
Zebrafish as a Model to Study Retinoic Acid Signaling in Development and Disease


Department of Biological Sciences, Center for Stem Cells and Regenerative Medicine, Center for Zebrafish Research, Boler-Parseghian Center for Rare and Neglected Diseases, Warren Center for Drug Discovery, University of Notre Dame, Notre Dame, IN 46556, USA
*
Author to whom correspondence should be addressed.
Biomedicines 2023, 11(4), 1180; https://doi.org/10.3390/biomedicines11041180
Submission received: 16 March 2023
/
Revised: 6 April 2023
/
Accepted: 13 April 2023
/
Published: 15 April 2023
(This article belongs to the Special Issue Zebrafish Models for Development and Disease 4.0)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Retinoic acid (RA) is a metabolite of vitamin A (retinol) that plays various roles in development to influence differentiation, patterning, and organogenesis. RA also serves as a crucial homeostatic regulator in adult tissues. The role of RA and its associated pathways are well conserved from zebrafish to humans in both development and disease. This makes the zebrafish a natural model for further interrogation into the functions of RA and RA-associated maladies for the sake of basic research, as well as human health. In this review, we explore both foundational and recent studies using zebrafish as a translational model for investigating RA from the molecular to the organismal scale.
Keywords:
retinoic acid; vitamin A; development; zebrafish; neurogenesis; kidney; heart; endoderm; cancer; regeneration1. Introduction
For nearly 100 years, vitamin A in the maternal diet has been linked to normal embryonic ontogeny in vertebrates. Initially, this was based on observations in female pigs and rats that subsistence on nourishments lacking vitamin A during pregnancy was associated with a plethora of birth defects in newborns ranging from eye abnormalities to genitourinary defects [1,2,3,4,5,6,7,8]. Continued nutrition research uncovered a complex spectrum of congenital malformations that occurred consequent to a maternal vitamin A deficient (VAD) diet, which came to be known as the VAD syndrome [9]. The malformations included defects in the central nervous system, eyes, ears, heart, lungs, limbs, skin, and urogenital system [9]. These and subsequent studies helped to stimulate continued research on how retinoids, the biologically active metabolites of vitamin A, modulate development. Many fundamental insights were uncovered by investigating the teratogenic effects of RA—how global or local exposure impacted normal processes [10,11,12,13,14,15,16]. RA signaling is now appreciated as being essential for the genesis of nearly every vertebrate tissue and organ [10,11,12,13,14,15,16]. The wide range of RA functions include patterning of the body axis, regional patterning of the central nervous system, neurogenesis, limb development, and pleiotropic roles in organogenesis [10,11,12,13,14,15,16].
The zebrafish, Danio rerio, is a powerful experimental model for studying the roles of RA signaling during ontogeny [17] due to its ex utero development, large embryonic size, and transparent nature—features which have facilitated classical and chemical genetic approaches [18,19]. Here, we provide an overview of RA biology and the zebrafish model, and discuss how zebrafish have been used to further our understanding about the roles of RA during embryogenesis of the neural plate, kidney, heart, blood, endoderm, and in disease states like cancer.
2. RA and Zebrafish, a Background
2.1. Evolutionary Perspective and Gene Regulation
RA-mediated morphogenic signaling was once thought to represent an evolutionary split between chordates and non-chordates, as RA is necessary for the anterior-posterior (AP) axis and later germ layer organization in early development [20]. However, analysis involving both genomic and proteomic methods has highlighted evidence that traces essential RA-related machinery to far more ancient phylogenies, such as mollusks [20]. Even within chordates, machinery slightly differs along the evolutionary timeline. Within tunicates and other invertebrate chordates, the number of RA receptors is markedly lower due to the two whole genome duplications associated with vertebrate evolution [21]. Despite the change in the number of receptors, the importance of RA in development is still highly conserved across phyla. Tunicates such as C. intestinalis treated with exogenous RA were found to have limited neural tube closure, a feature conserved through zebrafish and into humans [22,23,24].
Across species, researchers have linked RA, its receptors, and the accompanying response elements as upstream regulators to homeobox (hox) genes, a gene family highly associated with AP positioning, neural crest organization, and overall organogenesis later in development [25,26,27,28]. In the zebrafish genome, hox genes are isolated to clusters, and this multiple cluster model is conserved throughout vertebrates [29,30,31]. Though these clustered identities are well conserved, the number of clusters containing protein coding regions does differ. In comparison to mammals which have four of these clusters, zebrafish have seven, the odd number due to a condensation post genome-wide duplication [32,33,34]. Genes within each of these clusters are genetically positioned 3′ to 5′ in the order in which they are transcribed, meaning upstream elements represent coding regions for anterior cell fates [35]. This collinear positioning is also sensitive to RA, as exogenous RA treatments can lead to the initiation of transcription for upstream/anterior elements in locations which normally require posterior hox elements, and the opposite for downstream/posterior identities [36,37,38].
Other than hox gene regulation, RA has a regulatory footprint in various other signaling pathways that aid in developmental processes. One signaling family that should be duly noted is the Fibroblast Growth Factors (FGF) and the ensuing cascades. FGF works on an opposing gradient to RA in the presomitic mesoderm, among other tissues, and is vital for posteriorizing cell fates in cell populations such as the neural plate [39]. In the presence of exogenous RA or RA agonists, fgf8 expression within caudal neural plate explants is markedly decreased [39]. In these same experiments performed by Diez el Corral et al. utilizing vitamin-A deficient quail, the researchers found that fgf8 expression extends to more anterior fates, thus portraying RA as a negative regulator of FGFs [39]. In the converse manner, FGFs have the ability to increase RA signaling via cyp26 suppression. Through Cyp26 enzymes, fgf8 transcription works to create a margin between the anterior and posterior neural ectoderm in the earliest stages of gastrulation, thus allowing for early organization for future cell fates [40]. RA has a regulatory role on FGF signaling in other developmental processes as well, for example, in defining the cardiac and forelimb progenitor fields [10,11,12,13,14,15,16,17].
2.2. Biogenesis, Metabolism, and Receptors Involved
In zebrafish and other vertebrates, de novo RA synthesis does not occur, thus these species rely upon dietary intake of vitamin A or vitamin A preceptors for canonical RA signaling (Figure 1). Forms of vitamin A are present endogenously in the early embryo from maternal sources, e.g., transferred across the placental circulation in mammals or deposited within the egg yolk as carotenoid stores in oviparous species [41,42,43,44,45]. The forms include retinol, retinyl esters, and plant-derived provitamin A carotenoids like β-carotene and β-cryptoxanthin [41,42,43,44,45]. These plant-derived carotenoids must be converted to retinoids via oxygenases such as ββ-carotene-15,15-oxygenase (bcox) [46]. In zebrafish, knockdown of the bcox gene leads to phenotypes seen in VAD animals; thus, plant-derived carotenoids and their corresponding reducing enzymes are essential for proper development [46].
Once retinol has entered either the serum or the extracellular matrix, retinol binding proteins (RBPs) bind these free retinoids for escort to the cell membrane (Figure 1). This method of transport is found not only in the serum, but in the yolk and the yolk syncytial layer in early development. These bound RBPs act as ligands for the transmembrane transporters of retinol, STRA6 and STRA6l (stimulated by retinoic acid 6, stimulated by retinoic acid 6 like, formerly RBPR2) (Figure 1) [47,48,49,50]. STRA6/l serve as bidirectional gatekeepers of retinol, as RBPs bind to the extracellular facing domain, and cellular retinol binding proteins (CRBPs) bind the cytoplasmic domain [51,52].
Once retinol has bound CRBPs within the cytosol, one of two fates occurs (Figure 1) [11,12,13]. In the presence of lecithin-retinol acyltransferase (LRAT), CRBPII complexes are responsible for the transformations of retinol to retinyl esters [51,52]. These retinyl esters are a stable storage molecule that can be reduced back to retinol in times of RA deficiency [53]. The second fate for CRBP-bound retinol is the oxidation to retinaldehyde via retinol or alcohol dehydrogenases (RDHs or ADHs) (Figure 1) [54]. Once the retinal has been synthesized, the last oxidation of retinaldehyde to RA is facilitated by retinaldehyde dehydrogenases (RALDH, also known as ALDHs) [55,56].
Though several (R)ALDHs are sensitive to retinol, we wish to highlight the (R)ALDH1A family and the three proteins that reside within, two of which are present in zebrafish and other teleosts: aldh1a2 and aldh1a3 [57]. In humans, ALDH1A1 is not associated with development; rather, the enzyme has a role in alcohol metabolism and has been linked to several cancers [58]. In zebrafish, aldh1a2 represents the best described ALDH responsible for RA signaling in development. In the loss of aldh1a2, phenotypes in zebrafish development mimic those of VAD embryos in zebrafish and mammals [59,60,61,62,63,64,65]. For example, the death of neural crest cells at 24 h post fertilization (hpf), untidy hox expression patterns, and lack of pectoral fin skeletal structure were noted in the aldh1a2 null fish [59]. In zebrafish, aldh1a3 knock down via morpholino results in decreased eye size and overall aberrant eye morphology [66]. This phenotype of poor eye formation is thought to be conserved in humans, as mutations within aldh1a3 result in anophthalmia [67,68]. Transcriptionally within development, aldh1a2 and aldh1a3 expression show little temporal overlap [57,59,62,64,69,70]. In whole embryo qRT-PCR experiments performed by Xi et al., aldh1a2 expression begins during mid gastrulation and tapers off dramatically during segmentation activities [71]. In other work, aldh1a3 expression was shown to initiate in early somitogenesis/segmentation [57,69]. This falls nearly in step with the earlier work of Liang et al., who showed aldh1a3 expression to initiate at 14 hpf [69]. This mild discrepancy highlights the need for updated and further investigation of the regulatory networks involved in RA synthesis machinery through the continued application of effective methods, such as qRT-PCR [72,73], and whole mount in situ hybridization (WISH) [74,75,76,77].
Once synthesized, RA has two direct paths to take (Figure 1). The first of which is to bind to a cellular retinoic acid binding protein (CRABP) [78,79] that facilitates transport into the nucleus to bind an RA receptor (Figure 1) [80]. RA receptors represent a body of nuclear receptor superfamily members [81]. In humans and many other vertebrates, three RARs exist (ɑ, β, ɣ), and these RARs are accompanied by retinoid X receptors (RXRs, also: ɑ, β, ɣ) [81,82,83,84]. The coupling of these two receptors creates a complex that will subsequently bind DNA to regulate gene transcription at discrete sequences known as retinoic acid responsive elements (RAREs) (Figure 1) [85,86,87]. The important distinction between RARs and RXRs is the molecules that target them. RARs bind all-trans RA (ATRA) and 9-cis RA, while RXRs are solely targeted by the 9-cis RA isomer. Once the ligand and receptor are bound, a heterodimer forms between the twin receptors. Though it is thought that RARs only dimerize with RXRs, it is known that RXRs possess the ability to form homodimers with non-RARs such as Pparɣ to play roles in other pathways [88,89,90,91]. Interestingly, zebrafish, as a result of genome duplication and subsequent condensation, possess four RARs (two RARɑ and two RARɣ). Despite the loss of RARβ from the zebrafish genome, the corresponding RXRs were retained through the condensation, and are active in the regulation of various conserved developmental and regenerative processes [92,93,94,95,96,97,98,99].
Due to the need for RA signaling to occur in a gradient that is discrete in terms of concentration, an equally precise mechanism for RA degradation is a necessity, thus providing a second fate for newly synthesized RA (Figure 1). Primarily, three enzymes belonging to the cytochrome p450 26 subfamily (CYP26A1, CYP26B1, and CYP26C1) are tasked with RA metabolism [71,100,101,102,103,104,105,106,107,108]. These enzymes are crucial to tightly control RA distribution and prevent inappropriate signaling within the embryo [109,110]. Zebrafish similarly possess these three cyp26 genes (cyp26a1, cyp26b1, and cyp26c1), which are closely conserved with humans [71,100,101,102,103,104,105,106,107,108], and are likewise tightly controlled [111,112]. Adjunct functions of each of these Cyp26 enzymes do exist, however, overlaps in tissue-level spatial expression patterns are present [113]. In the course of development, the genes associated with these RA-metabolizing enzymes are found to be activated before synthesis machinery such as aldh1a2 and aldh1a3 is transcribed [71]. This accumulation of Cyp26 enzymes before gastrulation indicates potential priming of progenitor populations before the advent of zygotic RA synthesis [71]. This may also be evidence of a fourth enzyme with potential redundant and compensating retinal oxidizing capabilities in early development that is yet to be fully characterized [113]. The presence of Cyp26 family enzymes is also attributed to creating “RA sinks”, which are regions of tissue that act as collecting areas for diffuse/surplus RA [69,111,112,114,115]. These “sinks” occur in part due to RAREs present in the promoters of the Cyp26a1 gene expression [116,117,118,119,120], and they contribute to the creation of RA morphogen gradients in the early embryo [121,122,123,124,125,126,127].
2.3. Zebrafish as a Model Organism
The zebrafish presents as a natural model for developmental work [128] as well as disease modeling [129]. Zebrafish are non-amniotic vertebrates that are fertilized and develop externally within a transparent chorion [130]. This ontogeny can be witnessed readily from the single-cell stage, allowing for genetic manipulation via microinjection and other embryological approaches as well [131,132,133]. The transparent chorion and translucency of the early embryonic stages also enable the precise timing of stage-directed treatments. Cleavage stages transpire rapidly, occurring within the first six hours of development, followed by gastrulation within the first twelve, then subsequent organogenesis from the germ layers [130]. By 24 hpf, a full body plan exists [130]. Within two to three months, the zebrafish is able to breed.
