
Arc Regulates Transcription of Genes for Plasticity, Excitability and Alzheimer’s Disease

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Chemicals
2.2. Culturing Hippocampal and Cortical Neurons
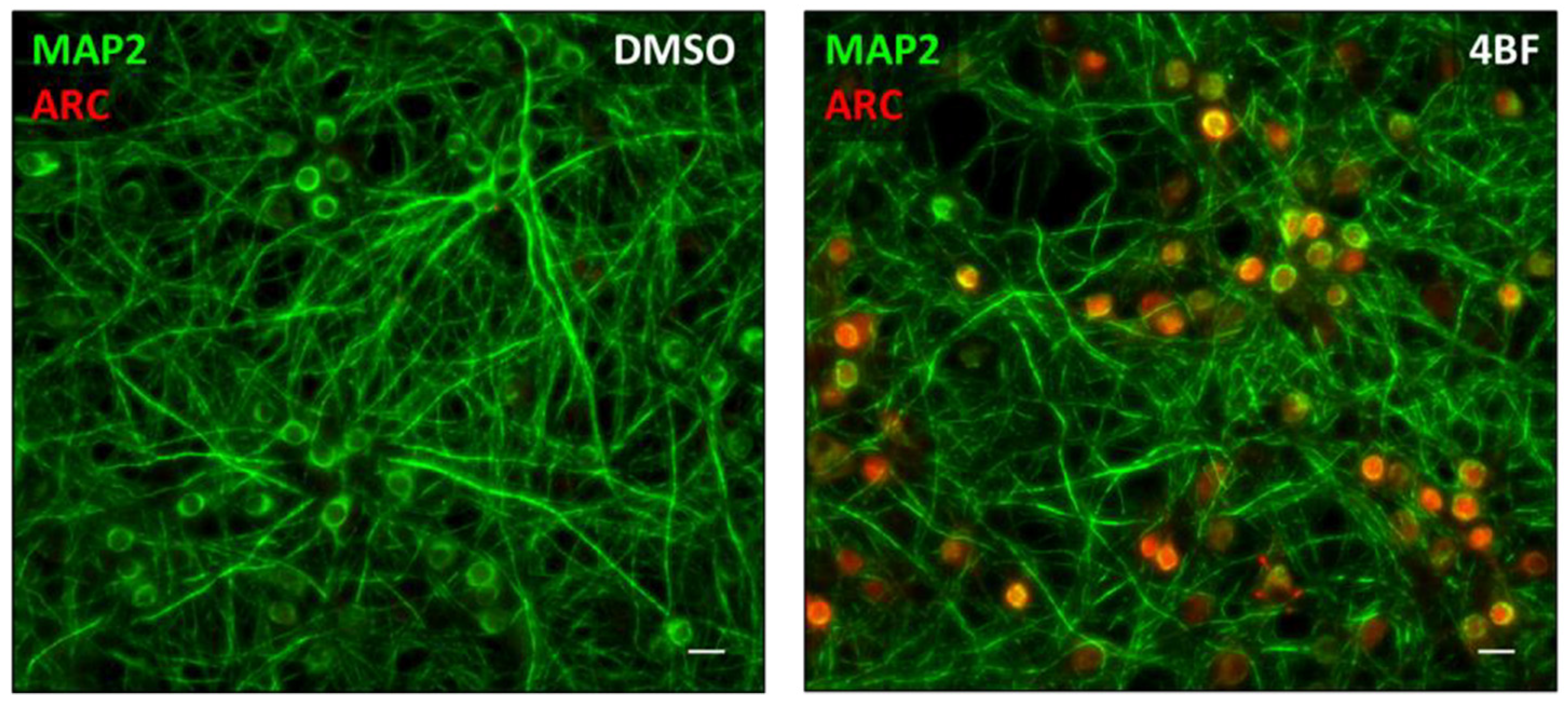
2.3. Pharmacological LTP Using 4BF
2.4. Immunofluorescence
2.5. Inhibition of Arc Expression by an shRNA
2.6. Transfection of Neuronal Cultures
2.7. Widefield Microscopy
2.8. Spinning Disc Confocal Microscopy
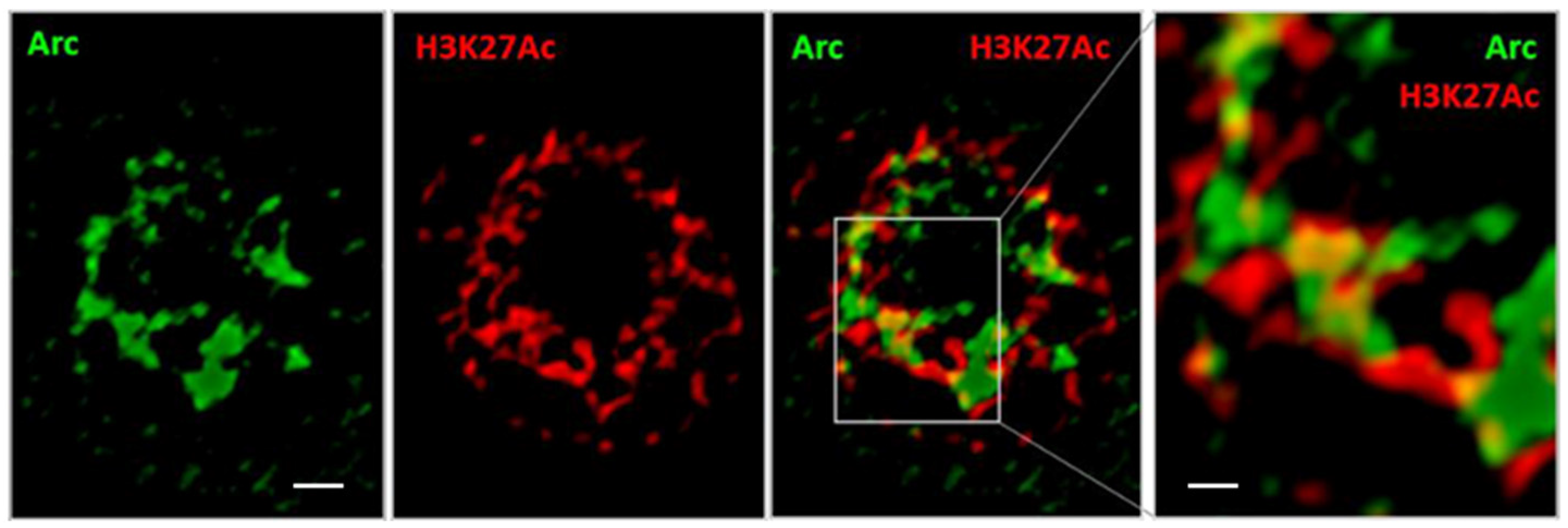
2.9. Stochastic Optical Reconstruction Microscopy (STORM)
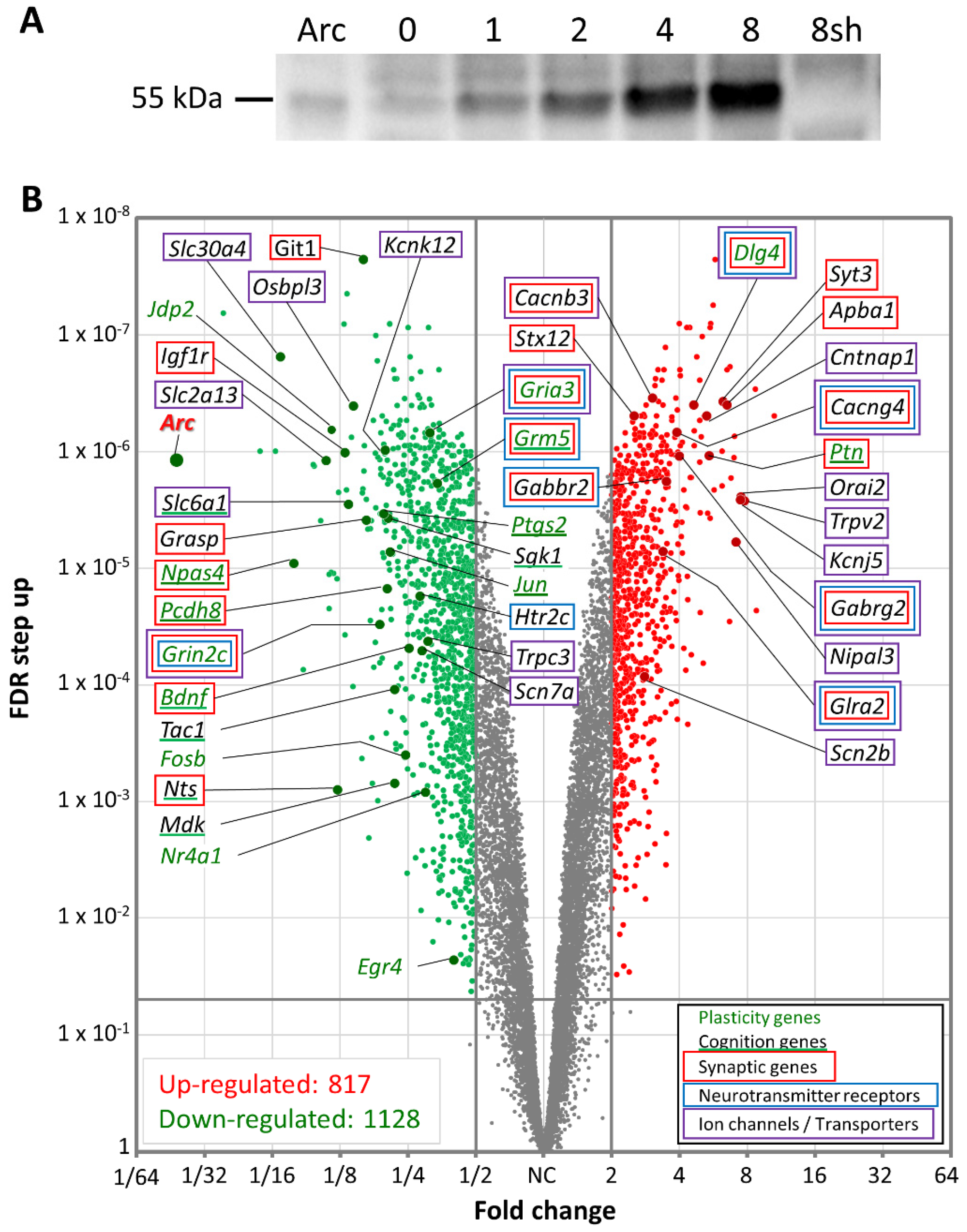
2.10. Cell lysate Preparation and Western Blotting
2.11. RNA Sample Preparation, Library Construction, RNA-Seq
2.12. Computational Analyses of RNA-Seq Data
2.13. Plasmid Construction for Arc Expression in HEK293T Cells
2.14. Transfection for Endogenous Arc Overexpression and Purification of mRNA
| Promoter | SgRNA Sequence |
| Human Arc (1) | GGGCGCTGGCGGG- GAGCCTG |
| Human Arc (2) | CCTCCCGTCCCTT- GCCGCCC |
| LacZ (1) | TTCCGGCTCGTATGTT- GTGT |
| LacZ (2) | GCTTTACACTTTATGCTTCC |
3. Results
3.1. Chromatin Reorganisation in Arc-Positive Neurons
3.2. Arc Associates with Dynamic Chromatin
3.3. Arc Associates with a Marker of Active Enhancers
3.4. Arc Associates with a Marker for Active Transcription
3.5. Arc Regulates Activity-Dependent Gene Transcription
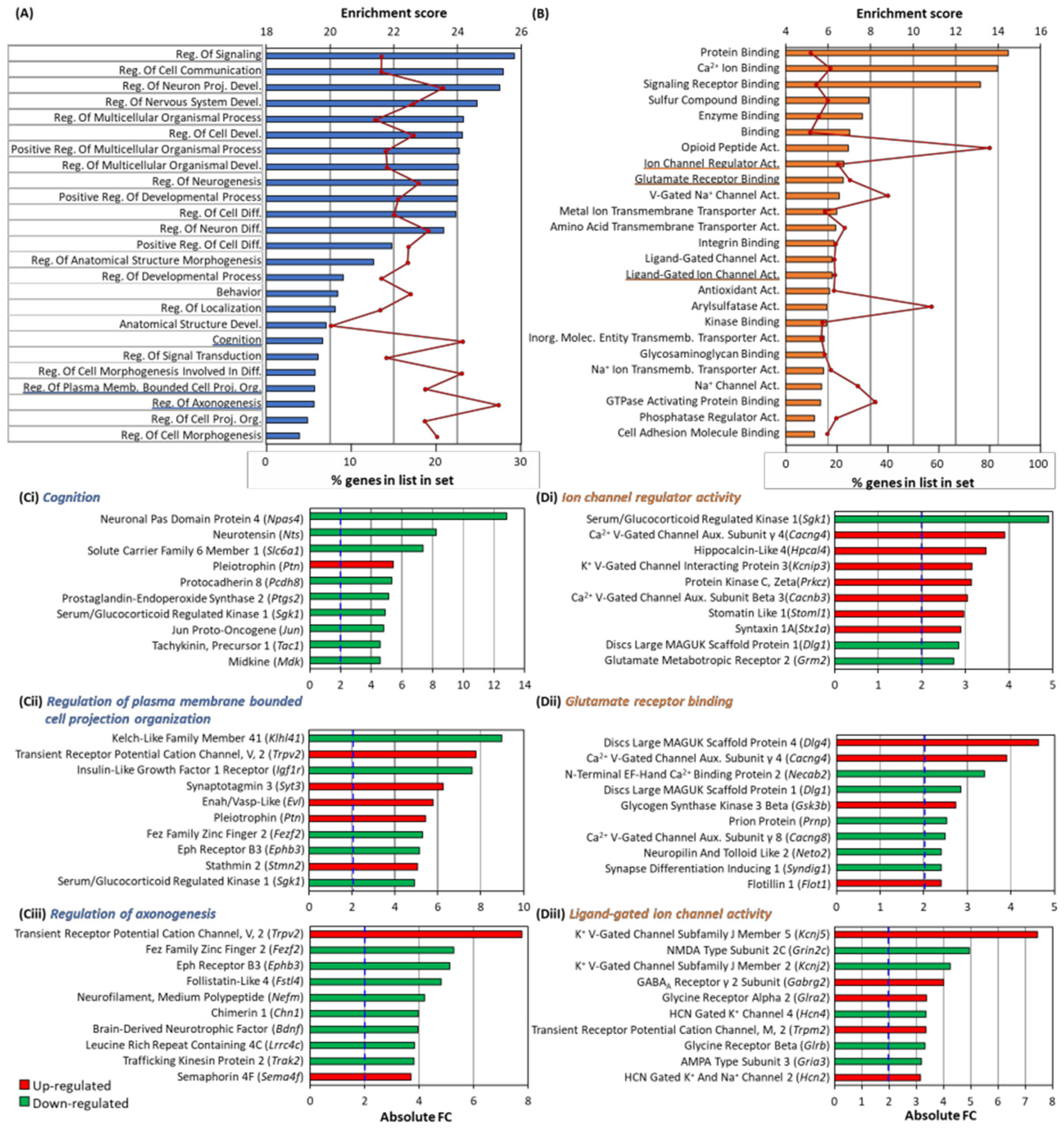
3.6. GO Analysis of Differentially Expressed Genes
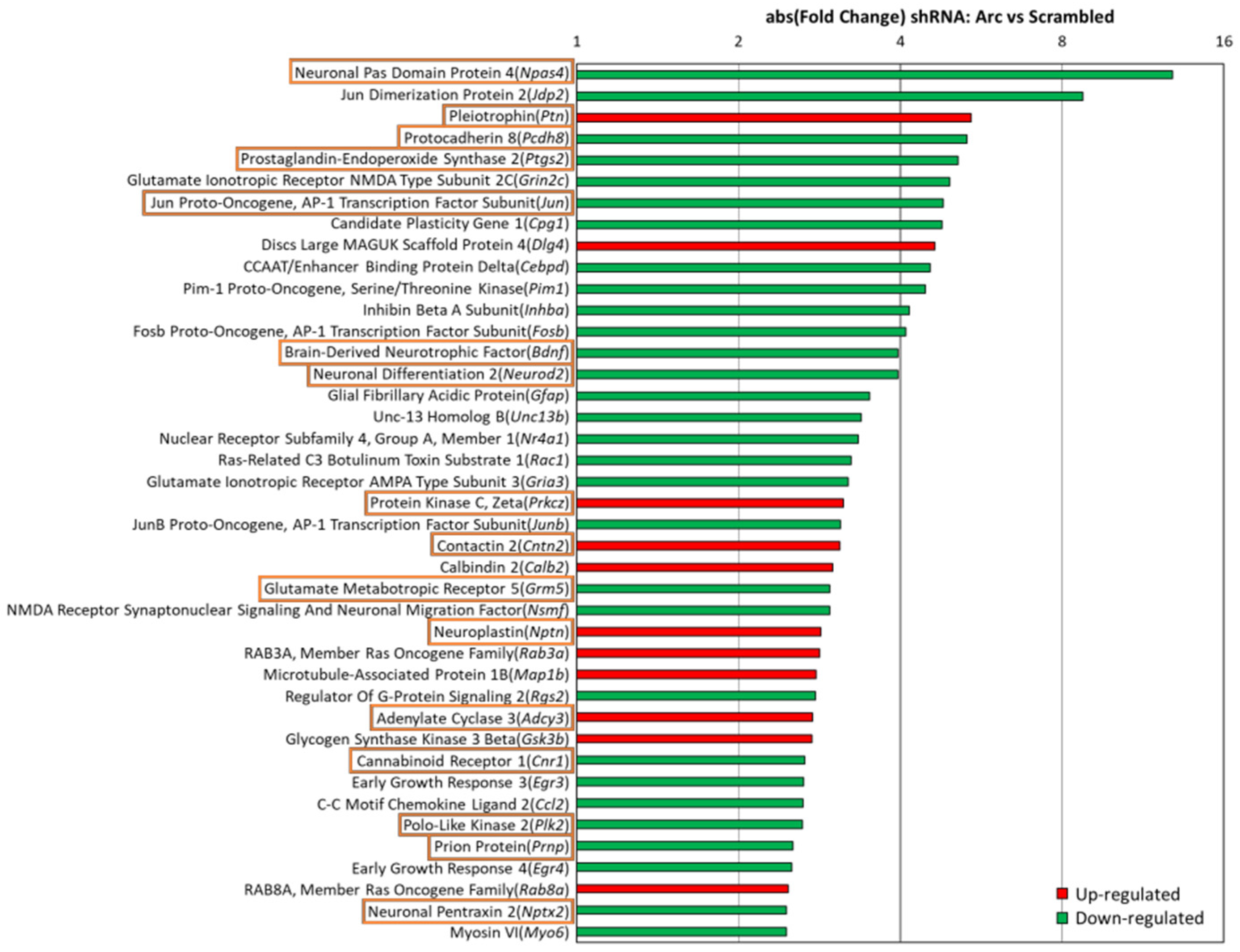
3.7. Arc Regulates Expression of Synaptic and Plasticity Genes
3.8. Arc Knockdown Altered Synaptogenesis, Synaptic Plasticity and Neuroinflammation Pathways
3.9. Arc Knockdown Changes the Expression of Alzheimer’s Disease Genes
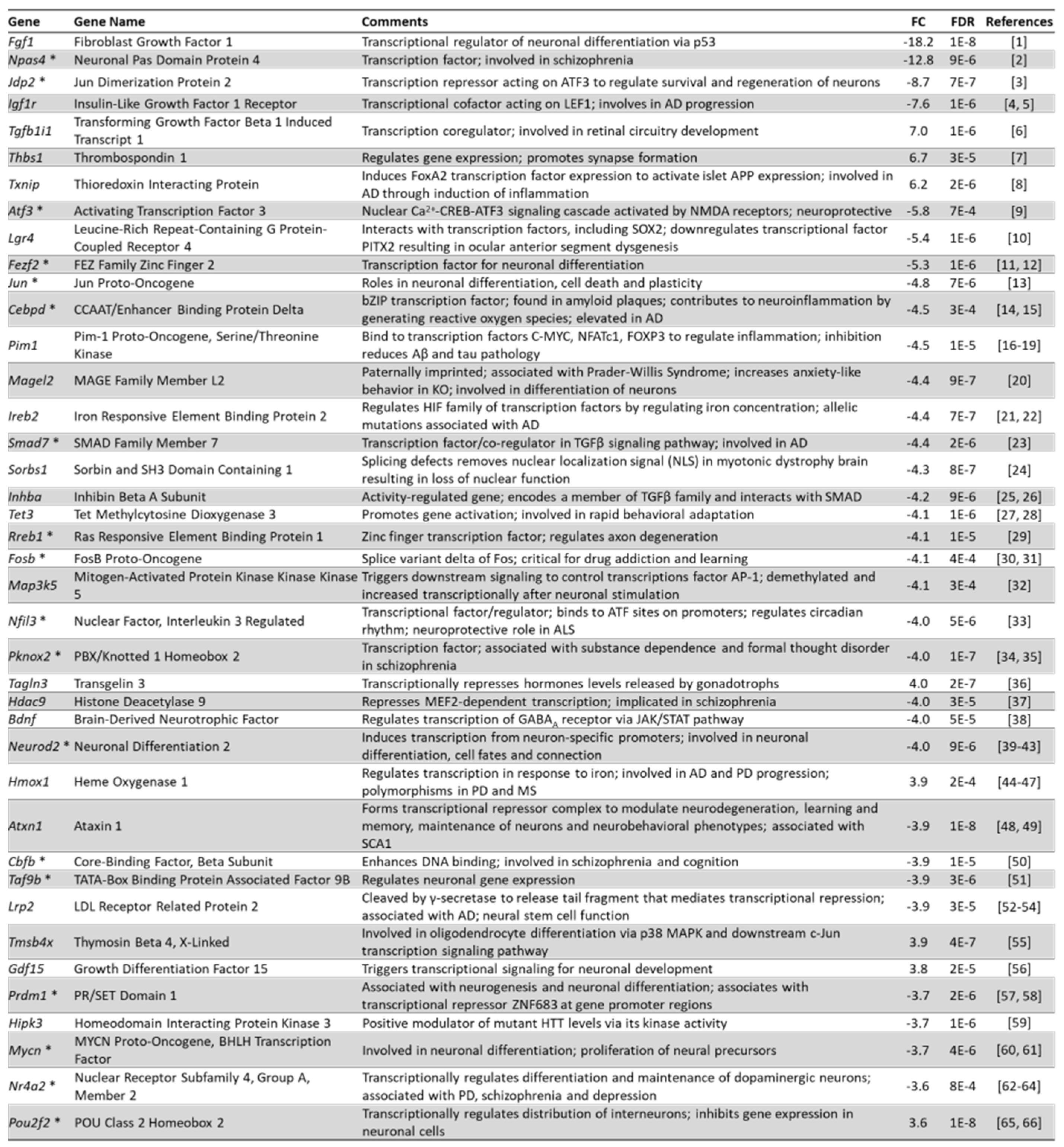
3.10. Arc Regulates the Expression of Transcription Factors
3.11. Upstream Regulators Associated with Arc-Dependent Genes
3.12. Arc Over-Expression Alters Gene Expression in Human Embryonic Kidney Cells
4. Discussion
4.1. Arc and Chromatin
4.2. How Does Arc Regulate Transcription?
4.3. Interactions among TIP60, NOTCH1 and APP
4.4. Arc’s Subcellular Localisation Determines Its Function
4.5. Arc Controls Synaptic Plasticity and Intrinsic Excitability
4.6. Arc and Alzheimer’s Disease
4.7. Arc and Ad Therapy
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lyford, G.L.; Yamagata, K.; Kaufmann, W.E.; Barnes, C.A.; Sanders, L.K.; Copeland, N.G.; Gilbert, D.J.; Jenkins, N.A.; Lanahan, A.A.; Worley, P.F. Arc, a growth factor and activity-regulated gene, encodes a novel cytoskeleton-associated protein that is enriched in neuronal dendrites. Neuron 1995, 14, 433–445. [Google Scholar] [CrossRef] [Green Version]
- Link, W.; Konietzko, U.; Kauselmann, G.; Krug, M.; Schwanke, B.; Frey, U.; Kuhl, D. Somatodendritic expression of an immediate early gene is regulated by synaptic activity. Proc. Natl. Acad. Sci. USA 1995, 92, 5734–5738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guzowski, J.F.; Lyford, G.L.; Stevenson, G.D.; Houston, F.P.; McGaugh, J.L.; Worley, P.F.; Barnes, C.A. Inhibition of activity-dependent Arc protein expression in the rat hippocampus impairs the maintenance of long-term potentiation and the consolidation of long-term memory. J. Neurosci. 2000, 20, 3993–4001. [Google Scholar] [CrossRef] [PubMed]
- Plath, N.; Ohana, O.; Dammermann, B.; Errington, M.L.; Schmitz, D.; Gross, C.; Mao, X.; Engelsberg, A.; Mahlke, C.; Welzl, H.; et al. Arc/Arg3.1 is essential for the consolidation of synaptic plasticity and memories. Neuron 2006, 52, 437–444. [Google Scholar] [CrossRef] [Green Version]
- Ploski, J.E.; Pierre, V.J.; Smucny, J.; Park, K.; Monsey, M.S.; Overeem, K.A.; Schafe, G.E. The activity-regulated cytoskeletal-associated protein (Arc/Arg3.1) is required for memory consolidation of pavlovian fear conditioning in the lateral amygdala. J. Neurosci. 2008, 28, 12383–12395. [Google Scholar] [CrossRef] [Green Version]
- Maddox, S.A.; Schafe, G.E. The activity-regulated cytoskeletal-associated protein (Arc/Arg3.1) is required for reconsolidation of a Pavlovian fear memory. J. Neurosci. 2011, 31, 7073–7082. [Google Scholar] [CrossRef]
- Guzowski, J.F.; McNaughton, B.L.; Barnes, C.A.; Worley, P.F. Environment-specific expression of the immediate-early gene Arc in hippocampal neuronal ensembles. Nat. Neurosci. 1999, 2, 1120–1124. [Google Scholar] [CrossRef]
- Guzowski, J.F.; Setlow, B.; Wagner, E.K.; McGaugh, J.L. Experience-dependent gene expression in the rat hippocampus after spatial learning: A comparison of the immediate-early genes Arc, c-fos, and zif268. J. Neurosci. 2001, 21, 5089–5098. [Google Scholar] [CrossRef] [Green Version]
- Chawla, M.K.; Guzowski, J.F.; Ramirez-Amaya, V.; Lipa, P.; Hoffman, K.L.; Marriott, L.K.; Worley, P.F.; McNaughton, B.L.; Barnes, C.A. Sparse, environmentally selective expression of Arc RNA in the upper blade of the rodent fascia dentata by brief spatial experience. Hippocampus 2005, 15, 579–586. [Google Scholar] [CrossRef]
- Ramirez-Amaya, V.; Vazdarjanova, A.; Mikhael, D.; Rosi, S.; Worley, P.F.; Barnes, C.A. Spatial exploration-induced Arc mRNA and protein expression: Evidence for selective, network-specific reactivation. J. Neurosci. 2005, 25, 1761–1768. [Google Scholar] [CrossRef] [Green Version]
- Vazdarjanova, A.; Ramirez-Amaya, V.; Insel, N.; Plummer, T.K.; Rosi, S.; Chowdhury, S.; Mikhael, D.; Worley, P.F.; Guzowski, J.F.; Barnes, C.A. Spatial exploration induces ARC, a plasticity-related immediate-early gene, only in calcium/calmodulin-dependent protein kinase II-positive principal excitatory and inhibitory neurons of the rat forebrain. J. Comp. Neurol. 2006, 498, 317–329. [Google Scholar] [CrossRef] [PubMed]
- Steward, O.; Wallace, C.S.; Lyford, G.L.; Worley, P.F. Synaptic activation causes the mRNA for the IEG Arc to localize selectively near activated postsynaptic sites on dendrites. Neuron 1998, 21, 741–751. [Google Scholar] [CrossRef] [Green Version]
- Steward, O.; Worley, P.F. Selective targeting of newly synthesized Arc mRNA to active synapses requires NMDA receptor activation. Neuron 2001, 30, 227–240. [Google Scholar] [CrossRef] [Green Version]
- Chowdhury, S.; Shepherd, J.D.; Okuno, H.; Lyford, G.; Petralia, R.S.; Plath, N.; Kuhl, D.; Huganir, R.L.; Worley, P.F. Arc/Arg3.1 interacts with the endocytic machinery to regulate AMPA receptor trafficking. Neuron 2006, 52, 445–459. [Google Scholar] [CrossRef] [Green Version]
- Shepherd, J.D.; Bear, M.F. New views of Arc, a master regulator of synaptic plasticity. Nat. Neurosci. 2011, 14, 279–284. [Google Scholar] [CrossRef]
- Okuno, H.; Minatohara, K.; Bito, H. Inverse synaptic tagging: An inactive synapse-specific mechanism to capture activity-induced Arc/arg3.1 and to locally regulate spatial distribution of synaptic weights. Semin. Cell Dev. Biol. 2018, 77, 43–50. [Google Scholar] [CrossRef]
- Shepherd, J.D.; Rumbaugh, G.; Wu, J.; Chowdhury, S.; Plath, N.; Kuhl, D.; Huganir, R.L.; Worley, P.F. Arc/Arg3.1 mediates homeostatic synaptic scaling of AMPA receptors. Neuron 2006, 52, 475–484. [Google Scholar] [CrossRef] [Green Version]
- Gao, M.; Sossa, K.; Song, L.; Errington, L.; Cummings, L.; Hwang, H.; Kuhl, D.; Worley, P.; Lee, H.K. A specific requirement of Arc/Arg3.1 for visual experience-induced homeostatic synaptic plasticity in mouse primary visual cortex. J. Neurosci. 2010, 30, 7168–7178. [Google Scholar] [CrossRef] [Green Version]
- Beique, J.C.; Na, Y.; Kuhl, D.; Worley, P.F.; Huganir, R.L. Arc-dependent synapse-specific homeostatic plasticity. Proc. Natl. Acad. Sci. USA 2011, 108, 816–821. [Google Scholar] [CrossRef] [Green Version]
- Buffington, S.A.; Huang, W.; Costa-Mattioli, M. Translational control in synaptic plasticity and cognitive dysfunction. Annu. Rev. Neurosci. 2014, 37, 17–38. [Google Scholar] [CrossRef] [Green Version]
- Collingridge, G.L.; Abraham, W.C. Glutamate receptors and synaptic plasticity: The impact of Evans and Watkins. Neuropharmacology 2022, 206, 108922. [Google Scholar] [CrossRef] [PubMed]
- Bloomer, W.A.; VanDongen, H.M.; VanDongen, A.M. Activity-regulated cytoskeleton-associated protein Arc/Arg3.1 binds to spectrin and associates with nuclear promyelocytic leukemia (PML) bodies. Brain Res. 2007, 1153, 20–33. [Google Scholar] [CrossRef] [PubMed]
- Bloomer, W.A.; VanDongen, H.M.; VanDongen, A.M. Arc/Arg3.1 translation is controlled by convergent N-methyl-D-aspartate and Gs-coupled receptor signaling pathways. J. Biol. Chem. 2008, 283, 582–592. [Google Scholar] [CrossRef] [Green Version]
- Oey, N.E.; Leung, H.W.; Ezhilarasan, R.; Zhou, L.; Beuerman, R.W.; VanDongen, H.M.A.; VanDongen, A.M.J. A Neuronal Activity-Dependent Dual Function Chromatin-Modifying Complex Regulates Arc Expression. eNeuro 2015, 2, ENEURO.0020-0014.2015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hedde, P.N.; Barylko, B.; Binns, D.D.; Jameson, D.M.; Albanesi, J.P. Differential Mobility and Self-Association of Arc/Arg3.1 in the Cytoplasm and Nucleus of Living Cells. ACS Chem. Neurosci. 2022, 13, 876–882. [Google Scholar] [CrossRef] [PubMed]
- Torok, D.; Ching, R.W.; Bazett-Jones, D.P. PML nuclear bodies as sites of epigenetic regulation. Front. Biosci. 2009, 14, 1325–1336. [Google Scholar] [CrossRef] [PubMed]
- Voglis, G.; Tavernarakis, N. The role of synaptic ion channels in synaptic plasticity. EMBO Rep. 2006, 7, 1104–1110. [Google Scholar] [CrossRef] [Green Version]
- Korb, E.; Wilkinson, C.L.; Delgado, R.N.; Lovero, K.L.; Finkbeiner, S. Arc in the nucleus regulates PML-dependent GluA1 transcription and homeostatic plasticity. Nat. Neurosci. 2013, 16, 874–883. [Google Scholar] [CrossRef] [Green Version]
- Wee, C.L.; Teo, S.; Oey, N.E.; Wright, G.D.; VanDongen, H.M.; VanDongen, A.M. Nuclear Arc Interacts with the Histone Acetyltransferase Tip60 to Modify H4K12 Acetylation. eNeuro 2014, 1, ENEURO.0019-14.2014. [Google Scholar] [CrossRef] [Green Version]
- Qi, D.; Jin, H.; Lilja, T.; Mannervik, M. Drosophila Reptin and other TIP60 complex components promote generation of silent chromatin. Genetics 2006, 174, 241–251. [Google Scholar] [CrossRef] [Green Version]
- Tea, J.S.; Luo, L. The chromatin remodeling factor Bap55 functions through the TIP60 complex to regulate olfactory projection neuron dendrite targeting. Neural Dev. 2011, 6, 5. [Google Scholar] [CrossRef] [Green Version]
- Rust, K.; Tiwari, M.D.; Mishra, V.K.; Grawe, F.; Wodarz, A. Myc and the Tip60 chromatin remodeling complex control neuroblast maintenance and polarity in Drosophila. EMBO J. 2018, 37, e98659. [Google Scholar] [CrossRef]
- Peleg, S.; Sananbenesi, F.; Zovoilis, A.; Burkhardt, S.; Bahari-Javan, S.; Agis-Balboa, R.C.; Cota, P.; Wittnam, J.L.; Gogol-Doering, A.; Opitz, L.; et al. Altered histone acetylation is associated with age-dependent memory impairment in mice. Science 2010, 328, 753–756. [Google Scholar] [CrossRef] [Green Version]
- Plagg, B.; Ehrlich, D.; Kniewallner, K.M.; Marksteiner, J.; Humpel, C. Increased Acetylation of Histone H4 at Lysine 12 (H4K12) in Monocytes of Transgenic ad Mice and in Human Patients. Curr. Alzheimer Res. 2015, 12, 752–760. [Google Scholar] [CrossRef]
- Hardingham, G.E.; Bading, H. The Yin and Yang of NMDA receptor signalling. Trends Neurosci. 2003, 26, 81–89. [Google Scholar] [CrossRef]
- Vanhoutte, P.; Bading, H. Opposing roles of synaptic and extrasynaptic NMDA receptors in neuronal calcium signalling and BDNF gene regulation. Curr. Opin. Neurobiol. 2003, 13, 366–371. [Google Scholar] [CrossRef]
- Hardingham, G.E.; Arnold, F.J.; Bading, H. Nuclear calcium signaling controls CREB-mediated gene expression triggered by synaptic activity. Nat. Neurosci. 2001, 4, 261–267. [Google Scholar] [CrossRef]
- Otmakhov, N.; Khibnik, L.; Otmakhova, N.; Carpenter, S.; Riahi, S.; Asrican, B.; Lisman, J. Forskolin-induced LTP in the CA1 hippocampal region is NMDA receptor dependent. J. Neurophysiol. 2004, 91, 1955–1962. [Google Scholar] [CrossRef]
- Gouty-Colomer, L.A.; Hosseini, B.; Marcelo, I.M.; Schreiber, J.; Slump, D.E.; Yamaguchi, S.; Houweling, A.R.; Jaarsma, D.; Elgersma, Y.; Kushner, S.A. Arc expression identifies the lateral amygdala fear memory trace. Mol. Psychiatry 2016, 21, 1153. [Google Scholar] [CrossRef] [Green Version]
- Minatohara, K.; Akiyoshi, M.; Okuno, H. Role of Immediate-Early Genes in Synaptic Plasticity and Neuronal Ensembles Underlying the Memory Trace. Front. Mol. Neurosci. 2015, 8, 78. [Google Scholar] [CrossRef] [Green Version]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Xing, Y.; Yu, T.; Wu, Y.N.; Roy, M.; Kim, J.; Lee, C. An expectation-maximization algorithm for probabilistic reconstructions of full-length isoforms from splice graphs. Nucleic Acids Res. 2006, 34, 3150–3160. [Google Scholar] [CrossRef] [Green Version]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef]
- Benjamini, Y.; Drai, D.; Elmer, G.; Kafkafi, N.; Golani, I. Controlling the false discovery rate in behavior genetics research. Behav. Brain Res. 2001, 125, 279–284. [Google Scholar] [CrossRef] [Green Version]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene ontology: Tool for the unification of biology. The Gene Ontology Consortium. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [Green Version]
- The Gene Ontology, C. The Gene Ontology Resource: 20 years and still GOing strong. Nucleic Acids Res. 2019, 47, D330–D338. [Google Scholar]
- Koch, C.M.; Chiu, S.F.; Akbarpour, M.; Bharat, A.; Ridge, K.M.; Bartom, E.T.; Winter, D.R. A Beginner’s Guide to Analysis of RNA Sequencing Data. Am. J. Respir. Cell Mol. Biol. 2018, 59, 145–157. [Google Scholar] [CrossRef]
- Huang da, W.; Sherman, B.T.; Lempicki, R.A. Bioinformatics enrichment tools: Paths toward the comprehensive functional analysis of large gene lists. Nucleic Acids Res. 2009, 37, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Huang da, W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef]
- Park, A.Y.; Park, Y.S.; So, D.; Song, I.K.; Choi, J.E.; Kim, H.J.; Lee, K.J. Activity-Regulated Cytoskeleton-Associated Protein (Arc/Arg3.1) is Transiently Expressed after Heat Shock Stress and Suppresses Heat Shock Factor 1. Sci. Rep. 2019, 9, 2592. [Google Scholar] [CrossRef] [Green Version]
- Salery, M.; Dos Santos, M.; Saint-Jour, E.; Moumne, L.; Pages, C.; Kappes, V.; Parnaudeau, S.; Caboche, J.; Vanhoutte, P. Activity-Regulated Cytoskeleton-Associated Protein Accumulates in the Nucleus in Response to Cocaine and Acts as a Brake on Chromatin Remodeling and Long-Term Behavioral Alterations. Biol. Psychiatry 2017, 81, 573–584. [Google Scholar] [CrossRef] [Green Version]
- Duvarci, S.; Nader, K.; LeDoux, J.E. De novo mRNA synthesis is required for both consolidation and reconsolidation of fear memories in the amygdala. Learn. Mem. 2008, 15, 747–755. [Google Scholar] [CrossRef] [Green Version]
- Pereira, L.M.; de Castro, C.M.; Guerra, L.T.L.; Queiroz, T.M.; Marques, J.T.; Pereira, G.S. Hippocampus and Prefrontal Cortex Modulation of Contextual Fear Memory Is Dissociated by Inhibiting De Novo Transcription During Late Consolidation. Mol. Neurobiol. 2019, 56, 5507–5519. [Google Scholar] [CrossRef]
- Deisseroth, K.; Bito, H.; Tsien, R.W. Signaling from synapse to nucleus: Postsynaptic CREB phosphorylation during multiple forms of hippocampal synaptic plasticity. Neuron 1996, 16, 89–101. [Google Scholar] [CrossRef] [Green Version]
- Thompson, K.R.; Otis, K.O.; Chen, D.Y.; Zhao, Y.; O’Dell, T.J.; Martin, K.C. Synapse to nucleus signaling during long-term synaptic plasticity; a role for the classical active nuclear import pathway. Neuron 2004, 44, 997–1009. [Google Scholar]
- Dieterich, D.C.; Karpova, A.; Mikhaylova, M.; Zdobnova, I.; Konig, I.; Landwehr, M.; Kreutz, M.; Smalla, K.H.; Richter, K.; Landgraf, P.; et al. Caldendrin-Jacob: A protein liaison that couples NMDA receptor signalling to the nucleus. PLoS Biol. 2008, 6, e34. [Google Scholar] [CrossRef] [Green Version]
- Kawashima, T.; Okuno, H.; Nonaka, M.; Adachi-Morishima, A.; Kyo, N.; Okamura, M.; Takemoto-Kimura, S.; Worley, P.F.; Bito, H. Synaptic activity-responsive element in the Arc/Arg3.1 promoter essential for synapse-to-nucleus signaling in activated neurons. Proc. Natl. Acad. Sci. USA 2009, 106, 316–321. [Google Scholar] [CrossRef] [Green Version]
- Bading, H. Nuclear calcium signalling in the regulation of brain function. Nat. Rev. Neurosci. 2013, 14, 593–608. [Google Scholar] [CrossRef]
- Tweedie-Cullen, R.Y.; Reck, J.M.; Mansuy, I.M. Comprehensive mapping of post-translational modifications on synaptic, nuclear, and histone proteins in the adult mouse brain. J. Proteome Res. 2009, 8, 4966–4982. [Google Scholar] [CrossRef]
- Kim, S.; Kaang, B.K. Epigenetic regulation and chromatin remodeling in learning and memory. Exp. Mol. Med. 2017, 49, e281. [Google Scholar] [CrossRef] [Green Version]
