In Vitro and In Vivo Effect of Poplar Bud Extracts on Phytophthora infestans: A New Effective Biological Method in Potato Late Blight Control


Abstract
:1. Introduction
2. Material and Methods
2.1. P. infestans Isolates
2.2. Plant Extracts Preparation
2.3. In Agaro Test of Poplar Bud Extracts on Mycelial Growth of Different Late Blight Strains
- v/v%-11.25 mL 20% populin + 140 g pea agar + 56.25 mL sterile distillate water.
- v/v%-22.5 mL 20% populin + 140 g pea agar + 45 mL sterile distillate water.
- v/v%-33.75 mL 20% populin + 140 g pea agar + 33.75 mL sterile distillate water.
- v/v%-45 mL 20% populin + 140 g pea agar + 22.5 mL sterile distillate water.
2.4. In Planta Test of Poplar Bud Extracts on Different Late Blight Strains
- Infinito 687 SC (62.5 g/L fluopicolid + 625 g/L propamocarb) doze: 1.4 L/300 L water (1.9 mL/200 mL water) synthetic fungicide.
- Valis M (60% mancozeb + 6% valifenalat) doze: 2.5 kg/300 L water (1.7 g/200 mL water) synthetic fungicide.
- Populin 10%: 100 mL extract 20% + 100 mL water.
- Populin 5%: 50 mL extract 20% + 150 mL water.
- 0% = healthy (0 foliar damage).
- 1% = up to 10 spots per plant or up to 1 leaflet in 10 attacked.
- 10% = up to 4 leaflets in 10 affected; plants still retaining normal form.
- 25% = nearly every leaflet with lesions but plants still retaining normal form; plot may look green though every plant affected.
- 50% = every plant affected and about half of leaf area destroyed by blight; plot looks green, flecked with brown.
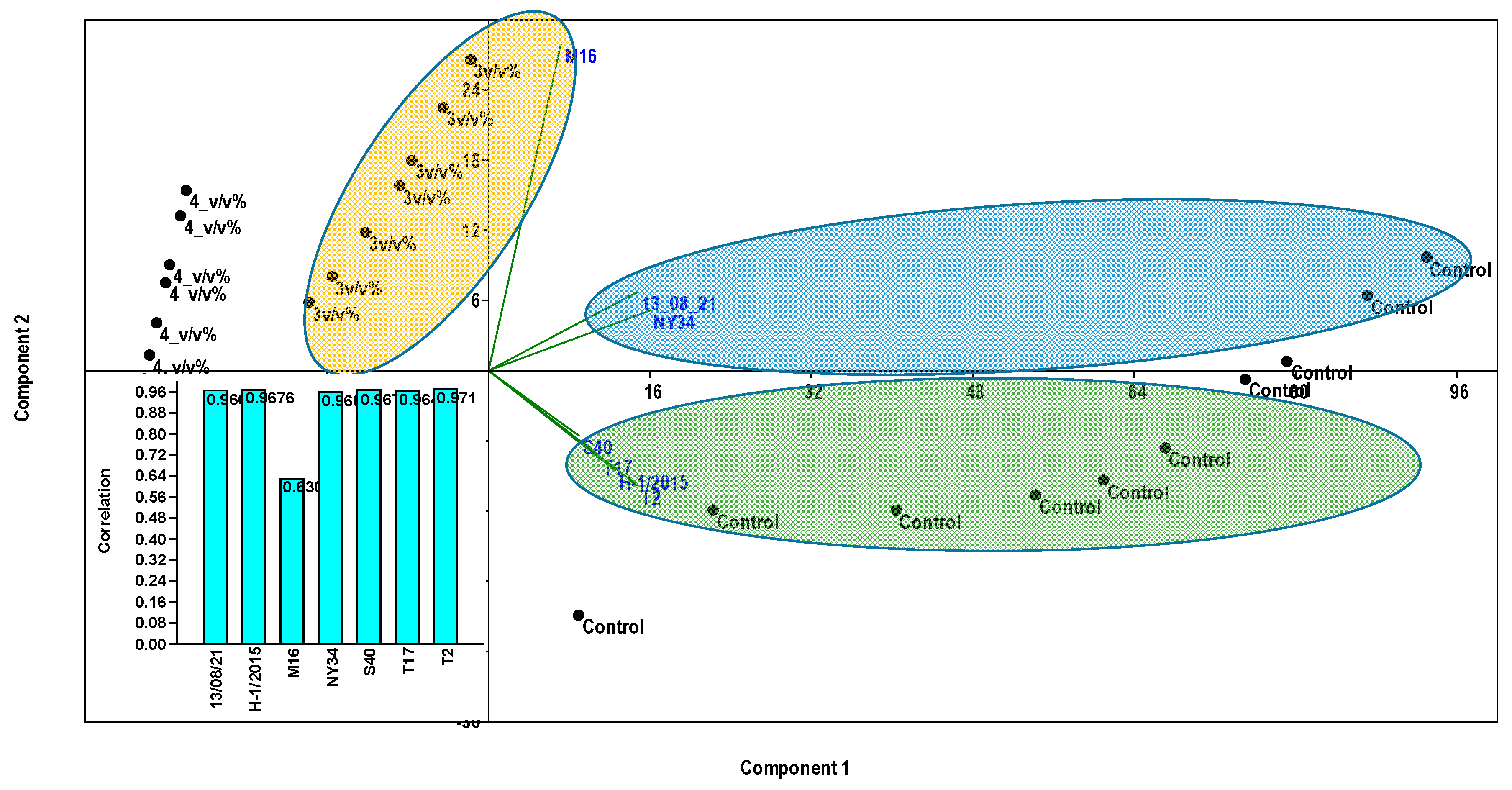
3. Data Analyses
4. Results
5. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Andrivon, D. Using Cultivar Resistance to Reduce Fungicide Inputs Against Late Blight. In From Science to Field, Potato Case Study—Guide Number 4, Endure, Food Quality and Safety 6th Framework Programme; EU Parlament: Bruxeles, Belgium, 2008; pp. 1–2. [Google Scholar]