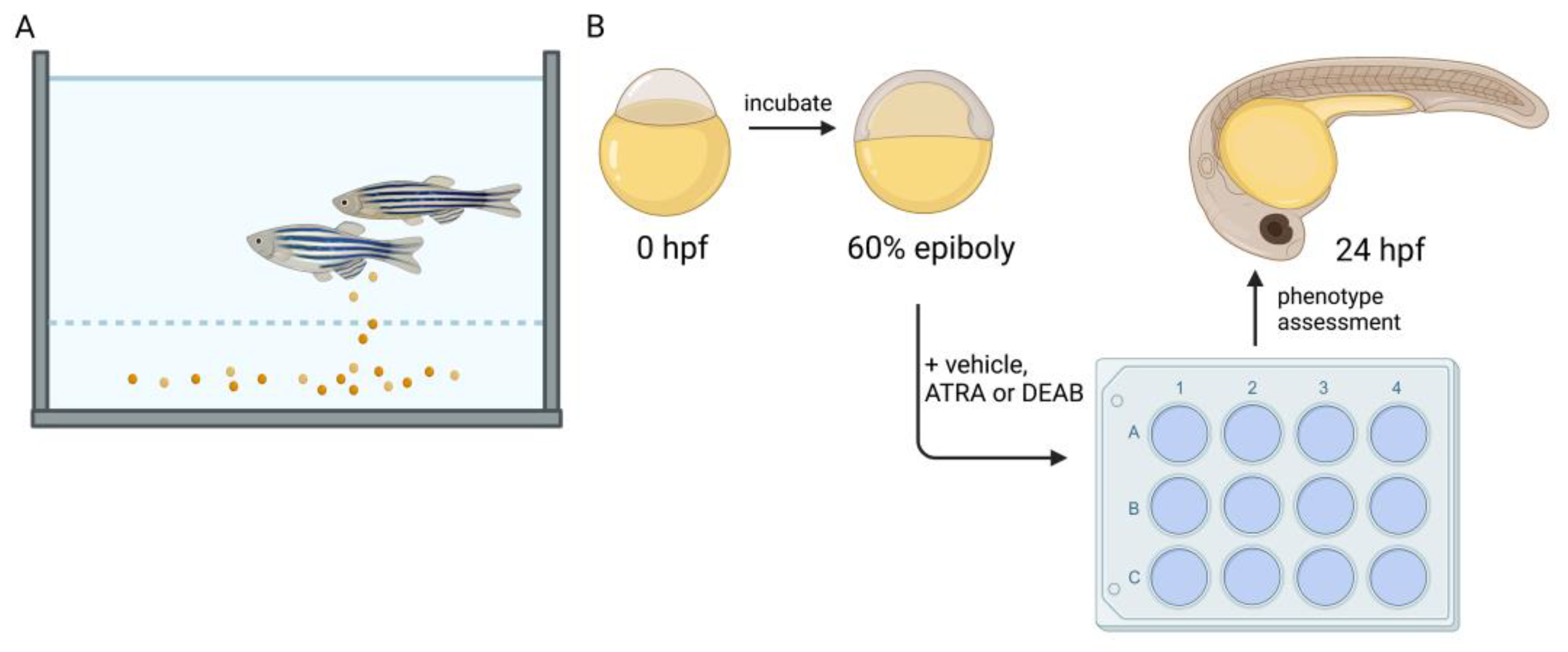
Zebrafish also possess a high fecundity, allowing for genetic screens [134,135,136,137,138,139,140,141,142,143,144] and high throughput assays to assess varied compounds and their role in developmental processes [145,146,147,148,149,150,151,152]. For example, RAR and RXR agonists/antagonists and their role in developmental processes can be explored by adding these compounds at various times (Figure 2). This short generation time paired with high fecundity also allows for large scale forward and reverse genetic screens. Further, in contrast to mammals, zebrafish possess the ability to regenerate many complex tissues and organs such as the kidney, heart, fin, and eye [153,154,155,156,157]. The zebrafish genome has been sequenced and is well annotated, allowing for conservation of biomarkers in health and disease [158]; and there are extensive centralized resources made possible by community efforts [159,160,161]. Genetically, ~70% of human protein coding genes share an ortholog with zebrafish [158]. In comparison to mammalian models, such as mouse and rat, the zebrafish is economically efficient in terms of the space needed to house adequate colonies. The use of zebrafish as a model has become increasingly important for providing insights into development and disease, given the nature and feasibility of many approaches that are unique to the use of this species. For the aforementioned reasons of rapid development, short generation time, and ease of use for screens that may utilize large chemical drug/libraries, zebrafish will continue to be a valuable model for elucidating the roles of RA in development.
3. RA in Development
3.1. Neural Plate Progenitors
Early hindbrain development is reliant on proper RA signaling. During this process, RA synthesis is thought to be acted on by pbx2 and pbx4 to regulate early hindbrain fate decisions [162]. In aldh1a2-deficient zebrafish neckless (nls) mutants [59], the down regulation of RARɑ leads to imprecise regulation of hoxb4. The neuroectoderm of 14-somite stage nls mutants lacks hoxb4 expression, despite ubiquitous expression in mesodermal tissues [59]. However, by the 16-somite stage (roughly one hour later), hoxb4 expression is restored to neural tube progenitors within nls mutants, thus implying a potential compensatory mechanism of RA-mediated hoxb4 expression. Both RA and FGF gradients work synergistically to develop positional identities within the rhombomeres, a group of vital cell populations necessary for early neural crest migration [163,164,165,166].
Through the exogenous treatment of embryos with the aldehyde dehydrogenase inhibitor molecule N,N-diethylaminobenzaldehyde (DEAB) (Figure 2) starting at the 4 hpf stage, rhombomere 5 and 6 identities were found to be lost at 16 hpf, implying the importance of RA in signaling fates in early hindbrain populations [166]. Other work performed in zebrafish showed that loss of RAR signaling via treatment with a pan-RAR antagonist led to not only a lack of posterior hindbrain identity, but also an increase in the domain of anterior rhombomeres two, three, and four [61]. This implies that the two tailed gradient of RA is opposed by Cyp26 enzymatic activity at both the far anterior and posterior ends of the developing fish. In work performed by Qiu et al., RA, FGF, and other morphogenic gradients were wonderfully computationally modeled to highlight the precision necessary for essential fates within the developing hindbrain and the associated rhombomeres within zebrafish [167].
In later points of development within the forebrain, loss of both aldh1a2 and aldh1a3 results in altered yet intact expression of fgf8 and shh, implying RA is not solely responsible for early forebrain organization in mice [168]. In zebrafish, depletion of hmx4, a homeobox gene and ortholog of the human HMX1, results in a significant decrease in aldh1a2 expression which leads to lack of neural tube closure [169]. In these same studies conducted by Gongal et al., the addition of exogenous RA to hmx4 morphants rescues gli3 expression and allows for regular forebrain development, suggesting that hmx4 works via RA to mediate Shh activation [169]. This work opens exciting new avenues for investigation into RA and its role in forebrain development.
Eye development in zebrafish as well as other vertebrates originates within the eye field, a primordial section of the neural plate. Transcriptionally, a grouping of factors known as eye field transcription factors (EFTF) are credited with this specification. These factors include homeodomain genes such as rx3, otx2, and lhx2, and pax6 [170]. In RNA-sequencing analysis of rx3-/- fish at 13 hpf, select RA-related orphan receptors were found to be transcriptionally downregulated [171]. Loss of this EFTF and the resulting down regulation of RA-associated machinery may imply transcriptional priming within these early eye field cells for RA that is used in later regulation of optic development.
3.2. Kidney
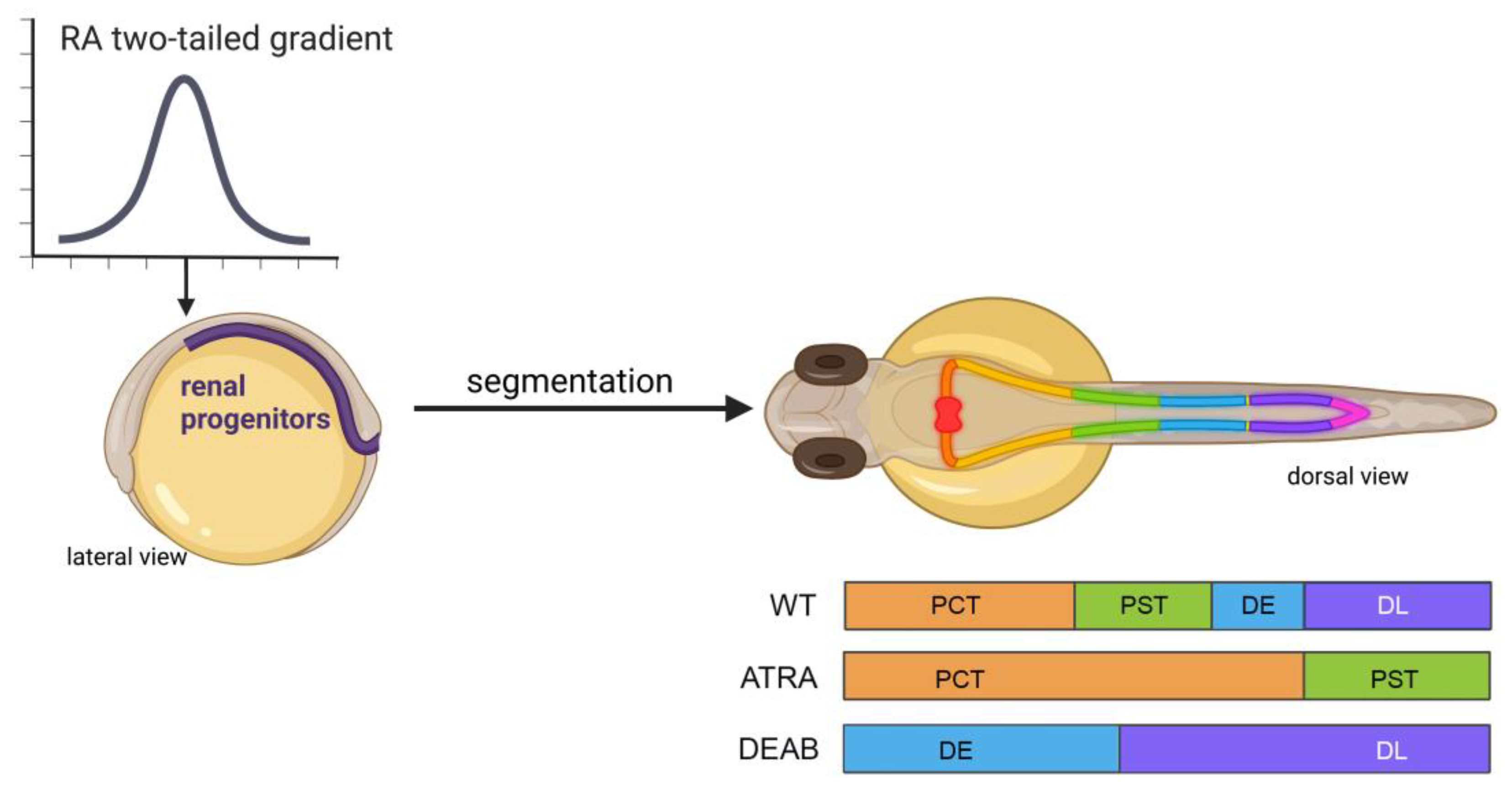
The nephron is the functional unit in the kidney, and it is tasked with several critical physiological roles: the filtering of the blood through the glomerulus, the facilitation of solutes in and out of circulation via the tubules, and the passage of waste for dismissal in the collecting duct [172]. In higher vertebrates, the pronephros is the first transient form of the kidney, followed by the mesonephros, and the final formation of the kidney is named the metanephros [172]. In zebrafish, only the first two, the pronephros and mesonephros, are formed [173]. Pronephros development in zebrafish begins during the process of intermediate mesoderm formation through the action of signals derived from Bone Morphogenic Proteins (BMPs) [174]. As the intermediate mesoderm forms, RA signals originating from the paraxial mesoderm orchestrate the genesis of the bilateral nephrons [64,175]. These movements can be visualized through the use of transgenic lines expressing renal progenitor field markers such as lh1xa, pax2a, and pax8 [176].
In the presence of exogenous (10−6 M) ATRA exposure from 60% epiboly/6 hpf stage to the six-somite stage, two of these same markers, pax2a and pax8, are found to have the non-discrete spatial expression pattern seen in regular development [175]. In this same line of experiments, use of the RA synthesis inhibitor DEAB at the same six-somite stage results in rostralization of evi1 (mecom) [175] a downstream mediator of Fgf signaling [177] and a marker for more distal tubule fates at the 28-somite/24 hpf stage [64,178]. Further analysis of the inhibition of RA synthesis through the comparison of aldh1a2 knockdown via morpholino oligonucleotide and chemical inhibition via DEAB shows that the time of RA synthesis is crucial to the tubule phenotype within the nephron; and it further confirms the redundancy within ALDH enzymatic expression patterns [175]. In aldh1a2 knockdown fish, all of the tubules form by 48 hpf; however, distal tubule fates expand rostrally at the cost of anterior segments [175]. In the chemical inhibition of RA synthesis via DEAB, time of addition experiments concluded that RA is most needed in tubule fate decisions between 60% epiboly and 15-somite stages, or approximately 6–16 hpf [175]. Lack of sufficient RA synthesis during this period of gastrulation and somitogenesis leads to the complete loss of proximal cell fates (Figure 3) [175]. This work concluded that RA works to posteriorize proximal cell fates in juxtaposition to Fgf signaling. Though loss of RA is associated with inefficient pronephros formation, overproduction or addition of exogenous RA can be equally detrimental to the organization of the tubules. At the same ATRA concentration (10−6 M) used to determine that intermediate mesoderm patterning is sensitive to opposing RA and FGF-inducing gradients, exposure to such high levels of ATRA from 90% epiboly to the 5-somite stage (5–10 hpf) leads to complete loss of distal segment fates and highly increased proximal convoluted tubule (PCT) and proximal straight tubule (PST) populations at the 24 hpf/28-somite stage (Figure 3) [175]. This work, and others involving the nephron and the associated tubule dependence on RA for regulation of segmentation, is well characterized, and the investigation of the downstream gene regulatory network has revealed a number of transcription factors crucial for fashioning segment fates, such as sim1a, tbx2a/b, and emx1, among others [64,179,180,181,182,183,184,185]. However, openings emerge for further interrogation due to the ever-evolving field of RA biology, the ease of performing chemical/drug/biologic screens in the zebrafish model, and the prevalence of next generation sequencing approaches in development.
Within the nephron, a subpopulation of aptly named multiciliated cells (MCCs) project clusters of motile cilia into the tubule lumen and govern fluid flow [186]. MCCs are detectable via WISH as early as the 10-somite stage [186]. Much like the nephron tubule populations, MCCs are dependent on RA signaling for differentiation from the renal progenitors [178]. In the proposed MCC regulatory network, RA works upstream to downregulate mecom, a positive regulator of Notch, and a factor for distal cell fates [178]. In knockdown of mecom and the subsequent addition of DEAB from late gastrulation to 24 hpf, MCC formation was almost completely rescued in terms of the domain they occupied within the nephrons and the position of this domain in the developing fish [178]. These experiments involving RA signaling and MCC fate determinants highlight exciting avenues of future interrogation into the potential regenerative capacities of MCCs after injury, which may lend potential insights into human disease.