- Almassalha, L.M.; Tiwari, A.; Ruhoff, P.T.; Stypula-Cyrus, Y.; Cherkezyan, L.; Matsuda, H.; Dela Cruz, M.A.; Chandler, J.E.; White, C.; Maneval, C.; et al. The Global Relationship between Chromatin Physical Topology, Fractal Structure, and Gene Expression. Sci. Rep. 2017, 7, 41061. [Google Scholar] [CrossRef] [Green Version]
- Gottesfeld, J.M.; Carey, M.F. Introduction to the Thematic Minireview Series: Chromatin and transcription. J. Biol. Chem. 2018, 293, 13775–13777. [Google Scholar] [CrossRef]
- Levenson, J.M.; O’Riordan, K.J.; Brown, K.D.; Trinh, M.A.; Molfese, D.L.; Sweatt, J.D. Regulation of histone acetylation during memory formation in the hippocampus. J. Biol. Chem. 2004, 279, 40545–40559. [Google Scholar] [CrossRef] [Green Version]
- Korzus, E.; Rosenfeld, M.G.; Mayford, M. CBP histone acetyltransferase activity is a critical component of memory consolidation. Neuron 2004, 42, 961–972. [Google Scholar] [CrossRef] [Green Version]
- Wood, M.A.; Hawk, J.D.; Abel, T. Combinatorial chromatin modifications and memory storage: A code for memory? Learn. Mem. 2006, 13, 241–244. [Google Scholar] [CrossRef] [Green Version]
- Graff, J.; Tsai, L.H. Histone acetylation: Molecular mnemonics on the chromatin. Nat. Rev. Neurosci. 2013, 14, 97–111. [Google Scholar] [CrossRef]
- Fischer, A. Epigenetic memory: The Lamarckian brain. EMBO J. 2014, 33, 945–967. [Google Scholar] [CrossRef]
- Zovkic, I.B.; Paulukaitis, B.S.; Day, J.J.; Etikala, D.M.; Sweatt, J.D. Histone H2A.Z subunit exchange controls consolidation of recent and remote memory. Nature 2014, 515, 582–586. [Google Scholar] [CrossRef] [Green Version]
- McNally, A.G.; Poplawski, S.G.; Mayweather, B.A.; White, K.M.; Abel, T. Characterization of a Novel Chromatin Sorting Tool Reveals Importance of Histone Variant H3.3 in Contextual Fear Memory and Motor Learning. Front. Mol. Neurosci. 2016, 9, 11. [Google Scholar] [CrossRef] [Green Version]
- Collins, B.E.; Greer, C.B.; Coleman, B.C.; Sweatt, J.D. Histone H3 lysine K4 methylation and its role in learning and memory. Epigenetics Chromatin 2019, 12, 7. [Google Scholar] [CrossRef]
- Deng, W.; Blobel, G.A. Do chromatin loops provide epigenetic gene expression states? Curr. Opin. Genet. Dev. 2010, 20, 548–554. [Google Scholar] [CrossRef] [Green Version]
- Gondor, A. Dynamic chromatin loops bridge health and disease in the nuclear landscape. Semin. Cancer Biol. 2013, 23, 90–98. [Google Scholar] [CrossRef]
- Hansen, A.S.; Cattoglio, C.; Darzacq, X.; Tjian, R. Recent evidence that TADs and chromatin loops are dynamic structures. Nucleus 2018, 9, 20–32. [Google Scholar] [CrossRef] [Green Version]
- Janssen, K.A.; Sidoli, S.; Garcia, B.A. Recent Achievements in Characterizing the Histone Code and Approaches to Integrating Epigenomics and Systems Biology. Methods Enzymol. 2017, 586, 359–378. [Google Scholar]
- Creyghton, M.P.; Cheng, A.W.; Welstead, G.G.; Kooistra, T.; Carey, B.W.; Steine, E.J.; Hanna, J.; Lodato, M.A.; Frampton, G.M.; Sharp, P.A.; et al. Histone H3K27ac separates active from poised enhancers and predicts developmental state. Proc. Natl. Acad. Sci. USA 2010, 107, 21931–21936. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Day, D.S.; Ho, J.W.; Song, L.; Cao, J.; Christodoulou, D.; Seidman, J.G.; Crawford, G.E.; Park, P.J.; Pu, W.T. A dynamic H3K27ac signature identifies VEGFA-stimulated endothelial enhancers and requires EP300 activity. Genome Res. 2013, 23, 917–927. [Google Scholar] [CrossRef] [Green Version]
- Deb, M.; Kar, S.; Sengupta, D.; Shilpi, A.; Parbin, S.; Rath, S.K.; Londhe, V.A.; Patra, S.K. Chromatin dynamics: H3K4 methylation and H3 variant replacement during development and in cancer. Cell Mol. Life Sci. 2014, 71, 3439–3463. [Google Scholar] [CrossRef]
- Esnault, C.; Gualdrini, F.; Horswell, S.; Kelly, G.; Stewart, A.; East, P.; Matthews, N.; Treisman, R. ERK-Induced Activation of TCF Family of SRF Cofactors Initiates a Chromatin Modification Cascade Associated with Transcription. Mol. Cell 2017, 65, 1081–1095.e1085. [Google Scholar] [CrossRef] [Green Version]
- Rust, M.J.; Bates, M.; Zhuang, X. Sub-diffraction-limit imaging by stochastic optical reconstruction microscopy (STORM). Nat. Methods 2006, 3, 793–795. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, S.; Brickner, J.H. Regulation and epigenetic control of transcription at the nuclear periphery. Trends Genet. 2007, 23, 396–402. [Google Scholar] [CrossRef]
- Kalverda, B.; Röling, M.D.; Fornerod, M. Chromatin organization in relation to the nuclear periphery. FEBS Lett. 2008, 582, 2017–2022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, Y.T.; Kazui, H.; Ikeda, M.; Huang, C.W.; Huang, S.H.; Hsu, S.W.; Chang, W.N.; Chang, C.C. Genetic Interaction of APOE and FGF1 is Associated with Memory Impairment and Hippocampal Atrophy in Alzheimer’s Disease. Aging Dis. 2019, 10, 510–519. [Google Scholar] [CrossRef] [Green Version]
- Tao, Q.Q.; Sun, Y.M.; Liu, Z.J.; Ni, W.; Yang, P.; Li, H.L.; Lu, S.J.; Wu, Z.Y. A variant within FGF1 is associated with Alzheimer’s disease in the Han Chinese population. Am. J. Med. Genet. B Neuropsychiatr Genet. 2014, 165B, 131–136. [Google Scholar] [CrossRef] [PubMed]
- Yamagata, H.; Chen, Y.; Akatsu, H.; Kamino, K.; Ito, J.; Yokoyama, S.; Yamamoto, T.; Kosaka, K.; Miki, T.; Kondo, I. Promoter polymorphism in fibroblast growth factor 1 gene increases risk of definite Alzheimer’s disease. Biochem. Biophys. Res. Commun. 2004, 321, 320–323. [Google Scholar] [CrossRef] [PubMed]
- Alazami, A.M.; Patel, N.; Shamseldin, H.E.; Anazi, S.; Al-Dosari, M.S.; Alzahrani, F.; Hijazi, H.; Alshammari, M.; Aldahmesh, M.A.; Salih, M.A.; et al. Accelerating novel candidate gene discovery in neurogenetic disorders via whole-exome sequencing of prescreened multiplex consanguineous families. Cell Rep. 2015, 10, 148–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lyubartseva, G.; Smith, J.L.; Markesbery, W.R.; Lovell, M.A. Alterations of zinc transporter proteins ZnT-1, ZnT-4 and ZnT-6 in preclinical Alzheimer’s disease brain. Brain Pathol. 2010, 20, 343–350. [Google Scholar] [CrossRef] [Green Version]
- Fan, W.; Long, Y.; Lai, Y.; Wang, X.; Chen, G.; Zhu, B. NPAS4 Facilitates the Autophagic Clearance of Endogenous Tau in Rat Cortical Neurons. J. Mol. Neurosci. 2016, 58, 401–410. [Google Scholar] [CrossRef]
- Miyashita, A.; Hatsuta, H.; Kikuchi, M.; Nakaya, A.; Saito, Y.; Tsukie, T.; Hara, N.; Ogishima, S.; Kitamura, N.; Akazawa, K.; et al. Genes associated with the progression of neurofibrillary tangles in Alzheimer’s disease. Transl. Psychiatry 2014, 4, e396. [Google Scholar] [CrossRef] [Green Version]
- Ramamoorthi, K.; Fropf, R.; Belfort, G.M.; Fitzmaurice, H.L.; McKinney, R.M.; Neve, R.L.; Otto, T.; Lin, Y. Npas4 regulates a transcriptional program in CA3 required for contextual memory formation. Science 2011, 334, 1669–1675. [Google Scholar] [CrossRef] [Green Version]
- Ito, S.; Kimura, K.; Haneda, M.; Ishida, Y.; Sawada, M.; Isobe, K. Induction of matrix metalloproteinases (MMP3, MMP12 and MMP13) expression in the microglia by amyloid-beta stimulation via the PI3K/Akt pathway. Exp. Gerontol. 2007, 42, 532–537. [Google Scholar] [CrossRef]
- Zhu, B.L.; Long, Y.; Luo, W.; Yan, Z.; Lai, Y.J.; Zhao, L.G.; Zhou, W.H.; Wang, Y.J.; Shen, L.L.; Liu, L.; et al. MMP13 inhibition rescues cognitive decline in Alzheimer transgenic mice via BACE1 regulation. Brain 2019, 142, 176–192. [Google Scholar] [CrossRef] [PubMed]
- De Souza Silva, M.A.; Lenz, B.; Rotter, A.; Biermann, T.; Peters, O.; Ramirez, A.; Jessen, F.; Maier, W.; Hull, M.; Schroder, J.; et al. Neurokinin3 receptor as a target to predict and improve learning and memory in the aged organism. Proc. Natl. Acad. Sci. USA 2013, 110, 15097–15102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foroud, T.; Wetherill, L.F.; Kramer, J.; Tischfield, J.A.; Nurnberger, J.I., Jr.; Schuckit, M.A.; Xuei, X.; Edenberg, H.J. The tachykinin receptor 3 is associated with alcohol and cocaine dependence. Alcohol. Clin. Exp. Res. 2008, 32, 1023–1030. [Google Scholar] [CrossRef] [PubMed]
- Teipel, S.J.; Grothe, M.J.; Wittfeld, K.; Hoffmann, W.; Hegenscheid, K.; Volzke, H.; Homuth, G.; Grabe, H.J. Association of a neurokinin 3 receptor polymorphism with the anterior basal forebrain. Neurobiol. Aging 2015, 36, 2060–2067. [Google Scholar] [CrossRef] [PubMed]
- Semerdjieva, S.; Abdul-Razak, H.H.; Salim, S.S.; Yanez-Munoz, R.J.; Chen, P.E.; Tarabykin, V.; Alifragis, P. Activation of EphA receptors mediates the recruitment of the adaptor protein Slap, contributing to the downregulation of N-methyl-D-aspartate receptors. Mol. Cell Biol. 2013, 33, 1442–1455. [Google Scholar] [CrossRef] [Green Version]
- Yaman, E.; Gasper, R.; Koerner, C.; Wittinghofer, A.; Tazebay, U.H. RasGEF1A and RasGEF1B are guanine nucleotide exchange factors that discriminate between Rap GTP-binding proteins and mediate Rap2-specific nucleotide exchange. FEBS J. 2009, 276, 4607–4616. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, M.; Hao, W.; Mihaljevic, I.; Liu, X.; Xie, K.; Walter, S.; Fassbender, K. Matrix metalloproteinase-12 contributes to neuroinflammation in the aged brain. Neurobiol. Aging 2013, 34, 1231–1239. [Google Scholar] [CrossRef]
- Teranishi, Y.; Inoue, M.; Yamamoto, N.G.; Kihara, T.; Wiehager, B.; Ishikawa, T.; Winblad, B.; Schedin-Weiss, S.; Frykman, S.; Tjernberg, L.O. Proton myo-inositol cotransporter is a novel gamma-secretase associated protein that regulates Abeta production without affecting Notch cleavage. FEBS J. 2015, 282, 3438–3451. [Google Scholar] [CrossRef]
- Wang, J.G.; Strong, J.A.; Xie, W.; Yang, R.H.; Coyle, D.E.; Wick, D.M.; Dorsey, E.D.; Zhang, J.M. The chemokine CXCL1/growth related oncogene increases sodium currents and neuronal excitability in small diameter sensory neurons. Mol. Pain 2008, 4, 38. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.; Zhao, Y.; Sun, X.-H. Notch-Regulated Periphery B Cell Differentiation Involves Suppression of E Protein Function. J. Immunol. 2013, 191, 726–736. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.F.; Zhao, Y.F.; Zhu, S.W.; Huang, W.J.; Luo, Y.; Chen, Q.Y.; Ge, L.J.; Li, R.S.; Wang, J.F.; Sun, M.; et al. CXCL1 Triggers Caspase-3 Dependent Tau Cleavage in Long-Term Neuronal Cultures and in the Hippocampus of Aged Mice: Implications in Alzheimer’s Disease. J. Alzheimers Dis. 2015, 48, 89–104. [Google Scholar] [CrossRef] [PubMed]
- Davis, W., Jr. The ATP-Binding Cassette Transporter-2 (ABCA2) Overexpression Modulates Sphingosine Levels and Transcription of the Amyloid Precursor Protein (APP) Gene. Curr. Alzheimer Res. 2015, 12, 847–859. [Google Scholar] [CrossRef]
- Guedea, A.L.; Schrick, C.; Guzman, Y.F.; Leaderbrand, K.; Jovasevic, V.; Corcoran, K.A.; Tronson, N.C.; Radulovic, J. ERK-associated changes of AP-1 proteins during fear extinction. Mol. Cell Neurosci. 2011, 47, 137–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorbacheva, L.; Davidova, O.; Sokolova, E.; Ishiwata, S.; Pinelis, V.; Strukova, S.; Reiser, G. Endothelial protein C receptor is expressed in rat cortical and hippocampal neurons and is necessary for protective effect of activated protein C at glutamate excitotoxicity. J. Neurochem. 2009, 111, 967–975. [Google Scholar] [CrossRef]
- Guo, H.; Liu, D.; Gelbard, H.; Cheng, T.; Insalaco, R.; Fernandez, J.A.; Griffin, J.H.; Zlokovic, B.V. Activated protein C prevents neuronal apoptosis via protease activated receptors 1 and 3. Neuron 2004, 41, 563–572. [Google Scholar] [CrossRef] [Green Version]
- Gahete, M.D.; Rubio, A.; Cordoba-Chacon, J.; Gracia-Navarro, F.; Kineman, R.D.; Avila, J.; Luque, R.M.; Castano, J.P. Expression of the ghrelin and neurotensin systems is altered in the temporal lobe of Alzheimer’s disease patients. J. Alzheimers Dis. 2010, 22, 819–828. [Google Scholar] [CrossRef]
- Woodworth, H.L.; Batchelor, H.M.; Beekly, B.G.; Bugescu, R.; Brown, J.A.; Kurt, G.; Fuller, P.M.; Leinninger, G.M. Neurotensin Receptor-1 Identifies a Subset of Ventral Tegmental Dopamine Neurons that Coordinates Energy Balance. Cell Rep. 2017, 20, 1881–1892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, Z.; Cilz, N.I.; Kurada, L.; Hu, B.; Yang, C.; Wada, E.; Combs, C.K.; Porter, J.E.; Lesage, F.; Lei, S. Activation of neurotensin receptor 1 facilitates neuronal excitability and spatial learning and memory in the entorhinal cortex: Beneficial actions in an Alzheimer’s disease model. J. Neurosci. 2014, 34, 7027–7042. [Google Scholar] [CrossRef] [Green Version]
- D’Haene, E.; Jacobs, E.Z.; Volders, P.J.; De Meyer, T.; Menten, B.; Vergult, S. Identification of long non-coding RNAs involved in neuronal development and intellectual disability. Sci. Rep. 2016, 6, 28396. [Google Scholar] [CrossRef] [Green Version]
- Katanosaka, K.; Takatsu, S.; Mizumura, K.; Naruse, K.; Katanosaka, Y. TRPV2 is required for mechanical nociception and the stretch-evoked response of primary sensory neurons. Sci. Rep. 2018, 8, 16782. [Google Scholar] [CrossRef]
- Shibasaki, K.; Murayama, N.; Ono, K.; Ishizaki, Y.; Tominaga, M. TRPV2 enhances axon outgrowth through its activation by membrane stretch in developing sensory and motor neurons. J. Neurosci. 2010, 30, 4601–4612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duits, F.H.; Hernandez-Guillamon, M.; Montaner, J.; Goos, J.D.; Montanola, A.; Wattjes, M.P.; Barkhof, F.; Scheltens, P.; Teunissen, C.E.; van der Flier, W.M. Matrix Metalloproteinases in Alzheimer’s Disease and Concurrent Cerebral Microbleeds. J. Alzheimers Dis. 2015, 48, 711–720. [Google Scholar] [CrossRef] [PubMed]
- Lakhan, S.E.; Kirchgessner, A.; Tepper, D.; Leonard, A. Matrix metalloproteinases and blood-brain barrier disruption in acute ischemic stroke. Front. Neurol. 2013, 4, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- George, C.; Gontier, G.; Lacube, P.; Francois, J.C.; Holzenberger, M.; Aid, S. The Alzheimer’s disease transcriptome mimics the neuroprotective signature of IGF-1 receptor-deficient neurons. Brain 2017, 140, 2012–2027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gontier, G.; George, C.; Chaker, Z.; Holzenberger, M.; Aid, S. Blocking IGF Signaling in Adult Neurons Alleviates Alzheimer’s Disease Pathology through Amyloid-beta Clearance. J. Neurosci. 2015, 35, 11500–11513. [Google Scholar] [CrossRef]
- Nieto Guil, A.F.; Oksdath, M.; Weiss, L.A.; Grassi, D.J.; Sosa, L.J.; Nieto, M.; Quiroga, S. IGF-1 receptor regulates dynamic changes in neuronal polarity during cerebral cortical migration. Sci. Rep. 2017, 7, 7703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pristera, A.; Blomeley, C.; Lopes, E.; Threlfell, S.; Merlini, E.; Burdakov, D.; Cragg, S.; Guillemot, F.; Ang, S.L. Dopamine neuron-derived IGF-1 controls dopamine neuron firing, skill learning, and exploration. Proc. Natl. Acad. Sci. USA 2019, 116, 3817–3826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moccia, F.; Zuccolo, E.; Soda, T.; Tanzi, F.; Guerra, G.; Mapelli, L.; Lodola, F.; D’Angelo, E. Stim and Orai proteins in neuronal Ca(2+) signaling and excitability. Front. Cell Neurosci. 2015, 9, 153. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Sun, S.; Wu, L.; Pchitskaya, E.; Zakharova, O.; Fon Tacer, K.; Bezprozvanny, I. Store-Operated Calcium Channel Complex in Postsynaptic Spines: A New Therapeutic Target for Alzheimer’s Disease Treatment. J. Neurosci. 2016, 36, 11837–11850. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Moody, J.P.; Edgerly, C.K.; Bordiuk, O.L.; Cormier, K.; Smith, K.; Beal, M.F.; Ferrante, R.J. Mitochondrial loss, dysfunction and altered dynamics in Huntington’s disease. Hum. Mol. Genet. 2010, 19, 3919–3935. [Google Scholar] [CrossRef]
- Zhang, L.; Guo, X.Q.; Chu, J.F.; Zhang, X.; Yan, Z.R.; Li, Y.Z. Potential hippocampal genes and pathways involved in Alzheimer’s disease: A bioinformatic analysis. Genet. Mol. Res. 2015, 14, 7218–7232. [Google Scholar] [CrossRef] [PubMed]
- Wickman, K.; Karschin, C.; Karschin, A.; Picciotto, M.R.; Clapham, D.E. Brain localization and behavioral impact of the G-protein-gated K+ channel subunit GIRK4. J. Neurosci. 2000, 20, 5608–5615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carvill, G.L.; McMahon, J.M.; Schneider, A.; Zemel, M.; Myers, C.T.; Saykally, J.; Nguyen, J.; Robbiano, A.; Zara, F.; Specchio, N.; et al. Mutations in the GABA Transporter SLC6A1 Cause Epilepsy with Myoclonic-Atonic Seizures. Am. J. Hum. Genet. 2015, 96, 808–815. [Google Scholar] [CrossRef] [Green Version]
- Thoeringer, C.K.; Ripke, S.; Unschuld, P.G.; Lucae, S.; Ising, M.; Bettecken, T.; Uhr, M.; Keck, M.E.; Mueller-Myhsok, B.; Holsboer, F.; et al. The GABA transporter 1 (SLC6A1): A novel candidate gene for anxiety disorders. J. Neural Transm. 2009, 116, 649–657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fromer, M.; Pocklington, A.J.; Kavanagh, D.H.; Williams, H.J.; Dwyer, S.; Gormley, P.; Georgieva, L.; Rees, E.; Palta, P.; Ruderfer, D.M.; et al. De novo mutations in schizophrenia implicate synaptic networks. Nature 2014, 506, 179–184. [Google Scholar] [CrossRef] [Green Version]