- Schumann, G.L. The Irish potato famine and the birth of plant pathology. In Plant Diseases: Their Biology and Social Impact; American Phytopathological Society: St. Paul, MN, USA, 1991; pp. 1–24. [Google Scholar]
- Agrios, G.N. Plant Pathology, 5th ed.; Academic Press: Cambridge, MA, USA, 2005; p. 952. [Google Scholar]
- Pettitt, T.; Keane, G.; John, S.; Cooke, D.; Žerjav, M. Atypical late blight symptoms following first recorded infections by Phytophthora infestans genotype EU_39_A1 in UK vine tomatoes. New Dis. Rep. 2019, 39, 16. [Google Scholar] [CrossRef] [Green Version]
- Kiiker, R.; Skrabule, I.; Ronis, A.; Cooke, D.E.L.; Hansen, J.G.; Williams, I.H.; Mänd, M.; Runno-Paurson, E. Diversity of populations of Phytophthora infestans in relation to patterns of potato crop management in Latvia and Lithuania. Plant Pathol. 2019, 68, 1207–1214. [Google Scholar] [CrossRef]
- Davidse, L.C.; Danial, D.L.; Westen, C.J. Resistance to metalaxyl in Phytophthora infestans in the Netherlands. Eur. J. Plant Pathol. 1983, 89, 1–20. [Google Scholar] [CrossRef]
- Gisi, U.; Cohen, Y. Resistance to Phenylamide Fungicides: A Case Study with Phytophthora infestans Involving Mating Type and Race Structure. Annu. Rev. Phytopathol. 1996, 34, 549–572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, G.-N.; Huang, F.-X.; Feng, L.-X.; Qin, B.-X.; Yang, Y.-H.; Chen, Y.-H.; Lu, X.-H. Sensitivities of Phytophthora infestans to Metalaxyl, Cymoxanil, and Dimethomorph. Agric. Sci. China 2008, 7, 831–840. [Google Scholar] [CrossRef]
- Mauch-Mani, B. Host Resistance to Downy Mildew Diseases. In Advances in Downy Mildew Research; Spencer-Phillips, P.T.N., Gisi, U., Lebeda, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2002; pp. 59–83. Available online: http://link.springer.com/chapter/10.1007/0-306-47914-1_2 (accessed on 26 November 2013).