3.3. Heart
From its genesis, the zebrafish heart is reliant on RA for correct spatial patterning of progenitors. Work performed by Keegan et al. delicately described both RA addition and the time in which RA is necessary for restricting progenitor fates [187]. When they treated embryos with a pan-RAR antagonist and then assessed cmlc2 and nkx2.5 expression, which marking cardiomyocytes precardiac mesoderm, respectively, they observed increased domains of expression at the 16-somite stage, suggesting RA works in restricting cardiac cell fates in post gastrulation fish [187]. This hypothesis was then confirmed through the use of exogenous ATRA treatments spanning a time point straddling gastrulation initiation leads to lessened cmlc2 expression at the 18-somite stage [187]. The mechanism of RA restricting the cardiac field was further characterized as being mediated by cdx4 and cdx1a expression. In this process, much like in hindbrain development, RA works through downregulating cdx4 expression, thereby activating cyp26a1 expression to metabolize RA within the cell [188,189]. This hypothesis further postulates the intricate nature of RA self-regulation, as RAREs exist as regulatory units of cyp26/Cyp26/CYP26 family expression [116,117]. Other work involving regulators of cardiac cell fate such as NR2F have been found to have RARES within these factors’ promoter regions, which in the presence of DEAB leads to a decreased nr2f1a domain. The loss of nr2f1a/b contributes to a relative expansion of the nkx2.5 domain, a commonality seen with RAR antagonism.
Further investigation into RA and its role in early heart field regulation revealed that RA works indirectly through the homeobox gene hoxb5b to limit cardiac progenitor populations [190]. Other work established that RA and FGF signaling worked in opposing manners to regulate heart field establishment, as the heat shock resulting in overexpression of FGF machinery in late gastrulation led to increased cardiac cell count, a phenotype also seen in DEAB-treated embryos [191,192]. Similarly, Aldh2 null mouse embryos bear a posteriorization and general domain increase of Fgf8 and its target Isl1 within the developing cardiac field, thus leading to improper chamber development [193,194].
RA signaling-associated machinery has also been a topic that has provided insights into RA and its role in heart development. One avenue of investigation included that of Cyp26 metabolic action for its role in maintaining RA equilibria. Opposite of a pan-RAR antagonist, Cyp26-deficient embryos result in a decreased heart field domain marked by nkx2.5, in course leading to decreased ventricular fates later in development [195]. Interestingly, the use of DEAB on these same cyp26 morphants results in a restoration of expression, concluding that loss of RA in early cardiac development results in increased heart field size, while increases in RA restrict these early progenitor populations and can alter later fate decisions [195]. These insights into RA’s role in cardiac development not only provide an understanding of base developmental processes, but also present a translational model for in utero VAD and accompanying congenital defects of the heart.
In work performed at later stages in development, Cyp26-deficient embryos were found to have increased mmp9 expression, leading to improper outflow tract morphogenesis due to the diminished addition from second heart field progenitors [196]. This work proposed a potential mechanism for RA-mediated outflow tract defects, and adds to the body of knowledge involving the relationship between vitamin A and mmp9. Further avenues of interrogation into processes associated with retinoid transport and synthesis within cardiac/cardiac progenitor cells remain open for insights, and make for exciting new lines of investigation.
One of the many reasons the zebrafish is such an attractive model is due to its regenerative capacity within the heart and other tissues [197]. In the ventricular amputation model [198], one hour after ventricular amputation of the adult zebrafish heart, aldh1a2 expression was remarkably abundant within the atrial endocardium, and within 3 days post amputation (dpa), expression of the RA-synthesizing enzyme was also localized to the epicardium, then to the epicardial cells that surround the wound clot by 7 dpa [199,200].
In this same line of experiments performed by Kikuchi et al., overexpression of cyp26a1, the enzyme responsible for RA degradation, led to dramatic decreases in cardiomyocyte proliferation at seven days post injury [200]. However, in the case of retinoid agonist supplementation, no increase in cardiomyocyte proliferation was induced, thus inferring that RA does not promote the regenerative process, but rather plays a permissive role [200]. Within the mouse model of acute damage to the heart, conflicting reports surround RA, and the protective effects it may or may not have in the case of myocardial infarction-induced cellular death [201,202].
3.4. Hematopoiesis
Research with the zebrafish has provided many fundamental insights into the genetic mechanisms of primitive and definitive hematopoiesis [203], from events involving hematopoietic stem cell (HSC) patterning [204] to fate choice [205,206] and differentiation [207,208,209,210]. A number of studies have contributed new insights into the effects of RA signaling on both the primitive and definitive hematopoietic waves, which are distinct in the cells they produce; namely, red blood cells and macrophages during the initial wave, followed by the production of all lineages in the subsequent wave, respectively [203].
Exogenous ATRA treatment of embryos from the late gastrula to the 5-somite stage elicits a dose-dependent inhibition of primitive erythropoiesis, based on reduced expression of the erythroid-specific transcription factor gata1 [211], and inhibition of primitive myelopoiesis, based on reduced expression of several markers [212]. Conversely, DEAB treatment increased specification of hematopoietic stem and progenitor cells, leading to elevated primitive erythropoiesis [213]. Further, interference in RA signaling with DEAB or though aldh1a2 knockdown has been associated with reductions in definitive blood cell production based on reduced expression of stem cell markers such as cmyb and thymic rag1/ikaros expression [214]. These findings parallel the ability of RA to enhance progenitor formation in culture [215], and the requirement for Aldh1a2 expression in the endothelium to produce definitive HSCs [216].
3.5. Pancreas, Liver, and Intestine
Within the endoderm, several organs/tissues have been characterized to have RA-dependent morphogenesis. Within the developing liver, RA positively regulates liver fates, as exogenous treatment of RA during varied significant points during hepatic differentiation led to increased expression of fabp1a (lfabp) at 72 hpf, while DEAB treatment at like times results in decreased fabp1a+ domain [217]. In this same line, fish treated with RA show increased levels of proliferation within the liver primordium via BrdU at 24 hpf [217]. Further investigation into the RA pathway by Garnaas et al. showcased rargb as being a regulator of early heart and liver morphogenesis, as rargb morphants and antagonists led to aberrant phenotypes in both tissues, falling in line with RAR agonist work involving cardiac cell proliferation [217,218]. In the developing intestines, decreases in RA biosynthesis, specifically in retinol dehydrogenase expression, were found to restrict gut marker expression domains in 96 hpf fish [219,220]. In these same experiments, interestingly, exogenous treatment of RA was found to only partially rescue the rdh1l morphants [220]. These results postulate that RA is a positive mediation of these two foregut tissues. These findings also suggest exciting avenues and niches for discovery within the realm of retinoid biology, especially with regard to downstream targets of R(A/X)Rs.
In the mouse model, aldh1a2 null animals were found to lack dorsal pancreatic tissues [221]. In fish, aldh1a2 null animals do not retain this same phenotype, as pdx1, a marker for beta cell progenitor populations was found to be expressed at 48 hpf, and other pancreatic markers were expressed at 72 hpf [63,222]. Further work utilizing both splice blocking and ATG-binding morpholinos, as well as DEAB, suggested that aldh1a2 is maternally deposited within fish due to DEAB and ATG-binding morphants resulting in the loss of pancreatic marker expression; meanwhile, aldh1a2 null and splice blocking MO-treated fish retained a domain, however aberrant [222]. During pancreatic development, loss of RA signaling via RAR pan-antagonist BMS493 before late gastrulation (90% epiboly) results in lack of insulin expression [223,224,225,226]. Conversely, ectopic ATRA (10−6 M treated from 9–10 hpf) results in the upregulation of insulin expression within 24 hpf animals, as well as increasing the insulin+ domain anteriorly, thus furthering the idea of RA-mediated beta cell proliferation across multiple models [223,224,225,226].
Recently, exciting new avenues thanks to multi-omic data have been revealed, and have opened for further investigation in RA/hepatopancreatic/endodermal biology fields. Insights into partially retained machinery across all three germ layers within the zebrafish were established. Among these observations, it was particularly interesting that hox clusters were established as being hosts to RAR sites, as this helps to confirm the idea of hox regulation of endodermal foregut tissues in the zebrafish model [227]. These same studies established RARs as mediating the expression of pioneer factors such as hnf1b and gata6 within the pancreas [227].
4. RA in Disease and Dysfunction
4.1. RA in Deficiency and Surplus in Humans
In addition to studying development, zebrafish have been a powerful system to study human disease (Figure 4) [18,19]. In work utilizing zebrafish, loss of RA associated with exposure to teratogenic compounds such as alcohol has been well documented. Zebrafish as a model for fetal alcohol syndrome has been attractive for the many reasons previously described [228,229]. In fish, when treated with 150 mM of ethanol from 3–24 hpf, severe phenotypes were observed within tissues and organs such as the eyes, otic vesicle, facial cartilage, and pericardial edema, among others [230]. These malformations fall in line with the severe defects seen clinically, as eye, heart, and improper craniofacial development are all found. These maladies are thought to be due to acetaldehyde, the byproduct of ethanol metabolism further being metabolized by an ALDH, thus reducing retinaldehydes’ ability to bind to ALDHs. In order to bypass this inhibition of RA synthesis, exogenous ATRA treatments at low levels (10−9 M) have been found to rescue the effects of ethanol exposure [230,231].
Much like insufficient RA is detrimental for fetal development in humans, an excess can be equally as harmful. Perhaps most notably, the dermatological drug isotretinoin, an RA isomer, is well characterized for the teratologic role it can have in pregnant individuals and their offspring. In utero exposure to the drug is estimated to have a 20–35% chance of congenital defects to many of the same tissues we have described as being RA-dependent in development. In terms of immeasurable maladies, it is estimated that potentially over 50% of individuals exposed to isotretinoin in utero may have cognitive impairment [233,234]. In zebrafish, isotretinoin has been used in neuroblastoma work, and has been successfully shown to have the capacity for decreasing tumor size in larval fish [235].
4.2. RA Pathways in Cancer
Within the realm of cancer biology, the zebrafish model has emerged as an efficient in vivo model, much for the same reasons why it is so popular in development. Thankfully, due to high amounts of conservation between the human and zebrafish genome, many oncologic markers are subsequently retained, making the fish useful for studying many aspects of cancer biology (Figure 4) [158,236,237]. The zebrafish immune system allows for tissue xenografts from mammalian donors, allowing for not only large-scale drug and genetic screens to take place, but also holding promise for a future in precision medicine [238,239,240,241].
In a zebrafish pancreatic cancer model, antagonization of RARs were used to downregulate miR-10a expression, resulting in a loss of invasive and metastatic phenotype; interestingly, it was also found that microRNA-10a (miR-10a) is an intermediate regulator between RARs and hox1 and hoxb3 [242]. This same work also showed that miR-10a is also upregulated in pancreatitis patient samples; this postulates the questions of whether RA is mediating the micro-RNA in both disease states, and if RA regulation of miR-10a is associated with normal development of the pancreas [242].
In neuroblastoma, loss of chromatin assembly factor 1 subunit p150 (CHAF1A) promotes oncogenesis/malignancy [243]. Alongside a mouse model, zebrafish were used to investigate chaf1a expression within regular neural crest cell development as well as carcinogenesis [244]. The use of ectopic chaf1a expression revealed that chaf1a plays roles in the critical fate determinant stages of neural crest cells in development. This work was in step with mouse work that was then used to hypothesize that RA could be used in combinatorial therapies for those diagnosed with neuroblastomas [244]. In the future, we believe zebrafish will further emerge as an in vivo model for oncological work as a supplement for early genetic and/or pharmacological screening methods, as well as for targeting therapeutics for individual patients [245].
5. Conclusions
Here, we have discussed how zebrafish research has been utilized to uncover new fundamental insights about the roles of RA signaling in the development of several tissues and organs, as well as in regeneration and disease. Continuing technological advances in multi-omics, combined with the tractability of zebrafish for pharmacological treatments, have facilitated these advances, and offer many prospects for future studies.
Our knowledge about the molecular targets of RA remains in its infancy, and more work is needed to decipher these gene regulatory networks, and to understand how the respective targets act to confer RA’s effects on cell fate specification and differentiation. Appreciating these facets may uncover new prospects for retinoids in clinical therapeutics. Indeed, the next century will be an exciting time in RA research, and the zebrafish will continue to proffer unique opportunities to narrow our knowledge gap about the many roles of this versatile and potent molecule in living systems.
