
Modulating Morphological and Redox/Glycative Alterations in the PCOS Uterus: Effects of Carnitines in PCOS Mice
 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
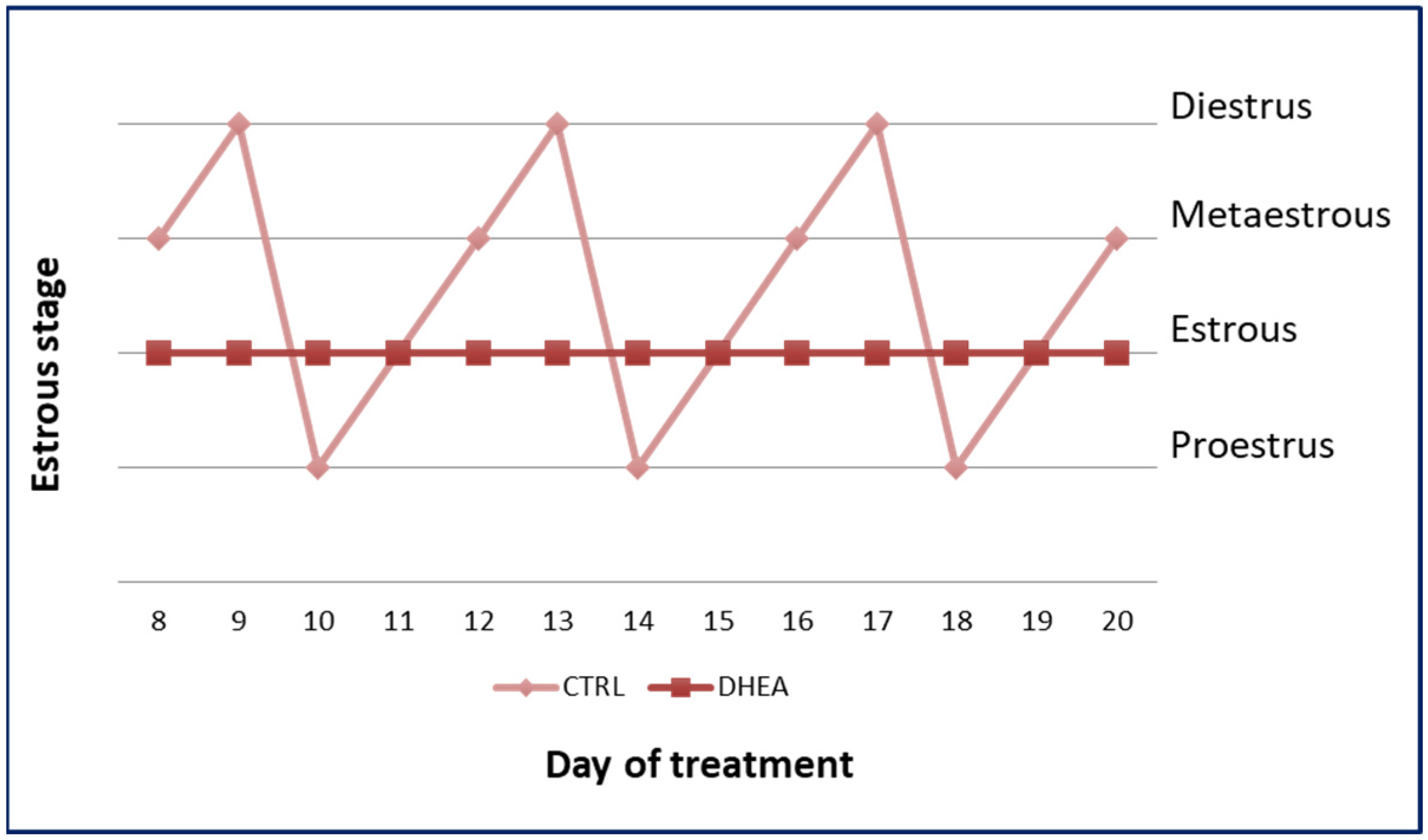
2.2. Estrous Cycle Determination
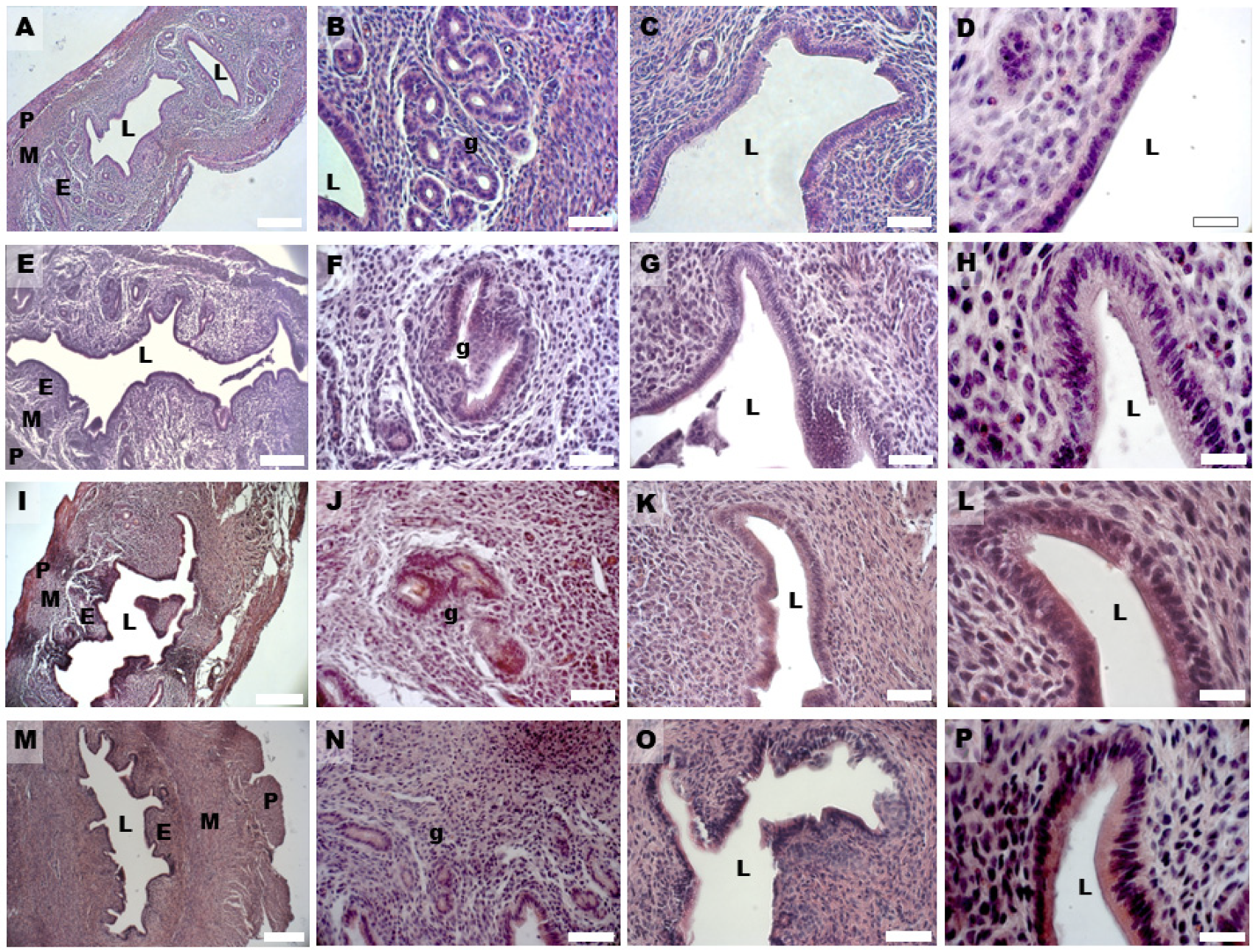
2.3. H & E Staining and Morphometric Analysis
2.4. Mallory Trichrome Staining
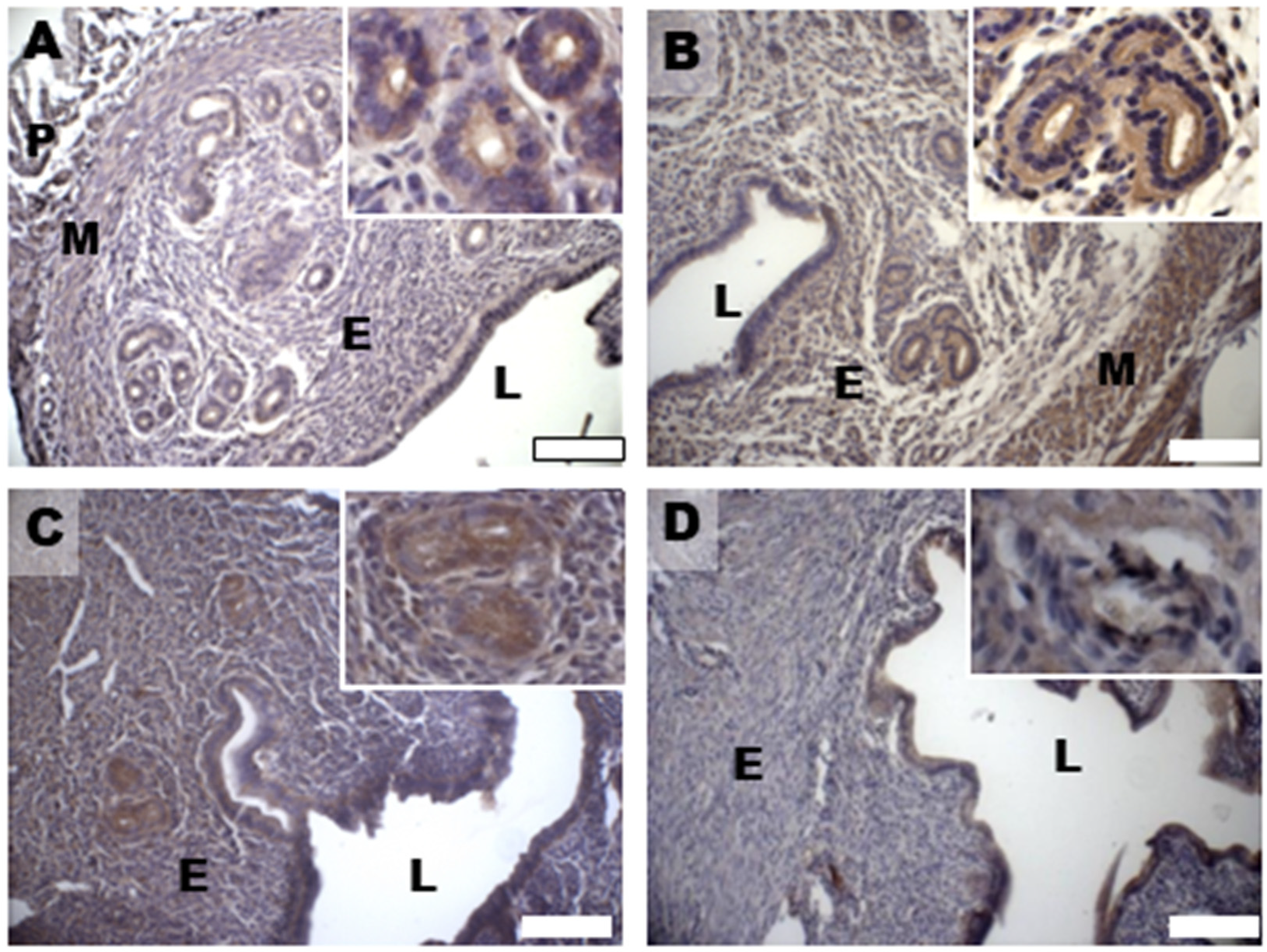
2.5. Immunohistochemical Analysis: MG-AGE and 4-HNE
2.6. Immunohistochemical Analysis
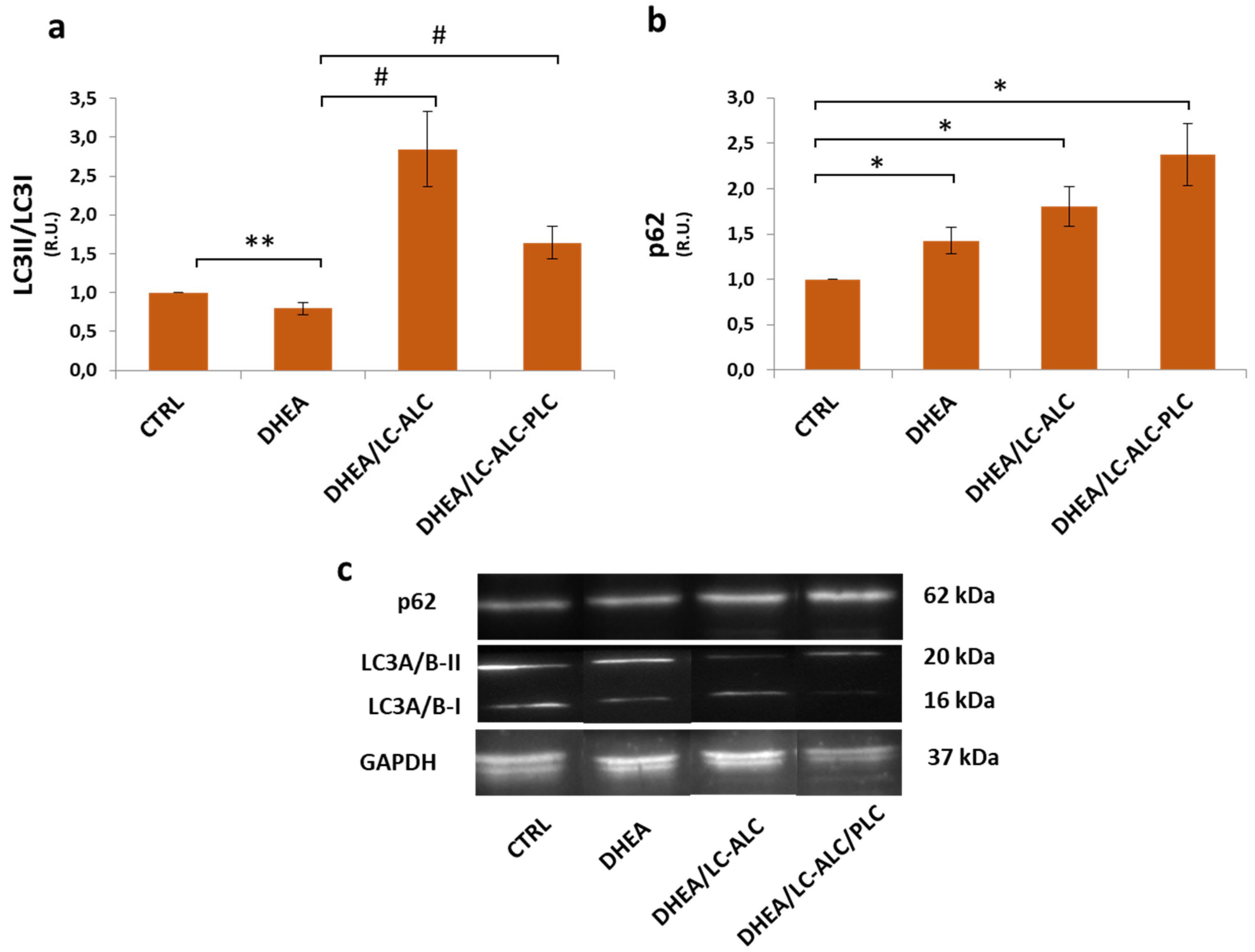
2.7. Western Blot Analysis
2.8. Statistical Analysis
3. Results
3.1. PCOS Phenotype
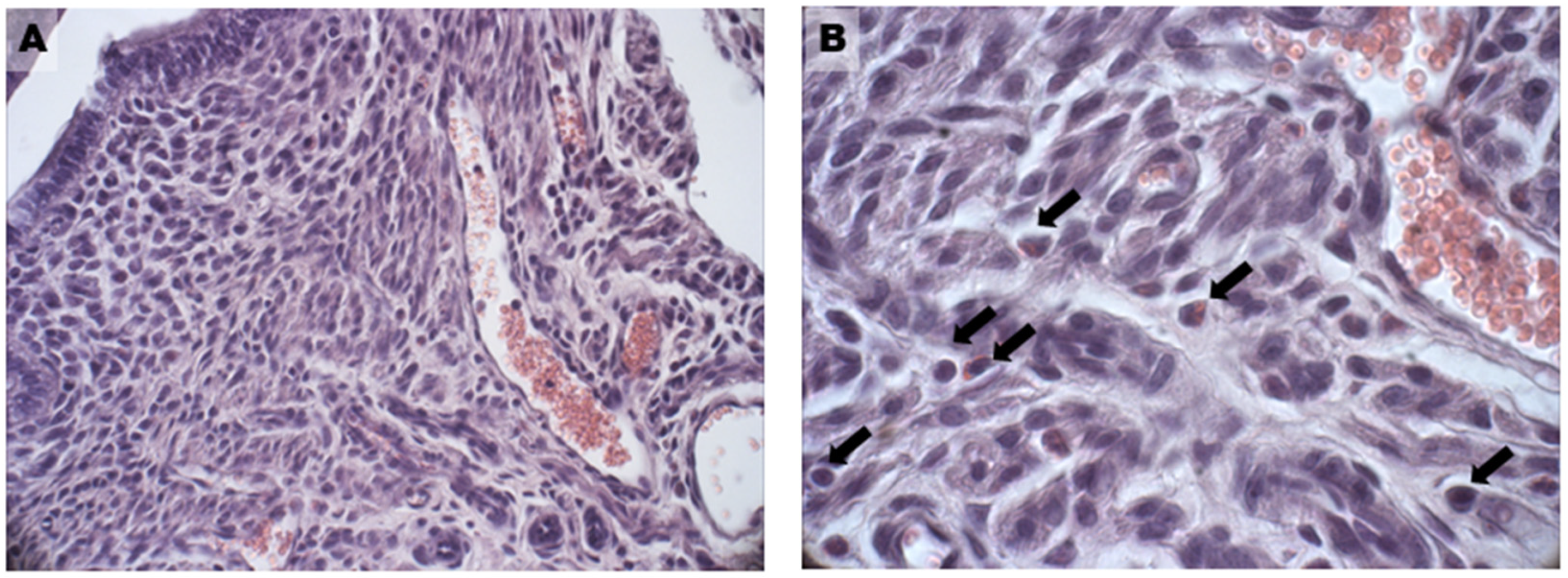
3.2. Uterine Hyperplasia and Fibrosis
3.3. Autophagic Flux
3.4. Steroidogenesis
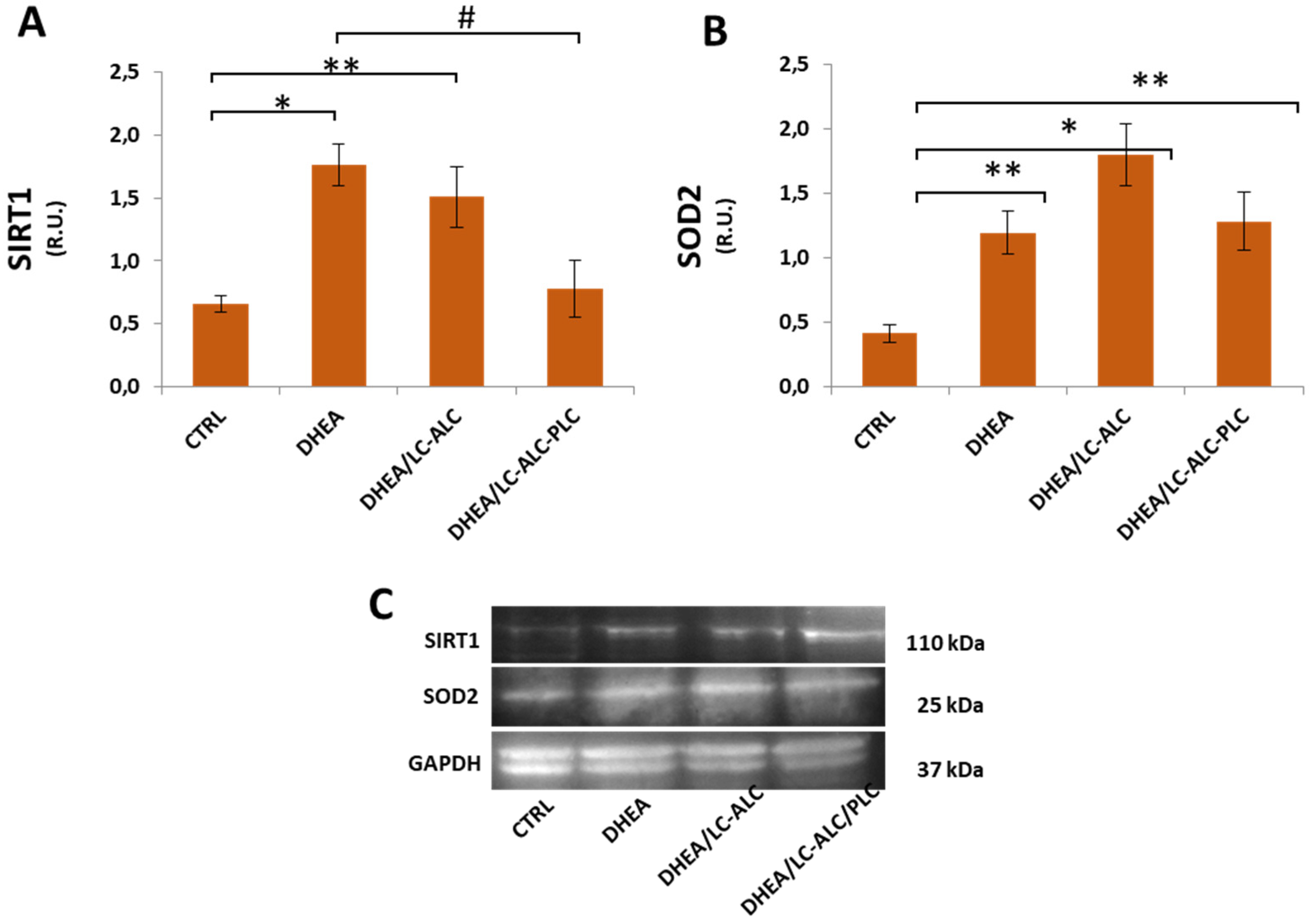
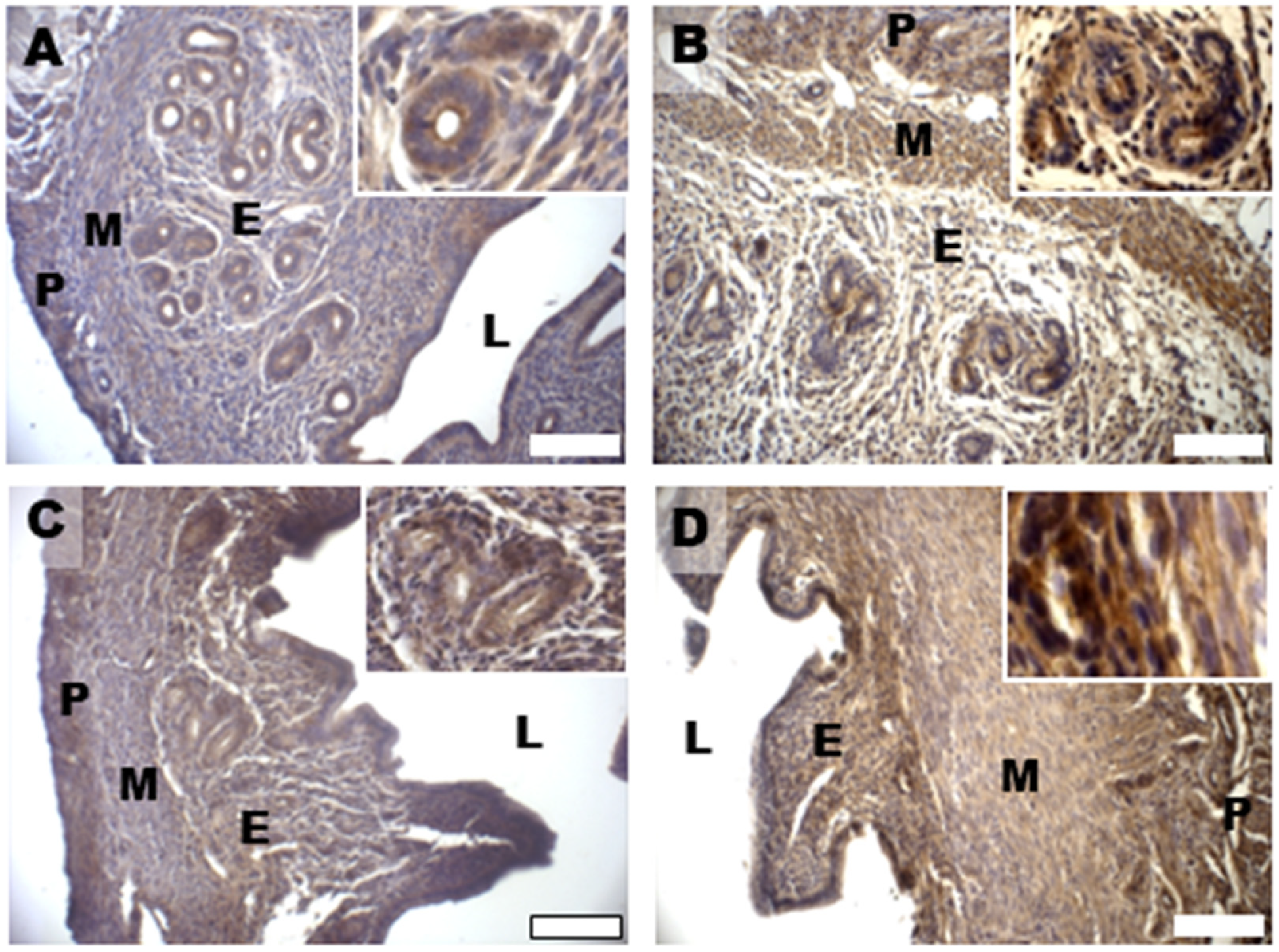
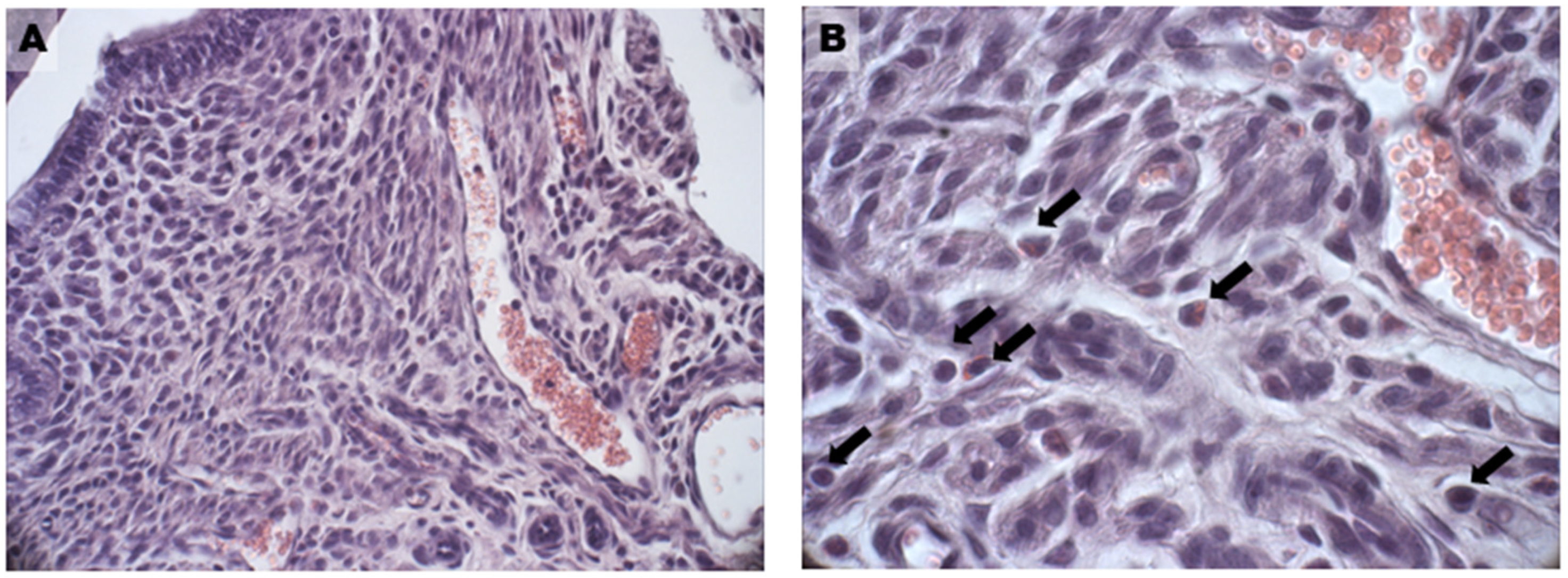
3.5. Oxidative Damage and Signaling
3.6. Mitochondrial Damage and Signaling
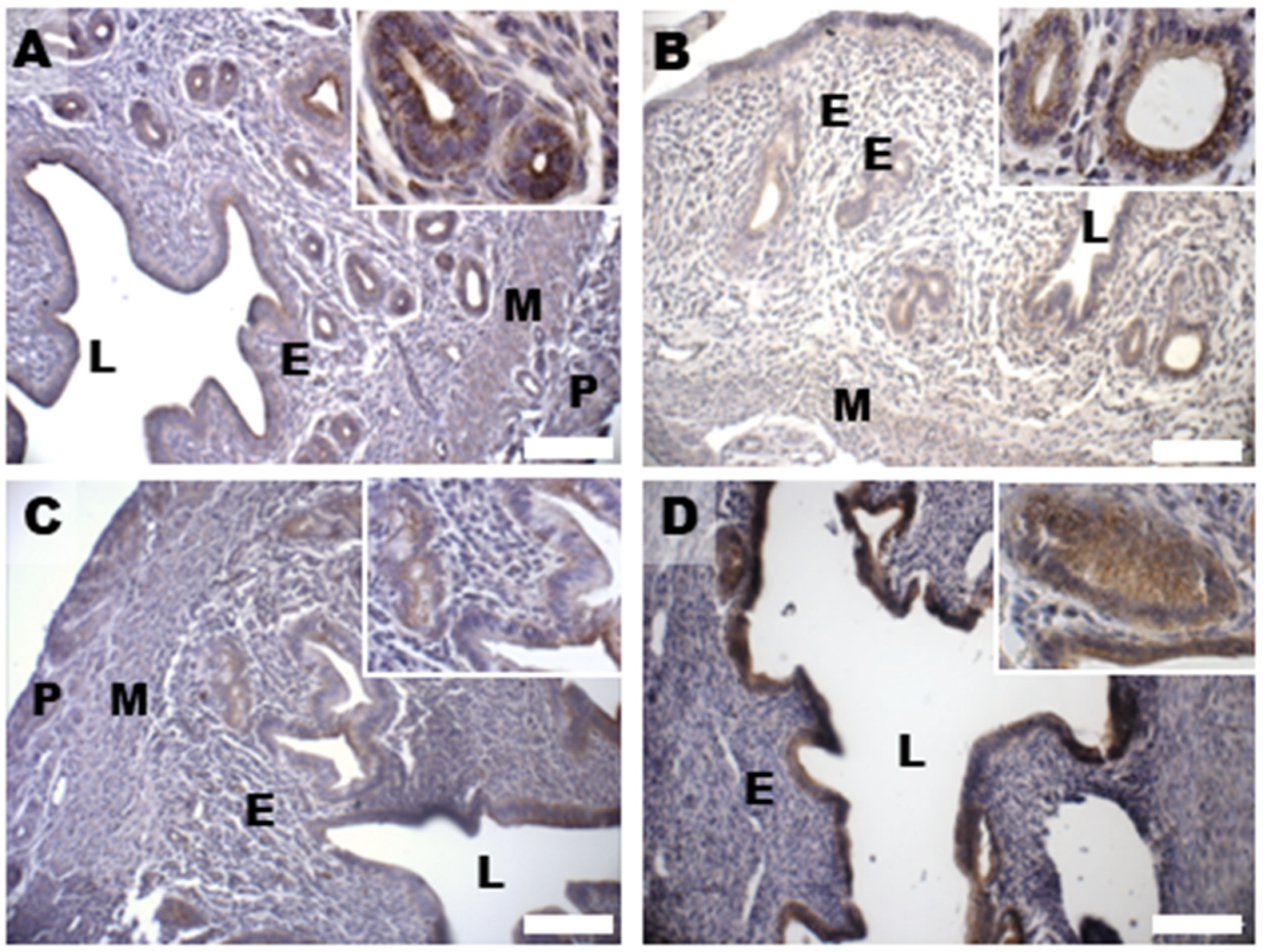
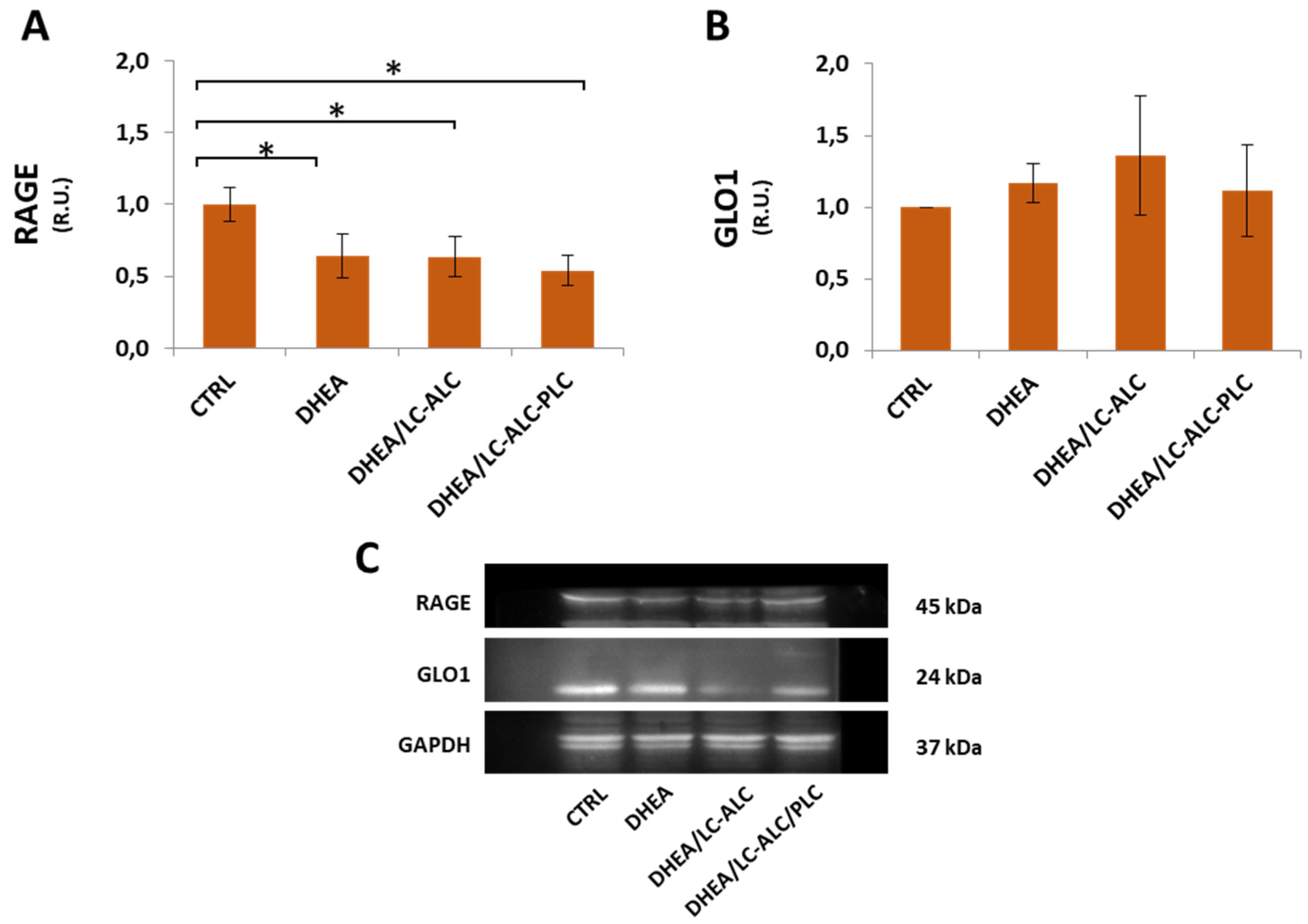
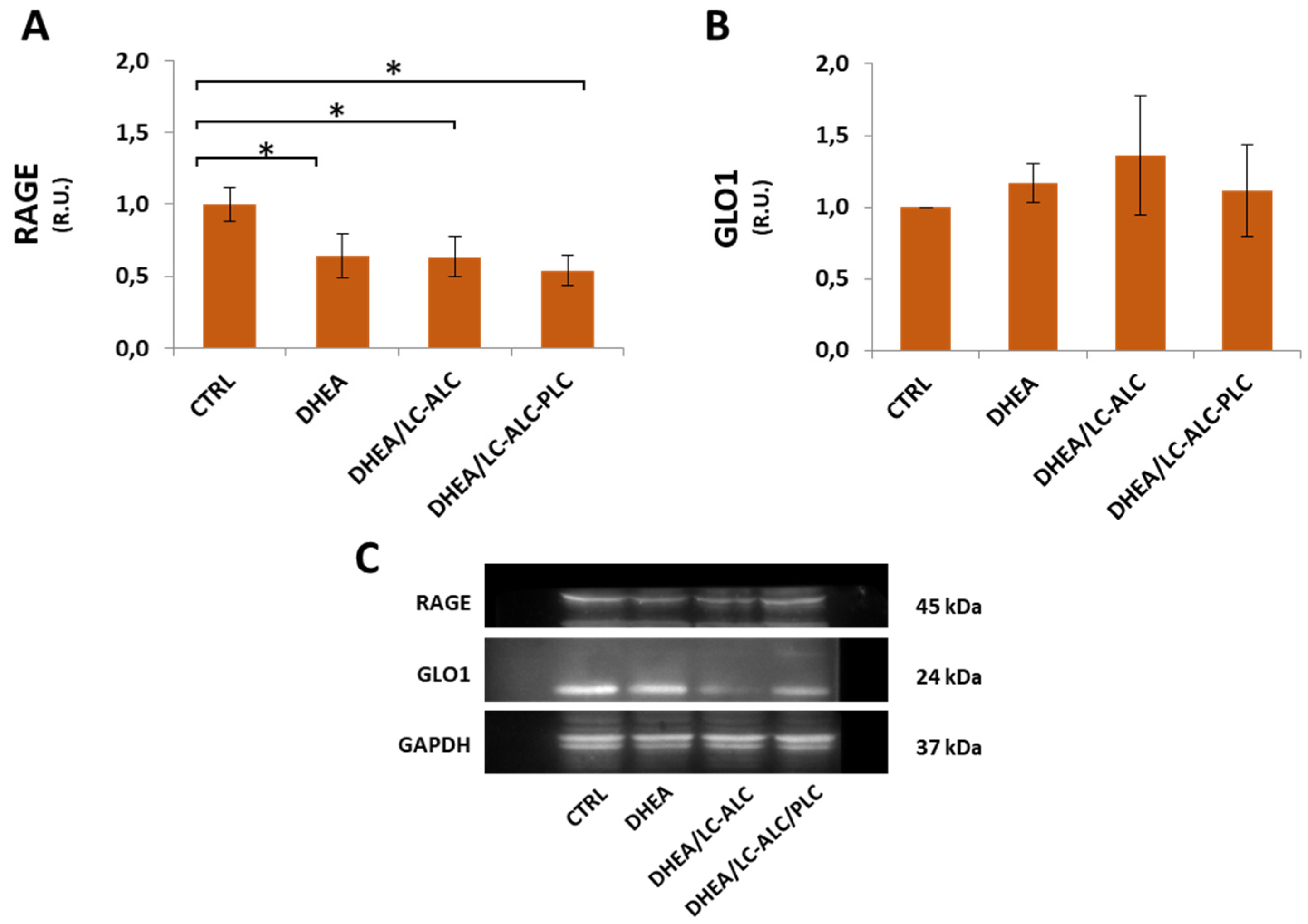
3.7. Glycative Damage and Signalling
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Skiba, M.A.; Islam, R.M.; Bell, R.J.; Davis, S.R. Understanding Variation in Prevalence Estimates of Polycystic Ovary Syndrome: A Systematic Review and Meta-Analysis. Hum. Reprod. Update 2018, 24, 694–709. [Google Scholar] [CrossRef] [PubMed]
- The Rotterdam ESHRE/ASRM-Sponsored PCOS Consensus Workshop Group. Revised 2003 Consensus on Diagnostic Criteria and Long-term Health Risks Related to Polycystic Ovary Syndrome (PCOS). Hum. Reprod. 2004, 19, 41–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azziz, R.; Carmina, E.; Dewailly, D.; Diamanti-Kandarakis, E.; Escobar-Morreale, H.F.; Futterweit, W.; Janssen, O.E.; Legro, R.S.; Norman, R.J.; Taylor, A.E.; et al. The Androgen Excess and PCOS Society Criteria for the Polycystic Ovary Syndrome: The Complete Task Force Report. Fertil. Steril. 2009, 91, 456–488. [Google Scholar] [CrossRef] [PubMed]
- Sirmans, S.M.; Pate, K.A. Epidemiology, Diagnosis, and Management of Polycystic Ovary Syndrome. Clin. Epidemiol. 2013, 6, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohammadi, M. Oxidative Stress and Polycystic Ovary Syndrome: A Brief Review. Int. J. Prev. Med. 2019, 10, 86. [Google Scholar] [CrossRef]