- Konstantinidou-Doltsinis, S.; Markellou, E.; Kasselaki, A.-M.; Fanouraki, M.N.; Koumaki, C.M.; Schmitt, A.; Liopa-Tsakalidis, A.; Malathrakis, N.E. Efficacy of Milsana®, a Formulated Plant Extract from Reynoutria sachalinensis, against Powdery Mildew of Tomato (Leveillula taurica). BioControl 2006, 51, 375–392. [Google Scholar] [CrossRef]
- Konstantinidou-Doltsinis, S.; Schmit, A. Impact of treatment with plant extracts from Reynoutria sachalinensis (F. Schmidt) Nakai on intensity of powdery mildew severity and yield in cucumber under high disease pressure. Crop. Prot. 1998, 17, 649–656. [Google Scholar] [CrossRef]
- Slusarenko, A.J.; Patel, A.; Portz, D. Control of plant diseases by natural products: Allicin from garlic as a case study. Eur. J. Plant Pathol. 2008, 121, 313–322. [Google Scholar] [CrossRef]
- Curtis, H.; Noll, U.; Störmann, J.; Slusarenko, A.J. Broad-spectrum activity of the volatile phytoanticipin allicin in extracts of garlic (Allium sativum L.) against plant pathogenic bacteria, fungi and Oomycetes. Physiol. Mol. Plant Pathol. 2004, 65, 79–89. [Google Scholar] [CrossRef]
- Portz, D.; Koch, E.; Slusarenko, A.J. Effects of garlic (Allium sativum) juice containing allicin on Phytophthora infestans and downy mildew of cucumber caused by Pseudoperonospora cubensis. Eur. J. Plant Pathol. 2008, 122, 197–206. [Google Scholar] [CrossRef]
- Bálint, J.; Nagy, S.; Thiesz, R.; Nyárádi, I.-I.; Balog, A. Using plant extracts to reduce asexual reproduction of apple scab (Venturia inaequalis). Turk. J. Agric. For. 2014, 38, 91–98. [Google Scholar] [CrossRef]
- Bálint, J.; Turóczi, B.; Máthé, I.; Benedek, K.; Szabó, K.-A.; Balog, A. In vitro and in vivo effect of poplar bud (Populi gemma) Extracts on late blight (Phytophthora infestans). Acta Univ. Sapientiae Agric. Environ. 2014, 6, 5–12. [Google Scholar]
- Thiesz, R.; Balog, A.; Ferencz, L.; Albert, J. The effects of plant extracts on apple scab (Venturia inaequalis Cooke) under laboratory conditions. Rom. Biotechnol. Lett. 2007, 12, 3295–3302. [Google Scholar]
- Bakonyi, J.; Láday, M.; Dula, T.; Érsek, T. Characterisation of Isolates of Phytophthora infestans From Hungary. Eur. J. Plant Pathol. 2002, 108, 139–146. [Google Scholar] [CrossRef]
- EPPO Standards. Phytophthora Infestans (PHYTIN) [Overview]|EPPO Global Database. 2019. Available online: https://gd.eppo.int/taxon/PHYTIN (accessed on 8 September 2019).
- Farr, D.F.; Rossman, A.Y. Fungal Databases, U.S. National Fungus Collections, ARS, USDA. 2020. Available online: https://nt.ars-grin.gov/fungaldatabases/ (accessed on 2 January 2020).
- Erwin, D.C.; Ribeiro, O.K. Phytophthora Diseases Worldwide; APS Press: St. Paul, MN, USA, 1996; p. 562. [Google Scholar]
- Andrivon, D. The origin of Phytophthora infestans populations present in Europe in the 1840s: A critical review of historical and scientific evidence. Plant Pathol. 1996, 45, 1027–1035. [Google Scholar] [CrossRef]
- Botero, K.; Restrepo, S.; Pinzón, A. A genome-scale metabolic model of potato late blight suggests a photosynthesis suppression mechanism. BMC Genom. 2018, 19, 863. [Google Scholar] [CrossRef] [Green Version]
- Olesen, M.H.; Jensen, P.K. Collection and evaluation of relevant information on crop interception. EFSA Support. Publ. 2013, 10, 438. [Google Scholar] [CrossRef]
- R Core Team (2013) R: A language and environment for statistical computing. Vienna, Austria. The R Foundation for Statistical Computing. 2019. Available online: http://www.R-project.org/ (accessed on 2 August 2019).
- Scheuerell, S.; Mahaffee, W. Compost Tea: Principles and Prospects For Plant Disease Control. Compos. Sci. Util. 2002, 10, 313–338. [Google Scholar] [CrossRef]
- Goufo, P.; Mofor, C.T.; Fontem, D.A.; Ngnokam, D. High efficacy of extracts of Cameroon plants against tomato late blight disease. Agron. Sustain. Dev. 2008, 28, 567–573. [Google Scholar] [CrossRef]
- Rashid, A.; Ahmad, I.; Iram, S.; Mirza, J.I. Efficiency of Different Neem (Azadirachta Indica A.Juss) Productsagainst Various Life Stages of Phytophthora Infestans (Mont.) DE Bary. Pak. J. Bot. 2004. Available online: http://agris.fao.org/agris-search/search.do?recordID=PK2005000971 (accessed on 20 September 2019).
