Figure 1.
Evaluation of T-lymphocytes transduction efficiency by lentiviral particles. (A)—dot plot: T cell population on the forward and side scatter plot; (B)—dot plot: population of GFP+ transduced T-lymphocytes (CAR-T cells); and (C)—dot plot: population of CAR+ T cells (CAR-T cells) in GFP+ population assessed via biotinylated protein L bound to streptavidin-Pacific Blue conjugate.
Figure 1.
Evaluation of T-lymphocytes transduction efficiency by lentiviral particles. (A)—dot plot: T cell population on the forward and side scatter plot; (B)—dot plot: population of GFP+ transduced T-lymphocytes (CAR-T cells); and (C)—dot plot: population of CAR+ T cells (CAR-T cells) in GFP+ population assessed via biotinylated protein L bound to streptavidin-Pacific Blue conjugate.
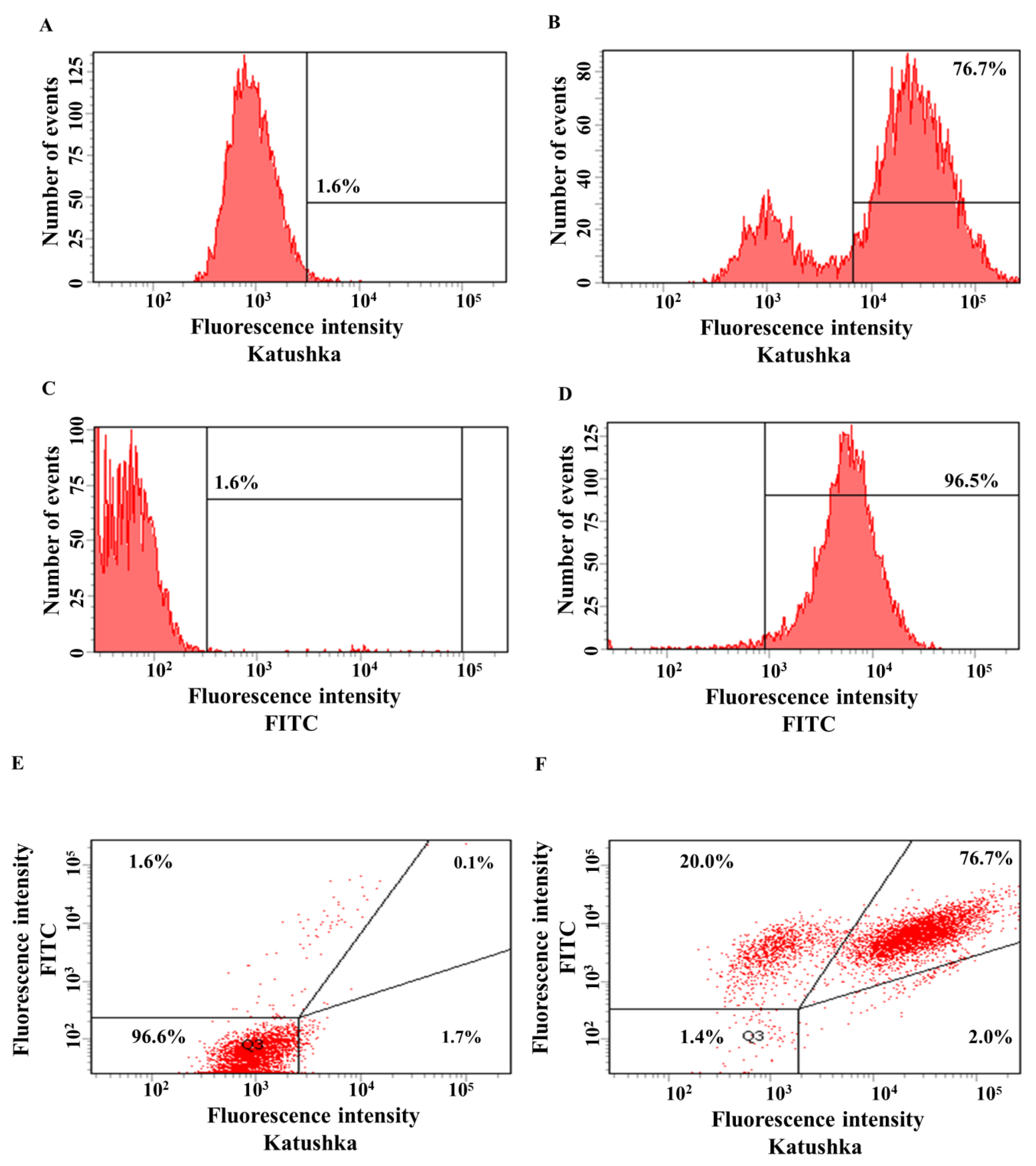
Figure 2.
Evaluation of the transduction efficiency of H522 cells with lentiviruses encoding Katushka2S and/or CD19. Flow cytometry, representative histograms (A–D), and dot plots (E,F) are presented. (A)—negative control, non-transduced cells; (B)—H522(Kat+) cells expressing red fluorescent protein Katushka2S; (C)—negative control, non-transduced cells; (D)—H522(CD19+) cells expressing the CD19 antigen; (E)—negative control, non-transduced cells are shown in the upper right corner; and (F)—H522(Kat+CD19+) cells simultaneously expressing both red fluorescent protein Katushka2S and the CD19 antigen are shown in the upper right corner.
Figure 2.
Evaluation of the transduction efficiency of H522 cells with lentiviruses encoding Katushka2S and/or CD19. Flow cytometry, representative histograms (A–D), and dot plots (E,F) are presented. (A)—negative control, non-transduced cells; (B)—H522(Kat+) cells expressing red fluorescent protein Katushka2S; (C)—negative control, non-transduced cells; (D)—H522(CD19+) cells expressing the CD19 antigen; (E)—negative control, non-transduced cells are shown in the upper right corner; and (F)—H522(Kat+CD19+) cells simultaneously expressing both red fluorescent protein Katushka2S and the CD19 antigen are shown in the upper right corner.