- Tatone, C.; Di Emidio, G.; Placidi, M.; Rossi, G.; Ruggieri, S.; Taccaliti, C.; D’Alfonso, A.; Amicarelli, F.; Guido, M. AGEs-Related Dysfunctions in PCOS: Evidence from Animal and Clinical Research. J. Endocrinol. 2021, 251, R1–R9. [Google Scholar] [CrossRef]
- Rudnicka, E.; Suchta, K.; Grymowicz, M.; Calik-Ksepka, A.; Smolarczyk, K.; Duszewska, A.M.; Smolarczyk, R.; Meczekalski, B. Chronic Low Grade Inflammation in Pathogenesis of PCOS. Int. J. Mol. Sci. 2021, 22, 3789. [Google Scholar] [CrossRef]
- Murri, M.; Insenser, M.; Escobar-Morreale, H.F. Metabolomics in Polycystic Ovary Syndrome. Clin. Chim. Acta 2014, 429, 181–188. [Google Scholar] [CrossRef]
- Bañuls, C.; Rovira-Llopis, S.; Martinez de Marañon, A.; Veses, S.; Jover, A.; Gomez, M.; Rocha, M.; Hernandez-Mijares, A.; Victor, V.M. Metabolic Syndrome Enhances Endoplasmic Reticulum, Oxidative Stress and Leukocyte-Endothelium Interactions in PCOS. Metabolism 2017, 71, 153–162. [Google Scholar] [CrossRef]
- Lai, Q.; Xiang, W.; Li, Q.; Zhang, H.; Li, Y.; Zhu, G.; Xiong, C.; Jin, L. Oxidative Stress in Granulosa Cells Contributes to Poor Oocyte Quality and IVF-ET Outcomes in Women with Polycystic Ovary Syndrome. Front. Med. 2018, 12, 518–524. [Google Scholar] [CrossRef]
- Advani, K.; Batra, M.; Tajpuriya, S.; Gupta, R.; Saraswat, A.; Nagar, H.D.; Makwana, L.; Kshirsagar, S.; Kaul, P.; Ghosh, A.K.; et al. Efficacy of Combination Therapy of Inositols, Antioxidants and Vitamins in Obese and Non-Obese Women with Polycystic Ovary Syndrome: An Observational Study. J. Obstet. Gynaecol. 2020, 40, 96–101. [Google Scholar] [CrossRef]
- Furat Rencber, S.; Kurnaz Ozbek, S.; Eraldemır, C.; Sezer, Z.; Kum, T.; Ceylan, S.; Guzel, E. Effect of Resveratrol and Metformin on Ovarian Reserve and Ultrastructure in PCOS: An Experimental Study. J. Ovarian Res. 2018, 11, 55. [Google Scholar] [CrossRef] [Green Version]
