The Use of Proteomic Tools to Address Challenges Faced in Clonal Propagation of Tropical Crops through Somatic Embryogenesis

Abstract
:1. Introduction
2. Tissue Culture in Tropical Crops
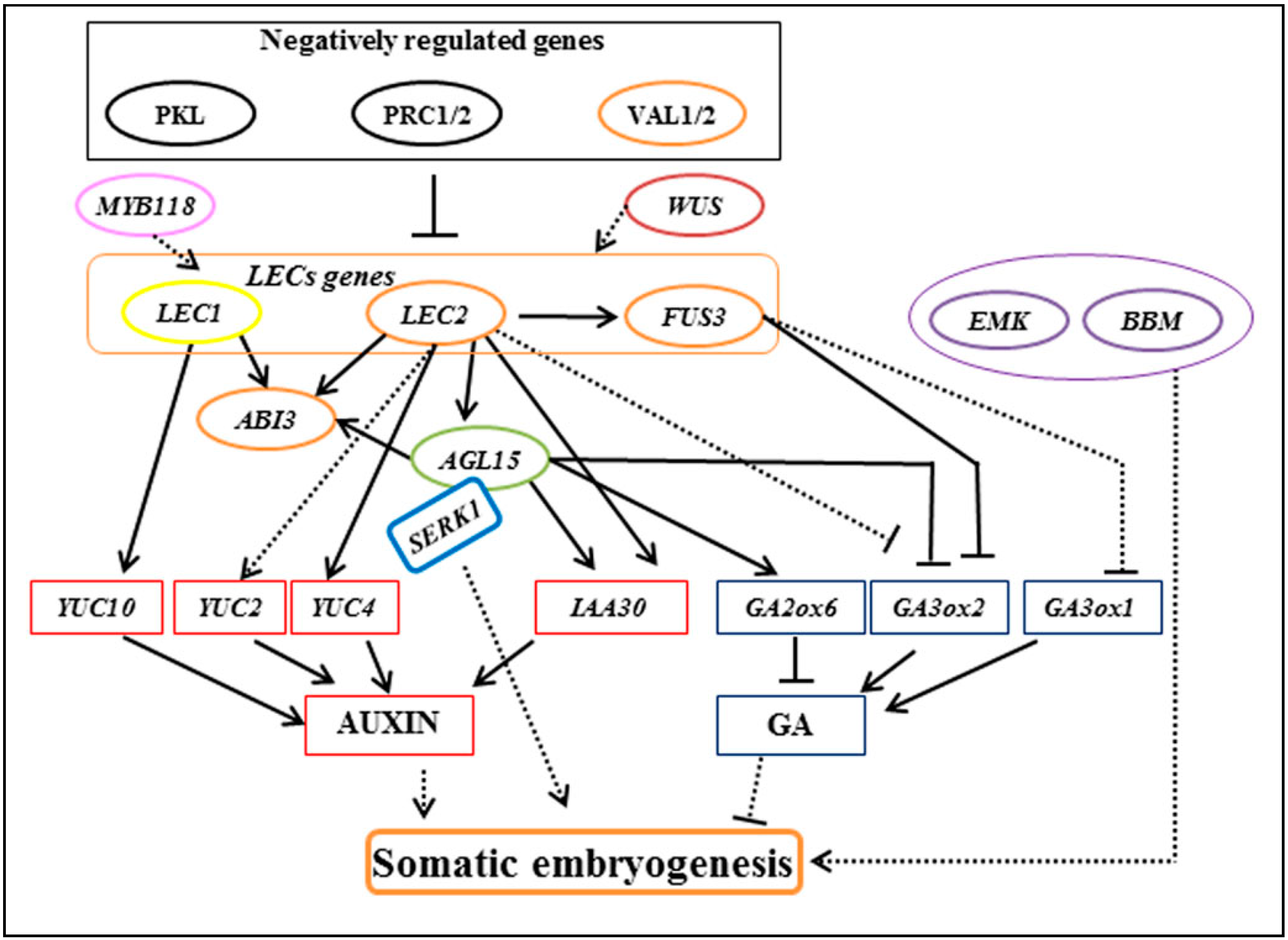
3. Regulatory Genes in Somatic Embryogenesis
4. The Development of Proteomic Technology in Crops
5. Proteomics in Somatic Embryogenesis
6. Application of Proteomics in Tissue Culture of Tropical Crops
7. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Wilkinson, A. Expanding tropics will play greater global role, report predicts. Science 2014, 344, 194. [Google Scholar]
- Thorpe, T.A. History of Plant Tissue Culture; Humana Press Inc.: Totowa, NJ, USA, 2007. [Google Scholar]
- Sugimoto, K.; Gordon, S.P.; Meyerowitz, E.M. Regeneration in plants and animals: Dedifferentiation, transdifferentiation, or just differentiation? Trends Cell Biol. 2011, 21, 212–218. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.C.W.; Thorpe, T.A. Crop improvement through tissue culture. World J. Microbiol. Biotechnol. 1995, 11, 409–415. [Google Scholar] [CrossRef] [PubMed]
- Staritsky, G. Tissue culture of the oil palm (Elaeis guineensis Jacq.) as a tool for its vegetative propagation. Euphytica 1970, 19, 288–292. [Google Scholar] [CrossRef]
- Alvard, D.; Cote, F.; Teisson, C. Comparison of methods of liquid medium culture for banana Micropropagation—Effects of temporary immersion of explants. Plant Cell Tissue Organ Cult. 1993, 32, 55–60. [Google Scholar] [CrossRef]
- Sripaoraya, S.; Marchant, R.; Power, J.B.; Davey, M.R. Plant regeneration by somatic embryogenesis and organogenesis in commercial pineapple (Ananas Comosus L.). In Vitro Cell. Dev. Biol. Plant 2003, 39, 450–454. [Google Scholar] [CrossRef]
- Zhao, H.; Peng, M.; Wang, X.; Zeng, H.C.; Chen, X.T. Micropropagation of rubber tree (Hevea brasiliensis) by employing mature stem as explants. Genom. Appl. Biol. 2009, 28, 1169–1176. [Google Scholar]
- Gavinlertvatana, P. Commercial micropropagation of tropical fruit trees. Acta Hortic. 1992, 321, 574–578. [Google Scholar] [CrossRef]
- Vasil, I.K. Plant tissue culture and molecular biology as tools in understanding plant development and in plant improvement. Curr. Opin. Biotechnol. 1991, 2, 158–163. [Google Scholar] [CrossRef]
- Namasivayam, P. Acquisition of embryogenic competence during somatic embryogenesis. Plant Cell Tissue Organ Cult. 2007, 90, 1–8. [Google Scholar] [CrossRef]
- Feher, A.; Pasternak, T.P.; Dudits, D. Transition of somatic plant cells to an embryogenic state. Plant Cell Tissue Organ Cult. 2003, 74, 201–228. [Google Scholar] [CrossRef]