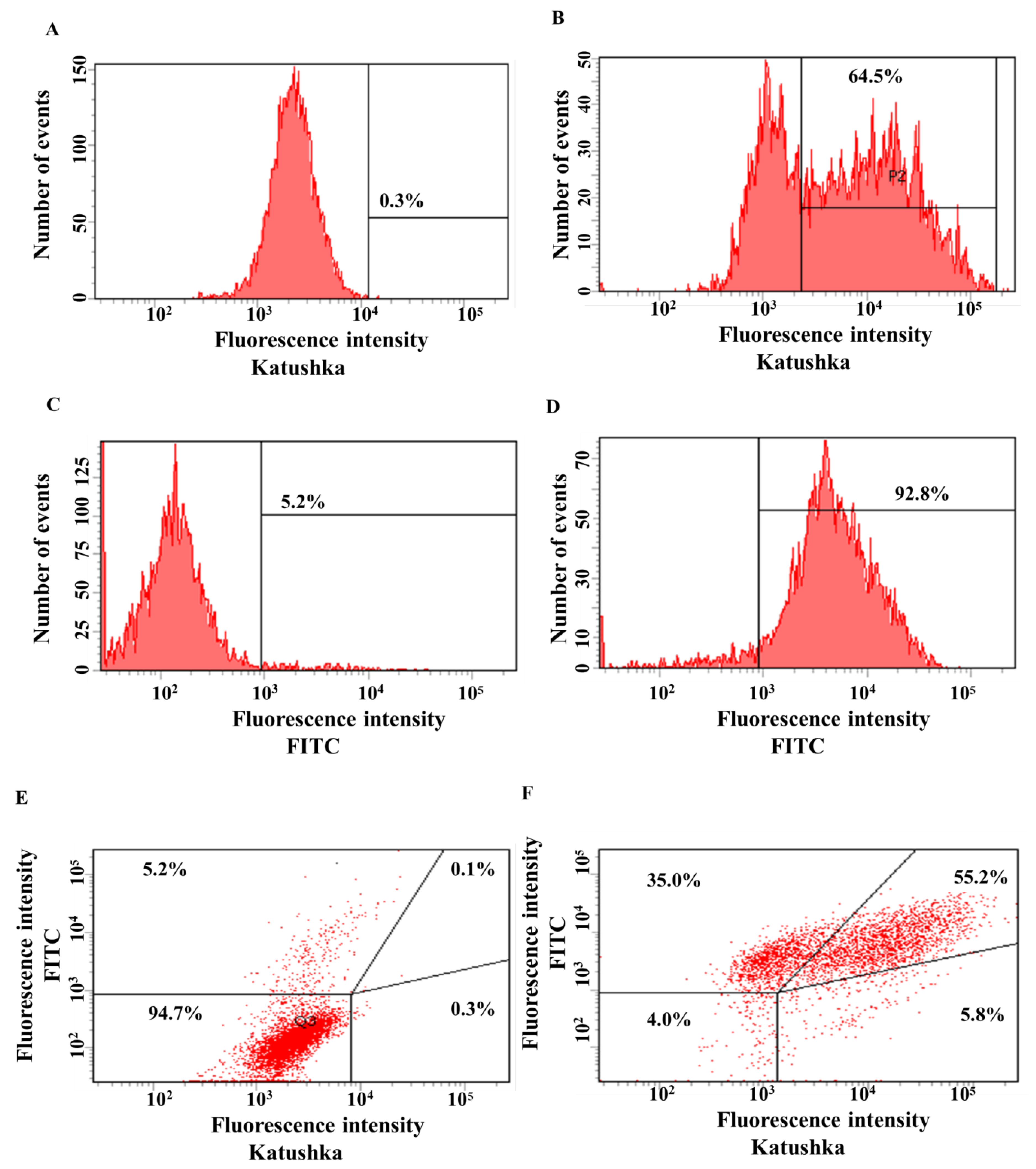
Figure 3.
Evaluation of the transduction efficiency of PC-3M cells by lentiviruses encoding Katushka2S and/or CD19. Flow cytometry, representative histograms (A–D), and scatter plots (E,F) are presented. (A)—negative control, non-transduced cells; (B)—PC-3M(Kat+) cells expressing red fluorescent protein Katushka2S; (C)—negative control, non-transduced cells; (D)—PC-3M(CD19+) cells expressing CD19 antigen; (E)—negative control, non-transduced cells are shown in the upper right corner; and (F)—PC-3M(Kat+CD19+) cells expressing both red fluorescent protein Katushka2S and CD19 antigen are shown in the upper right corner.
Figure 3.
Evaluation of the transduction efficiency of PC-3M cells by lentiviruses encoding Katushka2S and/or CD19. Flow cytometry, representative histograms (A–D), and scatter plots (E,F) are presented. (A)—negative control, non-transduced cells; (B)—PC-3M(Kat+) cells expressing red fluorescent protein Katushka2S; (C)—negative control, non-transduced cells; (D)—PC-3M(CD19+) cells expressing CD19 antigen; (E)—negative control, non-transduced cells are shown in the upper right corner; and (F)—PC-3M(Kat+CD19+) cells expressing both red fluorescent protein Katushka2S and CD19 antigen are shown in the upper right corner.