- Grzmil, P.; Konietzko, J.; Boehm, D.; Holter, S.M.; Aguilar-Pimentel, A.; Javaheri, A.; Kalaydjiev, S.; Adler, T.; Bolle, I.; Adham, I.; et al. Targeted disruption of the mouse Npal3 gene leads to deficits in behavior, increased IgE levels, and impaired lung function. Cytogenet. Genome Res. 2009, 125, 186–200. [Google Scholar] [CrossRef]
- Huang, R.; Chen, M.; Yang, L.; Wagle, M.; Guo, S.; Hu, B. MicroRNA-133b Negatively Regulates Zebrafish Single Mauthner-Cell Axon Regeneration through Targeting tppp3 in Vivo. Front. Mol. Neurosci. 2017, 10, 375. [Google Scholar] [CrossRef]
- Meyer, M.A. Identification of 17 Highly Expressed Genes within Mouse Lumbar Spinal Cord Anterior Horn Region from an In-Situ Hybridization Atlas of 3430 Genes: Implications for Motor Neuron Disease. Neurol. Int. 2014, 6, 5367. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Xiao, H.; Huang, Z.; Hu, Z.; Qi, T.; Zhang, B.; Tao, X.; Liu, S.H. MicroRNA124 regulate cell growth of prostate cancer cells by targeting iASPP. Int. J. Clin. Exp. Pathol. 2014, 7, 2283–2290. [Google Scholar]
- Carneiro, A.M.; Blakely, R.D. Serotonin-, protein kinase C-, and Hic-5-associated redistribution of the platelet serotonin transporter. J. Biol. Chem. 2006, 281, 24769–24780. [Google Scholar] [CrossRef] [Green Version]
- Carneiro, A.M.; Ingram, S.L.; Beaulieu, J.M.; Sweeney, A.; Amara, S.G.; Thomas, S.M.; Caron, M.G.; Torres, G.E. The multiple LIM domain-containing adaptor protein Hic-5 synaptically colocalizes and interacts with the dopamine transporter. J. Neurosci. 2002, 22, 7045–7054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.; Lim, S.; Ha, T.; Song, Y.H.; Sohn, Y.I.; Park, D.J.; Paik, S.S.; Kim-Kaneyama, J.R.; Song, M.R.; Leung, A.; et al. The LIM protein complex establishes a retinal circuitry of visual adaptation by regulating Pax6 alpha-enhancer activity. eLife 2017, 6, e21303. [Google Scholar] [CrossRef] [PubMed]
- Stern, S.; Debre, E.; Stritt, C.; Berger, J.; Posern, G.; Knoll, B. A nuclear actin function regulates neuronal motility by serum response factor-dependent gene transcription. J. Neurosci. 2009, 29, 4512–4518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ueda, K.; Serajee, F.; Huq, A.M. A Mutation in the ACTA1 gene Manifesting Nemaline Myopathy with Central Nervous System Lesions. J. Clin. Neurol. 2017, 13, 300–302. [Google Scholar] [CrossRef] [Green Version]
- Chander, P.; Kennedy, M.J.; Winckler, B.; Weick, J.P. Neuron-Specific Gene 2 (NSG2) Encodes an AMPA Receptor Interacting Protein That Modulates Excitatory Neurotransmission. eNeuro 2019, 6, ENEURO.0292-18.2018. [Google Scholar] [CrossRef]
- Yap, C.C.; Digilio, L.; McMahon, L.; Winckler, B. The endosomal neuronal proteins Nsg1/NEEP21 and Nsg2/P19 are itinerant, not resident proteins of dendritic endosomes. Sci. Rep. 2017, 7, 10481. [Google Scholar] [CrossRef] [Green Version]
- Cheng, C.; Lau, S.K.; Doering, L.C. Astrocyte-secreted thrombospondin-1 modulates synapse and spine defects in the fragile X mouse model. Mol. Brain 2016, 9, 74. [Google Scholar] [CrossRef] [Green Version]
- Tyzack, G.E.; Sitnikov, S.; Barson, D.; Adams-Carr, K.L.; Lau, N.K.; Kwok, J.C.; Zhao, C.; Franklin, R.J.; Karadottir, R.T.; Fawcett, J.W.; et al. Astrocyte response to motor neuron injury promotes structural synaptic plasticity via STAT3-regulated TSP-1 expression. Nat. Commun. 2014, 5, 4294. [Google Scholar] [CrossRef] [Green Version]
- Bray, E.R.; Yungher, B.J.; Levay, K.; Ribeiro, M.; Dvoryanchikov, G.; Ayupe, A.C.; Thakor, K.; Marks, V.; Randolph, M.; Danzi, M.C.; et al. Thrombospondin-1 Mediates Axon Regeneration in Retinal Ganglion Cells. Neuron 2019, 103, 642–657.e647. [Google Scholar] [CrossRef]
- Ho, A.; Morishita, W.; Atasoy, D.; Liu, X.; Tabuchi, K.; Hammer, R.E.; Malenka, R.C.; Sudhof, T.C. Genetic analysis of Mint/X11 proteins: Essential presynaptic functions of a neuronal adaptor protein family. J. Neurosci. 2006, 26, 13089–13101. [Google Scholar] [CrossRef] [Green Version]
- Simms, B.A.; Zamponi, G.W. Neuronal voltage-gated calcium channels: Structure, function, and dysfunction. Neuron 2014, 82, 24–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sullivan, S.E.; Dillon, G.M.; Sullivan, J.M.; Ho, A. Mint proteins are required for synaptic activity-dependent amyloid precursor protein (APP) trafficking and amyloid beta generation. J. Biol. Chem. 2014, 289, 15374–15383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, M.; Zuo, Y. Experience-dependent structural plasticity in the cortex. Trends Neurosci. 2011, 34, 177–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holtmaat, A.; Caroni, P. Functional and structural underpinnings of neuronal assembly formation in learning. Nat. Neurosci. 2016, 19, 1553–1562. [Google Scholar] [CrossRef]
- Shah, M.M.; Hammond, R.S.; Hoffman, D.A. Dendritic ion channel trafficking and plasticity. Trends Neurosci. 2010, 33, 307–316. [Google Scholar] [CrossRef] [Green Version]
- Scheefhals, N.; MacGillavry, H.D. Functional organization of postsynaptic glutamate receptors. Mol. Cell Neurosci. 2018, 91, 82–94. [Google Scholar] [CrossRef]
- Lin, Y.; Bloodgood, B.L.; Hauser, J.L.; Lapan, A.D.; Koon, A.C.; Kim, T.K.; Hu, L.S.; Malik, A.N.; Greenberg, M.E. Activity-dependent regulation of inhibitory synapse development by Npas4. Nature 2008, 455, 1198–1204. [Google Scholar] [CrossRef]
- Spiegel, I.; Mardinly, A.R.; Gabel, H.W.; Bazinet, J.E.; Couch, C.H.; Tzeng, C.P.; Harmin, D.A.; Greenberg, M.E. Npas4 regulates excitatory-inhibitory balance within neural circuits through cell-type-specific gene programs. Cell 2014, 157, 1216–1229. [Google Scholar] [CrossRef] [Green Version]
- Ferraro, L.; Beggiato, S.; Borroto-Escuela, D.O.; Ravani, L.; O’Connor, W.T.; Tomasini, M.C.; Borelli, A.C.; Agnati, L.F.; Antonelli, T.; Tanganelli, S.; et al. Neurotensin NTS1-dopamine D2 receptor-receptor interactions in putative receptor heteromers: Relevance for Parkinson’s disease and schizophrenia. Curr. Protein Pept. Sci. 2014, 15, 681–690. [Google Scholar] [CrossRef]
- Goehler, H.; Lalowski, M.; Stelzl, U.; Waelter, S.; Stroedicke, M.; Worm, U.; Droege, A.; Lindenberg, K.S.; Knoblich, M.; Haenig, C.; et al. A protein interaction network links GIT1, an enhancer of huntingtin aggregation, to Huntington’s disease. Mol. Cell 2004, 15, 853–865. [Google Scholar] [CrossRef]
- Awasthi, A.; Ramachandran, B.; Ahmed, S.; Benito, E.; Shinoda, Y.; Nitzan, N.; Heukamp, A.; Rannio, S.; Martens, H.; Barth, J.; et al. Synaptotagmin-3 drives AMPA receptor endocytosis, depression of synapse strength, and forgetting. Science 2019, 363, eaav1483. [Google Scholar] [CrossRef] [Green Version]
- Kitano, J.; Kimura, K.; Yamazaki, Y.; Soda, T.; Shigemoto, R.; Nakajima, Y.; Nakanishi, S. Tamalin, a PDZ domain-containing protein, links a protein complex formation of group 1 metabotropic glutamate receptors and the guanine nucleotide exchange factor cytohesins. J. Neurosci. 2002, 22, 1280–1289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skillback, T.; Mattsson, N.; Hansson, K.; Mirgorodskaya, E.; Dahlen, R.; van der Flier, W.; Scheltens, P.; Duits, F.; Hansson, O.; Teunissen, C.; et al. A novel quantification-driven proteomic strategy identifies an endogenous peptide of pleiotrophin as a new biomarker of Alzheimer’s disease. Sci. Rep. 2017, 7, 13333. [Google Scholar] [CrossRef] [Green Version]
- Yamagata, K.; Andreasson, K.I.; Sugiura, H.; Maru, E.; Dominique, M.; Irie, Y.; Miki, N.; Hayashi, Y.; Yoshioka, M.; Kaneko, K.; et al. Arcadlin is a neural activity-regulated cadherin involved in long term potentiation. J. Biol. Chem. 1999, 274, 19473–19979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yasuda, S.; Tanaka, H.; Sugiura, H.; Okamura, K.; Sakaguchi, T.; Tran, U.; Takemiya, T.; Mizoguchi, A.; Yagita, Y.; Sakurai, T.; et al. Activity-induced protocadherin arcadlin regulates dendritic spine number by triggering N-cadherin endocytosis via TAO2beta and p38 MAP kinases. Neuron 2007, 56, 456–471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menard, C.; Tse, Y.C.; Cavanagh, C.; Chabot, J.G.; Herzog, H.; Schwarzer, C.; Wong, T.P.; Quirion, R. Knockdown of prodynorphin gene prevents cognitive decline, reduces anxiety, and rescues loss of group 1 metabotropic glutamate receptor function in aging. J. Neurosci. 2013, 33, 12792–12804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kayser, M.S.; Nolt, M.J.; Dalva, M.B. EphB receptors couple dendritic filopodia motility to synapse formation. Neuron 2008, 59, 56–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olivares, D.; Deshpande, V.K.; Shi, Y.; Lahiri, D.K.; Greig, N.H.; Rogers, J.T.; Huang, X. N-methyl D-aspartate (NMDA) receptor antagonists and memantine treatment for Alzheimer’s disease, vascular dementia and Parkinson’s disease. Curr. Alzheimer Res. 2012, 9, 746–758. [Google Scholar] [CrossRef]
- Baser, M.E.; Kuramoto, L.; Woods, R.; Joe, H.; Friedman, J.M.; Wallace, A.J.; Ramsden, R.T.; Olschwang, S.; Bijlsma, E.; Kalamarides, M.; et al. The location of constitutional neurofibromatosis 2 (NF2) splice site mutations is associated with the severity of NF2. J. Med. Genet. 2005, 42, 540–546. [Google Scholar] [CrossRef] [Green Version]
- Chan, C.C.; Koch, C.A.; Kaiser-Kupfer, M.I.; Parry, D.M.; Gutmann, D.H.; Zhuang, Z.; Vortmeyer, A.O. Loss of heterozygosity for the NF2 gene in retinal and optic nerve lesions of patients with neurofibromatosis 2. J. Pathol. 2002, 198, 14–20. [Google Scholar] [CrossRef] [Green Version]
- Bereczki, E.; Branca, R.M.; Francis, P.T.; Pereira, J.B.; Baek, J.H.; Hortobagyi, T.; Winblad, B.; Ballard, C.; Lehtio, J.; Aarsland, D. Synaptic markers of cognitive decline in neurodegenerative diseases: A proteomic approach. Brain 2018, 141, 582–595. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Kim, R.; Cho, Y.S.; Song, W.S.; Kim, D.; Kim, K.; Roh, J.D.; Chung, C.; Park, H.; Yang, E.; et al. Lrfn2-Mutant Mice Display Suppressed Synaptic Plasticity and Inhibitory Synapse Development and Abnormal Social Communication and Startle Response. J. Neurosci. 2018, 38, 5872–5887. [Google Scholar] [CrossRef] [Green Version]
- Bustos, F.J.; Ampuero, E.; Jury, N.; Aguilar, R.; Falahi, F.; Toledo, J.; Ahumada, J.; Lata, J.; Cubillos, P.; Henriquez, B.; et al. Epigenetic editing of the Dlg4/PSD95 gene improves cognition in aged and Alzheimer’s disease mice. Brain 2017, 140, 3252–3268. [Google Scholar] [CrossRef] [Green Version]
- Li, K.; Nakajima, M.; Ibanez-Tallon, I.; Heintz, N. A Cortical Circuit for Sexually Dimorphic Oxytocin-Dependent Anxiety Behaviors. Cell 2016, 167, 60–72.e11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corradi, A.; Fadda, M.; Piton, A.; Patry, L.; Marte, A.; Rossi, P.; Cadieux-Dion, M.; Gauthier, J.; Lapointe, L.; Mottron, L.; et al. SYN2 is an autism predisposing gene: Loss-of-function mutations alter synaptic vesicle cycling and axon outgrowth. Hum. Mol. Genet. 2014, 23, 90–103. [Google Scholar] [CrossRef] [PubMed]
- Binda, A.; Rivolta, I.; Villa, C.; Chisci, E.; Beghi, M.; Cornaggia, C.M.; Giovannoni, R.; Combi, R. A Novel KCNJ2 Mutation Identified in an Autistic Proband Affects the Single Channel Properties of Kir2.1. Front. Cell Neurosci. 2018, 12, 76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez-Morillo, E.; Childs, C.; Garcia, B.P.; Alvarez Menendez, F.V.; Romaschin, A.D.; Cervellin, G.; Lippi, G.; Diamandis, E.P. Neurofilament medium polypeptide (NFM) protein concentration is increased in CSF and serum samples from patients with brain injury. Clin. Chem. Lab. Med. 2015, 53, 1575–1584. [Google Scholar] [CrossRef] [PubMed]
- Anitha, A.; Thanseem, I.; Nakamura, K.; Yamada, K.; Iwayama, Y.; Toyota, T.; Iwata, Y.; Suzuki, K.; Sugiyama, T.; Tsujii, M.; et al. Protocadherin alpha (PCDHA) as a novel susceptibility gene for autism. J. Psychiatry Neurosci. 2013, 38, 192–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubinstein, R.; Thu, C.A.; Goodman, K.M.; Wolcott, H.N.; Bahna, F.; Mannepalli, S.; Ahlsen, G.; Chevee, M.; Halim, A.; Clausen, H.; et al. Molecular logic of neuronal self-recognition through protocadherin domain interactions. Cell 2015, 163, 629–642. [Google Scholar] [CrossRef] [Green Version]
- Maselli, R.A.; Arredondo, J.; Vazquez, J.; Chong, J.X.; University of Washington Center for Mendelian Genomics; Bamshad, M.J.; Nickerson, D.A.; Lara, M.; Ng, F.; Lo, V.L.; et al. Presynaptic congenital myasthenic syndrome with a homozygous sequence variant in LAMA5 combines myopia, facial tics, and failure of neuromuscular transmission. Am. J. Med. Genet. A 2017, 173, 2240–2245. [Google Scholar] [CrossRef]
- Dong, W.; Radulovic, T.; Goral, R.O.; Thomas, C.; Suarez Montesinos, M.; Guerrero-Given, D.; Hagiwara, A.; Putzke, T.; Hida, Y.; Abe, M.; et al. CAST/ELKS Proteins Control Voltage-Gated Ca(2+) Channel Density and Synaptic Release Probability at a Mammalian Central Synapse. Cell Rep. 2018, 24, 284–293.e286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaya, N.; Alsagob, M.; D’Adamo, M.C.; Al-Bakheet, A.; Hasan, S.; Muccioli, M.; Almutairi, F.B.; Almass, R.; Aldosary, M.; Monies, D.; et al. KCNA4 deficiency leads to a syndrome of abnormal striatum, congenital cataract and intellectual disability. J. Med. Genet. 2016, 53, 786–792. [Google Scholar] [CrossRef]
- Kang, J.Q.; Macdonald, R.L. Molecular Pathogenic Basis for GABRG2 Mutations Associated with a Spectrum of Epilepsy Syndromes, From Generalized Absence Epilepsy to Dravet Syndrome. JAMA Neurol. 2016, 73, 1009–1016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lomash, R.M.; Gu, X.; Youle, R.J.; Lu, W.; Roche, K.W. Neurolastin, a Dynamin Family GTPase, Regulates Excitatory Synapses and Spine Density. Cell Rep. 2015, 12, 743–751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, S.H.; Bylykbashi, E.; Chatila, Z.K.; Lee, S.W.; Pulli, B.; Clemenson, G.D.; Kim, E.; Rompala, A.; Oram, M.K.; Asselin, C.; et al. Combined adult neurogenesis and BDNF mimic exercise effects on cognition in an Alzheimer’s mouse model. Science 2018, 361, eaan8821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiao, S.S.; Shen, L.L.; Zhu, C.; Bu, X.L.; Liu, Y.H.; Liu, C.H.; Yao, X.Q.; Zhang, L.L.; Zhou, H.D.; Walker, D.G.; et al. Brain-derived neurotrophic factor protects against tau-related neurodegeneration of Alzheimer’s disease. Transl. Psychiatry 2016, 6, e907. [Google Scholar] [CrossRef]
- Bormuth, I.; Yan, K.; Yonemasu, T.; Gummert, M.; Zhang, M.; Wichert, S.; Grishina, O.; Pieper, A.; Zhang, W.; Goebbels, S.; et al. Neuronal basic helix-loop-helix proteins Neurod2/6 regulate cortical commissure formation before midline interactions. J. Neurosci. 2013, 33, 641–651. [Google Scholar] [CrossRef] [Green Version]
- Pieper, A.; Rudolph, S.; Wieser, G.L.; Gotze, T.; Miessner, H.; Yonemasu, T.; Yan, K.; Tzvetanova, I.; Castillo, B.D.; Bode, U.; et al. NeuroD2 controls inhibitory circuit formation in the molecular layer of the cerebellum. Sci. Rep. 2019, 9, 1448. [Google Scholar] [CrossRef]
- Giza, J.; Urbanski, M.J.; Prestori, F.; Bandyopadhyay, B.; Yam, A.; Friedrich, V.; Kelley, K.; D’Angelo, E.; Goldfarb, M. Behavioral and cerebellar transmission deficits in mice lacking the autism-linked gene islet brain-2. J. Neurosci. 2010, 30, 14805–14816. [Google Scholar] [CrossRef] [PubMed]
- Mitz, A.R.; Philyaw, T.J.; Boccuto, L.; Shcheglovitov, A.; Sarasua, S.M.; Kaufmann, W.E.; Thurm, A. Identification of 22q13 genes most likely to contribute to Phelan McDermid syndrome. Eur. J. Hum. Genet. 2018, 26, 293–302. [Google Scholar] [CrossRef] [Green Version]
- Hatanaka, Y.; Watase, K.; Wada, K.; Nagai, Y. Abnormalities in synaptic dynamics during development in a mouse model of spinocerebellar ataxia type 1. Sci. Rep. 2015, 5, 16102. [Google Scholar] [CrossRef] [PubMed]
- Guan, F.; Zhang, T.; Liu, X.; Han, W.; Lin, H.; Li, L.; Chen, G.; Li, T. Evaluation of voltage-dependent calcium channel gamma gene families identified several novel potential susceptible genes to schizophrenia. Sci. Rep. 2016, 6, 24914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korber, C.; Werner, M.; Kott, S.; Ma, Z.L.; Hollmann, M. The transmembrane AMPA receptor regulatory protein gamma 4 is a more effective modulator of AMPA receptor function than stargazin (gamma 2). J. Neurosci. 2007, 27, 8442–8447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seigneur, E.; Sudhof, T.C. Genetic Ablation of All Cerebellins Reveals Synapse Organizer Functions in Multiple Regions Throughout the Brain. J. Neurosci. 2018, 38, 4774–4790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tao, W.; Diaz-Alonso, J.; Sheng, N.; Nicoll, R.A. Postsynaptic delta1 glutamate receptor assembles and maintains hippocampal synapses via Cbln2 and neurexin. Proc. Natl. Acad. Sci. USA 2018, 115, E5373–E5381. [Google Scholar] [CrossRef] [Green Version]