- Von Arnold, S.; Sabala, I.; Bozhkov, P.; Dyachok, J.; Filonova, L. Developmental pathways of somatic embryogenesis. Plant Cell Tissue Organ Cult. 2002, 69, 233–249. [Google Scholar] [CrossRef]
- Loschiavo, F.; Pitto, L.; Giuliano, G.; Torti, G.; Nuti-Ronchi, V.; Marazziti, D.; Vergara, R.; Orselli, S.; Terzi, M. DNA methylation of embryogenic carrot cell cultures and its variations as caused by maturation, differentiation, hormones and hypomethylation drugs. Theor. Appl. Genet. 1989, 77, 325–331. [Google Scholar] [CrossRef] [PubMed]
- Guan, Y.; Li, S.G.; Fan, X.F.; Su, Z.H. Applications of somatic embryogenesis in woody plants. Front. Plant Sci. 2016, 7, 938. [Google Scholar] [CrossRef] [PubMed]
- Elhiti, M.; Stasolla, C.; Wang, A. Molecular regulation of plant somatic embryogenesis. In Vitro Cell. Dev. Biol. Plant 2013, 49, 631–642. [Google Scholar] [CrossRef]
- Stone, S.L.; Braybrook, S.A.; Paula, S.L.; Kwong, L.W.; Meuser, J.; Pelletier, J.; Hsieh, T.F.; Fischer, R.L.; Goldberg, R.B.; Harada, J.J. Arabidopsis LEAFY COTYLEDON2 induces maturation traits and auxin activity: Implications for somatic embryogenesis. Proc. Natl. Acad. Sci. USA 2008, 105, 3151–3156. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.M.; Xu, Z.H.; Chua, N.H. Auxin polar transport is essential for the establishment of bilateral symmetry during early plant embryogenesis. Plant Cell 1993, 5, 621–630. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, E.D.; Guzzo, F.; Toonen, M.A.; de Vries, S.C. A leucine-rich repeat containing receptor-like kinase marks somatic plant cells competent to form embryos. Development 1997, 124, 2049–2062. [Google Scholar] [PubMed]
- Boutilier, K.; Offringa, R.; Sharma, V.K.; Kieft, H.; Ouellet, T.; Zhang, L.; Hattori, J.; Liu, C.M.; van Lammeren, A.A.; Miki, B.L.; et al. Ectopic expression of BABY BOOM triggers a conversion from vegetative to embryonic growth. Plant Cell 2002, 14, 1737–1749. [Google Scholar] [CrossRef] [PubMed]
- Zuo, J.; Niu, Q.W.; Frugis, G.; Chua, N.H. The WUSCHEL gene promotes vegetative-to-embryonic transition in Arabidopsis. Plant J. 2002, 30, 349–359. [Google Scholar] [CrossRef] [PubMed]
- Tobias, M.; Marc, G.; Luis, S. Correlation of mRNA and protein in complex biological samples. FEBS Lett. 2009, 583, 3966–3973. [Google Scholar] [CrossRef]
- Wilkins, M.R.; Gasteiger, E.; Gooley, A.A.; Appel, R.D.; Humphery-Smith, I.; Hochstrasser, D.F.; William, K.L. Progress with proteome projects: Why all protein expressed by a genome should be identified and how to do it. Biotechnol. Genet. Eng. Rev. 1996, 13, 19–50. [Google Scholar] [CrossRef] [PubMed]
- The Arabidopsis Genome Initiative. Analysis of the genome sequence of the flowering plant Arabidopsis thaliana. Nature 2000, 408, 796–815. [Google Scholar]