- Wang, S.; Hu, T.; Zhang, F.; Forrer, H.R.; Cao, K. Screening for plant extracts to control potato late blight. Front. Agric. China 2007, 1, 43–46. [Google Scholar] [CrossRef]
- Majeed, A.; Ahmad, H.; Chaudhry, Z.; Jan, G.; Alam, J.; Muhammad, Z. Assessment of leaf extracts of three medicinal plants against late blight of potato in Kaghan valley, Pakistan. Int. J. Agric. Technol. 2011, 7, 1155–1161. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Year of Isolation | Location | Mating Type | Coordinates |
|---|---|---|---|---|
| 13/08/21 | 2013 | Érd (HU) | Unknown | Unknown |
| H-1/2015 | 2015 | Érd (HU) | Unknown | Unknown |
| M16 | Unknown | Monorierdő (HU) | A1 | N47 18.772, E19 29.118 |
| NY34 | 2006 | Nagykálló (HU) | A1 | Unknown |
| S40 | 2006 | Solt (HU) | A1 | N46 46.867, E19 02.621 |
| T17 | 2010 | Tordas (HU) | A1 | N47 20.448, E18 45.604 |
| T2 | 2010 | Tordas (HU) | Unknown | N47 19.817, E18 45.707 |
| Assessment Periods (Days) | Mean | * | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | |||
| Growth (mm) of strain 13/08/21 | ||||||||||||
| Control | 11.1 | 14 | 19.1 | 22.6 | 27.8 | 32.6 | 37.8 | 40.9 | 40.9 | 41 | 28.78 | A |
| 1 v/v% | 3.7 | 12.7 | 17.3 | 18.8 | 22.4 | 24.7 | 24.9 | 26.7 | 28.6 | 29.3 | 20.91 | A |
| 2 v/v% | 0 | 0 | 16 | 17.9 | 20.9 | 22.7 | 24.2 | 25.4 | 27.1 | 28.6 | 18.28 | A |
| 3 v/v% | 0 | 0 | 0 | 0 | 11.3 | 12.3 | 14 | 14 | 14 | 14.1 | 7.97 | B |
| 4 v/v% | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | C |
| Growth (mm) of strain H-1/2015 | ||||||||||||
| Control | 12.7 | 18.5 | 23.8 | 27.3 | 33.3 | 39.5 | 43.3 | 55 | 55 | 55 | 36.34 | A |
| 1 v/v% | 10.7 | 13.8 | 17.3 | 20 | 22.8 | 24.1 | 27.5 | 29.6 | 29.8 | 31.6 | 22.72 | B |
| 2 v/v% | 10.1 | 11.3 | 13 | 15 | 16.8 | 17.3 | 17.6 | 20.1 | 20.1 | 20.8 | 16.21 | C |
| 3 v/v% | 0 | 0 | 9.4 | 10.1 | 11.8 | 12.1 | 11.9 | 12.4 | 13 | 13.3 | 9.4 | D |
| 4 v/v% | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | E |
| Growth (mm) of strain M16 | ||||||||||||
| Control | 3.2 | 11.1 | 17.2 | 22.2 | 31.3 | 38.6 | 45 | 55 | 55 | 55 | 33.36 | A |
| 1 v/v% | 0 | 11.4 | 18.3 | 22.5 | 28.1 | 35.6 | 42.5 | 55 | 55 | 55 | 32.34 | A |
| 2 v/v% | 0 | 0 | 15 | 16.3 | 20.5 | 26 | 30.7 | 37.5 | 45.1 | 46.2 | 23.73 | AB |
| 3 v/v% | 0 | 0 | 11.6 | 15.7 | 15.7 | 18.4 | 21.4 | 27.3 | 28.6 | 32.5 | 17.12 | BC |
| 4 v/v% | 0 | 0 | 0 | 0 | 16.5 | 19.8 | 21.7 | 23.9 | 29.2 | 32.5 | 14.36 | C |
| Growth (mm) of strain NY34 | ||||||||||||
| Control | 17.2 | 29.5 | 38.2 | 48.6 | 55 | 55 | 55 | 55 | 55 | 55 | 46.35 | A |
| 1 v/v% | 13.3 | 18.2 | 23 | 25.5 | 29.4 | 34.3 | 38.2 | 44.6 | 47.9 | 48.9 | 32.33 | B |