- Sadakata, T.; Washida, M.; Iwayama, Y.; Shoji, S.; Sato, Y.; Ohkura, T.; Katoh-Semba, R.; Nakajima, M.; Sekine, Y.; Tanaka, M.; et al. Autistic-like phenotypes in Cadps2-knockout mice and aberrant CADPS2 splicing in autistic patients. J. Clin. Investig. 2007, 117, 931–943. [Google Scholar] [CrossRef] [Green Version]
- Shinoda, Y.; Sadakata, T.; Akagi, T.; Sakamaki, Y.; Hashikawa, T.; Sano, Y.; Furuichi, T. Calcium-dependent activator protein for secretion 2 (CADPS2) deficiency causes abnormal synapse development in hippocampal mossy fiber terminals. Neurosci. Lett. 2018, 677, 65–71. [Google Scholar] [CrossRef]
- Armendariz, B.G.; Bribian, A.; Perez-Martinez, E.; Martinez, A.; de Castro, F.; Soriano, E.; Burgaya, F. Expression of Semaphorin 4F in neurons and brain oligodendrocytes and the regulation of oligodendrocyte precursor migration in the optic nerve. Mol. Cell Neurosci. 2012, 49, 54–67. [Google Scholar] [CrossRef]
- Minett, M.S.; Pereira, V.; Sikandar, S.; Matsuyama, A.; Lolignier, S.; Kanellopoulos, A.H.; Mancini, F.; Iannetti, G.D.; Bogdanov, Y.D.; Santana-Varela, S.; et al. Endogenous opioids contribute to insensitivity to pain in humans and mice lacking sodium channel Nav1.7. Nat. Commun. 2015, 6, 8967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shiina, N.; Tokunaga, M. RNA granule protein 140 (RNG140), a paralog of RNG105 localized to distinct RNA granules in neuronal dendrites in the adult vertebrate brain. J. Biol. Chem. 2010, 285, 24260–24269. [Google Scholar] [CrossRef] [Green Version]
- Ackerley, S.; James, P.A.; Kalli, A.; French, S.; Davies, K.E.; Talbot, K. A mutation in the small heat-shock protein HSPB1 leading to distal hereditary motor neuronopathy disrupts neurofilament assembly and the axonal transport of specific cellular cargoes. Hum. Mol. Genet. 2006, 15, 347–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baughman, H.E.R.; Clouser, A.F.; Klevit, R.E.; Nath, A. HspB1 and Hsc70 chaperones engage distinct tau species and have different inhibitory effects on amyloid formation. J. Biol. Chem. 2018, 293, 2687–2700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- King, M.; Nafar, F.; Clarke, J.; Mearow, K. The small heat shock protein Hsp27 protects cortical neurons against the toxic effects of beta-amyloid peptide. J. Neurosci. Res. 2009, 87, 3161–3175. [Google Scholar] [CrossRef] [PubMed]
- Li, M.D.; Mangold, J.E.; Seneviratne, C.; Chen, G.B.; Ma, J.Z.; Lou, X.Y.; Payne, T.J. Association and interaction analyses of GABBR1 and GABBR2 with nicotine dependence in European- and African-American populations. PLoS ONE 2009, 4, e7055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoo, Y.; Jung, J.; Lee, Y.N.; Lee, Y.; Cho, H.; Na, E.; Hong, J.; Kim, E.; Lee, J.S.; Lee, J.S.; et al. GABBR2 mutations determine phenotype in rett syndrome and epileptic encephalopathy. Ann. Neurol. 2017, 82, 466–478. [Google Scholar] [CrossRef]
- Kakegawa, W.; Mitakidis, N.; Miura, E.; Abe, M.; Matsuda, K.; Takeo, Y.H.; Kohda, K.; Motohashi, J.; Takahashi, A.; Nagao, S.; et al. Anterograde C1ql1 signaling is required in order to determine and maintain a single-winner climbing fiber in the mouse cerebellum. Neuron 2015, 85, 316–329. [Google Scholar] [CrossRef] [Green Version]
- Chew, K.S.; Fernandez, D.C.; Hattar, S.; Sudhof, T.C.; Martinelli, D.C. Anatomical and Behavioral Investigation of C1ql3 in the Mouse Suprachiasmatic Nucleus. J. Biol. Rhythms 2017, 32, 222–236. [Google Scholar] [CrossRef] [Green Version]
- Martinelli, D.C.; Chew, K.S.; Rohlmann, A.; Lum, M.Y.; Ressl, S.; Hattar, S.; Brunger, A.T.; Missler, M.; Sudhof, T.C. Expression of C1ql3 in Discrete Neuronal Populations Controls Efferent Synapse Numbers and Diverse Behaviors. Neuron 2016, 91, 1034–1051. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Wang, B.L.; Sun, F.R.; Li, J.Q.; Cao, X.P.; Tan, L. The role of UNC5C in Alzheimer’s disease. Ann. Transl. Med. 2018, 6, 178. [Google Scholar] [CrossRef]
- Schroeder, A.; de Wit, J. Leucine-rich repeat-containing synaptic adhesion molecules as organizers of synaptic specificity and diversity. Exp. Mol. Med. 2018, 50, 10. [Google Scholar] [CrossRef] [Green Version]
- Shao, Q.; Yang, T.; Huang, H.; Alarmanazi, F.; Liu, G. Uncoupling of UNC5C with Polymerized TUBB3 in Microtubules Mediates Netrin-1 Repulsion. J. Neurosci. 2017, 37, 5620–5633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Kesteren, R.E.; Spencer, G.E. The role of neurotransmitters in neurite outgrowth and synapse formation. Rev. Neurosci. 2003, 14, 217–231. [Google Scholar] [CrossRef]
- McCann, R.F.; Ross, D.A. A Fragile Balance: Dendritic Spines, Learning, and Memory. Biol. Psychiatry 2017, 82, e11–e13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abraham, W.C.; Jones, O.D.; Glanzman, D.L. Is plasticity of synapses the mechanism of long-term memory storage? NPJ Sci. Learn. 2019, 4, 9. [Google Scholar] [CrossRef]
- Bloom, G.S. Amyloid-beta and tau: The trigger and bullet in Alzheimer disease pathogenesis. JAMA Neurol. 2014, 71, 505–508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buniello, A.; MacArthur, J.A.L.; Cerezo, M.; Harris, L.W.; Hayhurst, J.; Malangone, C.; McMahon, A.; Morales, J.; Mountjoy, E.; Sollis, E.; et al. The NHGRI-EBI GWAS Catalog of published genome-wide association studies, targeted arrays and summary statistics 2019. Nucleic Acids Res. 2019, 47, D1005–D1012. [Google Scholar] [CrossRef] [Green Version]
- Harold, D.; Abraham, R.; Hollingworth, P.; Sims, R.; Gerrish, A.; Hamshere, M.L.; Pahwa, J.S.; Moskvina, V.; Dowzell, K.; Williams, A.; et al. Genome-wide association study identifies variants at CLU and PICALM associated with Alzheimer’s disease. Nat. Genet. 2009, 41, 1088–1093. [Google Scholar] [CrossRef] [Green Version]
- Jansen, I.E.; Savage, J.E.; Watanabe, K.; Bryois, J.; Williams, D.M.; Steinberg, S.; Sealock, J.; Karlsson, I.K.; Hagg, S.; Athanasiu, L.; et al. Genome-wide meta-analysis identifies new loci and functional pathways influencing Alzheimer’s disease risk. Nat. Genet. 2019, 51, 404–413. [Google Scholar] [CrossRef]
- Jun, G.; Ibrahim-Verbaas, C.A.; Vronskaya, M.; Lambert, J.C.; Chung, J.; Naj, A.C.; Kunkle, B.W.; Wang, L.S.; Bis, J.C.; Bellenguez, C.; et al. A novel Alzheimer disease locus located near the gene encoding tau protein. Mol. Psychiatry 2016, 21, 108–117. [Google Scholar] [CrossRef] [Green Version]
- Coon, K.D.; Myers, A.J.; Craig, D.W.; Webster, J.A.; Pearson, J.V.; Lince, D.H.; Zismann, V.L.; Beach, T.G.; Leung, D.; Bryden, L.; et al. A high-density whole-genome association study reveals that APOE is the major susceptibility gene for sporadic late-onset Alzheimer’s disease. J. Clin. Psychiatry 2007, 68, 613–618. [Google Scholar] [CrossRef]
- Lambert, J.C.; Ibrahim-Verbaas, C.A.; Harold, D.; Naj, A.C.; Sims, R.; Bellenguez, C.; DeStafano, A.L.; Bis, J.C.; Beecham, G.W.; Grenier-Boley, B.; et al. Meta-analysis of 74,046 individuals identifies 11 new susceptibility loci for Alzheimer’s disease. Nat. Genet. 2013, 45, 1452–1458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nikolaienko, O.; Patil, S.; Eriksen, M.S.; Bramham, C.R. Arc protein: A flexible hub for synaptic plasticity and cognition. Semin. Cell Dev. Biol. 2018, 77, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Gozdz, A.; Nikolaienko, O.; Urbanska, M.; Cymerman, I.A.; Sitkiewicz, E.; Blazejczyk, M.; Dadlez, M.; Bramham, C.R.; Jaworski, J. GSK3alpha and GSK3beta Phosphorylate Arc and Regulate its Degradation. Front. Mol. Neurosci. 2017, 10, 192. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Petralia, R.S.; Kurushima, H.; Patel, H.; Jung, M.Y.; Volk, L.; Chowdhury, S.; Shepherd, J.D.; Dehoff, M.; Li, Y.; et al. Arc/Arg3.1 regulates an endosomal pathway essential for activity-dependent beta-amyloid generation. Cell 2011, 147, 615–628. [Google Scholar] [CrossRef] [Green Version]
- Bertram, L.; Hiltunen, M.; Parkinson, M.; Ingelsson, M.; Lange, C.; Ramasamy, K.; Mullin, K.; Menon, R.; Sampson, A.J.; Hsiao, M.Y.; et al. Family-based association between Alzheimer’s disease and variants in UBQLN1. N. Engl. J. Med. 2005, 352, 884–894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rehker, J.; Rodhe, J.; Nesbitt, R.R.; Boyle, E.A.; Martin, B.K.; Lord, J.; Karaca, I.; Naj, A.; Jessen, F.; Helisalmi, S.; et al. Caspase-8, association with Alzheimer’s Disease and functional analysis of rare variants. PLoS ONE 2017, 12, e0185777. [Google Scholar] [CrossRef] [Green Version]
- Saha, P.; Biswas, S.C. Amyloid-beta induced astrocytosis and astrocyte death: Implication of FoxO3a-Bim-caspase3 death signaling. Mol. Cell Neurosci. 2015, 68, 203–211. [Google Scholar] [CrossRef]
- Sanphui, P.; Biswas, S.C. FoxO3a is activated and executes neuron death via Bim in response to beta-amyloid. Cell Death Dis. 2013, 4, e625. [Google Scholar] [CrossRef]
- Woods, Y.L.; Cohen, P.; Becker, W.; Jakes, R.; Goedert, M.; Wang, X.; Proud, C.G. The kinase DYRK phosphorylates protein-synthesis initiation factor eIF2Bepsilon at Ser539 and the microtubule-associated protein tau at Thr212: Potential role for DYRK as a glycogen synthase kinase 3-priming kinase. Biochem. J. 2001, 355, 609–615. [Google Scholar] [CrossRef]
- Inoue, M.; Hur, J.Y.; Kihara, T.; Teranishi, Y.; Yamamoto, N.G.; Ishikawa, T.; Wiehager, B.; Winblad, B.; Tjernberg, L.O.; Schedin-Weiss, S. Human brain proteins showing neuron-specific interactions with gamma-secretase. FEBS J. 2015, 282, 2587–2599. [Google Scholar] [CrossRef]
- Kolsch, H.; Wagner, M.; Bilkei-Gorzo, A.; Toliat, M.R.; Pentzek, M.; Fuchs, A.; Kaduszkiewicz, H.; van den Bussche, H.; Riedel-Heller, S.G.; Angermeyer, M.C.; et al. Gene polymorphisms in prodynorphin (PDYN) are associated with episodic memory in the elderly. J. Neural Transm. 2009, 116, 897–903. [Google Scholar] [CrossRef] [PubMed]
- Menard, C.; Herzog, H.; Schwarzer, C.; Quirion, R. Possible role of dynorphins in Alzheimer’s disease and age-related cognitive deficits. Neurodegener. Dis. 2014, 13, 82–85. [Google Scholar] [CrossRef] [PubMed]
- Yakovleva, T.; Marinova, Z.; Kuzmin, A.; Seidah, N.G.; Haroutunian, V.; Terenius, L.; Bakalkin, G. Dysregulation of dynorphins in Alzheimer disease. Neurobiol. Aging 2007, 28, 1700–1708. [Google Scholar] [CrossRef] [PubMed]
- Abdullah, L.; Ait-Ghezala, G.; Crawford, F.; Crowell, T.A.; Barker, W.W.; Duara, R.; Mullan, M. The cyclooxygenase 2 -765 C promoter allele is a protective factor for Alzheimer’s disease. Neurosci. Lett. 2006, 395, 240–243. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Liang, B.; Wang, Z.; Cheng, X.; Huang, Y.; Liu, Y.; Huang, Z. Influence of four polymorphisms in ABCA1 and PTGS2 genes on risk of Alzheimer’s disease: A meta-analysis. Neurol. Sci. 2016, 37, 1209–1220. [Google Scholar] [CrossRef] [PubMed]
- Guan, P.P.; Wang, P. Integrated communications between cyclooxygenase-2 and Alzheimer’s disease. FASEB J. 2019, 33, 13–33. [Google Scholar] [CrossRef] [PubMed]
- Ma, S.L.; Tang, N.L.; Zhang, Y.P.; Ji, L.D.; Tam, C.W.; Lui, V.W.; Chiu, H.F.; Lam, L.C. Association of prostaglandin-endoperoxide synthase 2 (PTGS2) polymorphisms and Alzheimer’s disease in Chinese. Neurobiol. Aging 2008, 29, 856–860. [Google Scholar] [CrossRef]
- Vito, P.; Lacana, E.; D’Adamio, L. Interfering with apoptosis: Ca(2+)-binding protein ALG-2 and Alzheimer’s disease gene ALG-3. Science 1996, 271, 521–525. [Google Scholar] [CrossRef] [Green Version]
- Nelson, C.D.; Sheng, M. Gpr3 stimulates Abeta production via interactions with APP and beta-arrestin2. PLoS ONE 2013, 8, e74680. [Google Scholar] [CrossRef] [Green Version]
- Thathiah, A.; Spittaels, K.; Hoffmann, M.; Staes, M.; Cohen, A.; Horre, K.; Vanbrabant, M.; Coun, F.; Baekelandt, V.; Delacourte, A.; et al. The orphan G protein-coupled receptor 3 modulates amyloid-beta peptide generation in neurons. Science 2009, 323, 946–951. [Google Scholar] [CrossRef]
- Huang, Y.; Skwarek-Maruszewska, A.; Horré, K.; Vandewyer, E.; Wolfs, L.; Snellinx, A.; Saito, T.; Radaelli, E.; Corthout, N.; Colombelli, J.; et al. Loss of GPR3 reduces the amyloid plaque burden and improves memory in Alzheimer’s disease mouse models. Sci. Transl. Med. 2015, 7, 309ra164. [Google Scholar] [CrossRef]
- Campolongo, P.; Ratano, P.; Ciotti, M.T.; Florenzano, F.; Nori, S.L.; Marolda, R.; Palmery, M.; Rinaldi, A.M.; Zona, C.; Possenti, R.; et al. Systemic administration of substance P recovers beta amyloid-induced cognitive deficits in rat: Involvement of Kv potassium channels. PLoS ONE 2013, 8, e78036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pieri, M.; Amadoro, G.; Carunchio, I.; Ciotti, M.T.; Quaresima, S.; Florenzano, F.; Calissano, P.; Possenti, R.; Zona, C.; Severini, C. SP protects cerebellar granule cells against beta-amyloid-induced apoptosis by down-regulation and reduced activity of Kv4 potassium channels. Neuropharmacology 2010, 58, 268–276. [Google Scholar] [CrossRef]
- Beyer, K.; Lao, J.I.; Gomez, M.; Riutort, N.; Latorre, P.; Mate, J.L.; Ariza, A. Alzheimer’s disease and the cystatin C gene polymorphism: An association study. Neurosci. Lett. 2001, 315, 17–20. [Google Scholar] [CrossRef]
- Finckh, U.; von der Kammer, H.; Velden, J.; Michel, T.; Andresen, B.; Deng, A.; Zhang, J.; Muller-Thomsen, T.; Zuchowski, K.; Menzer, G.; et al. Genetic association of a cystatin C gene polymorphism with late-onset Alzheimer disease. Arch. Neurol. 2000, 57, 1579–1583. [Google Scholar] [CrossRef] [PubMed]
- Kaur, G.; Levy, E. Cystatin C in Alzheimer’s disease. Front. Mol. Neurosci. 2012, 5, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hettiarachchi, N.; Dallas, M.; Al-Owais, M.; Griffiths, H.; Hooper, N.; Scragg, J.; Boyle, J.; Peers, C. Heme oxygenase-1 protects against Alzheimer’s amyloid-beta(1–42)-induced toxicity via carbon monoxide production. Cell Death Dis. 2014, 5, e1569. [Google Scholar] [CrossRef] [Green Version]
- Hettiarachchi, N.T.; Boyle, J.P.; Dallas, M.L.; Al-Owais, M.M.; Scragg, J.L.; Peers, C. Heme oxygenase-1 derived carbon monoxide suppresses Abeta1–42 toxicity in astrocytes. Cell Death Dis. 2017, 8, e2884. [Google Scholar] [CrossRef]
- Allen, M.; Zou, F.; Chai, H.S.; Younkin, C.S.; Miles, R.; Nair, A.A.; Crook, J.E.; Pankratz, V.S.; Carrasquillo, M.M.; Rowley, C.N.; et al. Glutathione S-transferase omega genes in Alzheimer and Parkinson disease risk, age-at-diagnosis and brain gene expression: An association study with mechanistic implications. Mol. Neurodegener. 2012, 7, 13. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.J.; Oliveira, S.A.; Xu, P.; Martin, E.R.; Stenger, J.E.; Scherzer, C.R.; Hauser, M.A.; Scott, W.K.; Small, G.W.; Nance, M.A.; et al. Glutathione S-transferase omega-1 modifies age-at-onset of Alzheimer disease and Parkinson disease. Hum. Mol. Genet. 2003, 12, 3259–3267. [Google Scholar] [CrossRef] [Green Version]
- Moon, M.; Jung, E.S.; Jeon, S.G.; Cha, M.Y.; Jang, Y.; Kim, W.; Lopes, C.; Mook-Jung, I.; Kim, K.S. Nurr1 (NR4A2) regulates Alzheimer’s disease-related pathogenesis and cognitive function in the 5XFAD mouse model. Aging Cell 2019, 18, e12866. [Google Scholar] [CrossRef] [PubMed]
- Meilandt, W.J.; Yu, G.Q.; Chin, J.; Roberson, E.D.; Palop, J.J.; Wu, T.; Scearce-Levie, K.; Mucke, L. Enkephalin elevations contribute to neuronal and behavioral impairments in a transgenic mouse model of Alzheimer’s disease. J. Neurosci. 2008, 28, 5007–5017. [Google Scholar] [CrossRef]
- Guo, Q.; Fu, W.; Xie, J.; Luo, H.; Sells, S.F.; Geddes, J.W.; Bondada, V.; Rangnekar, V.M.; Mattson, M.P. Par-4 is a mediator of neuronal degeneration associated with the pathogenesis of Alzheimer disease. Nat. Med. 1998, 4, 957–962. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Guo, Q. PAR-4 is involved in regulation of beta-secretase cleavage of the Alzheimer amyloid precursor protein. J. Biol. Chem. 2005, 280, 13824–13832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishiki, A.; Kamada, M.; Kawamura, Y.; Terao, C.; Shimoda, F.; Tomita, N.; Arai, H.; Furukawa, K. Glial fibrillar acidic protein in the cerebrospinal fluid of Alzheimer’s disease, dementia with Lewy bodies, and frontotemporal lobar degeneration. J. Neurochem. 2016, 136, 258–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oeckl, P.; Halbgebauer, S.; Anderl-Straub, S.; Steinacker, P.; Huss, A.M.; Neugebauer, H.; von Arnim, C.A.F.; Diehl-Schmid, J.; Grimmer, T.; Kornhuber, J.; et al. Glial Fibrillary Acidic Protein in Serum is Increased in Alzheimer’s Disease and Correlates with Cognitive Impairment. J. Alzheimers Dis. 2019, 67, 481–488. [Google Scholar] [CrossRef]