- Goff, S.A.; Ricke, D.; Lan, T.H.; Presting, G.; Wang, R.; Dunn, M.; Glazebrook, J.; Sessions, A.; Oeller, P.; Varma, H.; et al. A Draft Sequence of the Rice Genome (Oryza sativa L. ssp. japonica). Science 2002, 296, 92–100. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Hu, S.; Wang, J.; Wong, G.K.S.; Li, S.; Liu, B.; Deng, Y.; Dai, L.; Zhou, Y.; Zhang, X.; et al. A Draft Sequence of the Rice Genome (Oryza sativa L. ssp. indica). Science 2002, 296, 79–92. [Google Scholar] [CrossRef] [PubMed]
- Majeran, W.; Friso, G.; Ponnala, L.; Connolly, B.; Huang, M.; Reidel, E.; Zhang, C.; Asakura, Y.; Bhuiyan, N.H.; Sun, Q.; et al. Structural and Metabolic Transitions of C4 Leaf Development and Differentiation Defined by Microscopy and Quantitative Proteomics in Maize. Plant Cell 2010, 22, 3509–3542. [Google Scholar] [CrossRef] [PubMed]
- Peng, Z.; Wang, M.; Li, F.; Lv, H.; Li, C.; Xia, G. A proteomic study of the response to salinity and drought stress in an introgression strain of bread wheat. Mol. Cell. Proteom. 2009, 8, 2676–2686. [Google Scholar] [CrossRef] [PubMed]
- Moller, A.L.B.; Pedas, P.; Andersen, B.; Svensson, B.; Schjoerring, J.K.; Finnie, C. Responses of barley root and shoot proteomes to long-term nitrogen deficiency, short-term nitrogen starvation and ammonium. Plant Cell Environ. 2011, 34, 2024–2037. [Google Scholar] [CrossRef] [PubMed]
- Rasoulnia, A.; Bihamta, M.R.; Peyghambari, S.A.; Alizadeh, H.; Rahnama, A. Proteomic response of barley leaves to salinity. Mol. Biol. Rep. 2011, 38, 5055–5063. [Google Scholar] [CrossRef] [PubMed]
- Komatsu, S.; Makino, T.; Yasue, H. Proteomic and Biochemical Analyses of the Cotyledon and Root of Flooding-Stressed Soybean Plants. PLoS ONE 2013, 8, e65301. [Google Scholar] [CrossRef] [PubMed]
- Subba, P.; Kumar, R.; Gayali, S.; Shekhar, S.; Parveen, S.; Pandey, A.; Datta, A.; Chakraborty, S.; Chakraborty, N. Characterisation of the nuclear proteome of a dehydration-sensitive cultivar of chickpea and comparative proteomic analysis with a tolerant cultivar. Proteomics 2013, 13, 1973–1992. [Google Scholar] [CrossRef] [PubMed]
- Marondedze, C.; Gehring, C.; Thomas, L. Dynamic changes in the date palm fruit proteome during development and ripening. Hortic. Res. 2014, 1, 14039. [Google Scholar] [CrossRef] [PubMed]
- Jorrín-Novo, J.V.; Pascual, J.; Sánchez-Lucas, R.; Romero-Rodríguez, M.C.; Rodríguez-Ortega, M.J.; Lenz, C.; Valledor, L. Fourteen years of plant proteomics reflected in Proteomics: Moving from model species and 2DE-based approaches to orphan species and gel-free platforms. Proteomics 2015, 15, 1089–1112. [Google Scholar] [CrossRef] [PubMed]