- Heshmati, J.; Golab, F.; Morvaridzadeh, M.; Potter, E.; Akbari-Fakhrabadi, M.; Farsi, F.; Tanbakooei, S.; Shidfar, F. The Effects of Curcumin Supplementation on Oxidative Stress, Sirtuin-1 and Peroxisome Proliferator Activated Receptor γ Coactivator 1α Gene Expression in Polycystic Ovarian Syndrome (PCOS) Patients: A Randomized Placebo-Controlled Clinical Trial. Diabetes Metab. Syndr. 2020, 14, 77–82. [Google Scholar] [CrossRef]
- Yarmolinskaya, M.; Bulgakova, O.; Abashova, E.; Borodina, V.; Tral, T. The Effectiveness of Resveratrol in Treatment of PCOS on the Basis of Experimental Model in Rats. Gynecol. Endocrinol. 2021, 37, 54–57. [Google Scholar] [CrossRef] [PubMed]
- Hosseinzadeh, P.; Barsky, M.; Gibbons, W.E.; Blesson, C.S. Polycystic Ovary Syndrome and the Forgotten Uterus. F&S Rev. 2021, 2, 11–20. [Google Scholar] [CrossRef]
- Palomba, S.; Piltonen, T.T.; Giudice, L.C. Endometrial Function in Women with Polycystic Ovary Syndrome: A Comprehensive Review. Hum. Reprod. Update 2021, 27, 584–618. [Google Scholar] [CrossRef]
- Tal, R.; Seifer, D.B.; Arici, A. The Emerging Role of Angiogenic Factor Dysregulation in the Pathogenesis of Polycystic Ovarian Syndrome. Semin. Reprod. Med. 2015, 33, 195–207. [Google Scholar] [CrossRef]
- Bai, X.; Zheng, L.; Li, D.; Xu, Y. Research Progress of Endometrial Receptivity in Patients with Polycystic Ovary Syndrome: A Systematic Review. Reprod. Biol. Endocrinol. 2021, 19, 122. [Google Scholar] [CrossRef]
- Leonhardt, H.; Gull, B.; Kishimoto, K.; Kataoka, M.; Nilsson, L.; Janson, P.O.; Stener-Victorin, E.; Hellström, M. Uterine Morphology and Peristalsis in Women with Polycystic Ovary Syndrome. Acta Radiol. 2012, 53, 1195–1201. [Google Scholar] [CrossRef] [Green Version]
- Kido, A.; Togashi, K. Uterine Anatomy and Function on Cine Magnetic Resonance Imaging. Reprod. Med. Biol. 2016, 15, 191–199. [Google Scholar] [CrossRef]
- Bracho, G.S.; Altamirano, G.A.; Kass, L.; Luque, E.H.; Bosquiazzo, V.L. Hyperandrogenism Induces Histo-Architectural Changes in the Rat Uterus. Reprod. Sci. 2019, 26, 657–668. [Google Scholar] [CrossRef] [PubMed]
- Hansda, S.R.; Haldar, C. Uterine Anomalies in Cell Proliferation, Energy Homeostasis and Oxidative Stress in PCOS Hamsters, M. Auratus: Therapeutic Potentials of Melatonin. Life Sci. 2021, 281, 119755. [Google Scholar] [CrossRef] [PubMed]