Author Contributions
Conceptualization, M.R.H. and R.A.W.; writing—original draft preparation, M.R.H. and R.A.W.; writing—review and editing, M.R.H. and R.A.W.; visualization, M.R.H. and R.A.W.; supervision, R.A.W.; project administration, R.A.W.; funding acquisition, M.R.H. and R.A.W. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by funds from the University of Notre Dame College of Science (to R.A.W.) and a University of Notre Dame Graduate School Arthur J. Schmitt Leadership Fellowship (to M.R.H.). We are deeply grateful to the Gallagher family, most especially Elizabeth and Michael Gallagher, for their generous gift to the University of Notre Dame that enables the ongoing support for novel stem cell research. The funders had no role in the study design, data collection and analysis, the decision to publish, or manuscript preparation.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We thank the University of Notre Dame Department of Biological Sciences for their support. We express gratitude to the Freimann Life Science Center and the Center for Zebrafish Research at the University of Notre Dame for outstanding care of our zebrafish aquarium. R.A.W. thanks G.R.W., as well as B.C., C.G.M., K.P.E., and M.M. for unconditional support. Finally, we thank all the past and current members of our lab for making it a special place for the pursuit of science, and of course our zebrafish, for helping to unlock the mysteries of the world around us.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hale, F. The relation of vitamin A to anophthalmos in pigs. Am. J. Ophth. 1935, 18, 1087–1093. [Google Scholar] [CrossRef]
- Warkany, J.; Schraffenberger, E. Congenital malformations of the eyes induced in rats by maternal A deficiency. V. Effects of a purified diet lacking riboflavin. Proc. Soc. Exp. Biol. Med. 1943, 54, 92–94. [Google Scholar] [CrossRef]
- Warkany, J.; Schraffenberger, E. Congenital malformations of the eyes induced in rats by maternal A deficiency. Proc. Soc. Exp. Biol. Med. 1944, 57, 49–52. [Google Scholar] [CrossRef]
- Warkany, J.; Schraffenberger, E. Congenital malformations induced in rats by maternal vitamin A deficiency: I. Defects of the eye. Arrh. Ophthal. 1946, 35, 150–169. [Google Scholar] [CrossRef]
- Wilson, J.G.; Warkany, J. Epithelial keratinization as evidence of fetal vitamin A deficiency. Proc. Soc. Exp. Biol. Med. 1947, 64, 419–422. [Google Scholar] [CrossRef]
- Wilson, J.G.; Warkany, J. Abnormalities of the genitourinary tract induced by maternal A deficiency in fetal rats. Anat. Rec. 1947, 97, 396. [Google Scholar]
- Warkany, J.; Roth, C.B. Congenital malformations induced in rats by maternal vitamin A deficiency: II. Effect of varying the preparatory diet upon the yield of abnormal young. J. Nutr. 1948, 35, 1–11. [Google Scholar] [CrossRef]
- Wilson, J.G.; Warkany, J. Malformations in the genito-urinary tract induced by maternal vitamin A deficiency in the rat. Am. J. Anat. 1948, 83, 357–407. [Google Scholar] [CrossRef]
- Wilson, J.G.; Roth, C.B.; Warkany, J. An analysis of the syndrome of malformations induced by maternal vitamin A deficiency. Effects of restoration of vitamin A at various times during gestation. Am. J. Anat. 1953, 92, 189–217. [Google Scholar] [CrossRef]
- Clagett-Dame, M.; DeLuca, H.F. The role of vitamin A in mammalian reproduction and embryonic development. Annu. Rev. Nutr. 2002, 22, 347–381. [Google Scholar] [CrossRef]
- Niederreither, K.; Dolle, P. Retinoic acid in development: Towards an integrated view. Nat. Rev. Genet. 2008, 9, 541–553. [Google Scholar] [CrossRef]
- Duester, G. Retinoic acid synthesis and signaling during early organogenesis. Cell 2008, 134, 921–931. [Google Scholar] [CrossRef] [Green Version]
- Rhinn, M.; Dollé, P. Retinoic acid signalling during development. Development 2012, 139, 843–858. [Google Scholar] [CrossRef] [Green Version]
- Cunningham, T.J.; Duester, G. Mechanisms of retinoic acid signalling and its roles in organ and limb development. Nat. Rev. Mol. Cell. Biol. 2015, 16, 110–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghyselinck, N.B.; Duester, G. Retinoic acid signaling pathways. Development 2019, 146, dev167502. [Google Scholar] [CrossRef] [Green Version]
- Mezquita, B.; Mezquita, C. Two opposing faces of retinoic acid: Induction of stemness or induction of differentiation depending on cell-type. Biomolecules 2019, 9, 567. [Google Scholar] [CrossRef] [Green Version]
- Samarut, E.; Fraher, D.; Laudet, V.; Gibert, Y. ZebRA: An overview of retinoic acid signaling during zebrafish development. Biochim. Biophys. Acta 2015, 1849, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Lieschke, G.J.; Currie, P.D. Animal models of human disease: Zebrafish swim into view. Nat. Rev. Genet. 2007, 8, 353–367. [Google Scholar] [CrossRef] [PubMed]
- Patton, E.E.; Zon, L.I.; Langenau, D.M. Zebrafish disease models in drug discovery: From preclinical modelling to clinical trials. Nat. Rev. Drug Discov. 2021, 20, 611–628. [Google Scholar] [CrossRef]
- Albalat, R.; Cañestro, C. Identification of Aldh1a, Cyp26 and RAR orthologs in protostomes pushes back the retinoic acid genetic machinery in evolutionary time to the bilaterian ancestor. Chem. Biol. Interact. 2009, 178, 188–196. [Google Scholar] [CrossRef] [PubMed]
- Dehal, P.; Boore, J.L. Two rounds of whole genome duplication in the ancestral vertebrate. PLoS Biol. 2005, 3, e314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagatomo, K.; Ishibashi, T.; Satou, Y.; Satoh, N.; Fujiwara, S. Retinoic acid affects gene expression and morphogenesis without upregulating the retinoic acid receptor in the ascidian Ciona intestinalis. Mech. Dev. 2003, 120, 363–372. [Google Scholar] [CrossRef] [PubMed]
- Nagatomo, K.; Fujiwara, S. Expression of Raldh2, Cyp26 and Hox-1 in normal and retinoic acid-treated Ciona intestinalis embryos. Gene Expr. Patterns 2003, 3, 273–277. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Zhang, J.; Chen, S.; Wang, F.; Zhang, T.; Niswander, L. Genetic contribution of retinoid-related genes to neural tube defects. Hum. Mutat. 2018, 39, 550–562. [Google Scholar] [CrossRef]
- Burke, A.C.; Nelson, C.E.; Morgan, B.A.; Tabin, C. Hox genes and the evolution of vertebrate axial morphology. Development 1995, 121, 333–346. [Google Scholar] [CrossRef]
- Ishihara, K.; Nakamoto, M.; Nakao, M. DNA methylation-independent removable insulator controls chromatin remodeling at the HOXA locus via retinoic acid signaling. Hum. Mol. Genet. 2016, 25, 5383–5394. [Google Scholar] [PubMed] [Green Version]
- Yamada, K.; Maeno, A.; Araki, S.; Kikuchi, M.; Suzuki, M.; Ishizaka, M.; Satoh, K.; Akama, K.; Kawabe, Y.; Suzuki, K.; et al. An atlas of seven zebrafish hox cluster mutants provides insights into sub/neofunctionalization of vertebrate Hox clusters. Development 2021, 148, dev198325. [Google Scholar] [CrossRef]
- Mallo, M.; Wellik, D.M.; Deschamps, J. Hox genes and regional patterning of the vertebrate body plan. Dev. Biol. 2010, 344, 7–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abbasi, A.A. Diversification of four human HOX gene clusters by step-wise evolution rather than ancient whole-genome duplications. Dev. Genes Evol. 2015, 225, 353–357. [Google Scholar] [CrossRef] [PubMed]
- Asrar, Z.; Haq, F.; Abbasi, A.A. Fourfold paralogy regions on human HOX-bearing chromosomes: Role of ancient segmental duplications in the evolution of vertebrate genome. Mol. Phylogenet. Evol. 2013, 66, 737–747. [Google Scholar] [CrossRef] [PubMed]
- Hoegg, S.; Meyer, A. Hox clusters as models for vertebrate genome evolution. Trends Genet. 2005, 21, 421–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amores, A.; Force, A.; Yan, Y.L.; Joly, L.; Amemiya, C.; Fritz, A.; Ho, R.K.; Langeland, J.; Prince, V.; Wang, Y.L.; et al. Zebrafish hox clusters and vertebrate genome evolution. Science 1998, 282, 1711–1714. [Google Scholar] [CrossRef] [PubMed]
- Stellwag, E.J. Hox gene duplication in fish. Semin. Cell Dev. Biol. 1999, 10, 531–540. [Google Scholar] [CrossRef]
- Kuraku, S.; Meyer, A. The evolution and maintenance of Hox gene clusters in vertebrates and the teleost-specific genome duplication. Int. J. Dev. Biol. 2009, 53, 765–773. [Google Scholar] [CrossRef] [Green Version]
- Graham, A.; Papalopulu, N.; Krumlauf, R. The murine and Drosophila homeobox gene complexes have common features of organization and expression. Cell 1989, 57, 367–378. [Google Scholar] [CrossRef]
- Dekker, E.J.; Pannese, M.; Houtzager, E.; Timmermans, A.; Boncinelli, E.; Durston, A. Xenopus Hox-2 genes are expressed sequentially after the onset of gastrulation and are differentially inducible by retinoic acid. Dev. Suppl. 1992, 116, 195–202. [Google Scholar] [CrossRef]
- Conlon, R.A.; Rossant, J. Exogenous retinoic acid rapidly induces anterior ectopic expression of murine Hox-2 genes in vivo. Development 1992, 116, 357–368. [Google Scholar] [CrossRef] [PubMed]
- Dekker, E.J.; Pannese, M.; Houtzager, E.; Boncinelli, E.; Durston, A. Colinearity in the Xenopus laevis Hox-2 complex. Mech. Dev. 1993, 40, 3–12. [Google Scholar] [CrossRef]
- del Corral, R.D.; Olivera-Martinez, I.; Goriely, A.; Gale, E.; Maden, M.; Storey, K. Opposing FGF and retinoid pathways control ventral neural pattern, neuronal differentiation, and segmentation during body axis extension. Neuron 2003, 40, 65–79. [Google Scholar] [CrossRef] [Green Version]
- Kudoh, T.; Wilson, S.W.; Dawid, I.B. Distinct roles for Fgf, Wnt and retinoic acid in posteriorizing the neural ectoderm. Development 2002, 129, 4335–4346. [Google Scholar] [CrossRef]
- Thaller, C.; Eichele, G. Identification and spatial distribution of retinoids in the developing chick limb bud. Nature 1987, 327, 625–628. [Google Scholar] [CrossRef] [PubMed]
- Durston, A.J.; Timmermans, J.P.M.; Hage, W.J.; Hendricks, H.F.J.; de Vries, N.J.; Heideveld, M.; Nieuwkoop, P.D. Retinoic acid causes an anteroposterior transformation in the developing nervous system. Nature 1989, 340, 140–144. [Google Scholar] [CrossRef] [PubMed]
- Horton, C.; Maden, M. Endogenous distribution of retinoids during normal development and teratogenesis in the mouse embryo. Dev. Dyn. 1995, 202, 312–323. [Google Scholar] [CrossRef] [PubMed]
- Costaridis, P.; Horton, C.; Zeitlinger, J.; Holder, N.; Maden, M. Endogenous retinoids in the zebrafish embryo and adult. Dev. Dyn. 1996, 205, 41–51. [Google Scholar] [CrossRef]
- Ulven, S.M.; Gundersen, T.E.; Weedon, M.S.; Landaas, V.O.; Sakhi, A.K.; Fromm, S.H.; Geronimo, B.A.; Moskaug, J.O.; Blomhoff, R. Identification of endogenous retinoids, enzymes, binding proteins, and receptors during early postimplantation development in mouse: Important role of retinal dehydrogenase type 2 in synthesis of all-trans-retinoic acid. Dev. Biol. 2000, 220, 379–391. [Google Scholar] [CrossRef] [Green Version]
- Lampert, J.M.; Holzschuh, J.; Hessel, S.; Driever, W.; Vogt, K.; von Lintig, J. Provitamin A conversion to retinal via the β,β-carotene-15,15’-oxygenase (bcox) is essential for pattern formation and differentiation during zebrafish embryogenesis. Development 2003, 130, 2173–2186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawaguchi, R.; Yu, J.; Honda, J.; Hu, J.; Whitelegge, J.; Ping, P.; Wiita, P.; Bok, D.; Sun, H. A membrane receptor for retinol binding protein mediates cellular uptake of vitamin A. Science 2007, 315, 820–825. [Google Scholar] [CrossRef]
- Sun, H. Membrane receptors and transporters involved in the function and transport of vitamin A and its derivatives. Biochim. Biophys. Acta 2012, 1821, 99–112. [Google Scholar] [CrossRef] [Green Version]
- Alapatt, P.; Guo, F.; Komanetsky, S.M.; Wang, S.; Cai, J.; Sargsyan, A.; Rodríguez Díaz, E.; Bacon, B.T.; Aryal, P.; Graham, T.E. Liver retinol transporter and receptor for serum retinol-binding protein (RBP4). J. Biol. Chem. 2013, 288, 1250–1265. [Google Scholar] [CrossRef] [Green Version]
- Shi, Y.; Obert, E.; Rahman, B.; Rohrer, B.; Lobo, G.P. The retinol binding protein receptor 2 (Rbpr2) is required for photoreceptor outer segment morphogenesis and visual function in zebrafish. Sci. Rep. 2017, 7, 16207. [Google Scholar] [CrossRef] [Green Version]
- Napoli, J.L. Functions of intracellular retinoid binding-proteins. Subcell. Biochem. 2016, 81, 21–76. [Google Scholar]
- Napoli, J.L.; Yoo, H.S. Retinoid metabolism and functions mediated by retinoid binding-proteins. Methods Enzymol. 2020, 637, 55–75. [Google Scholar]
- Isken, A.; Holzschuh, J.; Lampert, J.M.; Fischer, L.; Oberhauser, V.; Palczewski, K.; von Lintig, J. Sequestration of retinyl esters is essential for retinoid signaling in the zebrafish embryo. J. Biol. Chem. 2007, 282, 1144–1151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parés, X.; Farrés, J.; Kedishvili, N.; Duester, G. Medium- and short-chain dehydrogenase/reductase gene and protein families: Medium-chain and short-chain dehydrogenases/reductases in retinoid metabolism. Cell Mol. Life Sci. 2008, 65, 3936–3949. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Sandell, L.L.; Trainor, P.A.; Koentgen, F.; Duester, G. Alcohol and aldehyde dehydrogenases: Retinoid metabolic effects in mouse knockout models. Biochim. Biophys. Acta 2012, 1821, 198–205. [Google Scholar] [CrossRef] [Green Version]
- Kedishvili, N.Y. Retinoic acid synthesis and degradation. Subcell. Biochem. 2016, 81, 127–161. [Google Scholar]
- Pittlik, S.; Domingues, S.; Meyer, A.; Begemann, G. Expression of zebrafish aldh1a3 (raldh3) and absence of aldh1a1 in teleosts. Gene Expr. Patterns 2008, 8, 141–147. [Google Scholar] [CrossRef] [Green Version]
- Yue, H.; Hu, Z.; Hu, R.; Guo, Z.; Zheng, Y.; Wang, Y.; Zhou, Y. ALDH1A1 in cancers: Bidirectional function, drug resistance, and regulatory mechanism. Front. Oncol. 2022, 12, 918778. [Google Scholar] [CrossRef]