- O’Farrell, P.H. High resolution two-dimensional electrophoresis of proteins. J. Biol. Chem. 1975, 250, 4007–4021. [Google Scholar] [PubMed]
- Huang, B.R.; Xu, C.P.; Xu, Y. Protein extraction for two-dimensional gel electrophoresis of proteomic profiling in turfgrass. Crop Sci. 2008, 48, 1608–1614. [Google Scholar]
- Friedman, D.B.; Lilley, K.S. Difference gel electrophoresis (DIGE). In The Protein Protocols Handbook, 3rd ed.; John, W., Ed.; Human Press: Totowa, NJ, USA, 2009; pp. 379–408. [Google Scholar]
- Lilley, K.S.; Freidman, D.B. All about DIGE: Quantification technology for differential display 2D-gel proteomics. Expert Rev. Proteom. 2004, 1, 401–409. [Google Scholar] [CrossRef] [PubMed]
- Unlu, M.; Morgan, M.E.; Minden, J.S. Difference gel electrophoresis: A single gel method for detecting changes in protein extracts. Electrophoresis 1997, 18, 2027–2077. [Google Scholar] [CrossRef] [PubMed]
- Colignon, B.; Raes, M.; Dieu, M.; Delaive, E.; Mauro, S. Evaluation of three-dimensional gel electrophoresis to improve quantitative profiling of complex proteomes. Proteomics 2013, 13, 2077–2082. [Google Scholar] [CrossRef] [PubMed]
- Baerenfaller, K.; Grossmann, J.; Grobei, M.A.; Hull, R.; Hirsch-Hoffmann, M.; Yalovsky, S.; Zimmermann, P.; Grossniklaus, U.; Gruissem, W.; Baginsky, S. Genome-scale proteomics reveals Arabidopsis thaliana gene models and proteome dynamics. Science 2008, 320, 938–941. [Google Scholar] [CrossRef] [PubMed]
- Imin, N.; Kerim, T.; Weinman, J.J.; Rolfe, B.G. Characterisation of rice anther proteins expressed at the young microspore stage. Proteomics 2001, 1, 1149–1161. [Google Scholar] [CrossRef]
- Ferchab, A.; Capriotti, A.L.; Caruso, G.; Cavaliere, C.; Samperi, R.; Stampachiacchiere, S.; Laganà, A. Comparative analysis of metabolic proteome variation in ascorbate-primed and unprimed wheat seeds during germination under salt stress. J. Proteom. 2014, 108, 238–257. [Google Scholar] [CrossRef] [PubMed]
- Capriotti, A.L.; Borrelli, G.M.; Colapicchioni, V.; Papa, R.; Piovesana, S.; Samperi, R.; Stampachiacchiere, S.; Laganà, A. Proteomic study of a tolerant genotype of durum wheat under salt-stress conditions. Anal. Bioanal. Chem. 2014, 406, 1423–1435. [Google Scholar] [CrossRef] [PubMed]
- Fercha, A.; Capriotti, A.L.; Caruso, G.; Cavaliere, C.; Stampachiacchiere, S.; Chiozzi, R.Z.; Laganà, A. Shotgun proteomic analysis of soybean embryonic axes during germination under salt stress. Proteomics 2016, 16, 1537–1546. [Google Scholar] [CrossRef] [PubMed]
- Capriotti, A.L.; Caruso, G.; Cavaliere, C.; Samperi, R.; Stampachiacchiere, S.; Chiozzi, R.Z.; Laganà, A. Protein Profile of Mature Soybean Seeds and Prepared Soybean Milk. J. Agric. Food Chem. 2014, 62, 9893–9899. [Google Scholar] [CrossRef] [PubMed]