- Li, S.-Y.; Song, Z.; Song, M.-J.; Qin, J.-W.; Zhao, M.-L.; Yang, Z.-M. Impaired Receptivity and Decidualization in DHEA-Induced PCOS Mice. Sci. Rep. 2016, 6, 38134. [Google Scholar] [CrossRef] [PubMed]
- Placidi, M.; Di Emidio, G.; Virmani, A.; D’Alfonso, A.; Artini, P.G.; D’Alessandro, A.M.; Tatone, C. Carnitines as Mitochondrial Modulators of Oocyte and Embryo Bioenergetics. Antioxidants 2022, 11, 745. [Google Scholar] [CrossRef] [PubMed]
- Fenkci, S.M.; Fenkci, V.; Oztekin, O.; Rota, S.; Karagenc, N. Serum Total L-Carnitine Levels in Non-Obese Women with Polycystic Ovary Syndrome. Hum. Reprod. 2008, 23, 1602–1606. [Google Scholar] [CrossRef] [Green Version]
- Celik, F.; Kose, M.; Yilmazer, M.; Köken, G.N.; Arioz, D.T.; Pektas, M.K. Plasma L-Carnitine Levels of Obese and Non-Obese Polycystic Ovary Syndrome Patients. J. Obstet. Gynaecol. 2017, 37, 476–479. [Google Scholar] [CrossRef]
- Solarska, K.; Lewińska, A.; Karowicz-Bilińska, A.; Bartosz, G. The Antioxidant Properties of Carnitine In Vitro. Cell. Mol. Biol. Lett. 2010, 15, 90–97. [Google Scholar] [CrossRef]
- Di Emidio, G.; Rea, F.; Placidi, M.; Rossi, G.; Cocciolone, D.; Virmani, A.; Macchiarelli, G.; Palmerini, M.G.; D’Alessandro, A.M.; Artini, P.G.; et al. Regulatory Functions of L-Carnitine, Acetyl, and Propionyl L-Carnitine in a PCOS Mouse Model: Focus on Antioxidant/Antiglycative Molecular Pathways in the Ovarian Microenvironment. Antioxidants 2020, 9, 867. [Google Scholar] [CrossRef]
- Di Emidio, G.; Placidi, M.; Rea, F.; Rossi, G.; Falone, S.; Cristiano, L.; Nottola, S.; D’Alessandro, A.M.; Amicarelli, F.; Palmerini, M.G.; et al. Methylglyoxal-Dependent Glycative Stress and Deregulation of SIRT1 Functional Network in the Ovary of PCOS Mice. Cells 2020, 9, 209. [Google Scholar] [CrossRef] [Green Version]
- Caligioni, C.S. Assessing Reproductive Status/Stages in Mice. Curr. Protoc. Neurosci. 2009, 48, A.4I.1–A.4I.8. [Google Scholar] [CrossRef] [Green Version]
- Kuyucu, Y.; Çelik, L.S.; Kendirlinan, Ö.; Tap, Ö.; Mete, U.Ö. Investigation of the Uterine Structural Changes in the Experimental Model with Polycystic Ovary Syndrome and Effects of Vitamin D Treatment: An Ultrastructural and Immunohistochemical Study. Reprod. Biol. 2018, 18, 53–59. [Google Scholar] [CrossRef]