- Hoe, H.S.; Cooper, M.J.; Burns, M.P.; Lewis, P.A.; van der Brug, M.; Chakraborty, G.; Cartagena, C.M.; Pak, D.T.; Cookson, M.R.; Rebeck, G.W. The metalloprotease inhibitor TIMP-3 regulates amyloid precursor protein and apolipoprotein E receptor proteolysis. J. Neurosci. 2007, 27, 10895–10905. [Google Scholar] [CrossRef] [Green Version]
- Buxbaum, J.D.; Choi, E.K.; Luo, Y.; Lilliehook, C.; Crowley, A.C.; Merriam, D.E.; Wasco, W. Calsenilin: A calcium-binding protein that interacts with the presenilins and regulates the levels of a presenilin fragment. Nat. Med. 1998, 4, 1177–1181. [Google Scholar] [CrossRef]
- Crary, J.F.; Shao, C.Y.; Mirra, S.S.; Hernandez, A.I.; Sacktor, T.C. Atypical protein kinase C in neurodegenerative disease I: PKMzeta aggregates with limbic neurofibrillary tangles and AMPA receptors in Alzheimer disease. J. Neuropathol. Exp. Neurol. 2006, 65, 319–326. [Google Scholar] [CrossRef] [Green Version]
- Liang, D.; Han, G.; Feng, X.; Sun, J.; Duan, Y.; Lei, H. Concerted perturbation observed in a hub network in Alzheimer’s disease. PLoS ONE 2012, 7, e40498. [Google Scholar] [CrossRef]
- Park, B.; Lee, W.; Han, K. Modeling the interactions of Alzheimer-related genes from the whole brain microarray data and diffusion tensor images of human brain. BMC Bioinform. 2012, 13 (Suppl. 7), S10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desrumaux, C.; Pisoni, A.; Meunier, J.; Deckert, V.; Athias, A.; Perrier, V.; Villard, V.; Lagrost, L.; Verdier, J.M.; Maurice, T. Increased amyloid-beta peptide-induced memory deficits in phospholipid transfer protein (PLTP) gene knockout mice. Neuropsychopharmacology 2013, 38, 817–825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, W.; Albers, J.J.; Vuletic, S. Phospholipid transfer protein reduces phosphorylation of tau in human neuronal cells. J. Neurosci. Res. 2009, 87, 3176–3185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mansuy, M.; Baille, S.; Canet, G.; Borie, A.; Cohen-Solal, C.; Vignes, M.; Perrier, V.; Chevallier, N.; Le Guern, N.; Deckert, V.; et al. Deletion of plasma Phospholipid Transfer Protein (PLTP) increases microglial phagocytosis and reduces cerebral amyloid-beta deposition in the J20 mouse model of Alzheimer’s disease. Oncotarget 2018, 9, 19688–19703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tong, Y.; Sun, Y.; Tian, X.; Zhou, T.; Wang, H.; Zhang, T.; Zhan, R.; Zhao, L.; Kuerban, B.; Li, Z.; et al. Phospholipid transfer protein (PLTP) deficiency accelerates memory dysfunction through altering amyloid precursor protein (APP) processing in a mouse model of Alzheimer’s disease. Hum. Mol. Genet. 2015, 24, 5388–5403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vuletic, S.; Peskind, E.R.; Marcovina, S.M.; Quinn, J.F.; Cheung, M.C.; Kennedy, H.; Kaye, J.A.; Jin, L.W.; Albers, J.J. Reduced CSF PLTP activity in Alzheimer’s disease and other neurologic diseases; PLTP induces ApoE secretion in primary human astrocytes in vitro. J. Neurosci. Res. 2005, 80, 406–413. [Google Scholar] [CrossRef]
- Wang, H.; Yu, Y.; Chen, W.; Cui, Y.; Luo, T.; Ma, J.; Jiang, X.C.; Qin, S. PLTP deficiency impairs learning and memory capabilities partially due to alteration of amyloid-beta metabolism in old mice. J. Alzheimers Dis. 2014, 39, 79–88. [Google Scholar] [CrossRef]
- Hernandez, F.; Lucas, J.J.; Avila, J. GSK3 and tau: Two convergence points in Alzheimer’s disease. J. Alzheimers Dis. 2013, 33 (Suppl. 1), S141–S144. [Google Scholar] [CrossRef] [Green Version]
- Llorens-Martin, M.; Jurado, J.; Hernandez, F.; Avila, J. GSK-3beta, a pivotal kinase in Alzheimer disease. Front. Mol. Neurosci. 2014, 7, 46. [Google Scholar]
- Guedes, J.R.; Lao, T.; Cardoso, A.L.; El Khoury, J. Roles of Microglial and Monocyte Chemokines and Their Receptors in Regulating Alzheimer’s Disease-Associated Amyloid-beta and Tau Pathologies. Front. Neurol. 2018, 9, 549. [Google Scholar] [CrossRef]
- Naert, G.; Rivest, S. A deficiency in CCR2+ monocytes: The hidden side of Alzheimer’s disease. J. Mol. Cell Biol. 2013, 5, 284–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westin, K.; Buchhave, P.; Nielsen, H.; Minthon, L.; Janciauskiene, S.; Hansson, O. CCL2 is associated with a faster rate of cognitive decline during early stages of Alzheimer’s disease. PLoS ONE 2012, 7, e30525. [Google Scholar] [CrossRef] [PubMed]
- Khandelwal, P.J.; Dumanis, S.B.; Herman, A.M.; Rebeck, G.W.; Moussa, C.E. Wild type and P301L mutant Tau promote neuro-inflammation and alpha-Synuclein accumulation in lentiviral gene delivery models. Mol. Cell Neurosci. 2012, 49, 44–53. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.; Lee, J.S.; Lee, K.J.; Turner, R.S.; Hoe, H.S.; Pak, D.T.S. Polo-like kinase 2 phosphorylation of amyloid precursor protein regulates activity-dependent amyloidogenic processing. Neuropharmacology 2017, 117, 387–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seeburg, D.P.; Feliu-Mojer, M.; Gaiottino, J.; Pak, D.T.; Sheng, M. Critical role of CDK5 and Polo-like kinase 2 in homeostatic synaptic plasticity during elevated activity. Neuron 2008, 58, 571–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kern, S.; Syrjanen, J.A.; Blennow, K.; Zetterberg, H.; Skoog, I.; Waern, M.; Hagen, C.E.; van Harten, A.C.; Knopman, D.S.; Jack, C.R., Jr.; et al. Association of Cerebrospinal Fluid Neurofilament Light Protein with Risk of Mild Cognitive Impairment Among Individuals Without Cognitive Impairment. JAMA Neurol. 2018, 76, 187–193. [Google Scholar] [CrossRef]
- Lewczuk, P.; Ermann, N.; Andreasson, U.; Schultheis, C.; Podhorna, J.; Spitzer, P.; Maler, J.M.; Kornhuber, J.; Blennow, K.; Zetterberg, H. Plasma neurofilament light as a potential biomarker of neurodegeneration in Alzheimer’s disease. Alzheimers Res. Ther. 2018, 10, 71. [Google Scholar] [CrossRef]
- Preische, O.; Schultz, S.A.; Apel, A.; Kuhle, J.; Kaeser, S.A.; Barro, C.; Graber, S.; Kuder-Buletta, E.; LaFougere, C.; Laske, C.; et al. Serum neurofilament dynamics predicts neurodegeneration and clinical progression in presymptomatic Alzheimer’s disease. Nat. Med. 2019, 25, 277–283. [Google Scholar] [CrossRef]
- Mattsson, N.; Cullen, N.C.; Andreasson, U.; Zetterberg, H.; Blennow, K. Association Between Longitudinal Plasma Neurofilament Light and Neurodegeneration in Patients with Alzheimer Disease. JAMA Neurol. 2019, 76, 791–799. [Google Scholar] [CrossRef]
- Corcoran, N.M.; Martin, D.; Hutter-Paier, B.; Windisch, M.; Nguyen, T.; Nheu, L.; Sundstrom, L.E.; Costello, A.J.; Hovens, C.M. Sodium selenate specifically activates PP2A phosphatase, dephosphorylates tau and reverses memory deficits in an Alzheimer’s disease model. J. Clin. Neurosci. 2010, 17, 1025–1033. [Google Scholar] [CrossRef]
- Du, X.; Li, H.; Wang, Z.; Qiu, S.; Liu, Q.; Ni, J. Selenoprotein P and selenoprotein M block Zn2+—Mediated Abeta42 aggregation and toxicity. Metallomics 2013, 5, 861–870. [Google Scholar] [CrossRef] [PubMed]
- Song, G.; Zhang, Z.; Wen, L.; Chen, C.; Shi, Q.; Zhang, Y.; Ni, J.; Liu, Q. Selenomethionine ameliorates cognitive decline, reduces tau hyperphosphorylation, and reverses synaptic deficit in the triple transgenic mouse model of Alzheimer’s disease. J. Alzheimers Dis. 2014, 41, 85–99. [Google Scholar] [CrossRef] [PubMed]
- Van Eersel, J.; Ke, Y.D.; Liu, X.; Delerue, F.; Kril, J.J.; Gotz, J.; Ittner, L.M. Sodium selenate mitigates tau pathology, neurodegeneration, and functional deficits in Alzheimer’s disease models. Proc. Natl. Acad. Sci. USA 2010, 107, 13888–13893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.C.; Hu, J.; Zhao, N.; Wang, J.; Wang, N.; Cirrito, J.R.; Kanekiyo, T.; Holtzman, D.M.; Bu, G. Astrocytic LRP1 Mediates Brain Abeta Clearance and Impacts Amyloid Deposition. J. Neurosci. 2017, 37, 4023–4031. [Google Scholar] [CrossRef] [Green Version]
- Storck, S.E.; Hartz, A.M.S.; Bernard, J.; Wolf, A.; Kachlmeier, A.; Mahringer, A.; Weggen, S.; Pahnke, J.; Pietrzik, C.U. The concerted amyloid-beta clearance of LRP1 and ABCB1/P-gp across the blood-brain barrier is linked by PICALM. Brain Behav. Immun. 2018, 73, 21–33. [Google Scholar] [CrossRef]
- Storck, S.E.; Meister, S.; Nahrath, J.; Meissner, J.N.; Schubert, N.; Di Spiezio, A.; Baches, S.; Vandenbroucke, R.E.; Bouter, Y.; Prikulis, I.; et al. Endothelial LRP1 transports amyloid-beta(1–42) across the blood-brain barrier. J. Clin. Investig. 2016, 126, 123–136. [Google Scholar] [CrossRef] [Green Version]
- Kook, S.Y.; Jeong, H.; Kang, M.J.; Park, R.; Shin, H.J.; Han, S.H.; Son, S.M.; Song, H.; Baik, S.H.; Moon, M.; et al. Crucial role of calbindin-D28k in the pathogenesis of Alzheimer’s disease mouse model. Cell Death Differ. 2014, 21, 1575–1587. [Google Scholar] [CrossRef]
- Riascos, D.; de Leon, D.; Baker-Nigh, A.; Nicholas, A.; Yukhananov, R.; Bu, J.; Wu, C.K.; Geula, C. Age-related loss of calcium buffering and selective neuronal vulnerability in Alzheimer’s disease. Acta Neuropathol. 2011, 122, 565–576. [Google Scholar] [CrossRef] [Green Version]
- Park, D.; Na, M.; Kim, J.A.; Lee, U.; Cho, E.; Jang, M.; Chang, S. Activation of CaMKIV by soluble amyloid-beta1–42 impedes trafficking of axonal vesicles and impairs activity-dependent synaptogenesis. Sci. Signal. 2017, 10, eaam8661. [Google Scholar] [CrossRef]
- Kaden, D.; Voigt, P.; Munter, L.M.; Bobowski, K.D.; Schaefer, M.; Multhaup, G. Subcellular localization and dimerization of APLP1 are strikingly different from APP and APLP2. J. Cell Sci. 2009, 122, 368–377. [Google Scholar] [CrossRef] [Green Version]
- D’Addario, C.; Di Francesco, A.; Arosio, B.; Gussago, C.; Dell’Osso, B.; Bari, M.; Galimberti, D.; Scarpini, E.; Altamura, A.C.; Mari, D.; et al. Epigenetic regulation of fatty acid amide hydrolase in Alzheimer disease. PLoS ONE 2012, 7, e39186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montanari, S.; Scalvini, L.; Bartolini, M.; Belluti, F.; Gobbi, S.; Andrisano, V.; Ligresti, A.; Di Marzo, V.; Rivara, S.; Mor, M.; et al. Fatty Acid Amide Hydrolase (FAAH), Acetylcholinesterase (AChE), and Butyrylcholinesterase (BuChE): Networked Targets for the Development of Carbamates as Potential Anti-Alzheimer’s Disease Agents. J. Med. Chem. 2016, 59, 6387–6406. [Google Scholar] [CrossRef] [PubMed]
- Mulder, J.; Zilberter, M.; Pasquare, S.J.; Alpar, A.; Schulte, G.; Ferreira, S.G.; Kofalvi, A.; Martin-Moreno, A.M.; Keimpema, E.; Tanila, H.; et al. Molecular reorganization of endocannabinoid signalling in Alzheimer’s disease. Brain 2011, 134, 1041–1060. [Google Scholar] [CrossRef] [PubMed]
- Williamson, R.; van Aalten, L.; Mann, D.M.; Platt, B.; Plattner, F.; Bedford, L.; Mayer, J.; Howlett, D.; Usardi, A.; Sutherland, C.; et al. CRMP2 hyperphosphorylation is characteristic of Alzheimer’s disease and not a feature common to other neurodegenerative diseases. J. Alzheimers Dis. 2011, 27, 615–625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xing, H.; Lim, Y.A.; Chong, J.R.; Lee, J.H.; Aarsland, D.; Ballard, C.G.; Francis, P.T.; Chen, C.P.; Lai, M.K. Increased phosphorylation of collapsin response mediator protein-2 at Thr514 correlates with beta-amyloid burden and synaptic deficits in Lewy body dementias. Mol. Brain 2016, 9, 84. [Google Scholar] [CrossRef] [Green Version]
- Abdul-Hay, S.O.; Sahara, T.; McBride, M.; Kang, D.; Leissring, M.A. Identification of BACE2 as an avid ss-amyloid-degrading protease. Mol. Neurodegener. 2012, 7, 46. [Google Scholar] [CrossRef] [Green Version]
- Montibeller, L.; de Belleroche, J. Amyotrophic lateral sclerosis (ALS) and Alzheimer’s disease (AD) are characterised by differential activation of ER stress pathways: Focus on UPR target genes. Cell Stress Chaperones 2018, 23, 897–912. [Google Scholar] [CrossRef] [Green Version]
- Salminen, A.; Kauppinen, A.; Suuronen, T.; Kaarniranta, K.; Ojala, J. ER stress in Alzheimer’s disease: A novel neuronal trigger for inflammation and Alzheimer’s pathology. J. Neuroinflamm. 2009, 6, 41. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.C.; Liu, C.C.; Kanekiyo, T.; Xu, H.; Bu, G. Apolipoprotein E and Alzheimer disease: Risk, mechanisms and therapy. Nat. Rev. Neurol. 2013, 9, 106–118. [Google Scholar] [CrossRef] [Green Version]
- Miller, C.C.; McLoughlin, D.M.; Lau, K.F.; Tennant, M.E.; Rogelj, B. The X11 proteins, Abeta production and Alzheimer’s disease. Trends Neurosci. 2006, 29, 280–285. [Google Scholar] [CrossRef]
- Kim, J.; Miller, V.M.; Levites, Y.; West, K.J.; Zwizinski, C.W.; Moore, B.D.; Troendle, F.J.; Bann, M.; Verbeeck, C.; Price, R.W.; et al. BRI2 (ITM2b) inhibits Abeta deposition in vivo. J. Neurosci. 2008, 28, 6030–6036. [Google Scholar] [CrossRef] [PubMed]
- Delmas, E.; Jah, N.; Pirou, C.; Bouleau, S.; Le Floch, N.; Vayssiere, J.L.; Mignotte, B.; Renaud, F. FGF1 C-terminal domain and phosphorylation regulate intracrine FGF1 signaling for its neurotrophic and anti-apoptotic activities. Cell Death Dis. 2016, 7, e2079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coutellier, L.; Beraki, S.; Ardestani, P.M.; Saw, N.L.; Shamloo, M. Npas4: A neuronal transcription factor with a key role in social and cognitive functions relevant to developmental disorders. PLoS ONE 2012, 7, e46604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hunt, D.; Raivich, G.; Anderson, P.N. Activating transcription factor 3 and the nervous system. Front. Mol. Neurosci. 2012, 5, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warsito, D.; Sjostrom, S.; Andersson, S.; Larsson, O.; Sehat, B. Nuclear IGF1R is a transcriptional co-activator of LEF1/TCF. EMBO Rep. 2012, 13, 244–250. [Google Scholar] [CrossRef] [Green Version]
- Christopherson, K.S.; Ullian, E.M.; Stokes, C.C.; Mullowney, C.E.; Hell, J.W.; Agah, A.; Lawler, J.; Mosher, D.F.; Bornstein, P.; Barres, B.A. Thrombospondins are astrocyte-secreted proteins that promote CNS synaptogenesis. Cell 2005, 120, 421–433. [Google Scholar] [CrossRef] [Green Version]
- Jing, G.; Westwell-Roper, C.; Chen, J.; Xu, G.; Verchere, C.B.; Shalev, A. Thioredoxin-interacting protein promotes islet amyloid polypeptide expression through miR-124a and FoxA2. J. Biol. Chem. 2014, 289, 11807–11815. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.J.; Buchthal, B.; Lau, D.; Hayer, S.; Dick, O.; Schwaninger, M.; Veltkamp, R.; Zou, M.; Weiss, U.; Bading, H. A signaling cascade of nuclear calcium-CREB-ATF3 activated by synaptic NMDA receptors defines a gene repression module that protects against extrasynaptic NMDA receptor-induced neuronal cell death and ischemic brain damage. J. Neurosci. 2011, 31, 4978–4990. [Google Scholar] [CrossRef]
- Weng, J.; Luo, J.; Cheng, X.; Jin, C.; Zhou, X.; Qu, J.; Tu, L.; Ai, D.; Li, D.; Wang, J.; et al. Deletion of G protein-coupled receptor 48 leads to ocular anterior segment dysgenesis (ASD) through down-regulation of Pitx2. Proc. Natl. Acad. Sci. USA 2008, 105, 6081–6086. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Li, J.; Lea, R.; Vleminckx, K.; Amaya, E. Fezf2 promotes neuronal differentiation through localised activation of Wnt/beta-catenin signalling during forebrain development. Development 2014, 141, 4794–4805. [Google Scholar] [CrossRef] [Green Version]
- Zuccotti, A.; Le Magueresse, C.; Chen, M.; Neitz, A.; Monyer, H. The transcription factor Fezf2 directs the differentiation of neural stem cells in the subventricular zone toward a cortical phenotype. Proc. Natl. Acad. Sci. USA 2014, 111, 10726–10731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlingensiepen, K.H.; Wollnik, F.; Kunst, M.; Schlingensiepen, R.; Herdegen, T.; Brysch, W. The role of Jun transcription factor expression and phosphorylation in neuronal differentiation, neuronal cell death, and plastic adaptations in vivo. Cell Mol. Neurobiol. 1994, 14, 487–505. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Strohmeyer, R.; Liang, Z.; Lue, L.F.; Rogers, J. CCAAT/enhancer binding protein delta (C/EBPdelta) expression and elevation in Alzheimer’s disease. Neurobiol. Aging 2004, 25, 991–999. [Google Scholar] [CrossRef]
- Wang, S.M.; Lim, S.W.; Wang, Y.H.; Lin, H.Y.; Lai, M.D.; Ko, C.Y.; Wang, J.M. Astrocytic CCAAT/Enhancer-binding protein delta contributes to reactive oxygen species formation in neuroinflammation. Redox Biol. 2018, 16, 104–112. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Lin, F.; Zhuo, C.; Deng, G.; Chen, Z.; Yin, S.; Gao, Z.; Piccioni, M.; Tsun, A.; Cai, S.; et al. PIM1 kinase phosphorylates the human transcription factor FOXP3 at serine 422 to negatively regulate its activity under inflammation. J. Biol. Chem. 2014, 289, 26872–26881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rainio, E.M.; Sandholm, J.; Koskinen, P.J. Cutting edge: Transcriptional activity of NFATc1 is enhanced by the Pim-1 kinase. J. Immunol. 2002, 168, 1524–1527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Velazquez, R.; Shaw, D.M.; Caccamo, A.; Oddo, S. Pim1 inhibition as a novel therapeutic s trategy for Alzheimer’s disease. Mol. Neurodegener. 2016, 11, 52. [Google Scholar] [CrossRef] [Green Version]