- Aebersold, R.; Mann, M. Mass spectrometry-based proteomics. Nature 2003, 422, 198–207. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.H.; Sung, Z.R. Review and Perspectives: Two-dimensional gel analysis of carrot somatic embryonic proteins. Plant Mol. Biol. Rep. 1984, 2, 19–25. [Google Scholar] [CrossRef]
- Helleboid, S.; Hendriks, T.; Bauw, G.; Inze, D.; Vasseur, J.; Hilbert, J.-L. Three major somatic embryogenesis related proteins in Cichorium identified as PR proteins. J. Exp. Bot. 2000, 51, 1189–1200. [Google Scholar] [CrossRef] [PubMed]
- Gianazza, E.; De Ponti, P.; Scienza, A.; Villa, P.; Martinelli, L. Monitoring by two-dimensional electrophoresis somatic embryogenesis in leaf and petiole explants from Vitis. Electrophoresis 1992, 13, 203–209. [Google Scholar] [CrossRef] [PubMed]
- Sallandrouze, A.; Faurobert, M.; El Maataoui, M.; Espagnac, H. Two dimensional electrophoresis analysis of proteins associated with somatic embryogenesis development in Cupressus sempervirens L. Electrophoresis 1999, 20, 1109–1119. [Google Scholar] [CrossRef]
- Bian, F.; Zheng, C.; Qu, F.; Gong, X.; You, C. Proteomic Analysis of Somatic Embryogenesis in Cyclamen persicum Mill. Plant Mol. Biol. Rep. 2010, 28, 22–31. [Google Scholar] [CrossRef]
- Lippert, D.; Zhuang, J.; Ralph, S.; Ellis, D.E.; Gilbert, M.; Olafson, R.; Ritland, K.; Ellis, B.; Douglas, C.J.; Bohlmann, J. Proteome analysis of early somatic embryogenesis in Picea glauca. Proteomics 2005, 5, 461–473. [Google Scholar] [CrossRef] [PubMed]
- Flinn, B.S.; Roberts, D.R.; Newton, C.H.; Cyr, D.R.; Webster, F.B.; Taylor, I.E.P. Storage protein gene exprcssion in zygotic and somatic embryos of interior spruce. Physiol. Plant 1993, 89, 719–730. [Google Scholar] [CrossRef]
- Lal, S.K.; Lee, C.; Sachs, M.M. Differential regulation of enolase during anaerobiosis in maize. Plant Physiol. 1998, 118, 1285–1293. [Google Scholar] [CrossRef] [PubMed]
- Imin, N.; Nizamidin, M.; Daniher, D.; Nolan, K.E.; Rose, R.J.; Rolfe, B.G. Proteomic analysis of somatic embryogenesis in Medicago truncatula. Explant cultures grown under 6-Benzylaminopurine and 1-Napthaleacetic Acid treatment. Plant Physiol. 2005, 137, 1250–1260. [Google Scholar] [CrossRef] [PubMed]
- Jong, F.; Mathesius, U.; Imin, N.; Rolfe, B.G. A Proteome study of the proliferation of cultured Medicago truncatula protoplasts. Plant Proteom. 2007, 7, 722–736. [Google Scholar] [CrossRef] [PubMed]