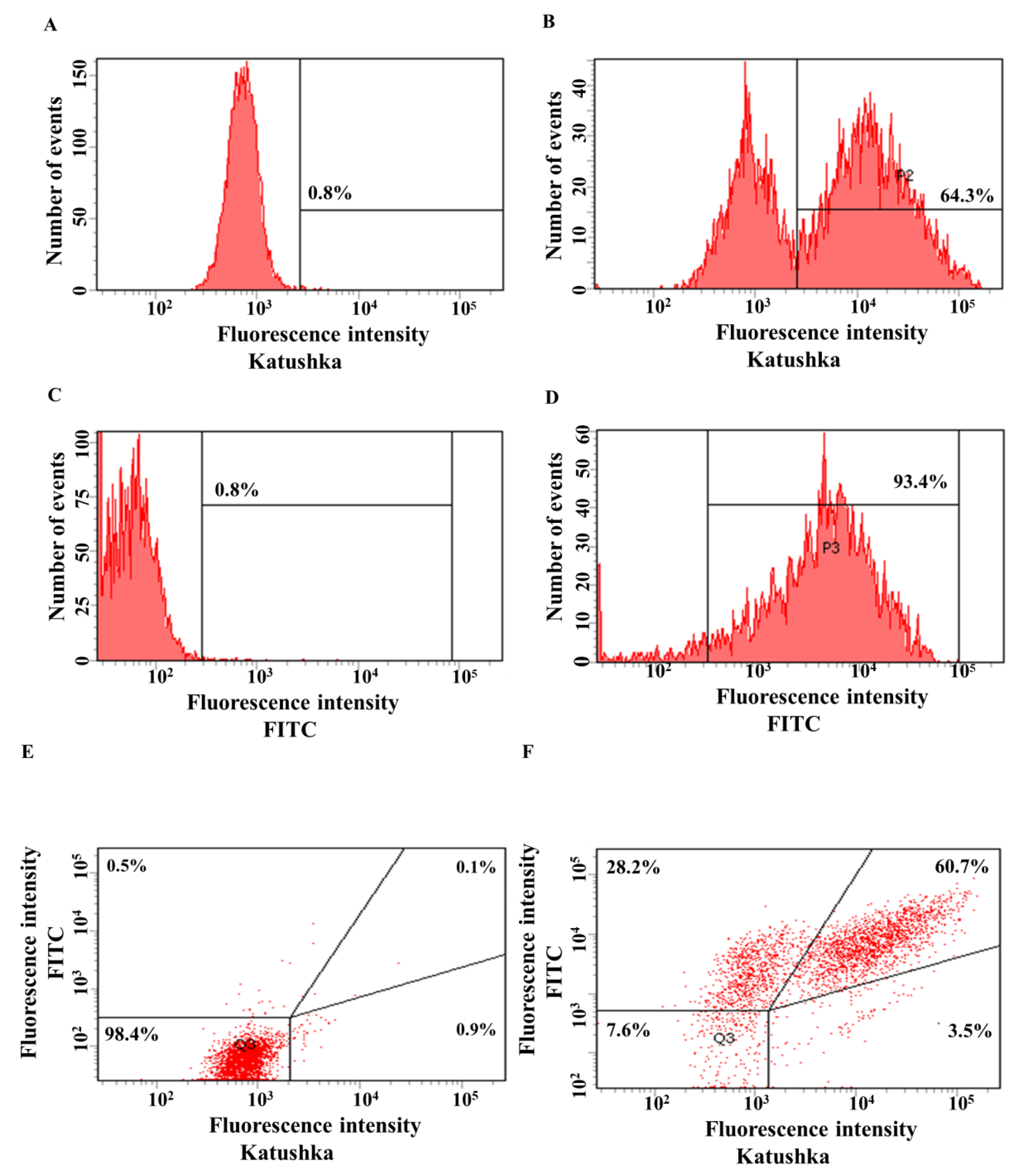
Figure 4.
Evaluation of the transduction efficiency of A431 cells by lentiviruses encoding Katushka2S and/or CD19. Flow cytometry, representative histograms (A–D), and scatter plots (E,F) are presented. (A)—negative control, non-transduced cells; (B)—A431(Kat+) cells expressing red fluorescent protein Katushka2S; (C)—negative control, non-transduced cells; (D)—A431(CD19+) cells expressing CD19 antigen; (E)—negative control, non-transduced cells are shown in the upper right corner; and (F)—A431(Kat+CD19+) cells simultaneously expressing Katushka2S and CD19 are shown in the upper right corner.
Figure 4.
Evaluation of the transduction efficiency of A431 cells by lentiviruses encoding Katushka2S and/or CD19. Flow cytometry, representative histograms (A–D), and scatter plots (E,F) are presented. (A)—negative control, non-transduced cells; (B)—A431(Kat+) cells expressing red fluorescent protein Katushka2S; (C)—negative control, non-transduced cells; (D)—A431(CD19+) cells expressing CD19 antigen; (E)—negative control, non-transduced cells are shown in the upper right corner; and (F)—A431(Kat+CD19+) cells simultaneously expressing Katushka2S and CD19 are shown in the upper right corner.
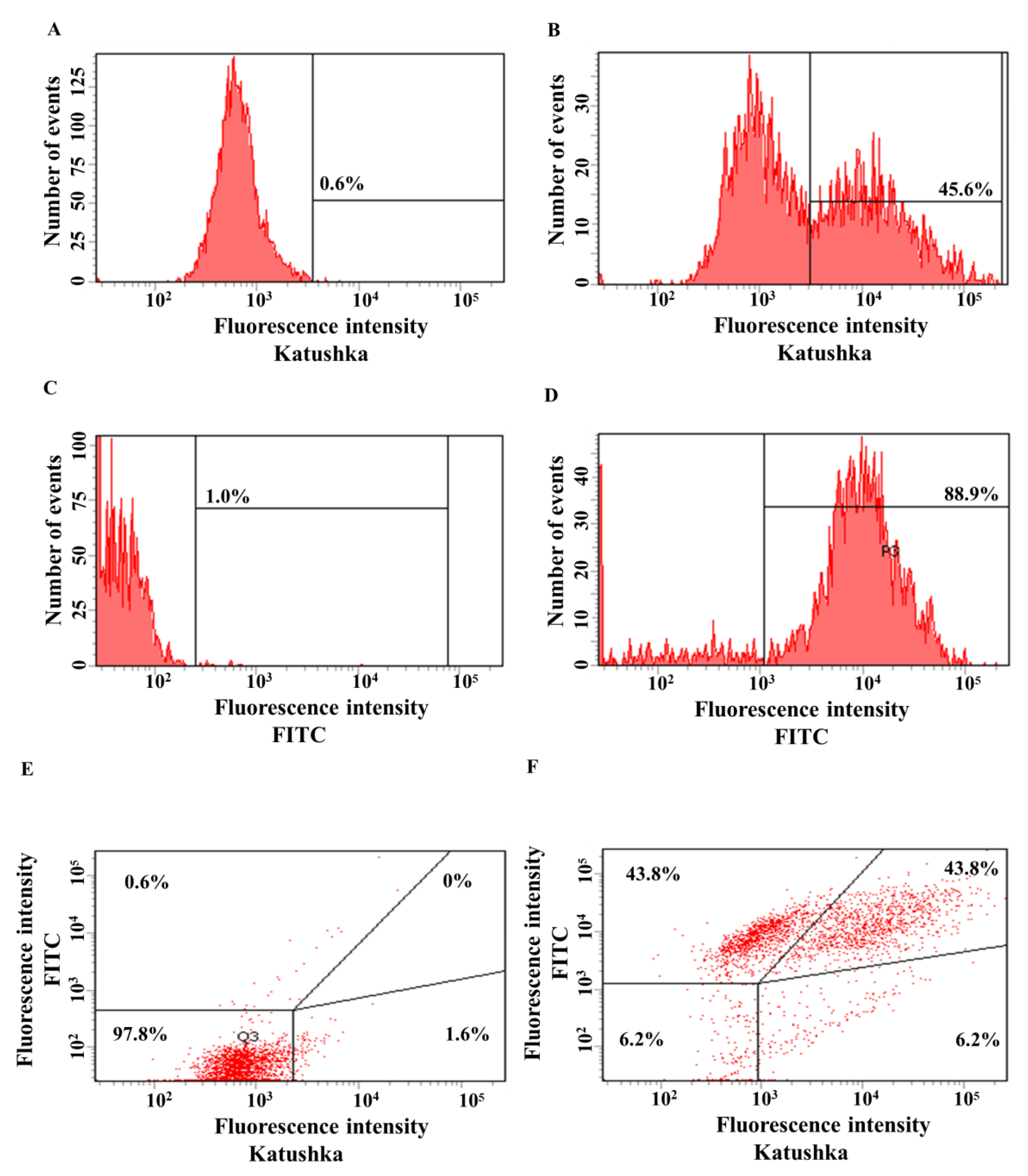
Figure 5.