- Zippo, A.; De Robertis, A.; Serafini, R.; Oliviero, S. PIM1-dependent phosphorylation of histone H3 at serine 10 is required for MYC-dependent transcriptional activation and oncogenic transformation. Nat. Cell Biol. 2007, 9, 932–944. [Google Scholar]
- Lee, A.K.; Potts, P.R. A Comprehensive Guide to the MAGE Family of Ubiquitin Ligases. J. Mol. Biol. 2017, 429, 1114–1142. [Google Scholar] [CrossRef] [Green Version]
- Coon, K.D.; Siegel, A.M.; Yee, S.J.; Dunckley, T.L.; Mueller, C.; Nagra, R.M.; Tourtellotte, W.W.; Reiman, E.M.; Papassotiropoulos, A.; Petersen, F.F.; et al. Preliminary demonstration of an allelic association of the IREB2 gene with Alzheimer’s disease. J. Alzheimers Dis. 2006, 9, 225–233. [Google Scholar] [CrossRef] [Green Version]
- Takahashi-Makise, N.; Ward, D.M.; Kaplan, J. On the mechanism of iron sensing by IRP2: New players, new paradigms. Nat. Chem. Biol. 2009, 5, 874–875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Von Bernhardi, R.; Cornejo, F.; Parada, G.E.; Eugenin, J. Role of TGFbeta signaling in the pathogenesis of Alzheimer’s disease. Front. Cell Neurosci. 2015, 9, 426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suenaga, K.; Lee, K.Y.; Nakamori, M.; Tatsumi, Y.; Takahashi, M.P.; Fujimura, H.; Jinnai, K.; Yoshikawa, H.; Du, H.; Ares, M., Jr.; et al. Muscleblind-like 1 knockout mice reveal novel splicing defects in the myotonic dystrophy brain. PLoS ONE 2012, 7, e33218. [Google Scholar] [CrossRef]
- Bluthgen, N.; van Bentum, M.; Merz, B.; Kuhl, D.; Hermey, G. Profiling the MAPK/ERK dependent and independent activity regulated transcriptional programs in the murine hippocampus in vivo. Sci. Rep. 2017, 7, 45101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Namwanje, M.; Brown, C.W. Activins and Inhibins: Roles in Development, Physiology, and Disease. Cold Spring Harb. Perspect. Biol. 2016, 8, a021881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Wei, W.; Zhao, Q.Y.; Widagdo, J.; Baker-Andresen, D.; Flavell, C.R.; D’Alessio, A.; Zhang, Y.; Bredy, T.W. Neocortical Tet3-mediated accumulation of 5-hydroxymethylcytosine promotes rapid behavioral adaptation. Proc. Natl. Acad. Sci. USA 2014, 111, 7120–7125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perera, A.; Eisen, D.; Wagner, M.; Laube, S.K.; Kunzel, A.F.; Koch, S.; Steinbacher, J.; Schulze, E.; Splith, V.; Mittermeier, N.; et al. TET3 is recruited by REST for context-specific hydroxymethylation and induction of gene expression. Cell Rep. 2015, 11, 283–294. [Google Scholar] [CrossRef] [Green Version]
- Farley, J.E.; Burdett, T.C.; Barria, R.; Neukomm, L.J.; Kenna, K.P.; Landers, J.E.; Freeman, M.R. Transcription factor Pebbled/RREB1 regulates injury-induced axon degeneration. Proc. Natl. Acad. Sci. USA 2018, 115, 1358–1363. [Google Scholar] [CrossRef] [Green Version]
- Cates, H.M.; Thibault, M.; Pfau, M.; Heller, E.; Eagle, A.; Gajewski, P.; Bagot, R.; Colangelo, C.; Abbott, T.; Rudenko, G.; et al. Threonine 149 phosphorylation enhances DeltaFosB transcriptional activity to control psychomotor responses to cocaine. J. Neurosci. 2014, 34, 11461–11469. [Google Scholar] [CrossRef] [Green Version]
- Eagle, A.L.; Gajewski, P.A.; Yang, M.; Kechner, M.E.; Al Masraf, B.S.; Kennedy, P.J.; Wang, H.; Mazei-Robison, M.S.; Robison, A.J. Experience-Dependent Induction of Hippocampal DeltaFosB Controls Learning. J. Neurosci. 2015, 35, 13773–13783. [Google Scholar] [CrossRef] [Green Version]
- Grassi, D.; Franz, H.; Vezzali, R.; Bovio, P.; Heidrich, S.; Dehghanian, F.; Lagunas, N.; Belzung, C.; Krieglstein, K.; Vogel, T. Neuronal Activity, TGFbeta-Signaling and Unpredictable Chronic Stress Modulate Transcription of Gadd45 Family Members and DNA Methylation in the Hippocampus. Cereb. Cortex 2017, 27, 4166–4181. [Google Scholar] [CrossRef] [PubMed]
- Tamai, S.; Imaizumi, K.; Kurabayashi, N.; Nguyen, M.D.; Abe, T.; Inoue, M.; Fukada, Y.; Sanada, K. Neuroprotective role of the basic leucine zipper transcription factor NFIL3 in models of amyotrophic lateral sclerosis. J. Biol. Chem. 2014, 289, 1629–1638. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Cho, K.; Singer, B.H.; Zhang, H. The nuclear transcription factor PKNOX2 is a candidate gene for substance dependence in European-origin women. PLoS ONE 2011, 6, e16002. [Google Scholar] [CrossRef]
- Wang, K.S.; Zhang, Q.; Liu, X.; Wu, L.; Zeng, M. PKNOX2 is associated with formal thought disorder in schizophrenia: A meta-analysis of two genome-wide association studies. J. Mol. Neurosci. 2012, 48, 265–272. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Lawson, M.A.; Melamed, P. A proteomic comparison of immature and mature mouse gonadotrophs reveals novel differentially expressed nuclear proteins that regulate gonadotropin gene transcription and RNA splicing. Biol. Reprod. 2008, 79, 546–561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lang, B.; Alrahbeni, T.M.; Clair, D.S.; Blackwood, D.H.; International Schizophrenia Consortium; McCaig, C.D.; Shen, S. HDAC9 is implicated in schizophrenia and expressed specifically in post-mitotic neurons but not in adult neural stem cells. Am. J. Stem Cells 2012, 1, 31–41. [Google Scholar]
- Lund, I.V.; Hu, Y.; Raol, Y.H.; Benham, R.S.; Faris, R.; Russek, S.J.; Brooks-Kayal, A.R. BDNF selectively regulates GABAA receptor transcription by activation of the JAK/STAT pathway. Sci Signal. 2008, 1, ra9. [Google Scholar] [CrossRef] [Green Version]
- Bayam, E.; Sahin, G.S.; Guzelsoy, G.; Guner, G.; Kabakcioglu, A.; Ince-Dunn, G. Genome-wide target analysis of NEUROD2 provides new insights into regulation of cortical projection neuron migration and differentiation. BMC Genomics 2015, 16, 681. [Google Scholar] [CrossRef]
- Chen, F.; Moran, J.T.; Zhang, Y.; Ates, K.M.; Yu, D.; Schrader, L.A.; Das, P.M.; Jones, F.E.; Hall, B.J. The transcription factor NeuroD2 coordinates synaptic innervation and cell intrinsic properties to control excitability of cortical pyramidal neurons. J. Physiol. 2016, 594, 3729–3744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, C.H.; Hansen, S.; Wang, Z.; Storm, D.R.; Tapscott, S.J.; Olson, J.M. The dosage of the neuroD2 transcription factor regulates amygdala development and emotional learning. Proc. Natl. Acad. Sci. USA 2005, 102, 14877–14882. [Google Scholar] [CrossRef] [Green Version]
- Agundez, J.A.; Garcia-Martin, E.; Martinez, C.; Benito-Leon, J.; Millan-Pascual, J.; Diaz-Sanchez, M.; Calleja, P.; Pisa, D.; Turpin-Fenoll, L.; Alonso-Navarro, H.; et al. Heme Oxygenase-1 and 2 Common Genetic Variants and Risk for Multiple Sclerosis. Sci. Rep. 2016, 6, 20830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ayuso, P.; Martinez, C.; Pastor, P.; Lorenzo-Betancor, O.; Luengo, A.; Jimenez-Jimenez, F.J.; Alonso-Navarro, H.; Agundez, J.A.; Garcia-Martin, E. An association study between Heme oxygenase-1 genetic variants and Parkinson’s disease. Front. Cell Neurosci. 2014, 8, 298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Q.; Weis, S.; Yang, G.; Weng, Y.H.; Helston, R.; Rish, K.; Smith, A.; Bordner, J.; Polte, T.; Gaunitz, F.; et al. Heme oxygenase-1 protein localizes to the nucleus and activates transcription factors important in oxidative stress. J. Biol. Chem. 2007, 282, 20621–20633. [Google Scholar] [CrossRef] [Green Version]
- Schipper, H.M.; Song, W. A heme oxygenase-1 transducer model of degenerative and developmental brain disorders. Int. J. Mol. Sci 2015, 16, 5400–5419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crespo-Barreto, J.; Fryer, J.D.; Shaw, C.A.; Orr, H.T.; Zoghbi, H.Y. Partial loss of ataxin-1 function contributes to transcriptional dysregulation in spinocerebellar ataxia type 1 pathogenesis. PLoS Genet. 2010, 6, e1001021. [Google Scholar] [CrossRef]
- Lu, H.C.; Tan, Q.; Rousseaux, M.W.; Wang, W.; Kim, J.Y.; Richman, R.; Wan, Y.W.; Yeh, S.Y.; Patel, J.M.; Liu, X.; et al. Disruption of the ATXN1-CIC complex causes a spectrum of neurobehavioral phenotypes in mice and humans. Nat. Genet. 2017, 49, 527–536. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, A.C.; Javidfar, B.; Pothula, V.; Ibi, D.; Shen, E.Y.; Peter, C.J.; Bicks, L.K.; Fehr, T.; Jiang, Y.; Brennand, K.J.; et al. MEF2C transcription factor is associated with the genetic and epigenetic risk architecture of schizophrenia and improves cognition in mice. Mol. Psychiatry 2018, 23, 123–132. [Google Scholar] [CrossRef] [Green Version]
- Herrera, F.J.; Yamaguchi, T.; Roelink, H.; Tjian, R. Core promoter factor TAF9B regulates neuronal gene expression. eLife 2014, 3, e02559. [Google Scholar] [CrossRef]
- Auderset, L.; Landowski, L.M.; Foa, L.; Young, K.M. Low Density Lipoprotein Receptor Related Proteins as Regulators of Neural Stem and Progenitor Cell Function. Stem Cells Int. 2016, 2016, 2108495. [Google Scholar] [CrossRef] [Green Version]
- Shah, M.; Baterina, O.Y., Jr.; Taupin, V.; Farquhar, M.G. ARH directs megalin to the endocytic recycling compartment to regulate its proteolysis and gene expression. J. Cell Biol. 2013, 202, 113–127. [Google Scholar] [CrossRef] [Green Version]
- Spuch, C.; Ortolano, S.; Navarro, C. LRP-1 and LRP-2 receptors function in the membrane neuron. Trafficking mechanisms and proteolytic processing in Alzheimer’s disease. Front. Physiol. 2012, 3, 269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santra, M.; Chopp, M.; Zhang, Z.G.; Lu, M.; Santra, S.; Nalani, A.; Santra, S.; Morris, D.C. Thymosin beta 4 mediates oligodendrocyte differentiation by upregulating p38 MAPK. Glia 2012, 60, 1826–1838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carrillo-Garcia, C.; Prochnow, S.; Simeonova, I.K.; Strelau, J.; Holzl-Wenig, G.; Mandl, C.; Unsicker, K.; von Bohlen Und Halbach, O.; Ciccolini, F. Growth/differentiation factor 15 promotes EGFR signalling, and regulates proliferation and migration in the hippocampus of neonatal and young adult mice. Development 2014, 141, 773–783. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Omori, Y.; Onishi, A.; Sato, S.; Kondo, M.; Furukawa, T. Blimp1 suppresses Chx10 expression in differentiating retinal photoreceptor precursors to ensure proper photoreceptor development. J. Neurosci. 2010, 30, 6515–6526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kinameri, E.; Inoue, T.; Aruga, J.; Imayoshi, I.; Kageyama, R.; Shimogori, T.; Moore, A.W. Prdm proto-oncogene transcription factor family expression and interaction with the Notch-Hes pathway in mouse neurogenesis. PLoS ONE 2008, 3, e3859. [Google Scholar] [CrossRef] [Green Version]
- Yu, M.; Fu, Y.; Liang, Y.; Song, H.; Yao, Y.; Wu, P.; Yao, Y.; Pan, Y.; Wen, X.; Ma, L.; et al. Suppression of MAPK11 or HIPK3 reduces mutant Huntingtin levels in Huntington’s disease models. Cell Res. 2017, 27, 1441–1465. [Google Scholar] [CrossRef]
- Knoepfler, P.S.; Cheng, P.F.; Eisenman, R.N. N-myc is essential during neurogenesis for the rapid expansion of progenitor cell populations and the inhibition of neuronal differentiation. Genes Dev. 2002, 16, 2699–2712. [Google Scholar] [CrossRef] [Green Version]
- Zinin, N.; Adameyko, I.; Wilhelm, M.; Fritz, N.; Uhlen, P.; Ernfors, P.; Henriksson, M.A. MYC proteins promote neuronal differentiation by controlling the mode of progenitor cell division. EMBO Rep. 2014, 15, 383–391. [Google Scholar] [CrossRef]
- Ancin, I.; Cabranes, J.A.; Vazquez-Alvarez, B.; Santos, J.L.; Sanchez-Morla, E.; Alaerts, M.; Del-Favero, J.; Barabash, A. NR4A2: Effects of an "orphan" receptor on sustained attention in a schizophrenic population. Schizophr. Bull. 2013, 39, 555–563. [Google Scholar] [CrossRef] [Green Version]
- Buervenich, S.; Carmine, A.; Arvidsson, M.; Xiang, F.; Zhang, Z.; Sydow, O.; Jonsson, E.G.; Sedvall, G.C.; Leonard, S.; Ross, R.G.; et al. NURR1 mutations in cases of schizophrenia and manic-depressive disorder. Am. J. Med. Genet. 2000, 96, 808–813. [Google Scholar] [CrossRef]
- Jacobs, F.M.; van der Linden, A.J.; Wang, Y.; von Oerthel, L.; Sul, H.S.; Burbach, J.P.; Smidt, M.P. Identification of Dlk1, Ptpru and Klhl1 as novel Nurr1 target genes in meso-diencephalic dopamine neurons. Development 2009, 136, 2363–2373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, A.; Masgutova, G.; Collin, A.; Toch, M.; Hidalgo-Figueroa, M.; Jacob, B.; Corcoran, L.M.; Francius, C.; Clotman, F. Onecut Factors and Pou2f2 Regulate the Distribution of V2 Interneurons in the Mouse Developing Spinal Cord. Front. Cell Neurosci. 2019, 13, 184. [Google Scholar] [CrossRef] [PubMed]
- Lillycrop, K.A.; Latchman, D.S. Alternative splicing of the Oct-2 transcription factor RNA is differentially regulated in neuronal cells and B cells and results in protein isoforms with opposite effects on the activity of octamer/TAATGARAT-containing promoters. J. Biol. Chem. 1992, 267, 24960–24965. [Google Scholar] [CrossRef]
- Wang, Y.T.; Linden, D.J. Expression of cerebellar long-term depression requires postsynaptic clathrin-mediated endocytosis. Neuron 2000, 25, 635–647. [Google Scholar] [CrossRef] [Green Version]
- Fowler, C.J. The contribution of cyclooxygenase-2 to endocannabinoid metabolism and action. Br. J. Pharmacol. 2007, 152, 594–601. [Google Scholar] [CrossRef] [Green Version]
- Aid, S.; Bosetti, F. Targeting cyclooxygenases-1 and -2 in neuroinflammation: Therapeutic implications. Biochimie 2011, 93, 46–51. [Google Scholar] [CrossRef] [Green Version]
- Woodling, N.S.; Colas, D.; Wang, Q.; Minhas, P.; Panchal, M.; Liang, X.; Mhatre, S.D.; Brown, H.; Ko, N.; Zagol-Ikapitte, I.; et al. Cyclooxygenase inhibition targets neurons to prevent early behavioural decline in Alzheimer’s disease model mice. Brain 2016, 139, 2063–2081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raivich, G. c-Jun expression, activation and function in neural cell death, inflammation and repair. J. Neurochem. 2008, 107, 898–906. [Google Scholar] [CrossRef]
- Savage, M.J.; Lin, Y.G.; Ciallella, J.R.; Flood, D.G.; Scott, R.W. Activation of c-Jun N-terminal kinase and p38 in an Alzheimer’s disease model is associated with amyloid deposition. J. Neurosci. 2002, 22, 3376–3385. [Google Scholar] [CrossRef] [Green Version]
- Yarza, R.; Vela, S.; Solas, M.; Ramirez, M.J. c-Jun N-terminal Kinase (JNK) Signaling as a Therapeutic Target for Alzheimer’s Disease. Front. Pharmacol. 2015, 6, 321. [Google Scholar] [CrossRef] [Green Version]
- Jiang, C.; Zou, X.; Zhu, R.; Shi, Y.; Wu, Z.; Zhao, F.; Chen, L. The correlation between accumulation of amyloid beta with enhanced neuroinflammation and cognitive impairment after intraventricular hemorrhage. J. Neurosurg. 2018, 131, 54–63. [Google Scholar] [CrossRef] [PubMed]
- Broadhead, M.J.; Horrocks, M.H.; Zhu, F.; Muresan, L.; Benavides-Piccione, R.; DeFelipe, J.; Fricker, D.; Kopanitsa, M.V.; Duncan, R.R.; Klenerman, D.; et al. PSD95 nanoclusters are postsynaptic building blocks in hippocampus circuits. Sci. Rep. 2016, 6, 24626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Craven, S.E.; El-Husseini, A.E.; Bredt, D.S. Synaptic targeting of the postsynaptic density protein PSD-95 mediated by lipid and protein motifs. Neuron 1999, 22, 497–509. [Google Scholar] [CrossRef] [Green Version]
- Medrihan, L.; Cesca, F.; Raimondi, A.; Lignani, G.; Baldelli, P.; Benfenati, F. Synapsin II desynchronizes neurotransmitter release at inhibitory synapses by interacting with presynaptic calcium channels. Nat. Commun. 2013, 4, 1512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Feng, D.; Liu, G.; Luo, Q.; Xu, Y.; Lin, S.; Fei, J.; Xu, L. Gamma-aminobutyric acid transporter 1 negatively regulates T cell-mediated immune responses and ameliorates autoimmune inflammation in the CNS. J. Immunol. 2008, 181, 8226–8236. [Google Scholar] [CrossRef] [Green Version]
- Bodnar, R.J. Endogenous Opiates and Behavior: 2016. Peptides 2018, 101, 167–212. [Google Scholar] [CrossRef]
- Kaplan, G.B.; Leite-Morris, K.A.; Fan, W.; Young, A.J.; Guy, M.D. Opiate sensitization induces FosB/DeltaFosB expression in prefrontal cortical, striatal and amygdala brain regions. PLoS ONE 2011, 6, e23574. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.J.; Jiang, M.; Zhou, H.; Liu, W.; Wang, C.; Kang, Z.; Han, B.; Zhang, Q.; Chen, X.; Xiao, J.; et al. TLR-stimulated IRAKM activates caspase-8 inflammasome in microglia and promotes neuroinflammation. J. Clin. Investig. 2018, 128, 5399–5412. [Google Scholar] [CrossRef]
- Crowley, T.; Cryan, J.F.; Downer, E.J.; O’Leary, O.F. Inhibiting neuroinflammation: The role and therapeutic potential of GABA in neuro-immune interactions. Brain Behav. Immun. 2016, 54, 260–277. [Google Scholar] [CrossRef]
- Haerian, B.S.; Baum, L.; Kwan, P.; Cherny, S.S.; Shin, J.G.; Kim, S.E.; Han, B.G.; Tan, H.J.; Raymond, A.A.; Tan, C.T.; et al. Contribution of GABRG2 Polymorphisms to Risk of Epilepsy and Febrile Seizure: A Multicenter Cohort Study and Meta-analysis. Mol. Neurobiol. 2016, 53, 5457–5467. [Google Scholar] [CrossRef]