- Varghese, F.; Bukhari, A.B.; Malhotra, R.; De, A. IHC Profiler: An Open Source Plugin for the Quantitative Evaluation and Automated Scoring of Immunohistochemistry Images of Human Tissue Samples. PLoS ONE 2014, 9, e96801. [Google Scholar] [CrossRef] [Green Version]
- Marcolongo, P.; Maellaro, E.; Luisi, S. Regulation of Autophagy in the Uterus: From Physiological Processes to Endometriosis and Uterine Fibroids. F&S Rev. 2022, 3, 69–75. [Google Scholar] [CrossRef]
- Zhao, X.; Jiang, Y.; Ren, J.; Wang, Y.; Zhao, Y.; Feng, X. Deciphering the Mechanism of Bushen Huoxue Decotion on Decidualization by Intervening Autophagy via AMPK/MTOR/ULK1: A Novel Discovery for URSA Treatment. Front. Pharmacol. 2022, 13, 794938. [Google Scholar] [CrossRef]
- Breitling, R.; Marijanović, Z.; Perović, D.; Adamski, J. Evolution of 17β-HSD Type 4, a Multifunctional Protein of β-Oxidation. Mol. Cell. Endocrinol. 2001, 171, 205–210. [Google Scholar] [CrossRef]
- Castro, J.P.; Jung, T.; Grune, T.; Siems, W. 4-Hydroxynonenal (HNE) Modified Proteins in Metabolic Diseases. Free. Radic. Biol. Med. 2017, 111, 309–315. [Google Scholar] [CrossRef] [Green Version]
- Singh, C.K.; Chhabra, G.; Ndiaye, M.A.; Garcia-Peterson, L.M.; Mack, N.J.; Ahmad, N. The Role of Sirtuins in Antioxidant and Redox Signaling. Antioxid. Redox Signal. 2018, 28, 643–661. [Google Scholar] [CrossRef]
- Tang, B.L. Sirt1 and the Mitochondria. Mol. Cells 2016, 39, 87–95. [Google Scholar] [CrossRef] [Green Version]
- Campbell, C.T.; Kolesar, J.E.; Kaufman, B.A. Mitochondrial Transcription Factor A Regulates Mitochondrial Transcription Initiation, DNA Packaging, and Genome Copy Number. Biochim. Biophys. Acta (BBA)—Gene Regul. Mech. 2012, 1819, 921–929. [Google Scholar] [CrossRef]
- Mirabolghasemi, G.; Kamyab, Z. Changes of The Uterine Tissue in Rats with Polycystic Ovary Syndrome Induced by Estradiol Valerate. Int. J. Fertil. Steril. 2017, 11, 47–55. [Google Scholar] [CrossRef]
- Wang, J.; Wu, D.; Guo, H.; Li, M. Hyperandrogenemia and Insulin Resistance: The Chief Culprit of Polycystic Ovary Syndrome. Life Sci. 2019, 236, 116940. [Google Scholar] [CrossRef] [PubMed]