Evaluation of the transduction efficiency of MDA-MB-231 cells by lentiviruses encoding red fluorescent protein Katushka2S and/or CD19 antigen. Flow cytometry, representative histograms (A–D), and scatter plots (E,F) are presented. (A)—negative control, non-transduced cells; (B)—MDA-MB-231(Kat+) cells expressing Katushka2S; (C)—negative control, non-transduced cells; (D)—MDA-MB-231(CD19+) cells expressing CD19; (E)—negative control, non-transduced cells are shown in the upper right corner; and (F)—MDA-MB-231(Kat+CD19+) cells expressing both Katushka2S and CD19 are shown in the upper right corner.
Figure 5.
Evaluation of the transduction efficiency of MDA-MB-231 cells by lentiviruses encoding red fluorescent protein Katushka2S and/or CD19 antigen. Flow cytometry, representative histograms (A–D), and scatter plots (E,F) are presented. (A)—negative control, non-transduced cells; (B)—MDA-MB-231(Kat+) cells expressing Katushka2S; (C)—negative control, non-transduced cells; (D)—MDA-MB-231(CD19+) cells expressing CD19; (E)—negative control, non-transduced cells are shown in the upper right corner; and (F)—MDA-MB-231(Kat+CD19+) cells expressing both Katushka2S and CD19 are shown in the upper right corner.
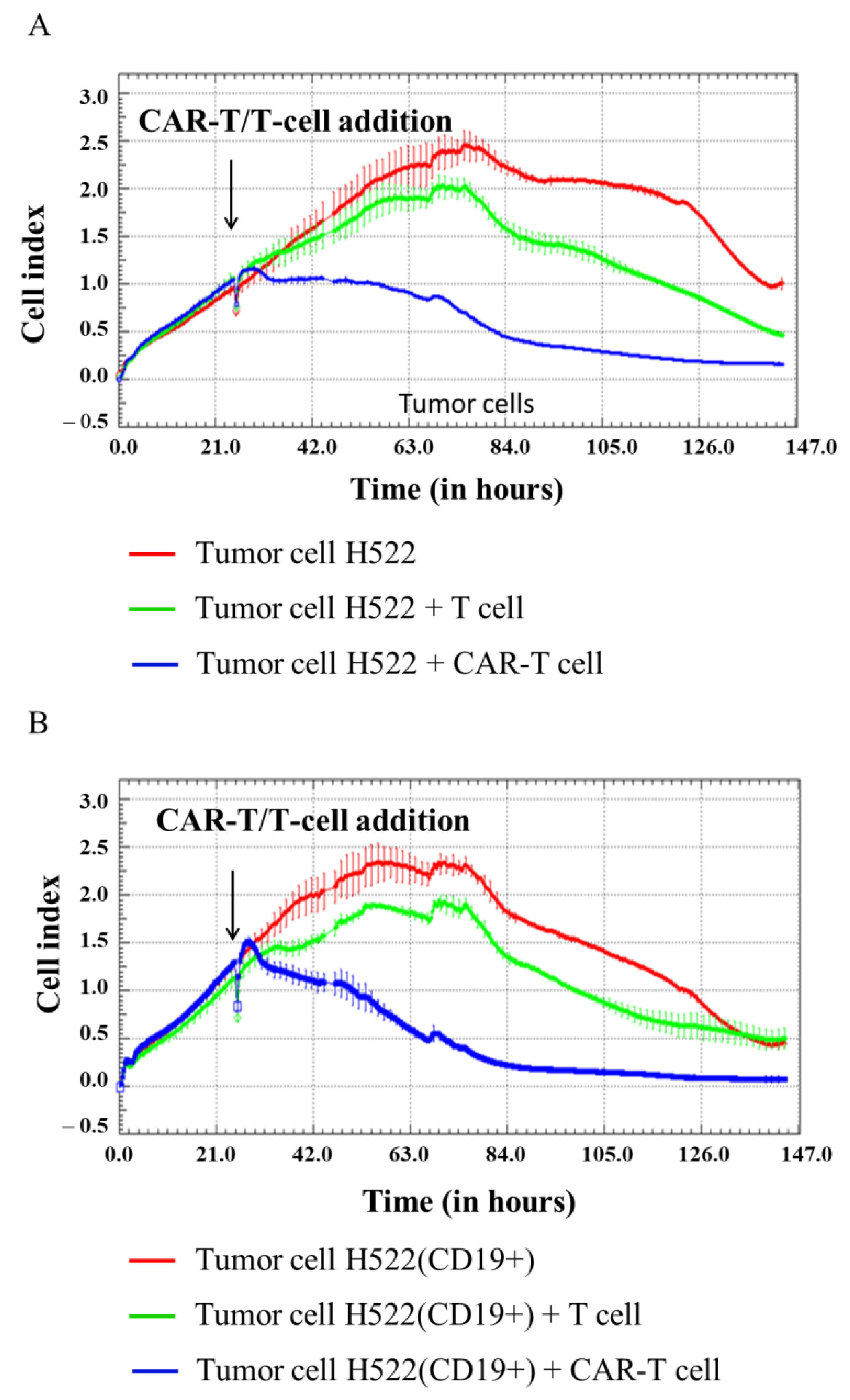
Figure 6.
Evaluation of the cytotoxicity of CAR-T cells against H522 and H522(CD19+) tumor cell lines using xCELLigence biosensor cell analyzer. Dynamic monitoring of proliferation of H522 and modified H522(CD19+) cells with and without the addition of T/CAR-T cells. Graphs of cell index versus time are presented as mean, the error bars indicate standard deviation (n = 3). (A)—H522 cells; (B)—H522(CD19+) cells.