- Ninan, I. Synaptic regulation of affective behaviors; role of BDNF. Neuropharmacology 2014, 76 Pt C, 684–695. [Google Scholar] [CrossRef] [Green Version]
- Cunha, C.; Brambilla, R.; Thomas, K.L. A simple role for BDNF in learning and memory? Front. Mol. Neurosci. 2010, 3, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, R.; Liu, Z.; Sun, N.; Liu, S.; Li, L.; Shen, Y.; Xiu, J.; Xu, Q. BDNF Alleviates Neuroinflammation in the Hippocampus of Type 1 Diabetic Mice via Blocking the Aberrant HMGB1/RAGE/NF-kappaB Pathway. Aging Dis. 2019, 10, 611–625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giuffrida, M.L.; Copani, A.; Rizzarelli, E. A promising connection between BDNF and Alzheimer’s disease. Aging 2018, 10, 1791–1792. [Google Scholar] [CrossRef] [PubMed]
- Charbogne, P.; Kieffer, B.L.; Befort, K. 15 years of genetic approaches in vivo for addiction research: Opioid receptor and peptide gene knockout in mouse models of drug abuse. Neuropharmacology 2014, 76 Pt B, 204–217. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Yin, C.; Zhang, T.; Li, F.; Yang, W.; Kaminski, R.; Fagan, P.R.; Putatunda, R.; Young, W.B.; Khalili, K.; et al. CRISPR/gRNA-directed synergistic activation mediator (SAM) induces specific, persistent and robust reactivation of the HIV-1 latent reservoirs. Sci. Rep. 2015, 5, 16277. [Google Scholar] [CrossRef] [PubMed]
- Seiliez, I.; Thisse, B.; Thisse, C. FoxA3 and goosecoid promote anterior neural fate through inhibition of Wnt8a activity before the onset of gastrulation. Dev. Biol. 2006, 290, 152–163. [Google Scholar] [CrossRef]
- Dal-Pra, S.; Thisse, C.; Thisse, B. FoxA transcription factors are essential for the development of dorsal axial structures. Dev. Biol. 2011, 350, 484–495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsieh, H.; Boehm, J.; Sato, C.; Iwatsubo, T.; Tomita, T.; Sisodia, S.; Malinow, R. AMPAR removal underlies Abeta-induced synaptic depression and dendritic spine loss. Neuron 2006, 52, 831–843. [Google Scholar] [CrossRef] [Green Version]
- Knobloch, M.; Mansuy, I.M. Dendritic spine loss and synaptic alterations in Alzheimer’s disease. Mol. Neurobiol. 2008, 37, 73–82. [Google Scholar] [CrossRef] [Green Version]
- Alberi, L.; Liu, S.; Wang, Y.; Badie, R.; Smith-Hicks, C.; Wu, J.; Pierfelice, T.J.; Abazyan, B.; Mattson, M.P.; Kuhl, D.; et al. Activity-induced Notch signaling in neurons requires Arc/Arg3.1 and is essential for synaptic plasticity in hippocampal networks. Neuron 2011, 69, 437–444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, Y.-H.; Ann, E.-J.; Yoon, J.-H.; Mo, J.-S.; Kim, M.-Y.; Park, H.-S. Calcium/calmodulin-dependent protein kinase IV (CaMKIV) enhances osteoclast differentiation via the up-regulation of Notch1 protein stability. Biochim. Et Biophys. Acta (BBA) Mol. Cell Res. 2013, 1833, 69–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, D.; Long, J.; Dai, F.; Liang, M.; Feng, X.-H.; Lin, X. BCL6 Represses Smad Signaling in Transforming Growth Factor-β Resistance. Cancer Res. 2008, 68, 783–789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, X.; Ito, A.; Kane, C.D.; Liao, T.-S.; Bolger, T.A.; Lemrow, S.M.; Means, A.R.; Yao, T.-P. The Modular Nature of Histone Deacetylase HDAC4 Confers Phosphorylation-dependent Intracellular Trafficking. J. Biol. Chem. 2001, 276, 35042–35048. [Google Scholar] [PubMed] [Green Version]
- Graziani, I.; Eliasz, S.; De Marco, M.A.; Chen, Y.; Pass, H.I.; De May, R.M.; Strack, P.R.; Miele, L.; Bocchetta, M. Opposite effects of Notch-1 and Notch-2 on mesothelioma cell survival under hypoxia are exerted through the Akt pathway. Cancer Res. 2008, 68, 9678–9685. [Google Scholar] [CrossRef] [Green Version]
- Fang, L.; Zhang, L.; Wei, W.; Jin, X.; Wang, P.; Tong, Y.; Li, J.; Du, J.X.; Wong, J. A methylation-phosphorylation switch determines Sox2 stability and function in ESC maintenance or differentiation. Mol. Cell 2014, 55, 537–551. [Google Scholar] [CrossRef] [Green Version]
- Jeong, C.H.; Cho, Y.Y.; Kim, M.O.; Kim, S.H.; Cho, E.J.; Lee, S.Y.; Jeon, Y.J.; Lee, K.Y.; Yao, K.; Keum, Y.S.; et al. Phosphorylation of Sox2 cooperates in reprogramming to pluripotent stem cells. Stem Cells 2010, 28, 2141–2150. [Google Scholar] [CrossRef]
- Zhou, L.; Zhang, N.; Song, W.; You, N.; Li, Q.; Sun, W.; Zhang, Y.; Wang, D.; Dou, K. The significance of Notch1 compared with Notch3 in high metastasis and poor overall survival in hepatocellular carcinoma. PLoS ONE 2013, 8, e57382. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.-M.; Jaskula-Sztul, R.; Georgen, M.R.; Aburjania, Z.; Somnay, Y.R.; Leverson, G.; Sippel, R.S.; Lloyd, R.V.; Johnson, B.P.; Chen, H. Notch1 Signaling Regulates the Aggressiveness of Differentiated Thyroid Cancer and Inhibits SERPINE1 Expression. Clin. Cancer Res. 2016, 22, 3582–3592. [Google Scholar] [CrossRef] [Green Version]
- Dekker, R.J.; Pannekoek, H.; Horrevoets, A.J.G. A steady-state competition model describes the modulating effects of thrombomodulin on thrombin inhibition by plasminogen activator inhibitor-1 in the absence and presence of vitronectin. Eur. J. Biochem. 2003, 270, 1942–1951. [Google Scholar] [CrossRef] [Green Version]
- Idogawa, M.; Masutani, M.; Shitashige, M.; Honda, K.; Tokino, T.; Shinomura, Y.; Imai, K.; Hirohashi, S.; Yamada, T. Ku70 and Poly(ADP-Ribose) Polymerase-1 Competitively Regulate β-Catenin and T-Cell Factor-4–Mediated Gene Transactivation: Possible Linkage of DNA Damage Recognition and Wnt Signaling. Cancer Res. 2007, 67, 911–918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hicks, C.; Pannuti, A.; Miele, L. Associating GWAS Information with the Notch Signaling Pathway Using Transcription Profiling. Cancer Inform. 2011, 10, CIN.S6072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jehn, B.M.; Bielke, W.; Pear, W.S.; Osborne, B.A. Cutting edge: Protective effects of notch-1 on TCR-induced apoptosis. J. Immunol. 1999, 162, 635–638. [Google Scholar]
- Kurakula, K.; Vos, M.; Logiantara, A.; Roelofs, J.J.; Nieuwenhuis, M.A.; Koppelman, G.H.; Postma, D.S.; van Rijt, L.S.; de Vries, C.J. Nuclear Receptor Nur77 Attenuates Airway Inflammation in Mice by Suppressing NF-kappaB Activity in Lung Epithelial Cells. J. Immunol. 2015, 195, 1388–1398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sade, H.; Krishna, S.; Sarin, A. The anti-apoptotic effect of Notch-1 requires p56lck-dependent, Akt/PKB-mediated signaling in T cells. J. Biol. Chem. 2004, 279, 2937–2944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaguchi, N.; Yamaguchi, N. The seventh zinc finger motif of A20 is required for the suppression of TNF-alpha-induced apoptosis. FEBS Lett. 2015, 589, 1369–1375. [Google Scholar] [CrossRef] [Green Version]
- Tapias, A.; Wang, Z.Q. Lysine Acetylation and Deacetylation in Brain Development and Neuropathies. Genom. Proteom. Bioinform. 2017, 15, 19–36. [Google Scholar] [CrossRef]
- Xu, S.; Wilf, R.; Menon, T.; Panikker, P.; Sarthi, J.; Elefant, F. Epigenetic control of learning and memory in Drosophila by Tip60 HAT action. Genetics 2014, 198, 1571–1586. [Google Scholar] [CrossRef] [Green Version]
- Pirooznia, S.K.; Sarthi, J.; Johnson, A.A.; Toth, M.S.; Chiu, K.; Koduri, S.; Elefant, F. Tip60 HAT activity mediates APP induced lethality and apoptotic cell death in the CNS of a Drosophila Alzheimer’s disease model. PLoS ONE 2012, 7, e41776. [Google Scholar] [CrossRef] [Green Version]
- Ai, W.; Zheng, H.; Yang, X.; Liu, Y.; Wang, T.C. Tip60 functions as a potential corepressor of KLF4 in regulation of HDC promoter activity. Nucleic Acids Res. 2007, 35, 6137–6149. [Google Scholar] [CrossRef] [Green Version]
- Jang, S.H.; Chen, H.; Gregersen, P.K.; Diamond, B.; Kim, S.J. Kruppel-like factor4 regulates PRDM1 expression through binding to an autoimmune risk allele. JCI Insight 2017, 2, e89569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, H.; Guo, M.; Xu, D.; Ding, Z.C.; Zhou, G.; Ding, H.F.; Zhang, J.; Tang, Y.; Yan, C. The stress-responsive gene ATF3 regulates the histone acetyltransferase Tip60. Nat. Commun. 2015, 6, 6752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, H.; Li, X.; Han, C.; Wang, Q.E.; Wang, H.; Ding, H.F.; Zhang, J.; Yan, C. The Stress-responsive Gene ATF3 Mediates Dichotomous UV Responses by Regulating the Tip60 and p53 Proteins. J. Biol. Chem. 2016, 291, 10847–10857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darlyuk-Saadon, I.; Weidenfeld-Baranboim, K.; Yokoyama, K.K.; Hai, T.; Aronheim, A. The bZIP repressor proteins, c-Jun dimerization protein 2 and activating transcription factor 3, recruit multiple HDAC members to the ATF3 promoter. Biochim. Biophys. Acta 2012, 1819, 1142–1153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolger, T.A.; Zhao, X.; Cohen, T.J.; Tsai, C.C.; Yao, T.P. The neurodegenerative disease protein ataxin-1 antagonizes the neuronal survival function of myocyte enhancer factor-2. J. Biol. Chem. 2007, 282, 29186–29192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serra, H.G.; Duvick, L.; Zu, T.; Carlson, K.; Stevens, S.; Jorgensen, N.; Lysholm, A.; Burright, E.; Zoghbi, H.Y.; Clark, H.B.; et al. RORα-Mediated Purkinje Cell Development Determines Disease Severity in Adult SCA1 Mice. Cell 2006, 127, 697–708. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.W.; Jang, S.M.; Kim, C.H.; An, J.H.; Kang, E.J.; Choi, K.H. New molecular bridge between RelA/p65 and NF-kappaB target genes via histone acetyltransferase TIP60 cofactor. J. Biol. Chem. 2012, 287, 7780–7791. [Google Scholar] [CrossRef] [Green Version]
- Koh, D.I.; Han, D.; Ryu, H.; Choi, W.I.; Jeon, B.N.; Kim, M.K.; Kim, Y.; Kim, J.Y.; Parry, L.; Clarke, A.R.; et al. KAISO, a critical regulator of p53-mediated transcription of CDKN1A and apoptotic genes. Proc. Natl. Acad. Sci. USA 2014, 111, 15078–15083. [Google Scholar] [CrossRef] [Green Version]
- Del Valle-Perez, B.; Casagolda, D.; Lugilde, E.; Valls, G.; Codina, M.; Dave, N.; de Herreros, A.G.; Dunach, M. Wnt controls the transcriptional activity of Kaiso through CK1epsilon-dependent phosphorylation of p120-catenin. J. Cell Sci. 2011, 124, 2298–2309. [Google Scholar] [CrossRef] [Green Version]
- Nagl, N.G., Jr.; Wang, X.; Patsialou, A.; Van Scoy, M.; Moran, E. Distinct mammalian SWI/SNF chromatin remodeling complexes with opposing roles in cell-cycle control. EMBO J. 2007, 26, 752–763. [Google Scholar] [CrossRef] [Green Version]
- Kim, B.R.; Coyaud, E.; Laurent, E.M.N.; St-Germain, J.; Van de Laar, E.; Tsao, M.S.; Raught, B.; Moghal, N. Identification of the SOX2 Interactome by BioID Reveals EP300 as a Mediator of SOX2—Dependent Squamous Differentiation and Lung Squamous Cell Carcinoma Growth. Mol. Cell Proteom. 2017, 16, 1864–1888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singhal, N.; Graumann, J.; Wu, G.; Arauzo-Bravo, M.J.; Han, D.W.; Greber, B.; Gentile, L.; Mann, M.; Scholer, H.R. Chromatin-Remodeling Components of the BAF Complex Facilitate Reprogramming. Cell 2010, 141, 943–955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krolewski, R.C.; Packard, A.; Jang, W.; Wildner, H.; Schwob, J.E. Ascl1 (Mash1) knockout perturbs differentiation of nonneuronal cells in olfactory epithelium. PLoS ONE 2012, 7, e51737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borromeo, M.D.; Meredith, D.M.; Castro, D.S.; Chang, J.C.; Tung, K.C.; Guillemot, F.; Johnson, J.E. A transcription factor network specifying inhibitory versus excitatory neurons in the dorsal spinal cord. Development 2014, 141, 2803–2812. [Google Scholar] [CrossRef] [Green Version]
- Brouillette, J.; Caillierez, R.; Zommer, N.; Alves-Pires, C.; Benilova, I.; Blum, D.; De Strooper, B.; Buee, L. Neurotoxicity and memory deficits induced by soluble low-molecular-weight amyloid-beta1–42 oligomers are revealed in vivo by using a novel animal model. J. Neurosci. 2012, 32, 7852–7861. [Google Scholar] [CrossRef] [Green Version]
- Paroni, G.; Mizzau, M.; Henderson, C.; Del Sal, G.; Schneider, C.; Brancolini, C. Caspase-dependent regulation of histone deacetylase 4 nuclear-cytoplasmic shuttling promotes apoptosis. Mol. Biol. Cell 2004, 15, 2804–2818. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.C.; Hsu, W.H.; Wang, C.C.; Chou, C.H.; Kuo, M.Y.; Lin, B.R.; Chen, S.T.; Tai, S.K.; Kuo, M.L.; Yang, M.H. Connective tissue growth factor activates pluripotency genes and mesenchymal-epithelial transition in head and neck cancer cells. Cancer Res. 2013, 73, 4147–4157. [Google Scholar] [CrossRef] [Green Version]
- Medeiros, R.; Prediger, R.D.; Passos, G.F.; Pandolfo, P.; Duarte, F.S.; Franco, J.L.; Dafre, A.L.; Di Giunta, G.; Figueiredo, C.P.; Takahashi, R.N.; et al. Connecting TNF-alpha signaling pathways to iNOS expression in a mouse model of Alzheimer’s disease: Relevance for the behavioral and synaptic deficits induced by amyloid beta protein. J. Neurosci. 2007, 27, 5394–5404. [Google Scholar]
- Manns, J.; Rico, M.; Mason, L.L.; Raul, D.L.C.A. Thrombospondin-1 (TSP1) Promotes Thrombin Generation on the Surface of Fibroblasts (HS-68) and Induces Up-Regulation of Connective Tissue Growth Factor (CTGF) Gene and Protein Expression. Blood 2006, 108, 1755. [Google Scholar] [CrossRef]
- Son, S.M.; Nam, D.W.; Cha, M.Y.; Kim, K.H.; Byun, J.; Ryu, H.; Mook-Jung, I. Thrombospondin-1 prevents amyloid beta-mediated synaptic pathology in Alzheimer’s disease. Neurobiol. Aging 2015, 36, 3214–3227. [Google Scholar] [CrossRef]
- Cao, X.; Sudhof, T.C. A transcriptionally active complex of APP with Fe65 and histone acetyltransferase Tip60. Science 2001, 293, 115–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Del Prete, D.; Rice, R.C.; Rajadhyaksha, A.M.; D’Adamio, L. Amyloid Precursor Protein (APP) May Act as a Substrate and a Recognition Unit for CRL4CRBN and Stub1 E3 Ligases Facilitating Ubiquitination of Proteins Involved in Presynaptic Functions and Neurodegeneration. J. Biol. Chem. 2016, 291, 17209–17227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fritsch, J.; Stephan, M.; Tchikov, V.; Winoto-Morbach, S.; Gubkina, S.; Kabelitz, D.; Schutze, S. Cell fate decisions regulated by K63 ubiquitination of tumor necrosis factor receptor 1. Mol. Cell Biol. 2014, 34, 3214–3228. [Google Scholar] [CrossRef] [Green Version]
- Marblestone, J.G.; Butt, S.; McKelvey, D.M.; Sterner, D.E.; Mattern, M.R.; Nicholson, B.; Eddins, M.J. Comprehensive ubiquitin E2 profiling of ten ubiquitin E3 ligases. Cell Biochem. Biophys. 2013, 67, 161–167. [Google Scholar] [CrossRef]
- Belova, L.; Sharma, S.; Brickley, D.R.; Nicolarsen, J.R.; Patterson, C.; Conzen, S.D. Ubiquitin-proteasome degradation of serum- and glucocorticoid-regulated kinase-1 (SGK-1) is mediated by the chaperone-dependent E3 ligase CHIP. Biochem. J. 2006, 400, 235–244. [Google Scholar] [CrossRef]
- Bogusz, A.M.; Brickley, D.R.; Pew, T.; Conzen, S.D. A novel N-terminal hydrophobic motif mediates constitutive degradation of serum- and glucocorticoid-induced kinase-1 by the ubiquitin-proteasome pathway. FEBS J. 2006, 273, 2913–2928. [Google Scholar] [CrossRef] [PubMed]
- Gao, D.; Wan, L.; Inuzuka, H.; Berg, A.H.; Tseng, A.; Zhai, B.; Shaik, S.; Bennett, E.; Tron, A.E.; Gasser, J.A.; et al. Rictor forms a complex with Cullin-1 to promote SGK1 ubiquitination and destruction. Mol. Cell 2010, 39, 797–808. [Google Scholar] [CrossRef] [Green Version]
- Mishra, A.; Dikshit, P.; Purkayastha, S.; Sharma, J.; Nukina, N.; Jana, N.R. E6-AP promotes misfolded polyglutamine proteins for proteasomal degradation and suppresses polyglutamine protein aggregation and toxicity. J. Biol. Chem. 2008, 283, 7648–7656. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Macfarlan, T.; Pittman, R.N.; Chakravarti, D. Ataxin-3 Is a Histone-binding Protein with Two Independent Transcriptional Corepressor Activities. J. Biol. Chem. 2002, 277, 45004–45012. [Google Scholar] [CrossRef] [Green Version]
- David, S.; Kalb, R.G. Serum/glucocorticoid-inducible kinase can phosphorylate the cyclic AMP response element binding protein, CREB. FEBS Lett. 2005, 579, 1534–1538. [Google Scholar] [CrossRef] [Green Version]
- Gundersen, B.B.; Briand, L.A.; Onksen, J.L.; Lelay, J.; Kaestner, K.H.; Blendy, J.A. Increased hippocampal neurogenesis and accelerated response to antidepressants in mice with specific deletion of CREB in the hippocampus: Role of cAMP response-element modulator tau. J. Neurosci. 2013, 33, 13673–13685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hummler, E.; Cole, T.J.; Blendy, J.A.; Ganss, R.; Aguzzi, A.; Schmid, W.; Beermann, F.; Schutz, G. Targeted mutation of the CREB gene: Compensation within the CREB/ATF family of transcription factors. Proc. Natl. Acad. Sci. USA 1994, 91, 5647–5651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.W.; Klemm, D.J.; Vinson, C.; Lane, M.D. Role of CREB in transcriptional regulation of CCAAT/enhancer-binding protein beta gene during adipogenesis. J. Biol. Chem. 2004, 279, 4471–4478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bohn, T.; Rapp, S.; Luther, N.; Klein, M.; Bruehl, T.J.; Kojima, N.; Aranda Lopez, P.; Hahlbrock, J.; Muth, S.; Endo, S.; et al. Tumor immunoevasion via acidosis-dependent induction of regulatory tumor-associated macrophages. Nat. Immunol. 2018, 19, 1319–1329. [Google Scholar] [CrossRef]