| 2 v/v% | 0 | 0 | 15 | 16.8 | 18.3 | 20.3 | 22.8 | 24.5 | 25.1 | 27.8 | 17.06 | C |
| 3 v/v% | 0 | 0 | 12 | 12.9 | 12.9 | 14.7 | 17.2 | 19.1 | 19.3 | 20.2 | 12.83 | D |
| 4 v/v% | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | E |
| Growth (mm) of strain S40 | ||||||||||||
| Control | 14 | 24.8 | 32.5 | 39.9 | 50.4 | 55 | 55 | 55 | 55 | 55 | 43.66 | A |
| 1 v/v% | 4.5 | 13 | 21.2 | 24.7 | 30.5 | 35.5 | 39.7 | 43.5 | 44.2 | 47.6 | 30.44 | B |
| 2 v/v% | 0 | 0 | 13.8 | 15.8 | 20.5 | 25.3 | 29.2 | 33.6 | 36.8 | 43 | 21.8 | C |
| 3 v/v% | 0 | 0 | 0 | 0 | 11.2 | 12.7 | 12.9 | 15.1 | 15.8 | 15.9 | 8.36 | D |
| 4 v/v% | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | E |
| Growth (mm) of strain T17 | ||||||||||||
| Control | 13.6 | 19.1 | 28.1 | 34.8 | 43.9 | 55 | 55 | 55 | 55 | 55 | 41.45 | A |
| 1 v/v% | 11.3 | 16.8 | 20.9 | 25.2 | 29.6 | 35.5 | 39.7 | 55 | 55 | 55 | 34.4 | A |
| 2 v/v% | 0 | 0 | 16.1 | 19.2 | 24.2 | 27.4 | 29.2 | 33.3 | 38.8 | 46.4 | 23.46 | B |
| 3 v/v% | 0 | 0 | 10.5 | 11.6 | 13.8 | 15.5 | 15.9 | 17 | 20.1 | 20.9 | 12.53 | C |
| 4 v/v% | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | D |
| Growth (mm) of strain T2 | ||||||||||||
| Control | 17 | 22.8 | 29 | 33.8 | 42.8 | 51 | 55 | 55 | 55 | 55 | 41.64 | A |
| 1 v/v% | 13 | 15.8 | 19.8 | 22 | 24.7 | 27.4 | 29 | 30.4 | 32.1 | 33.3 | 24.75 | B |
| 2 v/v% | 11.8 | 13.7 | 16.9 | 19.1 | 21.8 | 24 | 25.1 | 27.4 | 29.2 | 30.9 | 21.99 | B |
| 3 v/v% | 0 | 0 | 11.4 | 12.2 | 13.5 | 14.2 | 15.7 | 15.7 | 15.7 | 15.7 | 11.41 | C |
| 4 v/v% | 0 | 0 | 0 | 0 | 0 | 0 | 10.7 | 11.3 | 11.3 | 11.3 | 4.46 | D |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Turóczi, B.; Bakonyi, J.; Szabó, K.-A.; Bálint, J.; Máthé, I.; Lányi, S.; Balog, A. In Vitro and In Vivo Effect of Poplar Bud Extracts on Phytophthora infestans: A New Effective Biological Method in Potato Late Blight Control. Plants 2020, 9, 217. https://doi.org/10.3390/plants9020217
Turóczi B, Bakonyi J, Szabó K-A, Bálint J, Máthé I, Lányi S, Balog A. In Vitro and In Vivo Effect of Poplar Bud Extracts on Phytophthora infestans: A New Effective Biological Method in Potato Late Blight Control. Plants. 2020; 9(2):217. https://doi.org/10.3390/plants9020217
Chicago/Turabian StyleTuróczi, Botond, József Bakonyi, Károly-Attila Szabó, János Bálint, István Máthé, Szabolcs Lányi, and Adalbert Balog. 2020. "In Vitro and In Vivo Effect of Poplar Bud Extracts on Phytophthora infestans: A New Effective Biological Method in Potato Late Blight Control" Plants 9, no. 2: 217. https://doi.org/10.3390/plants9020217