Figure 6.
Evaluation of the cytotoxicity of CAR-T cells against H522 and H522(CD19+) tumor cell lines using xCELLigence biosensor cell analyzer. Dynamic monitoring of proliferation of H522 and modified H522(CD19+) cells with and without the addition of T/CAR-T cells. Graphs of cell index versus time are presented as mean, the error bars indicate standard deviation (n = 3). (A)—H522 cells; (B)—H522(CD19+) cells.
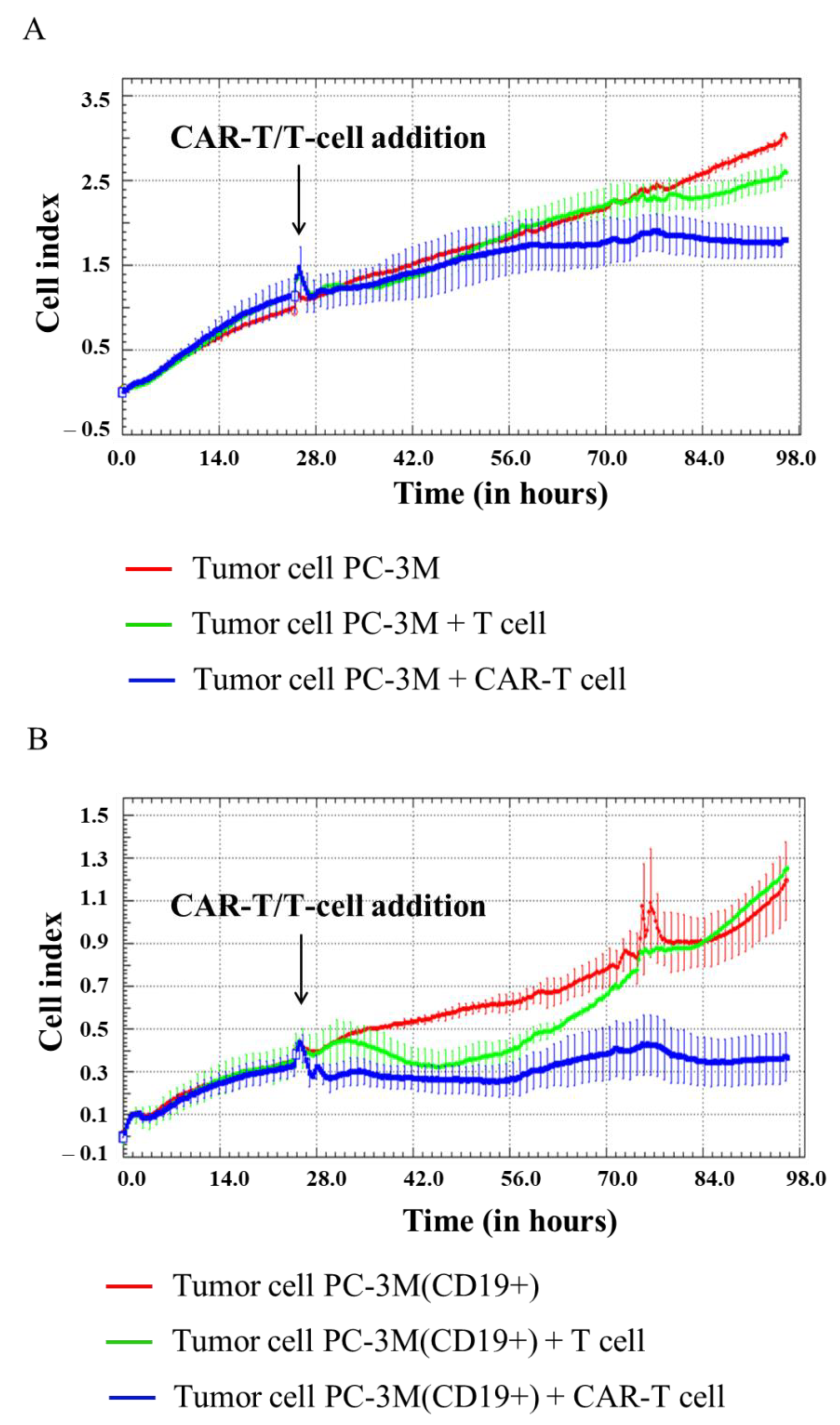
Figure 7.
Evaluation of the cytotoxicity of CAR-T cells against PC-3M and PC-3M(CD19+) tumor cell lines using xCELLigence biosensor cell analyzer. Dynamic monitoring of proliferation of PC-3M and modified PC-3M(CD19+) cells with and without the addition of T/CAR-T cells. Graphs of cell index versus time are presented as mean, the error bars indicate standard deviation (n = 3). (A)—PC-3M cells; (B)—PC-3M(CD19+) cells.
Figure 7.
Evaluation of the cytotoxicity of CAR-T cells against PC-3M and PC-3M(CD19+) tumor cell lines using xCELLigence biosensor cell analyzer. Dynamic monitoring of proliferation of PC-3M and modified PC-3M(CD19+) cells with and without the addition of T/CAR-T cells. Graphs of cell index versus time are presented as mean, the error bars indicate standard deviation (n = 3). (A)—PC-3M cells; (B)—PC-3M(CD19+) cells.
Figure 8.
Evaluation of the cytotoxicity of CAR-T cells against A431 and A431(CD19+) tumor cell lines using xCELLigence biosensor cell analyzer. Dynamic monitoring of proliferation of A431 and modified A431(CD19+) cells with and without the addition of T/CAR-T cells. Graphs of cell index versus time are presented as mean, the error bars indicate standard deviation (n = 3). (A)—A431 cells; (B)—A431(CD19+) cells.
Figure 8.
Evaluation of the cytotoxicity of CAR-T cells against A431 and A431(CD19+) tumor cell lines using xCELLigence biosensor cell analyzer. Dynamic monitoring of proliferation of A431 and modified A431(CD19+) cells with and without the addition of T/CAR-T cells. Graphs of cell index versus time are presented as mean, the error bars indicate standard deviation (n = 3). (A)—A431 cells; (B)—A431(CD19+) cells.
Figure 9.