- Begemann, G.; Schilling, T.F.; Rauch, G.J.; Geisler, R.; Ingham, P.W. The zebrafish neckless mutation reveals a requirement for raldh2 in mesodermal signals that pattern the hindbrain. Development 2001, 128, 3081–3094. [Google Scholar] [CrossRef] [PubMed]
- Begemann, G.; Marx, M.; Mebus, K.; Meyer, A.; Bastmeyer, M. Beyond the neckless phenotype: Influence of reduced retinoic acid signaling on motor neuron development in the zebrafish hindbrain. Dev. Biol. 2001, 271, 119–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linville, A.; Gumusaneli, E.; Chandraratna, R.A.; Schilling, T.F. Independent roles for retinoic acid in segmentation and neuronal differentiation in the zebrafish hindbrain. Dev. Biol. 2004, 270, 186–199. [Google Scholar] [CrossRef] [PubMed]
- Grandel, H.; Lun, K.; Rauch, G.J.; Rhinn, M.; Piotrowski, T.; Houart, C.; Sordino, P.; Küchler, A.M.; Schulte-Merker, S.; Geisler, R.; et al. Retinoic acid signaling in the zebrafish embryo is necessary during pre-segmentation stages to pattern the anterior-posterior axis of the CNS and to induce a pectoral limb bud. Development 2002, 129, 2851–2865. [Google Scholar] [CrossRef]
- Alexa, K.; Choe, S.K.; Hirsch, N.; Etheridge, L.; Laver, E.; Sagerstrom, C.G. Maternal and zygotic aldh1a2 activity is required for pancreas development in zebrafish. PLoS ONE 2009, 4, e8261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wingert, R.A.; Davidson, A.J. Zebrafish nephrogenesis involves dynamic spatiotemporal expression changes in renal progenitors and essential signals from retinoic acid and irx3b. Dev. Dyn. 2011, 240, 2011–2027. [Google Scholar] [CrossRef] [Green Version]
- White, J.C.; Highland, M.; Kaiser, M.; Clagett-Dame, M. Vitamin A deficiency results in the dose-dependent acquisition of anterior character and shortening of the caudal hindbrain of the rat embryo. Dev. Biol. 2000, 220, 263–284. [Google Scholar] [CrossRef] [PubMed]
- Yahyavi, M.; Abouzeid, H.; Gawdat, G.; de Preux, A.S.; Xiao, T.; Bardakjian, T.; Schneider, A.; Choi, A.; Jorgenson, E.; Baier, H.; et al. ALDH1A3 loss of function causes bilateral anophthalmia/ microphthalmia and hypoplasia of the optic nerve and optic chiasm. Hum. Mol. Genet. 2013, 22, 3250–3258. [Google Scholar] [CrossRef] [Green Version]
- Fares-Taie, L.; Gerber, S.; Chassaing, N.; Clayton-Smith, J.; Hanein, S.; Silva, E.; Serey, M.; Serre, V.; Gérard, X.; Baumann, C.; et al. ALDH1A3 mutations cause recessive anophthalmia and microphthalmia. Am. J. Hum. Genet. 2013, 92, 265–270. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.; Harlalka, G.V.; Hameed, A.; Reham, H.M.; Yasin, M.; Muhammad, N.; Khan, S.; Baple, E.L.; Crosby, A.H.; Saleha, S. Novel mutations in ALDH1A3 associated with autosomal recessive anophthalmia/microphthalmia, and review of the literature. BMC Med. Genet. 2018, 19, 160. [Google Scholar] [CrossRef] [Green Version]
- Liang, D.; Zhang, M.; Bao, J.; Zhang, L.; Xu, X.; Gao, X.; Zhao, Q. Expressions of Raldh3 and Raldh4 during zebrafish early development. Gene Expr. Patterns 2008, 8, 248–253. [Google Scholar] [CrossRef]
- Pittlik, S.; Begemann, G. New sources of retinoic acid synthesis revealed by live imaging of an Aldh1a2-GFP reporter fusion protein throughout zebrafish development. Dev. Dyn. 2012, 241, 1205–1216. [Google Scholar] [CrossRef]
- Xi, J.; Yue, J.; Yang, Z. Expression profiles of retinoic acid synthetases ALDH1As and metabolic enzymes CYP26s in adult and embryonic zebrafish (Danio rerio). Genet. Mol. Res. 2015, 14, 3948–3956. [Google Scholar] [CrossRef]
- Lan, C.C.; Tang, R.; Un San Leong, I.; Love, D.R. Quantitative real-time RT-PCR (qRT-PCR) of zebrafish transcripts: Optimization of RNA extraction, quality control considerations, and data analysis. Cold Spring Harb. Protoc. 2009, 2009, pdb.prot5314. [Google Scholar] [CrossRef]
- Vanhauwaert, S.; Lefever, S.; Coucke, P.; Speleman, F.; De Paepe, A.; Vandesompele, J.; Willaert, A. RT-qPCR gene expression analysis in zebrafish: Preanalytical precautions and use of expressed repetitive elements for normalization. Methods Cell Biol. 2016, 135, 329–342. [Google Scholar] [PubMed]
- Cheng, C.N.; Li, Y.; Marra, A.N.; Verdun, V.; Wingert, R.A. Flat mount preparation for observation and analysis of zebrafish embryo specimens stained by whole mount in situ hybridization. J. Vis. Exp. 2014, 89, 51604. [Google Scholar]
- Marra, A.N.; Ulrich, M.; White, A.; Springer, M.; Wingert, R.A. Visualizing multiciliated cells in the zebrafish through a combined protocol of whole mount fluorescent in situ hybridization and immunofluorescence. J. Vis. Exp. 2017, 129, 56261. [Google Scholar]
- Marra, A.N.; Chambers, B.E.; Chambers, J.M.; Drummond, B.E.; Adeeb, B.D.; Wesselman, H.M.; Morales, E.E.; Handa, N.; Pettini, T.; Ronshaugen, M.; et al. Visualizing gene expression during zebrafish pronephros development and regeneration. Methods Cell Biol. 2019, 154, 183–215. [Google Scholar]
- Wesselman, H.M.; Gatz, A.; Wingert, R.A. Visualizing multiciliated cells in the zebrafish. Methods Cell Biol. 2023, 175, 129–161. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.N. Cellular retinoic acid binding proteins: Genomic and non-genomic functions and their regulation. Subcell. Biochem. 2016, 81, 163–178. [Google Scholar] [PubMed]
- Napoli, J.L. Cellular retinoid binding-proteins, CRBP, CRABP, FABP5: Effects on retinoid metabolism, function and related diseases. Pharmacol. Ther. 2017, 173, 19–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dollé, P. Developmental expression of retinoic acid receptors (RARs). Nucl. Recept. Signal. 2009, 7, e006. [Google Scholar] [CrossRef] [Green Version]
- Petkovich, M.; Chambon, P. Retinoic acid receptors at 35 years. J. Mol. Endocrinol. 2022, 69, T13–T24. [Google Scholar] [CrossRef] [PubMed]
- Rochette-Egly, C.; Germain, P. Dynamic and combinatorial control of gene expression by nuclear retinoic acid receptors (RARs). Nucl. Recept. Signal. 2009, 7, e005. [Google Scholar] [CrossRef] [PubMed]
- Samarut, E.; Rochette-Egly, C. Nuclear retinoic acid receptors: Conductors of the retinoic acid symphony during development. Mol. Cell Endocrinol. 2012, 348, 348–360. [Google Scholar] [CrossRef] [PubMed]
- Rochette-Egly, C. Retinoic Acid-Regulated Target Genes During Development: Integrative Genomics Analysis. Subcell. Biochem. 2020, 95, 57–85. [Google Scholar]
- Petkovich, M.; Brand, N.J.; Krust, A.; Chambon, P. A human retinoic acid receptor which belongs to the family of nuclear receptors. Nature 1987, 330, 444–450. [Google Scholar] [CrossRef]
- Giguere, V.; Ong, E.S.; Segui, P.; Evans, R.M. Identification of a receptor for the morphogen retinoic acid. Nature 1987, 330, 624–629. [Google Scholar] [CrossRef]
- Brand, N.; Petkovich, M.; Krust, A.; Chambon, P.; de Thé, H.; Marchio, A.; Tiollais, P.; Dejean, A. Identification of a second human retinoic acid receptor. Nature 1988, 332, 850–853. [Google Scholar] [CrossRef]
- Desreumaux, P.; Dubuquoy, L.; Nutten, S.; Peuchmaur, M.; Englaro, W.; Schoonjans, K.; Derijard, B.; Desvergne, B.; Wahli, W.; Chambon, P.; et al. Attenuation of colon inflammation through activators of the retinoid X receptor (RXR)/peroxisome proliferator-activated receptor gamma (PPARgamma) heterodimer. A basis for new therapeutic strategies. J. Exp. Med. 2001, 193, 827–838. [Google Scholar] [CrossRef] [Green Version]
- Brtko, J.; Dvorak, Z. Natural and synthetic retinoid X receptor ligands and their role in selected nuclear receptor action. Biochimie 2020, 179, 157–168. [Google Scholar] [CrossRef]
- De Bosscher, K.; Desmet, S.J.; Clarisse, D.; Estébanez-Perpiña, E.; Brunsveld, L. Nuclear receptor crosstalk—Defining the mechanisms for therapeutic innovation. Nat. Rev. Endocrinol. 2020, 16, 363–377. [Google Scholar] [CrossRef]
- Chambers, J.M.; Wingert, R.A. PGC-1α in disease: Recent renal insights into a versatile metabolic regulator. Cells 2020, 9, 2234. [Google Scholar] [CrossRef] [PubMed]
- Hale, L.A.; Tallafuss, A.; Yan, Y.L.; Dudley, L.; Eisen, J.S.; Postlethwait, J.H. Characterization of the retinoic acid receptor genes raraa, rarab and rarg during zebrafish development. Gene Expr. Patterns 2006, 6, 546–555. [Google Scholar] [CrossRef] [PubMed]
- Tallafuss, A.; Hale, L.A.; Yan, Y.L.; Dudley, L.; Eisen, J.S.; Postlethwait, J.H. Characterization of retinoid-X receptor genes rxra, rxrba, rxrbb and rxrg during zebrafish development. Gene Expr. Patterns 2006, 6, 556–565. [Google Scholar] [CrossRef]
- Waxman, J.S.; Yelon, D. Comparison of the expression patterns of newly identified zebrafish retinoic acid and retinoid X receptors. Dev. Dyn. 2007, 236, 587–595. [Google Scholar] [CrossRef] [PubMed]
- Linville, A.; Radtke, K.; Waxman, J.S.; Yelon, D.; Schilling, T.F. Combinatorial roles for zebrafish retinoic acid receptors in the hindbrain, limbs and pharyngeal arches. Dev. Biol. 2009, 325, 60–70. [Google Scholar] [CrossRef] [Green Version]
- Waxman, J.S.; Yelon, D. Zebrafish retinoic acid receptors function as context-dependent transcriptional activators. Dev. Biol. 2011, 352, 128–140. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, E.; Casado, M.; Raldúa, D.; Soares, A.; Barata, C.; Piña, B. Retinoic acid receptors’ expression and function during zebrafish early development. J. Steroid Biochem. Mol. Biol. 2013, 138, 143–151. [Google Scholar] [CrossRef] [PubMed]
- D’Aniello, E.; Rydeen, A.B.; Anderson, J.L.; Mandal, A.; Waxman, J.S. Depletion of retinoic acid receptors initiates a novel positive feedback mechanism that promotes teratogenic increases in retinoic acid. PLoS Genet. 2013, 9, e1003689. [Google Scholar] [CrossRef] [Green Version]
- Samarut, E.; Gaudin, C.; Hughes, S.; Gillet, B.; de Bernard, S.; Jouve, P.E.; Buffat, L.; Allot, A.; Lecompte, O.; Berekelya, L.; et al. Retinoic acid receptor subtype-specific transcriptotypes in the early zebrafish embryo. Mol. Endocrinol. 2014, 28, 260–272. [Google Scholar] [CrossRef] [Green Version]
- White, J.A.; Guo, Y.D.; Baetz, K.; Beckett-Jones, B.; Bonasoro, J.; Hsu, K.E.; Dilworth, F.J.; Jones, G.; Petkovich, M. Identification of the retinoic acid-inducible all-trans-retinoic acid 4-hydroxylase. J. Biol. Chem. 1996, 271, 29922–29927. [Google Scholar] [CrossRef] [Green Version]
- Nelson, D.R. A second CYP26 P450 in humans and zebrafish: CYP26B1. Arch. Biochem. Biophys. 1999, 371, 345–347. [Google Scholar] [CrossRef]
- MacLean, G.; Abu-Abed, S.; Dollé, P.; Tahayato, A.; Chambon, P.; Petkovich, M. Cloning of a novel retinoic-acid metabolizing cytochrome P450, Cyp26B1, and comparative expression analysis with Cyp26A1 during early murine development. Mech. Dev. 2001, 107, 195–201. [Google Scholar] [CrossRef]
- Tahayato, A.; Dollé, P.; Petkovich, M. Cyp26C1 encodes a novel retinoic acid-metabolizing enzyme expressed in the hindbrain, inner ear, first branchial arch and tooth buds during murine development. Gene Expr. Patterns 2003, 3, 449–454. [Google Scholar] [CrossRef] [PubMed]
- Taimi, M.; Helvig, C.; Wisniewski, J.; Ramshaw, H.; White, J.; Amad, M.; Korczak, B.; Petkovich, M. A novel human cytochrome P450, CYP26C1, involved in metabolism of 9-cis and all-trans isomers of retinoic acid. J. Biol. Chem. 2004, 279, 77–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Q.; Dobbs-McAuliffe, B.; Linney, E. Expression of cyp26b1 during zebrafish early development. Gene Expr. Patterns 2005, 5, 363–369. [Google Scholar] [CrossRef] [PubMed]
- Gu, X.; Xu, F.; Wang, X.; Gao, X.; Zhao, Q. Molecular cloning and expression of a novel CYP26 gene (cyp26d1) during zebrafish early development. Gene Expr. Patterns 2005, 5, 733–739. [Google Scholar] [CrossRef]
- Gu, X.; Xu, F.; Song, W.; Wang, X.; Hu, P.; Yang, Y.; Gao, X.; Zhao, Q. A novel cytochrome P450, zebrafish Cyp26D1, is involved in metabolism of all-trans retinoic acid. Mol. Endocrinol. 2006, 20, 1661–1672. [Google Scholar] [CrossRef] [Green Version]
- Emoto, Y.; Wada, H.; Okamoto, H.; Kudo, A.; Imai, Y. Retinoic acid-metabolizing enzyme Cyp26a1 is essential for determining territories of hindbrain and spinal cord in zebrafish. Dev. Biol. 2005, 278, 415–427. [Google Scholar] [CrossRef] [Green Version]
- Pennimpede, T.; Cameron, D.A.; MacLean, G.A.; Li, H.; Abu-Abed, S.; Petkovich, M. The role of CYP26 enzymes in defining appropriate retinoic acid exposure during embryogenesis. Birth Defects Res. A Clin. Mol. Teratol. 2010, 88, 883–894. [Google Scholar] [CrossRef]
- Ross, A.C.; Zolfaghari, R. Cytochrome P450s in the regulation of cellular retinoic acid metabolism. Annu. Rev. Nutr. 2011, 31, 65–87. [Google Scholar] [CrossRef] [Green Version]
- Dobbs-McAuliffe, B.; Zhao, Q.; Linney, E. Feedback mechanisms regulate retinoic acid production and degradation in the zebrafish embryo. Mech. Dev. 2004, 121, 339–350. [Google Scholar] [CrossRef]
- Rydeen, A.; Voisin, N.; D’Aniello, E.; Ravisankar, P.; Devignes, C.S.; Waxman, J.S. Excessive feedback of Cyp26a1 promotes cell non-autonomous loss of retinoic acid signaling. Dev. Biol. 2015, 405, 47–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-Marí, A.; Cañestro, C.; BreMiller, R.A.; Catchen, J.M.; Yan, Y.L.; Postlethwait, J.H. Retinoic acid metabolic genes, meiosis, and gonadal sex differentiation in zebrafish. PLoS ONE 2013, 8, e73951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skromne, I.; Prince, V.E. Current perspectives in zebrafish reverse genetics: Moving forward. Dev. Dyn. 2008, 237, 861–882. [Google Scholar] [CrossRef] [Green Version]
- Bernheim, S.; Meilhac, S.M. Mesoderm patterning by a dynamic gradient of retinoic acid signaling. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2020, 375, 20190556. [Google Scholar] [CrossRef] [PubMed]
- Loudig, O.; Babichuk, C.; White, J.; Abu-Abed, S.; Mueller, C.; Petkovich, M. Cytochrome P450RAI(CYP26) promoter: A distinct composite retinoic acid response element underlies the complex regulation of retinoic acid metabolism. Mol. Endocrinol. 2000, 14, 1483–1497. [Google Scholar] [CrossRef]