- Ashary, N.; Tiwari, A.; Modi, D. Embryo Implantation: War in Times of Love. Endocrinology 2018, 159, 1188–1198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, N.-X.; Li, X.-L. The Disorders of Endometrial Receptivity in PCOS and Its Mechanisms. Reprod. Sci. 2022, 29, 2465–2476. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.; Li, J.; Zhang, Y.; Li, X.; Brännström, M.; Shao, L.R.; Billig, H. Endometrial Progesterone Receptor Isoforms in Women with Polycystic Ovary Syndrome. Am. J. Transl. Res. 2018, 10, 2696–2705. [Google Scholar] [PubMed]
- Savaris, R.F.; Groll, J.M.; Young, S.L.; DeMayo, F.J.; Jeong, J.-W.; Hamilton, A.E.; Giudice, L.C.; Lessey, B.A. Progesterone Resistance in PCOS Endometrium: A Microarray Analysis in Clomiphene Citrate-Treated and Artificial Menstrual Cycles. J. Clin. Endocrinol. Metab. 2011, 96, 1737–1746. [Google Scholar] [CrossRef] [Green Version]
- Xue, Z.; Li, J.; Feng, J.; Han, H.; Zhao, J.; Zhang, J.; Han, Y.; Wu, X.; Zhang, Y. Research Progress on the Mechanism between Polycystic Ovary Syndrome and Abnormal Endometrium. Front. Physiol. 2021, 12, 788772. [Google Scholar] [CrossRef]
- Hu, J.; Zeng, B.; Jiang, X.; Hu, L.; Meng, Y.; Zhu, Y.; Mao, M. The Expression of Marker for Endometrial Stem Cell and Fibrosis Was Increased in Intrauterine Adhesious. Int. J. Clin. Exp. Pathol. 2015, 8, 1525–1534. [Google Scholar]
- Kim, T.H.; Yoo, J.-Y.; Choi, K.-C.; Shin, J.-H.; Leach, R.E.; Fazleabas, A.T.; Young, S.L.; Lessey, B.A.; Yoon, H.-G.; Jeong, J.-W. Loss of HDAC3 Results in Nonreceptive Endometrium and Female Infertility. Sci. Transl. Med. 2019, 11, eaaf7533. [Google Scholar] [CrossRef]
- Aboeldalyl, S.; James, C.; Seyam, E.; Ibrahim, E.M.; Shawki, H.E.-D.; Amer, S. The Role of Chronic Inflammation in Polycystic Ovarian Syndrome—A Systematic Review and Meta-Analysis. Int. J. Mol. Sci. 2021, 22, 2734. [Google Scholar] [CrossRef]
- Secomandi, L.; Borghesan, M.; Velarde, M.; Demaria, M. The Role of Cellular Senescence in Female Reproductive Aging and the Potential for Senotherapeutic Interventions. Hum. Reprod. Update 2022, 28, 172–189. [Google Scholar] [CrossRef]
- Motta, A.B. Dehydroepiandrosterone to Induce Murine Models for the Study of Polycystic Ovary Syndrome. J. Steroid Biochem. Mol. Biol. 2010, 119, 105–111. [Google Scholar] [CrossRef]