Evaluation of the cytotoxicity of CAR-T cells against MDA-MB-231 and MDA-MB-231(CD19+) tumor cell lines using xCELLigence biosensor cell analyzer. Dynamic monitoring of proliferation of MDA-MB-231 and modified MDA-MB-231(CD19+) cells with and without the addition of T/CAR-T cells. Graphs of cell index versus time are presented as mean, the error bars indicate standard deviation (n = 3). (A)—MDA-MB-231 cells; (B)—MDA-MB-231(CD19+) cells.
Figure 9.
Evaluation of the cytotoxicity of CAR-T cells against MDA-MB-231 and MDA-MB-231(CD19+) tumor cell lines using xCELLigence biosensor cell analyzer. Dynamic monitoring of proliferation of MDA-MB-231 and modified MDA-MB-231(CD19+) cells with and without the addition of T/CAR-T cells. Graphs of cell index versus time are presented as mean, the error bars indicate standard deviation (n = 3). (A)—MDA-MB-231 cells; (B)—MDA-MB-231(CD19+) cells.
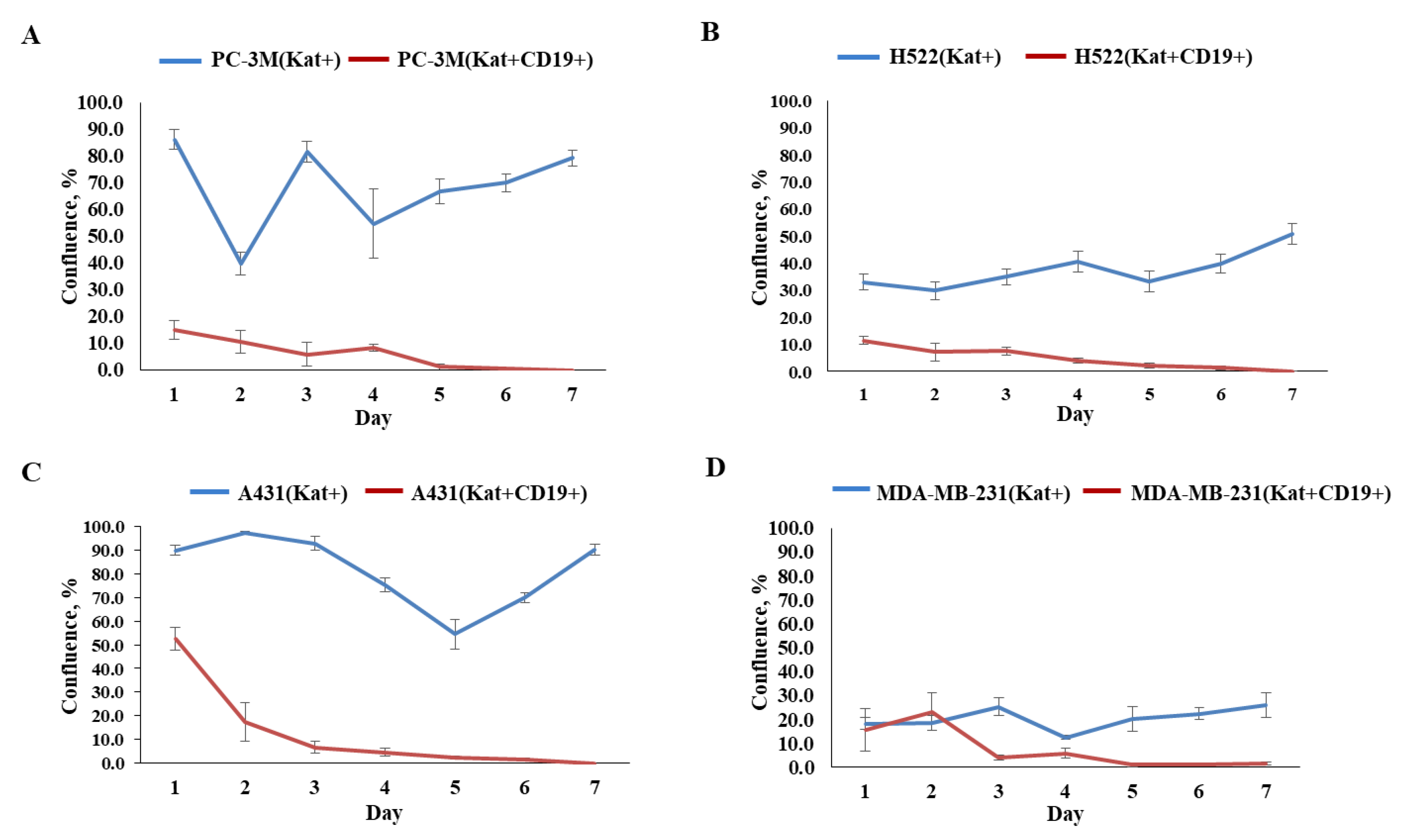
Figure 10.
Graphs representing comparative evaluation of CAR-T cells’ efficacy against modified tumor cells over 7-day period. (A)—comparative confluence of PC-3M(Kat+) and PC-3M(Kat+CD19+) cells after addition of CAR-T cells; (B)—comparative confluence of H522(Kat+) and H522(Kat+CD19+) cells after addition of CAR-T cells; (C)—comparative confluence of A431(Kat+) and A431(Kat+CD19+) cells after addition of CAR-T cells; (D)—comparative confluence of MDA-MB-231(Kat+) and MDA-MB-231(Kat+CD19+) cells after addition of CAR-T cells. The results are presented as mean, the error bars indicate standard deviation (n = 3, p < 0.05). Control tumor cells are shown in blue, modified CD19+ tumor cell lines in red.
Figure 10.
Graphs representing comparative evaluation of CAR-T cells’ efficacy against modified tumor cells over 7-day period. (A)—comparative confluence of PC-3M(Kat+) and PC-3M(Kat+CD19+) cells after addition of CAR-T cells; (B)—comparative confluence of H522(Kat+) and H522(Kat+CD19+) cells after addition of CAR-T cells; (C)—comparative confluence of A431(Kat+) and A431(Kat+CD19+) cells after addition of CAR-T cells; (D)—comparative confluence of MDA-MB-231(Kat+) and MDA-MB-231(Kat+CD19+) cells after addition of CAR-T cells. The results are presented as mean, the error bars indicate standard deviation (n = 3, p < 0.05). Control tumor cells are shown in blue, modified CD19+ tumor cell lines in red.
