | 1.038 | 0.302 | 0.100 | 0.051 | 1.946 | 0.055 | −2.082 | 1.766 | −1.179 | 0.242 |
| Ellenberg-L | −0.109 | 0.146 | −0.749 | 0.456 | −0.013 | 0.085 | −0.150 | 0.881 | 2.268 | 2.759 | 0.822 | 0.413 |
| Ellenberg-T | 0.446 | 0.141 | 3.152 | 0.002 ** | 0.106 | 0.085 | 1.250 | 0.215 | 2.732 | 2.770 | 0.986 | 0.327 |
| Sensitivity to Maize herbicides | 0.069 | 0.119 | 0.579 | 0.564 | 0.152 | 0.067 | 1.263 | 0.126 | −3.568 | 2.302 | −1.550 | 0.125 |
| Traits | Axis1 | Axis2 | Axis3 | Axis4 | Axis5 | Axis6 |
|---|---|---|---|---|---|---|
| Geophyte | 17.30 | −4.83 | −2.16 | 32.84 | −0.10 | −0.12 |
| Hemicryptophyte | 2.86 | −23.21 | 28.20 | −5.56 | 8.52 | 7.94 |
| Therophyte | −19.00 | 29.08 | −9.69 | −5.35 | −4.25 | −3.86 |
| Plant Height | 31.37 | −19.29 | −4.90 | −0.15 | −2.67 | 4.98 |
| Seed weight | 3.67 | −25.66 | −16.47 | −11.30 | −10.27 | −0.03 |
| SLA | −23.63 | 0.29 | −2.34 | 0.40 | −22.55 | 0.83 |
| Autumn-Winter-Spring | −24.66 | 0.00 | 5.41 | 6.44 | −0.42 | −0.06 |
| All-year-round | −8.25 | −0.16 | −32.73 | −5.71 | 3.61 | 0.07 |
| Spring | 0.08 | −17.00 | 11.47 | −21.01 | 0.11 | −5.46 |
| Spring-Summer | 8.70 | −6.85 | −0.24 | 35.25 | −0.72 | 1.30 |
| Summer | 25.28 | 40.32 | 0.36 | −15.34 | −0.09 | 0.26 |
| Flowering Onset | 51.05 | 1.40 | 0.00 | 0.03 | 1.40 | 0.80 |
| Flowering Duration | −35.68 | 1.85 | 19.92 | 1.08 | −4.76 | −0.04 |
| Fecundity | 3.67 | 21.96 | 2.93 | −5.82 | 0.30 | 25.12 |
| Seed longevity | −0.10 | −6.15 | 45.85 | −1.85 | −7.04 | −3.88 |
| Animals | −1.35 | −4.02 | −0.24 | −4.98 | 2.72 | −14.50 |
| Gravity | 1.35 | −0.01 | −1.90 | −12.07 | −11.34 | −1.02 |
| Wind | −0.01 | 3.61 | 3.19 | 29.23 | 3.24 | 19.58 |
| Ellenberg-N | −0.22 | −0.15 | 0.08 | −2.19 | −48.51 | 25.78 |
| Ellenberg-L | 37.11 | 3.78 | −0.76 | 0.31 | −6.68 | −14.20 |
| Ellenberg-T | 37.42 | 3.31 | 8.43 | 8.77 | −13.62 | −10.37 |
| C3 | −26.31 | −34.62 | −1.18 | 5.31 | −0.34 | 0.01 |
| C4 | 26.31 | 34.62 | 1.18 | −5.31 | 0.34 | −0.01 |
| Sensitivity to Herbicides | −21.30 | 34.98 | −2.58 | 0.12 | −1.25 | −3.56 |
| Regional Frequency | Local Abundance | Specificity to Maize | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Est. | S. E. | t val | p val | Est. | S. E. | t val | p val | Est. | S. E. | t val | p val | |
| HS 1 | 0.056 | 0.064 | 0.880 | 0.382 | 0.012 | 0.040 | 0.303 | 0.763 | 0.241 | 0.051 | 4.706 | 0.000 |
| HS 2 | 0.178 | 0.077 | 2.317 | 0.023 | 0.145 | 0.047 | 3.080 | 0.003 | 0.102 | 0.061 | 1.670 | 0.098 |