- Soares, D.M.; Junior, H.W.; Bittencourt, L.K.; Lopes, F.P.P.L.; de Oliveira, M.A.P. The Role of Cine MR Imaging in the Assessment of Uterine Function. Arch. Gynecol. Obstet. 2019, 300, 545–553. [Google Scholar] [CrossRef]
- Austin, S.; St-Pierre, J. PGC1α and Mitochondrial Metabolism—Emerging Concepts and Relevance in Ageing and Neurodegenerative Disorders. J. Cell Sci. 2012, 125, 4963–4971. [Google Scholar] [CrossRef] [Green Version]
- Merhi, Z. Crosstalk between Advanced Glycation End Products and Vitamin D: A Compelling Paradigm for the Treatment of Ovarian Dysfunction in PCOS. Mol. Cell. Endocrinol. 2019, 479, 20–26. [Google Scholar] [CrossRef]
- Lin, S.-Y.; Yang, Y.-C.; Chang, C.Y.-Y.; Lin, C.-C.; Hsu, W.-H.; Ju, S.-W.; Hsu, C.-Y.; Kao, C.-H. Risk of Polycystic Ovary Syndrome in Women Exposed to Fine Air Pollutants and Acidic Gases: A Nationwide Cohort Analysis. Int. J. Environ. Res. Public Health 2019, 16, 4816. [Google Scholar] [CrossRef] [Green Version]
- Rutkowska, A.Z.; Diamanti-Kandarakis, E. Do Advanced Glycation End Products (AGEs) Contribute to the Comorbidities of Polycystic Ovary Syndrome (PCOS)? Curr. Pharm. Des. 2016, 22, 5558–5571. [Google Scholar] [CrossRef]
- Papachristoforou, E.; Lambadiari, V.; Maratou, E.; Makrilakis, K. Association of Glycemic Indices (Hyperglycemia, Glucose Variability, and Hypoglycemia) with Oxidative Stress and Diabetic Complications. J. Diabetes Res. 2020, 2020, 7489795. [Google Scholar] [CrossRef]
- Sibbersen, C.; Johannsen, M. Dicarbonyl Derived Post-Translational Modifications: Chemistry Bridging Biology and Aging-Related Disease. Essays Biochem. 2020, 64, 97–110. [Google Scholar] [CrossRef]
- Tatone, C.; Eichenlaub-Ritter, U.; Amicarelli, F. Dicarbonyl Stress and Glyoxalases in Ovarian Function. Biochem. Soc. Trans. 2014, 42, 433–438. [Google Scholar] [CrossRef]
- Erusalimsky, J.D. The Use of the Soluble Receptor for Advanced Glycation-End Products (SRAGE) as a Potential Biomarker of Disease Risk and Adverse Outcomes. Redox Biol. 2021, 42, 101958. [Google Scholar] [CrossRef]