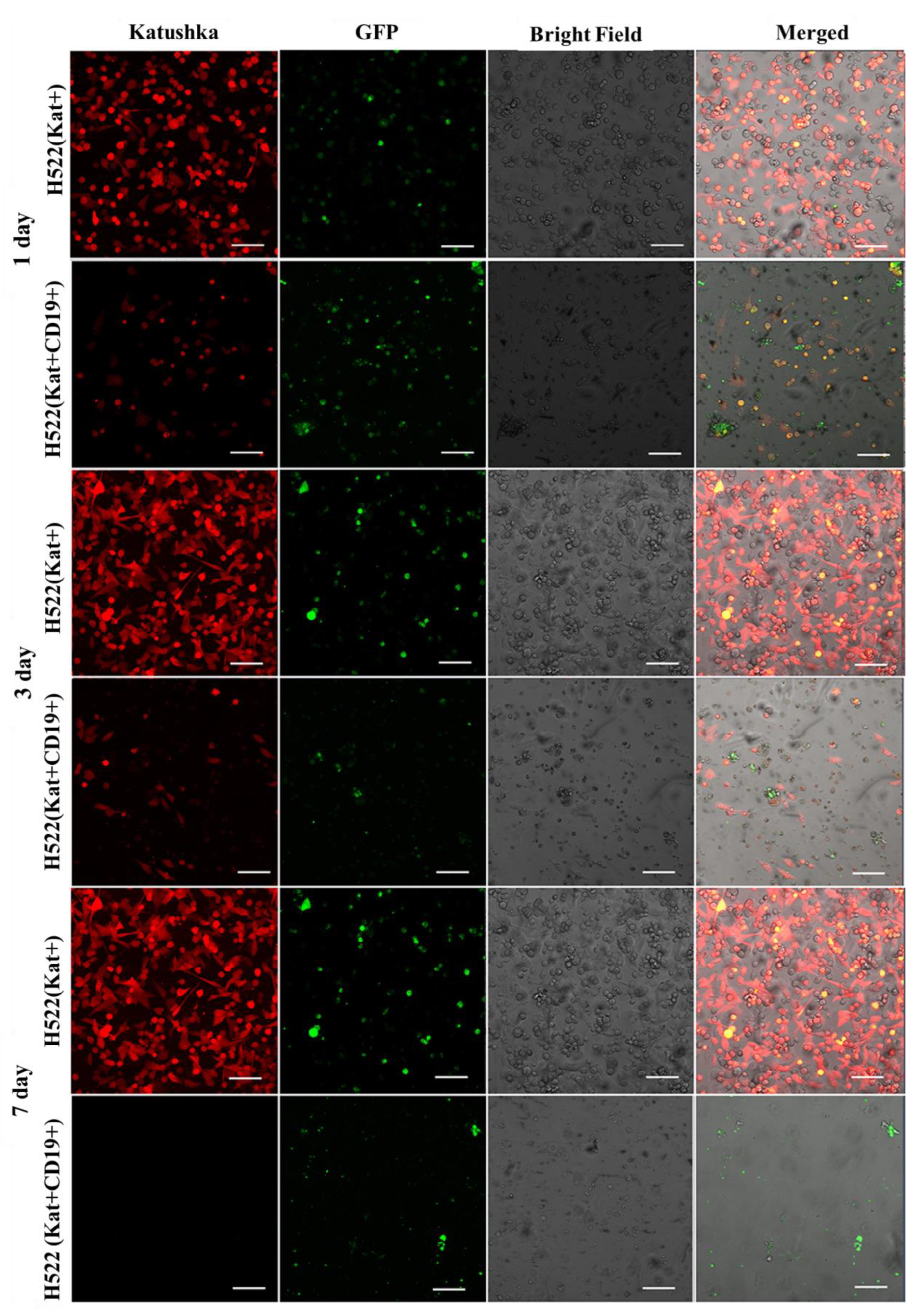
Figure 11.
Micrographs of H522(Kat+) and H522(Kat+CD19+) tumor cell monolayers treated with CAR-T cells for a period of 7 days. Light microscopy, fluorescence microscopy, and representative micrographs (n = 6) are presented, scale bar is equal to 100 µm. CAR-T cells—green fluorescence; H522(Kat+) and H522(Kat+CD19+) cells—red fluorescence.
Figure 11.
Micrographs of H522(Kat+) and H522(Kat+CD19+) tumor cell monolayers treated with CAR-T cells for a period of 7 days. Light microscopy, fluorescence microscopy, and representative micrographs (n = 6) are presented, scale bar is equal to 100 µm. CAR-T cells—green fluorescence; H522(Kat+) and H522(Kat+CD19+) cells—red fluorescence.
Figure 12.
Micrographs of PC-3M(Kat+) and PC-3M(Kat+CD19+) tumor cell monolayers treated with CAR-T cells for a period of 7 days. Light microscopy, fluorescence microscopy, and representative micrographs (n = 6) are presented, scale bar is equal to 100 µm. CAR-T cells—green fluorescence; PC-3M(Kat+) and PC-3M(Kat+CD19+) cells—red fluorescence.
Figure 12.
Micrographs of PC-3M(Kat+) and PC-3M(Kat+CD19+) tumor cell monolayers treated with CAR-T cells for a period of 7 days. Light microscopy, fluorescence microscopy, and representative micrographs (n = 6) are presented, scale bar is equal to 100 µm. CAR-T cells—green fluorescence; PC-3M(Kat+) and PC-3M(Kat+CD19+) cells—red fluorescence.
Figure 13.
Micrographs of A431(Kat+) and A431(Kat+CD19+) tumor cell monolayers treated with CAR-T cells for a period of 7 days. Light microscopy, fluorescence microscopy, and representative micrographs (n = 6) are presented, scale bar is equal to 100 µm. CAR-T cells—green fluorescence; A431(Kat+) and A431(Kat+CD19+) cells—red fluorescence.
Figure 13.
Micrographs of A431(Kat+) and A431(Kat+CD19+) tumor cell monolayers treated with CAR-T cells for a period of 7 days. Light microscopy, fluorescence microscopy, and representative micrographs (n = 6) are presented, scale bar is equal to 100 µm. CAR-T cells—green fluorescence; A431(Kat+) and A431(Kat+CD19+) cells—red fluorescence.
Figure 14.