- Kibbey, M.C.; Jucker, M.; Weeks, B.S.; Neve, R.L.; Van Nostrand, W.E.; Kleinman, H.K. beta-Amyloid precursor protein binds to the neurite-promoting IKVAV site of laminin. Proc. Natl. Acad. Sci. USA 1993, 90, 10150–10153. [Google Scholar] [CrossRef] [Green Version]
- Adair-Kirk, T.L.; Atkinson, J.J.; Kelley, D.G.; Arch, R.H.; Miner, J.H.; Senior, R.M. A chemotactic peptide from laminin alpha 5 functions as a regulator of inflammatory immune responses via TNF alpha-mediated signaling. J. Immunol. 2005, 174, 1621–1629. [Google Scholar] [CrossRef] [Green Version]
- Takashima, A.; Noguchi, K.; Sato, K.; Hoshino, T.; Imahori, K. Tau protein kinase I is essential for amyloid beta-protein-induced neurotoxicity. Proc. Natl. Acad. Sci. USA 1993, 90, 7789–7793. [Google Scholar] [CrossRef] [Green Version]
- Takashima, A.; Noguchi, K.; Michel, G.; Mercken, M.; Hoshi, M.; Ishiguro, K.; Imahori, K. Exposure of rat hippocampal neurons to amyloid beta peptide (25–35) induces the inactivation of phosphatidyl inositol-3 kinase and the activation of tau protein kinase I/glycogen synthase kinase-3 beta. Neurosci. Lett. 1996, 203, 33–36. [Google Scholar] [CrossRef]
- Reifert, J.; Hartung-Cranston, D.; Feinstein, S.C. Amyloid beta-mediated cell death of cultured hippocampal neurons reveals extensive Tau fragmentation without increased full-length tau phosphorylation. J. Biol. Chem. 2011, 286, 20797–20811. [Google Scholar] [CrossRef] [Green Version]
- Tyson, D.R.; Swarthout, J.T.; Jefcoat, S.C.; Partridge, N.C. PTH induction of transcriptional activity of the cAMP response element-binding protein requires the serine 129 site and glycogen synthase kinase-3 activity, but not casein kinase II sites. Endocrinology 2002, 143, 674–682. [Google Scholar] [CrossRef]
- Verma, N.K.; Fazil, M.H.; Ong, S.T.; Chalasani, M.L.; Low, J.H.; Kottaiswamy, A.; P, P.; Kizhakeyil, A.; Kumar, S.; Panda, A.K.; et al. LFA-1/ICAM-1 Ligation in Human T Cells Promotes Th1 Polarization through a GSK3beta Signaling-Dependent Notch Pathway. J. Immunol. 2016, 197, 108–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pigino, G.; Morfini, G.; Pelsman, A.; Mattson, M.P.; Brady, S.T.; Busciglio, J. Alzheimer’s presenilin 1 mutations impair kinesin-based axonal transport. J. Neurosci. 2003, 23, 4499–4508. [Google Scholar] [CrossRef] [Green Version]
- Song, B.; Lai, B.; Zheng, Z.; Zhang, Y.; Luo, J.; Wang, C.; Chen, Y.; Woodgett, J.R.; Li, M. Inhibitory phosphorylation of GSK-3 by CaMKII couples depolarization to neuronal survival. J. Biol. Chem. 2010, 285, 41122–41134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okuno, H.; Akashi, K.; Ishii, Y.; Yagishita-Kyo, N.; Suzuki, K.; Nonaka, M.; Kawashima, T.; Fujii, H.; Takemoto-Kimura, S.; Abe, M.; et al. Inverse synaptic tagging of inactive synapses via dynamic interaction of Arc/Arg3.1 with CaMKIIbeta. Cell 2012, 149, 886–898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, X.; Sato, S.; Murayama, O.; Murayama, M.; Park, J.M.; Yamaguchi, H.; Takashima, A. Lithium inhibits amyloid secretion in COS7 cells transfected with amyloid precursor protein C100. Neurosci. Lett. 2002, 321, 61–64. [Google Scholar] [CrossRef]
- Pierrot, N.; Santos, S.F.; Feyt, C.; Morel, M.; Brion, J.P.; Octave, J.N. Calcium-mediated transient phosphorylation of tau and amyloid precursor protein followed by intraneuronal amyloid-beta accumulation. J. Biol. Chem. 2006, 281, 39907–39914. [Google Scholar] [CrossRef] [Green Version]
- Pinnix, I.; Ghiso, J.A.; Pappolla, M.A.; Sambamurti, K. Major carboxyl terminal fragments generated by gamma-secretase processing of the Alzheimer amyloid precursor are 50 and 51 amino acids long. Am. J. Geriatr. Psychiatry 2013, 21, 474–483. [Google Scholar] [CrossRef] [Green Version]
- Von Rotz, R.C.; Kohli, B.M.; Bosset, J.; Meier, M.; Suzuki, T.; Nitsch, R.M.; Konietzko, U. The APP intracellular domain forms nuclear multiprotein complexes and regulates the transcription of its own precursor. J. Cell Sci. 2004, 117, 4435–4448. [Google Scholar] [CrossRef] [Green Version]
- Muller, T.; Schrotter, A.; Loosse, C.; Pfeiffer, K.; Theiss, C.; Kauth, M.; Meyer, H.E.; Marcus, K. A ternary complex consisting of AICD, FE65, and TIP60 down-regulates Stathmin1. Biochim. Biophys. Acta 2013, 1834, 387–394. [Google Scholar] [CrossRef]
- Kim, H.S.; Kim, E.M.; Lee, J.P.; Park, C.H.; Kim, S.; Seo, J.H.; Chang, K.A.; Yu, E.; Jeong, S.J.; Chong, Y.H.; et al. C-terminal fragments of amyloid precursor protein exert neurotoxicity by inducing glycogen synthase kinase-3beta expression. FASEB J. 2003, 17, 1951–1953. [Google Scholar] [CrossRef]
- Hass, M.R.; Yankner, B.A. A 1-secretase-independent mechanism of signal transduction by the amyloid precursor protein. J. Biol. Chem. 2005, 280, 36895–36904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kinoshita, A.; Whelan, C.M.; Berezovska, O.; Hyman, B.T. The gamma secretase-generated carboxyl-terminal domain of the amyloid precursor protein induces apoptosis via Tip60 in H4 cells. J. Biol. Chem. 2002, 277, 28530–28536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sumioka, A.; Nagaishi, S.; Yoshida, T.; Lin, A.; Miura, M.; Suzuki, T. Role of 14–3-3gamma in FE65-dependent gene transactivation mediated by the amyloid beta-protein precursor cytoplasmic fragment. J. Biol. Chem. 2005, 280, 42364–42374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.Y.; Kim, M.Y.; Mo, J.S.; Park, H.S. Notch1 intracellular domain suppresses APP intracellular domain-Tip60-Fe65 complex mediated signaling through physical interaction. Biochim. Biophys. Acta 2007, 1773, 736–746. [Google Scholar] [CrossRef] [Green Version]
- Fischer, D.F.; van Dijk, R.; Sluijs, J.A.; Nair, S.M.; Racchi, M.; Levelt, C.N.; van Leeuwen, F.W.; Hol, E.M. Activation of the Notch pathway in Down syndrome: Cross-talk of Notch and APP. FASEB J. 2005, 19, 1451–1458. [Google Scholar]
- Berezovska, O.; Jack, C.; Deng, A.; Gastineau, N.; Rebeck, G.W.; Hyman, B.T. Notch1 and amyloid precursor protein are competitive substrates for presenilin1-dependent gamma-secretase cleavage. J. Biol. Chem. 2001, 276, 30018–30023. [Google Scholar] [CrossRef] [Green Version]
- Cao, C.; Rioult-Pedotti, M.S.; Migani, P.; Yu, C.J.; Tiwari, R.; Parang, K.; Spaller, M.R.; Goebel, D.J.; Marshall, J. Impairment of TrkB-PSD-95 signaling in Angelman syndrome. PLoS Biol. 2013, 11, e1001478. [Google Scholar] [CrossRef]
- Nair, R.R.; Patil, S.; Tiron, A.; Kanhema, T.; Panja, D.; Schiro, L.; Parobczak, K.; Wilczynski, G.; Bramham, C.R. Dynamic Arc SUMOylation and Selective Interaction with F-Actin-Binding Protein Drebrin A in LTP Consolidation In Vivo. Front. Synaptic Neurosci. 2017, 9, 8. [Google Scholar] [CrossRef] [Green Version]
- Pastuzyn, E.D.; Shepherd, J.D. Activity-Dependent Arc Expression and Homeostatic Synaptic Plasticity Are Altered in Neurons from a Mouse Model of Angelman Syndrome. Front. Mol. Neurosci. 2017, 10, 234. [Google Scholar] [CrossRef] [Green Version]
- Yoshii, A.; Constantine-Paton, M. Postsynaptic BDNF-TrkB signaling in synapse maturation, plasticity, and disease. Dev. Neurobiol. 2010, 70, 304–322. [Google Scholar] [CrossRef] [Green Version]
- Martin-Avila, A.; Medina-Tamayo, J.; Ibarra-Sanchez, A.; Vazquez-Victorio, G.; Castillo-Arellano, J.I.; Hernandez-Mondragon, A.C.; Rivera, J.; Madera-Salcedo, I.K.; Blank, U.; Macias-Silva, M.; et al. Protein Tyrosine Kinase Fyn Regulates TLR4-Elicited Responses on Mast Cells Controlling the Function of a PP2A-PKCalpha/beta Signaling Node Leading to TNF Secretion. J. Immunol. 2016, 196, 5075–5088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Irie, Y.; Yamagata, K.; Gan, Y.; Miyamoto, K.; Do, E.; Kuo, C.H.; Taira, E.; Miki, N. Molecular cloning and characterization of Amida, a novel protein which interacts with a neuron-specific immediate early gene product arc, contains novel nuclear localization signals, and causes cell death in cultured cells. J. Biol. Chem. 2000, 275, 2647–2653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, Y.; Jin, J.; Yao, T.; Gottschalk, A.J.; Swanson, S.K.; Wu, S.; Shi, Y.; Washburn, M.P.; Florens, L.; Conaway, R.C.; et al. YY1 functions with INO80 to activate transcription. Nat. Struct. Mol. Biol. 2007, 14, 872–874. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Ooi, S.K.; Conaway, R.C.; Conaway, J.W. Generation and purification of human INO80 chromatin remodeling complexes and subcomplexes. J. Vis. Exp. 2014, 92, e51720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alberini, C.M.; Kandel, E.R. The regulation of transcription in memory consolidation. Cold Spring Harb Perspect Biol. 2014, 7, a021741. [Google Scholar] [CrossRef] [Green Version]
- Rial Verde, E.M.; Lee-Osbourne, J.; Worley, P.F.; Malinow, R.; Cline, H.T. Increased expression of the immediate-early gene arc/arg3.1 reduces AMPA receptor-mediated synaptic transmission. Neuron 2006, 52, 461–474. [Google Scholar] [CrossRef] [Green Version]
- Waung, M.W.; Pfeiffer, B.E.; Nosyreva, E.D.; Ronesi, J.A.; Huber, K.M. Rapid translation of Arc/Arg3.1 selectively mediates mGluR-dependent LTD through persistent increases in AMPAR endocytosis rate. Neuron 2008, 59, 84–97. [Google Scholar] [CrossRef] [Green Version]
- Park, S.; Park, J.M.; Kim, S.; Kim, J.A.; Shepherd, J.D.; Smith-Hicks, C.L.; Chowdhury, S.; Kaufmann, W.; Kuhl, D.; Ryazanov, A.G.; et al. Elongation factor 2 and fragile X mental retardation protein control the dynamic translation of Arc/Arg3.1 essential for mGluR-LTD. Neuron 2008, 59, 70–83. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Ardiles, A.O.; Yang, S.; Tran, T.; Posada-Duque, R.; Valdivia, G.; Baek, M.; Chuang, Y.A.; Palacios, A.G.; Gallagher, M.; et al. Metabotropic Glutamate Receptors Induce a Form of LTP Controlled by Translation and Arc Signaling in the Hippocampus. J. Neurosci. 2016, 36, 1723–1729. [Google Scholar] [CrossRef] [Green Version]
- Wilkerson, J.R.; Albanesi, J.P.; Huber, K.M. Roles for Arc in metabotropic glutamate receptor-dependent LTD and synapse elimination: Implications in health and disease. Semin. Cell Dev. Biol. 2018, 77, 51–62. [Google Scholar] [CrossRef]
- Dunn, A.R.; Kaczorowski, C.C. Regulation of intrinsic excitability: Roles for learning and memory, aging and Alzheimer’s disease, and genetic diversity. Neurobiol. Learn. Mem. 2019, 164, 107069. [Google Scholar] [CrossRef]
- Mozzachiodi, R.; Lorenzetti, F.D.; Baxter, D.A.; Byrne, J.H. Changes in neuronal excitability serve as a mechanism of long-term memory for operant conditioning. Nat. Neurosci. 2008, 11, 1146–1148. [Google Scholar] [CrossRef] [Green Version]
- Otis, J.M.; Fitzgerald, M.K.; Yousuf, H.; Burkard, J.L.; Drake, M.; Mueller, D. Prefrontal Neuronal Excitability Maintains Cocaine-Associated Memory During Retrieval. Front. Behav. Neurosci. 2018, 12, 119. [Google Scholar] [CrossRef] [PubMed]
- Scheltens, P.; Blennow, K.; Breteler, M.M.; de Strooper, B.; Frisoni, G.B.; Salloway, S.; Van der Flier, W.M. Alzheimer’s disease. Lancet 2016. [Google Scholar] [CrossRef]
- Ballatore, C.; Lee, V.M.; Trojanowski, J.Q. Tau-mediated neurodegeneration in Alzheimer’s disease and related disorders. Nat. Rev. Neurosci. 2007, 8, 663–672. [Google Scholar] [CrossRef] [PubMed]
- Kerrigan, T.L.; Randall, A.D. A new player in the "synaptopathy" of Alzheimer’s disease—Arc/arg 3.1. Front. Neurol. 2013, 4, 9. [Google Scholar] [CrossRef] [Green Version]
- Palop, J.J.; Mucke, L. Amyloid-beta-induced neuronal dysfunction in Alzheimer’s disease: From synapses toward neural networks. Nat. Neurosci. 2010, 13, 812–818. [Google Scholar] [CrossRef]
- Prince, M.; Bryce, R.; Albanese, E.; Wimo, A.; Ribeiro, W.; Ferri, C.P. The global prevalence of dementia: A systematic review and metaanalysis. Alzheimers Dement. 2013, 9, 63–75.e62. [Google Scholar] [CrossRef]
- Collaborators, G.B.D.D. Global, regional, and national burden of Alzheimer’s disease and other dementias, 1990–2016: A systematic analysis for the Global Burden of Disease Study 2016. Lancet Neurol. 2019, 18, 88–106. [Google Scholar]
- Lacor, P.N.; Buniel, M.C.; Chang, L.; Fernandez, S.J.; Gong, Y.; Viola, K.L.; Lambert, M.P.; Velasco, P.T.; Bigio, E.H.; Finch, C.E.; et al. Synaptic targeting by Alzheimer’s-related amyloid beta oligomers. J. Neurosci. 2004, 24, 10191–10200. [Google Scholar] [CrossRef] [Green Version]
- Rudinskiy, N.; Hawkes, J.M.; Betensky, R.A.; Eguchi, M.; Yamaguchi, S.; Spires-Jones, T.L.; Hyman, B.T. Orchestrated experience-driven Arc responses are disrupted in a mouse model of Alzheimer’s disease. Nat. Neurosci. 2012, 15, 1422–1429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landgren, S.; von Otter, M.; Palmer, M.S.; Zetterstrom, C.; Nilsson, S.; Skoog, I.; Gustafson, D.R.; Minthon, L.; Wallin, A.; Andreasen, N.; et al. A novel ARC gene polymorphism is associated with reduced risk of Alzheimer’s disease. J. Neural Transm. 2012, 119, 833–842. [Google Scholar] [CrossRef] [PubMed]
- Morin, J.P.; Ceron-Solano, G.; Velazquez-Campos, G.; Pacheco-Lopez, G.; Bermudez-Rattoni, F.; Diaz-Cintra, S. Spatial Memory Impairment is Associated with Intraneural Amyloid-beta Immunoreactivity and Dysfunctional Arc Expression in the Hippocampal-CA3 Region of a Transgenic Mouse Model of Alzheimer’s Disease. J. Alzheimers Dis. 2016, 51, 69–79. [Google Scholar] [CrossRef]
- Rosi, S. Neuroinflammation and the plasticity-related immediate-early gene Arc. Brain Behav. Immun. 2011, 25 (Suppl. 1), S39–S49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Long, J.M.; Holtzman, D.M. Alzheimer Disease: An Update on Pathobiology and Treatment Strategies. Cell 2019, 179, 312–339. [Google Scholar] [CrossRef] [PubMed]
- Abeysinghe, A.; Deshapriya, R.; Udawatte, C. Alzheimer’s disease; a review of the pathophysiological basis and therapeutic interventions. Life Sci. 2020, 256, 117996. [Google Scholar] [CrossRef] [PubMed]
- Du, X.; Wang, X.; Geng, M. Alzheimer’s disease hypothesis and related therapies. Transl. Neurodegener. 2018, 7, 2. [Google Scholar] [CrossRef] [Green Version]
- Liu, P.-P.; Xie, Y.; Meng, X.-Y.; Kang, J.-S. History and progress of hypotheses and clinical trials for Alzheimer’s disease. Signal. Transduct. Targeted Ther. 2019, 4, 29. [Google Scholar] [CrossRef]
- Hara, Y.; McKeehan, N.; Fillit, H.M. Translating the biology of aging into novel therapeutics for Alzheimer disease. Neurology 2019, 92, 84–93. [Google Scholar] [CrossRef] [Green Version]
- Pei, Q.; Zetterstrom, T.S.; Sprakes, M.; Tordera, R.; Sharp, T. Antidepressant drug treatment induces Arc gene expression in the rat brain. Neuroscience 2003, 121, 975–982. [Google Scholar] [CrossRef]
- Thomsen, M.S.; Hansen, H.H.; Mikkelsen, J.D. Opposite effect of phencyclidine on activity-regulated cytoskeleton-associated protein (Arc) in juvenile and adult limbic rat brain regions. Neurochem. Int. 2010, 56, 270–275. [Google Scholar] [CrossRef] [PubMed]
- McReynolds, J.R.; Donowho, K.; Abdi, A.; McGaugh, J.L.; Roozendaal, B.; McIntyre, C.K. Memory-enhancing corticosterone treatment increases amygdala norepinephrine and Arc protein expression in hippocampal synaptic fractions. Neurobiol. Learn. Mem. 2010, 93, 312–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Judes, G.; Rifai, K.; Ngollo, M.; Daures, M.; Bignon, Y.J.; Penault-Llorca, F.; Bernard-Gallon, D. A bivalent role of TIP60 histone acetyl transferase in human cancer. Epigenomics 2015, 7, 1351–1363. [Google Scholar] [CrossRef] [PubMed]
- Venkatesh, V.; Nataraj, R.; Thangaraj, G.S.; Karthikeyan, M.; Gnanasekaran, A.; Kaginelli, S.B.; Kuppanna, G.; Kallappa, C.G.; Basalingappa, K.M. Targeting Notch signalling pathway of cancer stem cells. Stem Cell Investig. 2018, 5, 5. [Google Scholar] [CrossRef] [Green Version]




















Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leung, H.-W.; Foo, G.; VanDongen, A. Arc Regulates Transcription of Genes for Plasticity, Excitability and Alzheimer’s Disease. Biomedicines 2022, 10, 1946. https://doi.org/10.3390/biomedicines10081946
Leung H-W, Foo G, VanDongen A. Arc Regulates Transcription of Genes for Plasticity, Excitability and Alzheimer’s Disease. Biomedicines. 2022; 10(8):1946. https://doi.org/10.3390/biomedicines10081946
Chicago/Turabian StyleLeung, How-Wing, Gabriel Foo, and Antonius VanDongen. 2022. "Arc Regulates Transcription of Genes for Plasticity, Excitability and Alzheimer’s Disease" Biomedicines 10, no. 8: 1946. https://doi.org/10.3390/biomedicines10081946