- Loudig, O.; Maclean, G.A.; Dore, N.L.; Luu, L.; Petkovich, M. Transcriptional co-operativity between distant retinoic acid response elements in regulation of Cyp26A1 inducibility. Biochem. J. 2005, 392, 241–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernandez, R.E.; Putzke, A.P.; Myers, J.P.; Margaretha, L.; Moens, C.B. Cyp26 enzymes generate the retinoic acid response pattern necessary for hindbrain development. Development 2007, 134, 177–187. [Google Scholar]
- Hu, P.; Tian, M.; Bao, J.; Xing, G.; Gu, X.; Gao, X.; Linney, E.; Zhao, Q. Retinoid regulation of the zebrafish cyp26a1 promoter. Dev. Dyn. 2008, 237, 3798–3808. [Google Scholar]
- White, R.J.; Nie, Q.; Lander, A.D.; Schilling, T.F. Complex regulation of cyp26a1 creates a robust retinoic acid gradient in the zebrafish embryo. PLoS Biol. 2007, 5, e304. [Google Scholar] [CrossRef]
- Shimozono, S.; Iimura, T.; Kitaguchi, T.; Higashijima, S.; Miyawaki, A. Visualization of an endogenous retinoic acid gradient across embryonic development. Nature 2013, 496, 363–366. [Google Scholar] [CrossRef] [PubMed]
- Mey, J.; McCaffery, P.; Klemeit, M. Sources and sink of retinoic acid in the embryonic chick retina: Distribution of aldehyde dehydrogenase activities, CRABP-I, and sites of retinoic acid inactivation. Brain Res. Dev. Brain Res. 2001, 127, 135–148. [Google Scholar] [CrossRef] [PubMed]
- Aulehla, A.; Pourquié, O. Signaling gradients during paraxial mesoderm development. Cold Spring Harb. Perspect. Biol. 2010, 2, a000869. [Google Scholar] [CrossRef] [Green Version]
- Schilling, T.F.; Nie, Q.; Lander, A.D. Dynamics and precision in retinoic acid morphogen gradients. Curr. Opin. Genet. Dev. 2012, 22, 562–569. [Google Scholar] [CrossRef] [Green Version]
- Dubey, A.; Rose, R.E.; Jones, D.R.; Saint-Jeannet, J.P. Generating retinoic acid gradients by local degradation during craniofacial development: One cell’s cue is another cell’s poison. Genesis 2018, 56, e23091. [Google Scholar] [CrossRef]
- Simsek, M.F.; Özbudak, E.M. Patterning principles of morphogen gradients. Open. Biol. 2022, 12, 220224. [Google Scholar] [CrossRef] [PubMed]
- Gudas, L.J. Retinoid metabolism: New insights. J. Mol. Endocrinol. 2022, 69, T37–T49. [Google Scholar] [CrossRef] [PubMed]
- Irion, U.; Nüsslein-Volhard, C. Developmental genetics with model organisms. Proc. Natl. Acad. Sci. USA 2022, 119, e2122148119. [Google Scholar] [CrossRef]
- Shin, J.T.; Fishman, M.C. From zebrafish to human: Modular medical models. Annu. Rev. Genomics Hum. Genet. 2002, 3, 311–340. [Google Scholar] [CrossRef]
- Kimmel, C.B.; Ballard, W.W.; Kimmel, S.R.; Ullmann, B.; Schilling, T.F. Stages of embryonic development of the zebrafish. Dev. Dyn. 1995, 203, 253–310. [Google Scholar] [CrossRef] [PubMed]
- Hogan, B.M.; Verkade, H.; Lieschke, G.J.; Heath, J.K. Manipulation of gene expression during zebrafish embryonic development using transient approaches. Methods Mol. Biol. 2008, 469, 273–300. [Google Scholar]
- Lawson, N.D.; Wolfe, S.A. Forward and reverse genetic approaches for the analysis of vertebrate development in the zebrafish. Dev. Cell 2011, 21, 48–64. [Google Scholar] [CrossRef] [Green Version]
- Gerlach, G.F.; Morales, E.E.; Wingert, R.A. Microbead implantation in the zebrafish embryo. J. Vis. Exp. 2015, 101, e52943. [Google Scholar]
- Haffter, P.; Granato, M.; Brand, M.; Mullins, M.C.; Hammerschmidt, M.; Kane, D.A.; Odenthal, J.; van Eeden, F.J.; Jiang, Y.J.; Heisenberg, C.P.; et al. The identification of genes with unique and essential functions in the development of the zebrafish, Danio rerio. Development 1996, 123, 1–36. [Google Scholar] [CrossRef] [PubMed]
- Driever, W.; Solnica-Krezel, L.; Schier, A.F.; Neuhauss, S.C.; Malicki, J.; Stemple, D.L.; Stainier, D.Y.; Zwartkruis, F.; Abdelilah, S.; Rangini, Z.; et al. A genetic screen for mutations affecting embryogenesis in zebrafish. Development 1996, 123, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Patton, E.E.; Zon, L.I. The art and design of genetic screens: Zebrafish. Nat. Rev. Genet. 2001, 2, 956–966. [Google Scholar] [CrossRef]
- Golling, G.; Amsterdam, A.; Sun, Z.; Antonelli, M.; Maldonado, E.; Chen, W.; Burgess, S.; Haldi, M.; Artzt, K.; Farrington, S.; et al. Insertional mutagenesis in zebrafish rapidly identifies genes essential for early vertebrate development. Nat. Genet. 2002, 31, 135–140. [Google Scholar] [CrossRef]
- Sun, Z.; Amsterdam, A.; Pazour, G.J.; Cole, D.G.; Miller, M.S.; Hopkins, N. A genetic screen in zebrafish identifies cilia genes as a principal cause of cystic kidney. Development 2004, 131, 4085–4093. [Google Scholar] [CrossRef] [Green Version]
- Kroeger, P.T.; Poureetezadi, S.J.; McKee, R.; Jou, J.; Miceli, R.; Wingert, R.A. Production of haploid zebrafish embryos by in vitro fertilization. J. Vis. Exp. 2014, 89, 51708. [Google Scholar]
- Ebarasi, L.; Ashraf, S.; Bierzynska, A.; Gee, H.Y.; McCarthy, H.J.; Lovric, S.; Sadowski, C.E.; Pabst, W.; Vega-Warner, V.; Fang, H.; et al. Defects of CRB2 cause steroid-resistant nephrotic syndrome. Am. J. Hum. Genet. 2015, 96, 153–161. [Google Scholar] [CrossRef] [Green Version]
- Kroeger, P.T.; Drummond, B.E.; Miceli, R.; McKernan, M.; Gerlach, G.F.; Marra, A.N.; Fox, A.; McCampbell, K.K.; Leshchiner, I.; Rodriguez-Mari, A.; et al. The zebrafish kidney mutant zeppelin reveals that brca2/fancd1 is essential for pronephros development. Dev. Biol. 2017, 428, 148–163. [Google Scholar] [CrossRef] [PubMed]
- Gehrig, J.; Pandey, G.; Westhoff, J.H. Zebrafish as a model for drug screening in genetic kidney diseases. Front. Pediatr. 2018, 6, 183. [Google Scholar] [CrossRef]
- Chambers, B.E.; Gerlach, G.F.; Clark, E.G.; Chen, K.H.; Levesque, A.E.; Leshchiner, I.; Goessling, W.; Wingert, R.A. Tfap2a is a novel gatekeeper of nephron differentiation during kidney development. Development 2019, 146, dev172387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drummond, B.E.; Chambers, B.E.; Wesselman, H.M.; Gibson, S.; Arceri, L.; Ulrich, M.N.; Gerlach, G.F.; Kroeger, P.T.; Leshchiner, I.; Goessling, W.; et al. osr1 maintains renal progenitors and regulates podocyte development by promoting wnt2ba via the antagonism of hand2. Biomedicines 2022, 10, 2868. [Google Scholar] [CrossRef] [PubMed]
- Peterson, R.T.; Link, B.A.; Dowling, J.E.; Schreiber, S.L. Small molecule developmental screens reveal the logic and timing of vertebrate development. Proc. Natl. Acad. Sci. USA 2000, 97, 12965–12969. [Google Scholar] [CrossRef] [Green Version]
- Rennekamp, A.J.; Peterson, R.T. 15 years of zebrafish chemical screening. Curr. Opin. Chem. Biol. 2015, 24, 58–70. [Google Scholar] [CrossRef] [Green Version]
- Poureetezadi, S.J.; Donahue, E.K.; Wingert, R.A. A manual small molecule screen approaching high-throughput using zebrafish embryos. J. Vis. Exp. 2014, 93, e52063. [Google Scholar]
- Poureetezadi, S.J.; Cheng, C.N.; Chambers, J.M.; Drummond, B.E.; Wingert, R.A. Prostaglandin signaling regulates nephron segment patterning of renal progenitors during zebrafish kidney development. Elife 2016, 5, e17551. [Google Scholar] [CrossRef]
- Chambers, J.M.; Poureetezadi, S.J.; Addiego, A.; Lahne, M.; Wingert, R.A. ppargc1a controls nephron segmentation during zebrafish embryonic kidney ontogeny. Elife 2018, 7, e40266. [Google Scholar] [CrossRef]
- Marra, A.N.; Adeeb, B.D.; Chambers, B.E.; Drummond, B.E.; Ulrich, M.; Addiego, A.; Springer, M.; Poureetezadi, S.J.; Chambers, J.M.; Ronshaugen, M.; et al. Prostaglandin signaling regulates renal multiciliated cell specification and maturation. Proc. Natl. Acad. Sci. USA 2019, 116, 8409–8418. [Google Scholar] [CrossRef] [Green Version]
- Larijani, B.; Hamidpour, S.K.; Tayanloo-Beik, A.; Shahbazbadr, A.; Yavari, H.; Namazi, N.; Biglar, M.; Arjmand, B. An overview of zebrafish modeling methods in drug discovery and development. Adv. Exp. Med. Biol. 2022, 1387, 145–169. [Google Scholar] [PubMed]
- Wesselman, H.M.; Gatz, A.E.; Pfaff, M.R.; Arceri, L.; Wingert, R.A. Estrogen signaling influences nephron segmentation of the zebrafish embryonic kidney. Cells 2023, 12, 666. [Google Scholar] [CrossRef]
- Gemberling, M.; Bailey, T.J.; Hyde, D.R.; Poss, K.D. The zebrafish as a model for complex tissue regeneration. Trends Genet. 2013, 29, 611–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCampbell, K.K.; Wingert, R.A. New tides: Using zebrafish to study renal regeneration. Transl. Res. 2014, 163, 109–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKee, R.A.; Wingert, R.A. Zebrafish renal pathology: Emerging models of acute kidney injury. Curr. Pathobiol. Rep. 2015, 3, 171–181. [Google Scholar] [CrossRef] [Green Version]
- Ross Stewart, K.M.; Walker, S.L.; Baker, A.H.; Riley, P.R.; Brittan, M. Hooked on heart regeneration: The zebrafish guide to recovery. Cardiovasc. Res. 2022, 118, 1667–1679. [Google Scholar] [CrossRef]
- Sehring, I.; Weidinger, G. Zebrafish fin: Complex molecular interactions and cellular mechanisms guiding regeneration. Cold Spring Harb. Perspect. Biol. 2022, 14, a040758. [Google Scholar]
- Howe, K.; Clark, M.; Torroja, C.; Torrance, J.; Berthelot, C.; Muffato, M.; Collins, J.E.; Humphray, S.; McLaren, K.; Matthews, L.; et al. The zebrafish reference genome sequence and its relationship to the human genome. Nature 2013, 496, 498–503. [Google Scholar]
- Howe, D.G.; Bradford, Y.M.; Eagle, A.; Fashena, D.; Frazer, K.; Kalita, P.; Mani, P.; Martin, R.; Moxon, S.T.; Paddock, H.; et al. The Zebrafish Model Organism Database: New support for human disease models, mutation details, gene expression phenotypes and searching. Nucleic Acids Res. 2017, 45, D758–D768. [Google Scholar] [CrossRef] [Green Version]
- Ruzicka, L.; Howe, D.G.; Ramachandran, S.; Toro, S.; Van Slyke, C.E.; Bradford, Y.M.; Eagle, A.; Fashena, D.; Frazer, K.; Kalita, P.; et al. The Zebrafish Information Network: New support for non-coding genes, richer Gene Ontology annotations and the Alliance of Genome Resources. Nucleic Acids Res. 2019, 47, D867–D873. [Google Scholar] [CrossRef] [Green Version]
- Bradford, Y.M.; Van Slyke, C.E.; Ruzicka, L.; Singer, A.; Eagle, A.; Fashena, D.; Howe, D.G.; Frazer, K.; Martin, R.; Paddock, H.; et al. Zebrafish Information Network, the knowledgebase for Danio rerio research. Genetics 2022, 220, iyac016. [Google Scholar] [CrossRef]
- Selland, L.G.; Koch, S.; Laraque, M.; Waskiewicz, A.J. Coordinate regulation of retinoic acid synthesis by pbx genes and fibroblast growth factor signaling by hoxb1b is required for hindbrain patterning and development. Mech. Dev. 2018, 150, 28–41. [Google Scholar] [CrossRef] [PubMed]
- Moens, C.B.; Prince, V.E. Constructing the hindbrain: Insights from the zebrafish. Dev. Dyn. 2002, 224, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Maves, L.; Kimmel, C.B. Dynamic and sequential patterning of the zebrafish posterior hindbrain by retinoic acid. Dev. Biol. 2005, 285, 593–605. [Google Scholar] [CrossRef] [Green Version]
- Hernandez, R.E.; Rikhof, H.A.; Bachmann, R.; Moens, C.B. vhnf1 integrates global RA patterning and local FGF signals to direct posterior hindbrain development in zebrafish. Development 2004, 131, 4511–4520. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, P.; Maurer, J.M.; Sagerström, C.G. Analysis of novel caudal hindbrain genes reveals different regulatory logic for gene expression in rhombomere 4 versus 5/6 in embryonic zebrafish. Neural Dev. 2018, 13, 13. [Google Scholar] [CrossRef] [Green Version]
- Qiu, Y.; Fung, L.; Schilling, T.F.; Nie, Q. Multiple morphogens and rapid elongation promote segmental patterning during development. PLoS Comput. Biol. 2021, 17, e1009077. [Google Scholar] [CrossRef]
- Molotkova, N.; Molotkov, A.; Duester, G. Role of retinoic acid during forebrain development begins late when Raldh3 generates retinoic acid in the ventral subventricular zone. Dev. Biol. 2007, 303, 601–610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gongal, P.A.; March, L.D.; Holly, V.L.; Pillay, L.M.; Berry-Wynne, K.M.; Kagechika, H.; Waskiewicz, A.J. Hmx4 regulates Sonic hedgehog signaling through control of retinoic acid synthesis during forebrain patterning. Dev. Biol. 2011, 355, 55–64. [Google Scholar] [CrossRef] [Green Version]
- Buono, L.; Corbacho, J.; Naranjo, S.; Almuedo-Castillo, M.; Moreno-Marmol, T.; de la Cerda, B.; Sanabria-Reinoso, E.; Polvillo, R.; Díaz-Corrales, F.J.; Bogdanovic, O.; et al. Analysis of gene network bifurcation during optic cup morphogenesis in zebrafish. Nat. Commun. 2021, 12, 3866. [Google Scholar] [CrossRef]
- Yin, J.; Morrissey, M.E.; Shine, L.; Kennedy, C.; Higgins, D.G.; Kennedy, B.N. Genes and signaling networks regulated during zebrafish optic vesicle morphogenesis. BMC Genomics. 2014, 15, 825. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, T.K.; Petrikas, M.; Chambers, B.E.; Wingert, R.A. Principles of nephron segment development. J. Dev. Biol. 2023, 11, 14. [Google Scholar] [CrossRef] [PubMed]
- Gerlach, G.F.; Wingert, R.A. Kidney organogenesis in the zebrafish: Insights into vertebrate nephrogenesis and regeneration. Wiley Interdiscip. Rev. Dev. Biol. 2013, 2, 559–585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naylor, R.W.; Skvarca, L.B.; Thisse, C.; Thisse, B.; Hukriede, N.A.; Davidson, A.J. BMP and retinoic acid regulate anterior-posterior patterning of the non-axial mesoderm across the dorsal-ventral axis. Nat. Commun. 2016, 7, 12197. [Google Scholar] [CrossRef] [PubMed]
- Wingert, R.A.; Selleck, R.; Yu, J.; Song, H.; Chen, Z.; Song, A.; Zhou, Y.; Thisse, B.; Thisse, C.; McMahon, A.P.; et al. The cdx genes and retinoic acid control the positioning and segmentation of the zebrafish pronephros. PLoS Genet. 2007, 3, 1922–1938. [Google Scholar] [CrossRef] [Green Version]