- Qin, J.; Goswami, R.; Dawson, S.; Dawson, G. Expression of the Receptor for Advanced Glycation End Products in Oligodendrocytes in Response to Oxidative Stress. J. Neurosci. Res. 2008, 86, 2414–2422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mingorance, C.; Rodriguez-Rodriguez, R.; Justo, M.L.; Herrera, M.D.; de Sotomayor, M.A. Pharmacological Effects and Clinical Applications of Propionyl-L-Carnitine. Nutr. Rev. 2011, 69, 279–290. [Google Scholar] [CrossRef]
- Yang, T.; Zhao, J.; Liu, F.; Li, Y. Lipid Metabolism and Endometrial Receptivity. Hum. Reprod. Update 2022, 28, 858–889. [Google Scholar] [CrossRef] [PubMed]
- Houten, S.M.; Wanders, R.J.A.; Ranea-Robles, P. Metabolic Interactions between Peroxisomes and Mitochondria with a Special Focus on Acylcarnitine Metabolism. Biochim. Biophys. Acta Mol. Basis Dis. 2020, 1866, 165720. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control (µm) | DHEA (µm) | DHEA/LC-ALC (µm) | DHEA/LC-ALC-PLC (µm) | |
|---|---|---|---|---|
| Endometrium | 141.615 ± 13.266 a | 345.584 ± 17.199 b | 237.225 ± 40.884 c | 171.683 ± 8.181 a,c |
| Epithelium | 12.464 ± 1.685 a | 25.119 ± 2.370 b | 19.435 ± 2.582 a,b | 13.747 ± 1.494 a |
| Stroma | 129.864 ± 13.419 a | 321.645 ± 14.989 b | 209.886 ± 38.386 a | 159.004 ± 7.373 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Palmerini, M.G.; Macchiarelli, G.; Cocciolone, D.; Mascitti, I.A.; Placidi, M.; Vergara, T.; Di Emidio, G.; Tatone, C. Modulating Morphological and Redox/Glycative Alterations in the PCOS Uterus: Effects of Carnitines in PCOS Mice. Biomedicines 2023, 11, 374. https://doi.org/10.3390/biomedicines11020374
Palmerini MG, Macchiarelli G, Cocciolone D, Mascitti IA, Placidi M, Vergara T, Di Emidio G, Tatone C. Modulating Morphological and Redox/Glycative Alterations in the PCOS Uterus: Effects of Carnitines in PCOS Mice. Biomedicines. 2023; 11(2):374. https://doi.org/10.3390/biomedicines11020374
Chicago/Turabian StylePalmerini, Maria Grazia, Guido Macchiarelli, Domenica Cocciolone, Ilaria Antenisca Mascitti, Martina Placidi, Teresa Vergara, Giovanna Di Emidio, and Carla Tatone. 2023. "Modulating Morphological and Redox/Glycative Alterations in the PCOS Uterus: Effects of Carnitines in PCOS Mice" Biomedicines 11, no. 2: 374. https://doi.org/10.3390/biomedicines11020374