- Amsterdam, A.; Pitzer, F.; Baumeister, W. Changes in intracellular localization of proteasomes in immortalized ovarian granulosa cells during mitosis associated with a role in cell cycle control. Proc. Natl. Acad. Sci. USA 1993, 90, 99–103. [Google Scholar] [CrossRef] [PubMed]
- Ito, H.; Iwabuchi, M.; Ogawa, K. The sugar-metabolic enzymes aldolase and triose-phosphate isomerase are targets of glutathionylation in Arabidopsis thaliana: Detection using biotinylated glutathione. Plant Cell Physiol. 2003, 44, 655–660. [Google Scholar] [CrossRef] [PubMed]
- Heringer, A.S.; Santa-Catarina, C.; Silveira, V. Insights from proteomic studies into plant somatic embryogenesis. Proteomics 2018, 18, e1700265. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, G.K.; Job, D.; Zivy, M.; Agrawal, V.P.; Bradshaw, R.A.; Dunn, M.J.; Haynes, P.A.; Wijk, K.J.V.; Kikichi, S.; Renaut, J.; et al. Time to articulate a vision for the future of plant proteomics—A global perspective: An initiative for establishing the International Plant Proteomics Organization (INPPO). Proteomics 2011, 11, 1559–1568. [Google Scholar] [CrossRef] [PubMed]
- Subhra, C.; Ghasem, H.S.; Yang, P.F.; Woo, S.H.; Chin, C.F.; Chris, G.; Paul, A.H.; Mehdi, M.; Komatsu, S. Proteomics of Important Food crops in the Asia Oceania Region: Current Status and Future Perspectives. J. Proteome Res. 2015, 14, 2723–2744. [Google Scholar]
- Campos, N.A.; Paiva, L.V.; Panis, B.; Carpentier, S.C. The proteome profile of embryogenic cell suspensions of Coffea arabica L. Proteomics 2016, 16, 1001–1005. [Google Scholar] [CrossRef] [PubMed]
- Correia, S.; Vinhas, R.; Manadas, B.; Lourenço, A.S.; Veríssimo, P.; Canhoto, J.M. Comparative proteomic analysis of auxin-induced embryogenic and nonembryogenic tissues of the Solanaceous tree Cyphomandra betacea (Tamarillo). J. Proteome Res. 2012, 11, 1666–1675. [Google Scholar] [CrossRef] [PubMed]
- Tan, H.S.; Liddell, S.; Abdullah, M.O.; Wong, W.C.; Chin, C.F. Differential proteomic analysis of embryogenic lines in oil palm (Elaeis guineensis Jacq). J. Proteom. 2016, 143, 334–345. [Google Scholar] [CrossRef] [PubMed]
- De Carvalho Silva, R.; Travassos Carmo, L.S.; Gomes Luis, Z.; Silva, L.P.; Scherwinski-Pereira, J.E.; Mehta, A. Proteomic identification of differentially expressed proteins during the acquisition of somatic embryogenesis in oil palm (Elaeis guineensis Jacq.). J. Proteom. 2014, 104, 112–127. [Google Scholar] [CrossRef] [PubMed]
- Baba, A.; Nogueira, F.; Pinheiro, C.; Brasil, J.; Jereissati, E.; Juca, T.; Soares, A.; Santos, M.; Domont, G.; Campos, F. Proteome analysis of secondary somatic embryogenesis in cassava (Manihot esculenta). Plant Sci. 2008, 175, 717–723. [Google Scholar] [CrossRef]