- de Groh, E.D.; Swanhart, L.M.; Cosentino, C.C.; Jackson, R.L.; Dai, W.; Kitchens, C.A.; Day, B.W.; Smithgall, T.E.; Hukriede, N.A. Inhibition of histone deacetylase expands the renal progenitor cell population. J. Am. Soc. Nephrol. 2010, 21, 794–802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Celá, P.; Balková, S.M.; Bryjová, A.; Horáková, D.; Míšek, I.; Richman, J.M.; Buchtová, M. Expression, function and regulation of Evi-1 during embryonic avian development. Gene Expr. Patterns 2013, 13, 343–353. [Google Scholar] [CrossRef]
- Li, Y.; Cheng, C.N.; Verdun, V.A.; Wingert, R.A. Zebrafish nephrogenesis is regulated by interactions between retinoic acid, mecom, and Notch signaling. Dev. Biol. 2014, 386, 111–122. [Google Scholar] [CrossRef] [Green Version]
- Cheng, C.N.; Wingert, R.A. Nephron proximal tubule patterning and corpuscles of Stannius formation are regulated by the sim1a transcription factor and retinoic acid in zebrafish. Dev. Biol. 2015, 399, 100–116. [Google Scholar] [CrossRef] [Green Version]
- Marra, A.N.; Wingert, R.A. Epithelial cell fate in the nephron tubule is mediated by the ETS transcription factors etv5a and etv4 during zebrafish kidney development. Dev. Biol. 2016, 411, 231–245. [Google Scholar] [CrossRef] [Green Version]
- Drummond, B.E.; Li, Y.; Marra, A.N.; Cheng, C.N.; Wingert, R.A. The tbx2a/b transcription factors direct pronephros segmentation and corpuscle of Stannius formation in zebrafish. Dev. Biol. 2017, 421, 52–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morales, E.E.; Handa, N.; Drummond, B.E.; Chambers, J.M.; Marra, A.N.; Addiego, A.; Wingert, R.A. Homeogene emx1 is required for nephron distal segment development in zebrafish. Sci. Rep. 2018, 8, 18038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marra, A.N.; Cheng, C.N.; Adeeb, B.; Addiego, A.; Wesselman, H.M.; Chambers, B.E.; Chambers, J.M.; Wingert, R.A. Iroquois transcription factor irx2a is required for multiciliated and transporter cell fate decisions during zebrafish pronephros development. Sci. Rep. 2019, 9, 6454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chambers, B.E.; Clark, E.G.; Gatz, A.E.; Wingert, R.A. Kctd15 regulates nephron segment development by repressing Tfap2a activity. Development 2020, 147, dev191973. [Google Scholar] [CrossRef] [PubMed]
- Weaver, N.E.; Healy, A.; Wingert, R.A. gldc is essential for renal progenitor patterning during kidney development. Biomedicines 2022, 10, 3220. [Google Scholar] [CrossRef]
- Wesselman, H.M.; Nguyen, T.K.; Chambers, J.M.; Drummond, B.E.; Wingert, R.A. Advances in understanding the genetic mechanisms of zebrafish renal multiciliated cell development. J. Dev. Biol. 2022, 11, 1. [Google Scholar] [CrossRef]
- Keegan, B.R.; Feldman, J.L.; Begemann, G.; Ingham, P.W.; Yelon, D. Retinoic acid signaling restricts the cardiac progenitor pool. Science 2005, 307, 247–249. [Google Scholar] [CrossRef]
- Lengerke, C.; Wingert, R.; Beeretz, M.; Grauer, M.; Schmidt, A.G.; Konantz, M.; Daley, G.Q.; Davidson, A.J. Interactions between Cdx genes and retinoic acid modulate early cardiogenesis. Dev. Biol. 2011, 354, 134–142. [Google Scholar] [CrossRef] [Green Version]
- Chang, J.; Skromne, I.; Ho, R.K. CDX4 and retinoic acid interact to position the hindbrain-spinal cord transition. Dev. Biol. 2016, 410, 178–189. [Google Scholar] [CrossRef]
- Waxman, J.S.; Keegan, B.R.; Roberts, R.W.; Poss, K.D.; Yelon, D. Hoxb5b acts downstream of retinoic acid signaling in the forelimb field to restrict heart field potential in zebrafish. Dev. Cell 2008, 15, 923–934. [Google Scholar] [CrossRef] [Green Version]
- Marques, S.R.; Lee, Y.; Poss, K.D.; Yelon, D. Reiterative roles for FGF signaling in the establishment of size and proportion of the zebrafish heart. Dev. Biol. 2008, 321, 397–406. [Google Scholar] [CrossRef] [Green Version]
- Sorrell, M.R.; Waxman, J.S. Restraint of Fgf8 signaling by retinoic acid signaling is required for proper heart and forelimb formation. Dev. Biol. 2011, 358, 44–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niederreither, K.; Vermot, J.; Messaddeq, N.; Schuhbaur, B.; Chambon, P.; Dollé, P. Embryonic retinoic acid synthesis is essential for heart morphogenesis in the mouse. Development 2001, 128, 1019–1031. [Google Scholar] [CrossRef]
- Sirbu, I.O.; Zhao, X.; Duester, G. Retinoic acid controls heart anteroposterior patterning by down-regulating Isl1 through the Fgf8 pathway. Dev. Dyn. 2008, 237, 1627–1635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rydeen, A.B.; Waxman, J.S. Cyp26 enzymes are required to balance the cardiac and vascular lineages within the anterior lateral plate mesoderm. Development 2014, 141, 1638–1648. [Google Scholar] [CrossRef] [Green Version]
- Rydeen, A.B.; Waxman, J.S. Cyp26 enzymes facilitate second heart field progenitor addition and maintenance of ventricular integrity. PLoS Biol. 2016, 14, e2000504. [Google Scholar] [CrossRef] [Green Version]
- Goldman, J.A.; Poss, K.D. Gene regulatory programmes of tissue regeneration. Nat. Rev. Genet. 2020, 21, 511–525. [Google Scholar] [CrossRef] [PubMed]
- Poss, K.D.; Wilson, L.G.; Keating, M.T. Heart regeneration in zebrafish. Science 2002, 298, 2188–2190. [Google Scholar] [CrossRef] [PubMed]
- Lepilina, A.; Coon, A.N.; Kikuchi, K.; Holdway, J.E.; Roberts, R.W.; Burns, C.G.; Poss, K.D. A dynamic epicardial injury response supports progenitor cell activity during zebrafish heart regeneration. Cell 2006, 127, 607–619. [Google Scholar] [CrossRef] [Green Version]
- Kikuchi, K.; Holdway, J.E.; Major, R.J.; Blum, N.; Dahn, R.D.; Begemann, G.; Poss, K.D. Retinoic acid production by endocardium and epicardium is an injury response essential for zebrafish heart regeneration. Dev. Cell 2011, 20, 397–404. [Google Scholar] [CrossRef] [Green Version]
- Danzl, K.; Messner, B.; Doppler, C.; Nebert, C.; Abfalterer, A.; Sakic, A.; Temml, V.; Heinz, K.; Streitwieser, R.; Edelmann, T.; et al. Early inhibition of endothelial retinoid uptake upon myocardial infarction restores cardiac function and prevents cell, tissue, and animal death. J. Mol. Cell Cardiol. 2019, 126, 105–117. [Google Scholar] [CrossRef] [Green Version]
- Da Silva, F.; Jian Motamedi, F.; Weerasinghe Arachchige, L.C.; Tison, A.; Bradford, S.T.; Lefebvre, J.; Dolle, P.; Ghyselinck, N.B.; Wagner, K.D.; Schedl, A. Retinoic acid signaling is directly activated in cardiomyocytes and protects mouse hearts from apoptosis after myocardial infarction. Elife 2021, 10, e68280. [Google Scholar] [CrossRef]
- Gore, A.V.; Pillay, L.M.; Venero Galanternik, M.; Weinstein, B.M. The zebrafish: A fintastic model for hematopoietic development and disease. Wiley Interdiscip. Rev. Dev. Biol. 2018, 7, e312. [Google Scholar] [CrossRef]
- Ransom, D.G.; Bahary, N.; Niss, K.; Traver, D.; Burns, C.; Trede, N.S.; Paffett-Lugassy, N.; Saganic, W.J.; Lim, C.A.; Hersey, C.; et al. The zebrafish moonshine gene encodes transcriptional intermediary factor 1 gamma, an essential regulator of hematopoiesis. PLoS Biol. 2004, 2, E237. [Google Scholar] [CrossRef] [Green Version]
- Rhodes, J.; Hagen, A.; Hsu, K.; Deng, M.; Liu, T.X.; Look, A.T.; Kanki, J.P. Interplay of pu.1 and gata1 determines myelo-erythroid progenitor cell fate in zebrafish. Dev. Cell 2005, 8, 97–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galloway, J.L.; Wingert, R.A.; Thisse, C.; Thisse, B.; Zon, L.I. Loss of gata1 but not gata2 converts erythropoiesis to myelopoiesis in zebrafish embryos. Dev. Cell 2005, 8, 109–116. [Google Scholar] [CrossRef]
- Donovan, A.; Brownlie, A.; Zhou, Y.; Shepard, J.; Pratt, S.J.; Moynihan, J.; Paw, B.H.; Drejer, A.; Barut, B.; Zapata, A.; et al. Positional cloning of zebrafish ferroportin1 identifies a conserved vertebrate iron exporter. Nature 2000, 403, 776–781. [Google Scholar] [CrossRef] [PubMed]
- Paw, B.H.; Davidson, A.J.; Zhou, Y.; Li, R.; Pratt, S.J.; Lee, C.; Trede, N.S.; Brownlie, A.; Donovan, A.; Liao, E.C.; et al. Cell-specific mitotic defect and dyserythropoiesis associated with erythroid band 3 deficiency. Nat. Genet. 2003, 34, 59–64. [Google Scholar] [CrossRef]
- Wingert, R.A.; Galloway, J.L.; Barut, B.; Foott, H.; Fraenkel, P.; Axe, J.L.; Weber, G.J.; Dooley, K.; Davidson, A.J.; Schmidt, B.; et al. Deficiency of glutaredoxin 5 reveals Fe-S clusters are required for vertebrate haem synthesis. Nature 2005, 436, 1035–1039. [Google Scholar] [CrossRef] [PubMed]
- Shaw, G.C.; Cope, J.J.; Li, L.; Corson, K.; Hersey, C.; Ackermann, G.E.; Gwynn, B.; Lambert, A.J.; Wingert, R.A.; Traver, D.; et al. Mitoferrin is essential for erythroid iron assimilation. Nature 2006, 440, 96–100. [Google Scholar] [CrossRef]
- De Jong, J.L.O.; Davidson, A.J.; Wang, Y.; Palis, J.; Opara, P.; Pugach, E.; Daley, G.Q.; Zon, L.I. Interaction of retinoic acid and scl controls primitive blood development. Blood 2010, 116, 201–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, D.; Jia, W.; Li, J.; Li, K.; Zhao, Q. Retinoic acid signaling plays a restrictive role in zebrafish primitive myelopoiesis. PLoS ONE 2012, 7, e30865. [Google Scholar] [CrossRef] [PubMed]
- Ma, A.C.H.; Chung, M.I.S.; Liang, R.; Leung, A.Y.H. A DEAB-sensitive aldehyde dehydrogenase regulates hematopoietic stem and progenitor cells development during primitive hematopoiesis in zebrafish embryos. Leukemia 2010, 24, 2090–2099. [Google Scholar] [CrossRef] [PubMed]
- Pillay, L.M.; Mackowetzky, K.J.; Widen, S.A.; Waskiewicz, A.J. Somite-derived retinoic acid regulates zebrafish hematopoietic stem cell formation. PLoS ONE 2016, 11, e0166040. [Google Scholar] [CrossRef] [Green Version]
- Yu, C.; Liu, Y.; Miao, Z.; Yin, M.; Lu, W.; Lv, Y.; Ding, M.; Deng, H. Retinoic acid enhances the generation of hematopoietic progenitors from human embryonic stem cell-derived hemato-vascular precursors. Blood 2010, 116, 4786–4794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chanda, B.; Ditadi, A.; Iscove, N.N.; Keller, G. Retinoic acid signaling is essential for embryonic hematopoietic stem cell development. Cell 2013, 155, 215–227. [Google Scholar] [CrossRef] [Green Version]
- Garnaas, M.K.; Cutting, C.C.; Meyers, A.; Kelsey, P.B., Jr.; Harris, J.M.; North, T.E.; Goessling, W. Rargb regulates organ laterality in a zebrafish model of right atrial isomerism. Dev. Biol. 2012, 372, 178–189. [Google Scholar] [CrossRef] [Green Version]
- Drowley, L.; McPheat, J.; Nordqvist, A.; Peel, S.; Karlsson, U.; Martinsson, S.; Müllers, E.; Dellsén, A.; Knight, S.; Barrett, I.; et al. Discovery of retinoic acid receptor agonists as proliferators of cardiac progenitor cells through a phenotypic screening approach. Stem. Cells Transl. Med. 2020, 9, 47–60. [Google Scholar] [CrossRef] [Green Version]
- Nadauld, L.D.; Sandoval, I.T.; Chidester, S.; Yost, H.J.; Jones, D.A. Adenomatous polyposis coli control of retinoic acid biosynthesis is critical for zebrafish intestinal development and differentiation. J. Biol. Chem. 2004, 279, 51581–51589. [Google Scholar] [CrossRef] [Green Version]
- Nadauld, L.D.; Shelton, D.N.; Chidester, S.; Yost, H.J.; Jones, D.A. The zebrafish retinol dehydrogenase, rdh1l, is essential for intestinal development and is regulated by the tumor suppressor adenomatous polyposis coli. J. Biol. Chem. 2005, 280, 30490–30495. [Google Scholar] [CrossRef] [Green Version]
- Martín, M.; Gallego-Llamas, J.; Ribes, V.; Kedinger, M.; Niederreither, K.; Chambon, P.; Dollé, P.; Gradwohl, G. Dorsal pancreas agenesis in retinoic acid-deficient Raldh2 mutant mice. Dev. Biol. 2005, 284, 399–411. [Google Scholar] [CrossRef] [PubMed]
- Biemar, F.; Argenton, F.; Schmidtke, R.; Epperlein, S.; Peers, B.; Driever, W. Pancreas development in zebrafish: Early dispersed appearance of endocrine hormone expressing cells and their convergence to form the definitive islet. Dev. Biol. 2001, 230, 189–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stafford, D.; Prince, V.E. Retinoic acid signaling is required for a critical early step in zebrafish pancreatic development. Curr. Biol. 2002, 12, 1215–1220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stafford, D.; White, R.J.; Kinkel, M.D.; Linville, A.; Schilling, T.F.; Prince, V.E. Retinoids signal directly to zebrafish endoderm to specify insulin-expressing beta-cells. Development 2006, 133, 949–956. [Google Scholar] [CrossRef] [Green Version]
- Tehrani, Z.; Lin, S. Antagonistic interactions of hedgehog, Bmp and retinoic acid signals control zebrafish endocrine pancreas development. Development 2011, 138, 631–640. [Google Scholar] [CrossRef] [Green Version]
- Lorberbaum, D.S.; Kishore, S.; Rosselot, C.; Sarbaugh, D.; Brooks, E.P.; Aragon, E.; Xuan, S.; Simon, O.; Ghosh, D.; Mendelsohn, C.; et al. Retinoic acid signaling within pancreatic endocrine progenitors regulates mouse and human β cell specification. Development 2020, 147, dev189977. [Google Scholar] [CrossRef] [PubMed]
- Dalgin, G.; Prince, V.E. Midline morphogenesis of zebrafish foregut endoderm is dependent on Hoxb5b. Dev. Biol. 2021, 471, 1–9. [Google Scholar] [CrossRef]
- Fernandes, Y.; Lovely, C.B. Zebrafish models of fetal alcohol spectrum disorders. Genesis 2021, 59, e23460. [Google Scholar] [CrossRef]
- Manikandan, P.; Sarmah, S.; Marrs, J.A. Ethanol effects on early developmental stages studied using the zebrafish. Biomedicines 2022, 10, 2555. [Google Scholar] [CrossRef]
- Marrs, J.A.; Clendenon, S.G.; Ratcliffe, D.R.; Fielding, S.M.; Liu, Q.; Bosron, W.F. Zebrafish fetal alcohol syndrome model: Effects of ethanol are rescued by retinoic acid supplement. Alcohol 2010, 44, 707–715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muralidharan, P.; Sarmah, S.; Marrs, J.A. Zebrafish retinal defects induced by ethanol exposure are rescued by retinoic acid and folic acid supplement. Alcohol 2015, 49, 149–163. [Google Scholar] [CrossRef] [Green Version]
- Nordestgaard, L.; Ona, S. From Template “Risk Factors of Dementia”, by BioRender.com. 2023. Available online: https://app.biorender.com/biorender-templates (accessed on 1 March 2023).