| HS 3 | 0.165 | 0.089 | 1.847 | 0.068 | 0.027 | 0.051 | 0.527 | 0.600 | 0.297 | 0.071 | 4.183 | 0.000 |
| HS 4 | 0.172 | 0.092 | 1.872 | 0.065 | 0.000 | 0.053 | −0.005 | 0.996 | 0.001 | 0.073 | 0.010 | 0.992 |
| HS 5 | −0.288 | 0.100 | −2.880 | 0.005 | −0.216 | 0.058 | −3.722 | 0.000 | 0.017 | 0.080 | 0.212 | 0.833 |
| HS 6 | 0.060 | 0.106 | 0.567 | 0.572 | 0.022 | 0.061 | 0.362 | 0.718 | 0.037 | 0.084 | 0.438 | 0.662 |
| Traits | Units | Mean | Median (Min-Max) | Source |
| Quantitative traits | ||||
| Specific Leaf Area (SLA) | cm2/g | 28.1 | 27.4 (10.9–53.7) | [50] |
| Maximum Plant Height | cm | 101.3 | 80 (20–500) | [51] |
| Seed Weight | g | 3.1 | 0.8 (0.05–39.9) | [52] |
| Flowering Onset | month | 5.3 | 6 (1–8) | [51] |
| Flowering Duration | month | 4.9 | 4 (1–12) | [51] |
| Fecundity | average number of seeds per plant | 5972 | 4000 (30–40,000) | [53] |
| Seed Longevity | year | 33.7 | 26 (3–100) | [53] |
| Ellenberg-L | 7.1 | 7 (5–9) | [54] | |
| Ellenberg-N | 6.5 | 7 (1–9) | [54] | |
| Ellenberg-T | 6.7 | 7 (5–9) | [54] | |
| Sensitivity to Herbicides 1 | 4.0 | 4.2 (1.5–6) | [49] | |
| Qualitative traits | Modalities | N. of Species | ||
| Life Form | Geophytes | 10 | [54] | |
| Hemicryptophytes | 12 | |||
| Therophytes | 73 | |||
| Emergence Period | All-year-round | 29 | [55] | |
| Autumn, Winter & Spring | 15 | |||
| Spring | 9 | |||
| Spring & Summer | 24 | |||
| Summer | 18 | |||
| Means of Dispersal | Animal | 25 | [54] | |
| Gravity | 33 | |||
| Wind | 37 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fried, G.; Chauvel, B.; Munoz, F.; Reboud, X. Which Traits Make Weeds More Successful in Maize Crops? Insights from a Three-Decade Monitoring in France. Plants 2020, 9, 40. https://doi.org/10.3390/plants9010040
Fried G, Chauvel B, Munoz F, Reboud X. Which Traits Make Weeds More Successful in Maize Crops? Insights from a Three-Decade Monitoring in France. Plants. 2020; 9(1):40. https://doi.org/10.3390/plants9010040
Chicago/Turabian StyleFried, Guillaume, Bruno Chauvel, François Munoz, and Xavier Reboud. 2020. "Which Traits Make Weeds More Successful in Maize Crops? Insights from a Three-Decade Monitoring in France" Plants 9, no. 1: 40. https://doi.org/10.3390/plants9010040
APA StyleFried, G., Chauvel, B., Munoz, F., & Reboud, X. (2020). Which Traits Make Weeds More Successful in Maize Crops? Insights from a Three-Decade Monitoring in France. Plants, 9(1), 40. https://doi.org/10.3390/plants9010040