Micrographs of MDA-MB-231(Kat+) and MDA-MB231(Kat+CD19+) tumor cell line monolayers treated with CAR-T cells for a period of 7 days. Light microscopy, fluorescence microscopy, and representative micrographs (n = 6) are presented, scale bar is equal to 100 µm. CAR-T cells—green fluorescence; MDA-MB-231(Kat+) and MDA-MB-231(Kat+CD19+) cells—red fluorescence.
Figure 14.
Micrographs of MDA-MB-231(Kat+) and MDA-MB231(Kat+CD19+) tumor cell line monolayers treated with CAR-T cells for a period of 7 days. Light microscopy, fluorescence microscopy, and representative micrographs (n = 6) are presented, scale bar is equal to 100 µm. CAR-T cells—green fluorescence; MDA-MB-231(Kat+) and MDA-MB-231(Kat+CD19+) cells—red fluorescence.
Figure 15.
Orthotopic micrograph of 3D structures formed by tumor cells. (A)—H522(Kat+CD19+), (B)—PC-3M(Kat+CD19+), (C)—A431(Kat+CD19+), and (D)—MDA-MB-231(Kat+CD19+) tumor cells. Confocal fluorescence microscopy and representative micrographs are presented (n = 6). Circles indicate examples of single CAR-T cells (green fluorescence) and areas of their direct contact with tumor cells (red fluorescence).
Figure 15.
Orthotopic micrograph of 3D structures formed by tumor cells. (A)—H522(Kat+CD19+), (B)—PC-3M(Kat+CD19+), (C)—A431(Kat+CD19+), and (D)—MDA-MB-231(Kat+CD19+) tumor cells. Confocal fluorescence microscopy and representative micrographs are presented (n = 6). Circles indicate examples of single CAR-T cells (green fluorescence) and areas of their direct contact with tumor cells (red fluorescence).
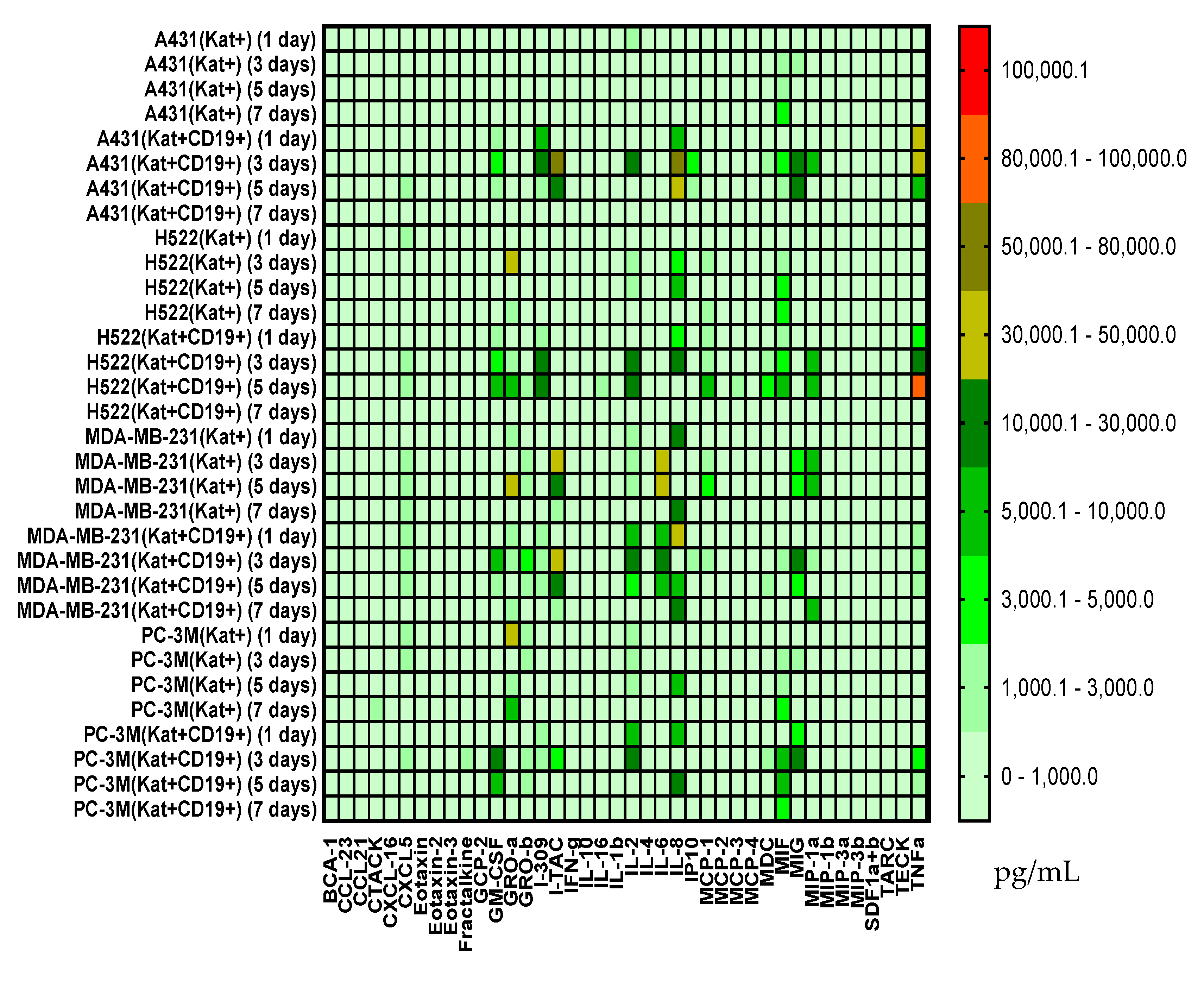
Figure 16.
Heat map of cytokines and chemokines multiplex arrays in the culture medium after co-cultivation of CAR-T cells with H522(Kat+), H522(Kat+CD19+), PC-3M(Kat+), PC-3M(Kat+CD19+), A431(Kat+), A431(Kat+CD19+), MDA-MB-231(Kat+), and MDA-MB-231(Kat+C19+) tumor cells at different time points (days 1, 3, 5, and 7).
Figure 16.
Heat map of cytokines and chemokines multiplex arrays in the culture medium after co-cultivation of CAR-T cells with H522(Kat+), H522(Kat+CD19+), PC-3M(Kat+), PC-3M(Kat+CD19+), A431(Kat+), A431(Kat+CD19+), MDA-MB-231(Kat+), and MDA-MB-231(Kat+C19+) tumor cells at different time points (days 1, 3, 5, and 7).

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}