- Kumaravel, M.; Uma, S.; Backiyarani, S.; Saraswathi, M.S.; Vaganan, M.M.; Muthusamy, M.; Sajith, K.P. Differential proteome analysis during early somatic embryogenesis in Musa spp. AAA cv. Grand Naine. Plant Cell Rep. 2017, 36, 163–178. [Google Scholar] [CrossRef] [PubMed]
- Guzmán-García, E.; Sánchez-Romero, C.; Panis, B.; Carpentier, S.C. The use of 2D-DIGE to understand the regeneration of somatic embryos in avocado. Proteomics 2013, 13, 3498–3507. [Google Scholar] [CrossRef] [PubMed]
- Sghaier-Hammami, B.; Driraa, N.; Jorrín-Novob, J.V. Comparative 2-DE proteomic analysis of date palm (Phoenix dactylifera L.) somatic and zygotic embryos. J. Proteom. 2009, 73, 161–177. [Google Scholar] [CrossRef] [PubMed]
- Heringer, A.S.; Reis, R.S.; Passamani, L.Z.; de Souza-Filho, G.A.; Santa-Catarina, C.; Silveira, V. Comparative proteomics analysis of the effect of combined red and blue lights on sugarcane somatic embryogenesis. Acta Physiol. Plant. 2017, 39, 52. [Google Scholar] [CrossRef]
- Niemenak, N.; Kaiser, E.; Maximova, S.N.; Laremore, T.; Guiltinan, M.J. Proteome analysis during pod, zygotic and somatic embryo maturation of Theobroma cacao. J. Plant Physiol. 2015, 180, 49–60. [Google Scholar] [CrossRef] [PubMed]
- Noaha, A.M.; Niemenaka, N.; Sunderhausb, S.; Haaseb, C.; Omokoloa, D.N.; Winkelmannc, T.; Braun, H.P. Comparative proteomic analysis of early somatic and zygotic embryogenesis in Theobroma cacao L. J. Porteom. 2013, 78, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Tan, B.C.; Chin, C.F.; Liddell, S.; Alderson, P. Proteomic analysis of callus development in Vanilla planifolia Andrews. Plant Mol. Biol. Rep. 2013, 31, 1220–1229. [Google Scholar] [CrossRef]
- Cassells, A.C.; Curry, R.F. Oxidative stress and physiological, epigenetic and genetic variability in plant tissue culture, implications for micropropagators and genetic engineers. Plant Cell Tissue Organ Cult. 2001, 64, 145–157. [Google Scholar] [CrossRef]


{kind=link}
| Scientific Name (Common Name) | Explants | Micropropagation Methods | Proteomic Techniques | References |
|---|---|---|---|---|
| Coffea arabica (coffee) | Leaf | SE | Q Exactive Orbitrap MS | [63] |
| Cyphomandra betacea (tamarillo) | Leaf | SE | LC MS/MS | [64] |
| Elaeis guineensis (oil palm) | (a) Leaf (b) Zygotic embryos | SE | 2-DE & MALDI ToF MS/MS | [65,66] |
| Manihot esculenta (cassava) | Green cotyledons of somatic Embryos | SE | 2-DE/MALDI-MSMS | [67] |
| Musa spp. (banana) | Immature male flower buds | SE | 2-DE & MALDI-Tof MS | [68] |
| Persea americana (avocado) | Immature zygotic embryos | SE | 2D-DIGE | [69] |
| Phoenix dactylifera (date palm) | Embryos | SE | 2-DE & MALDI-Tof MS/MS | [70] |