- Lammer, E.J.; Chen, D.T.; Hoar, R.M.; Agnish, N.D.; Benke, P.J.; Braun, J.T.; Curry, C.J.; Fernhoff, P.M.; Grix, A.W., Jr.; Lott, I.T.; et al. Retinoic acid embryopathy. N. Engl. J. Med. 1985, 313, 837–841. [Google Scholar] [CrossRef] [PubMed]
- Draghici, C.C.; Miulescu, R.G.; Petca, R.C.; Petca, A.; Dumitrașcu, M.C.; Șandru, F. Teratogenic effect of isotretinoin in both fertile females and males. Exp. Ther. Med. 2021, 21, 534. [Google Scholar] [CrossRef] [PubMed]
- He, S.; Mansour, M.R.; Zimmerman, M.W.; Ki, D.H.; Layden, H.M.; Akahane, K.; Gjini, E.; de Groh, E.D.; Perez-Atayde, A.R.; Zhu, S.; et al. Synergy between loss of NF1 and overexpression of MYCN in neuroblastoma is mediated by the GAP-related domain. Elife 2016, 5, e14713. [Google Scholar] [CrossRef]
- Lam, S.H.; Wu, Y.L.; Vega, V.B.; Miller, L.D.; Spitsbergen, J.; Tong, Y.; Zhan, H.; Govindarajan, K.R.; Lee, S.; Mathavan, S.; et al. Conservation of gene expression signatures between zebrafish and human liver tumors and tumor progression. Nat. Biotechnol. 2006, 24, 73–75. [Google Scholar] [CrossRef]
- Kobar, K.; Collett, K.; Prykhozhij, S.V.; Berman, J.N. Zebrafish cancer predisposition models. Front. Cell Dev. Biol. 2021, 9, 660069. [Google Scholar] [CrossRef]
- Haldi, M.; Ton, C.; Seng, W.L.; McGrath, P. Human melanoma cells transplanted into zebrafish proliferate, migrate, produce melanin, form masses and stimulate angiogenesis in zebrafish. Angiogenesis 2006, 9, 139–151. [Google Scholar] [CrossRef]
- Nicoli, S.; Ribatti, D.; Cotelli, F.; Presta, M. Mammalian tumor xenografts induce neovascularization in zebrafish embryos. Cancer Res. 2007, 67, 2927–2931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fazio, M.; Ablain, J.; Chuan, Y.; Langenau, D.M.; Zon, L.I. Zebrafish patient avatars in cancer biology and precision cancer therapy. Nat. Rev. Cancer 2020, 20, 263–273. [Google Scholar] [CrossRef]
- Mandelbaum, J.; Shestopalov, I.A.; Henderson, R.E.; Chau, N.G.; Knoechel, B.; Wick, M.J.; Zon, L.I. Zebrafish blastomere screen identifies retinoic acid suppression of MYB in adenoid cystic carcinoma. J. Exp. Med. 2018, 215, 2673–2685. [Google Scholar] [CrossRef]
- Weiss, F.U.; Marques, I.J.; Woltering, J.M.; Vlecken, D.H.; Aghdassi, A.; Partecke, L.I.; Heidecke, C.D.; Lerch, M.M.; Bagowski, C.P. Retinoic acid receptor antagonists inhibit miR-10a expression and block metastatic behavior of pancreatic cancer. Gastroenterology 2009, 137, 2136–2145.e1-e7. [Google Scholar] [CrossRef] [PubMed]
- Barbieri, E.; De Preter, K.; Capasso, M.; Chen, Z.; Hsu, D.M.; Tonini, G.P.; Lefever, S.; Hicks, J.; Versteeg, R.; Pession, A.; et al. Histone chaperone CHAF1A inhibits differentiation and promotes aggressive neuroblastoma. Cancer Res. 2014, 74, 765–774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tao, L.; Moreno-Smith, M.; Ibarra-García-Padilla, R.; Milazzo, G.; Drolet, N.A.; Hernandez, B.E.; Oh, Y.S.; Patel, I.; Kim, J.J.; Zorman, B.; et al. CHAF1A blocks neuronal differentiation and promotes neuroblastoma oncogenesis via metabolic reprogramming. Adv. Sci. 2021, 8, e2005047. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.; Glasgow, E.; Agarwal, S. Zebrafish xenografts for drug discovery and personalized medicine. Trends Cancer 2020, 6, 569–579. [Google Scholar] [CrossRef]
Figure 1.
Overview of RA biosynthesis and the canonical pathway of transcriptional activation. Schematic provides a view of the cell membrane, with flanking extracellular space (white) and the cytosol (blue), the latter containing the nuclear compartment (purple). Retinol enters the cell via STRA6(l) proteins, and is bound by CRBP. Next, it can be reversibly esterified for storage by LRAT, or processed by two sequential oxidation reactions by RDH and ALDH enzymes to form ATRA. ATRA can be bound and enter the nucleus upon interactions with RAR/RXR nuclear receptors, or degraded via the action of CYP26 enzymes. Figure created with BioRender.com.
Figure 1.
Overview of RA biosynthesis and the canonical pathway of transcriptional activation. Schematic provides a view of the cell membrane, with flanking extracellular space (white) and the cytosol (blue), the latter containing the nuclear compartment (purple). Retinol enters the cell via STRA6(l) proteins, and is bound by CRBP. Next, it can be reversibly esterified for storage by LRAT, or processed by two sequential oxidation reactions by RDH and ALDH enzymes to form ATRA. ATRA can be bound and enter the nucleus upon interactions with RAR/RXR nuclear receptors, or degraded via the action of CYP26 enzymes. Figure created with BioRender.com.

Figure 2.
Zebrafish as a model organism. (A) Zebrafish adults reach sexual maturity between 2–3 months of age and mate via broadcast spawning into the water column. (B) Thus, fertilized embryos can be collected non-invasively and grown in the lab. Embryos develop optimally at a temperature of 28.5 °C, as they are tropical fish, and can be grown in various formats. Here, we show the example of arraying the embryos in multi-well plates to conduct exogenous treatments with small molecules to interrogate developmental processes and for toxicology studies. The use of RA pathway agonists or antagonists, like ATRA or DEAB, can be used to interrogate how changes in RA biosynthesis or signaling affect ontogeny. Such treatments can begin at any timepoint of interest to the researcher, such as the 60% epiboly depicted here, and then embryos reared to the stage(s) of interest, such as 24 hpf, to examine the effects by any number of phenotype assessments, such as live assessment, WISH, or qRT-PCR. Figure created with BioRender.com.
Figure 2.
Zebrafish as a model organism. (A) Zebrafish adults reach sexual maturity between 2–3 months of age and mate via broadcast spawning into the water column. (B) Thus, fertilized embryos can be collected non-invasively and grown in the lab. Embryos develop optimally at a temperature of 28.5 °C, as they are tropical fish, and can be grown in various formats. Here, we show the example of arraying the embryos in multi-well plates to conduct exogenous treatments with small molecules to interrogate developmental processes and for toxicology studies. The use of RA pathway agonists or antagonists, like ATRA or DEAB, can be used to interrogate how changes in RA biosynthesis or signaling affect ontogeny. Such treatments can begin at any timepoint of interest to the researcher, such as the 60% epiboly depicted here, and then embryos reared to the stage(s) of interest, such as 24 hpf, to examine the effects by any number of phenotype assessments, such as live assessment, WISH, or qRT-PCR. Figure created with BioRender.com.

Figure 3.
RA levels influence proximo-distal nephron fates during embryonic kidney development in the zebrafish. (Left) Renal progenitors are exposed to a gradient of RA during gastrulation and axis formation, with the highest level at the rostral-most locations (arrow). (Right, top) As development proceeds, these RA levels exert an influence on the proximo-distal segmentation patterning. The zebrafish embryonic kidney comprises two bilateral nephrons with several unique cell populations, and these undergo morphogenesis events by the 48 stage to connect rostrally at a single midline blood filter and caudally at the cloaca where waste exits the body. (Right, bottom) The effects of RA are best understood for the 4 main tubule populations: the proximal convoluted tubule (PCT), proximal straight tubule (PST), distal early (DE), and distal late (DL). Embryos exposed to a high dosage of ATRA starting at 60% epiboly form nephrons with only proximal segments. Conversely, embryos exposed to the RA biosynthesis inhibitor DEAB starting at 60% epiboly form nephrons with only distal segments. Figure created with BioRender.com and based on [175].
Figure 3.
RA levels influence proximo-distal nephron fates during embryonic kidney development in the zebrafish. (Left) Renal progenitors are exposed to a gradient of RA during gastrulation and axis formation, with the highest level at the rostral-most locations (arrow). (Right, top) As development proceeds, these RA levels exert an influence on the proximo-distal segmentation patterning. The zebrafish embryonic kidney comprises two bilateral nephrons with several unique cell populations, and these undergo morphogenesis events by the 48 stage to connect rostrally at a single midline blood filter and caudally at the cloaca where waste exits the body. (Right, bottom) The effects of RA are best understood for the 4 main tubule populations: the proximal convoluted tubule (PCT), proximal straight tubule (PST), distal early (DE), and distal late (DL). Embryos exposed to a high dosage of ATRA starting at 60% epiboly form nephrons with only proximal segments. Conversely, embryos exposed to the RA biosynthesis inhibitor DEAB starting at 60% epiboly form nephrons with only distal segments. Figure created with BioRender.com and based on [175].

Figure 4.
Summary of zebrafish models of RA signaling in development and disease. Research using the zebrafish has been a powerful good for realizing new insights into the development of many tissues and organs. Numbering among them, and discussed in the present work, are the neural plate, eye, kidney, heart, blood, pancreas, liver, and intestine. Many others have been studied as well [17]. Our understanding about the teratogenic effects of RA in conditions such as fetal alcohol syndrome and its role in cancer have also been expanded through the use of zebrafish. Figure created with BioRender.com [232].
Figure 4.
Summary of zebrafish models of RA signaling in development and disease. Research using the zebrafish has been a powerful good for realizing new insights into the development of many tissues and organs. Numbering among them, and discussed in the present work, are the neural plate, eye, kidney, heart, blood, pancreas, liver, and intestine. Many others have been studied as well [17]. Our understanding about the teratogenic effects of RA in conditions such as fetal alcohol syndrome and its role in cancer have also been expanded through the use of zebrafish. Figure created with BioRender.com [232].

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Hawkins, M.R.; Wingert, R.A. Zebrafish as a Model to Study Retinoic Acid Signaling in Development and Disease. Biomedicines 2023, 11, 1180. https://doi.org/10.3390/biomedicines11041180
AMA Style
Hawkins MR, Wingert RA. Zebrafish as a Model to Study Retinoic Acid Signaling in Development and Disease. Biomedicines. 2023; 11(4):1180. https://doi.org/10.3390/biomedicines11041180
Chicago/Turabian StyleHawkins, Matthew R., and Rebecca A. Wingert. 2023. "Zebrafish as a Model to Study Retinoic Acid Signaling in Development and Disease" Biomedicines 11, no. 4: 1180. https://doi.org/10.3390/biomedicines11041180
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.