| Saccharum spp. (sugarcane) | nodal segments with axillary buds | SE | ESI-QTOF HDMS | [71] |
| Theobroma cacao (cocoa) | (a) Flower, zygotic embryos (b) Zygotic embryos | SE | (a) 2DE and nano-LC-MS (b) 2DE and EASY-nLC coupled with Micro-ToF-Q | [72,73] |
| Vanilla planifolia (vanilla) | nodal segments with axillary buds | OG | 2DE and MALDI Tof/Tof/MS | [74] |
| Plant Type | Protein | Cellular Functions | References |
|---|---|---|---|
| Cyphomandra betacea (tamarillo) | fructokinase | glycolysis | [64] |
| Pathogenesis-related proteins | Stress association | ||
| Heat shock 70 kDa | Stress association | ||
| enolase | glycolysis | ||
| Elaeis guineensis (oil palm) | triosephosphate isomerase | glycolysis | [65] |
| L- ascorbate peroxidase | Defence response | ||
| superoxide dismutase | defence response | ||
| Elaeis guineensis (oil palm) | type IIIa membrane protein cp-wap13 | cell wall degradation, loosening and biosynthesis | [66] |
| fructokinase | glycolysis | ||
| PR proteins (peroxidase and glutathione S-transferase) | Stress association | ||
| Musa spp. (banana) | indole-3-pyruvate monooxygenase | Auxin synthesis | [68] |
| adenylate isopentenyltransferase | Cytokinin synthesis | ||
| Acyl-acyl-carrier-protein desaturase | fatty acid biosynthesis | ||
| pectinesterase inhibitor | Inhibit pectin accumulation | ||
| Caffeoyl-CoA O-methyltransferase | lignin biosynthesis | ||
| Persea americana (avocado) | superoxide dismutase | Defence response | [69] |
| Heat shock 70 kDa | Stress association | ||
| Glutathione S-transferase | Detoxification process | ||
| Theobroma cacao (cocoa) | β-1,3 glucanases | Stress association | [72] |
| chitinase | Stress association | ||
| osmotin-like protein | Stress association |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chin, C.F.; Tan, H.S. The Use of Proteomic Tools to Address Challenges Faced in Clonal Propagation of Tropical Crops through Somatic Embryogenesis. Proteomes 2018, 6, 21. https://doi.org/10.3390/proteomes6020021
Chin CF, Tan HS. The Use of Proteomic Tools to Address Challenges Faced in Clonal Propagation of Tropical Crops through Somatic Embryogenesis. Proteomes. 2018; 6(2):21. https://doi.org/10.3390/proteomes6020021
Chicago/Turabian StyleChin, Chiew Foan, and Hooi Sin Tan. 2018. "The Use of Proteomic Tools to Address Challenges Faced in Clonal Propagation of Tropical Crops through Somatic Embryogenesis" Proteomes 6, no. 2: 21. https://doi.org/10.3390/proteomes6020021
APA StyleChin, C. F., & Tan, H. S. (2018). The Use of Proteomic Tools to Address Challenges Faced in Clonal Propagation of Tropical Crops through Somatic Embryogenesis. Proteomes, 6(2), 21. https://doi.org/10.3390/proteomes6020021