The Competition of Yin and Yang: Exploring the Role of Wild-Type and Mutant p53 in Tumor Progression




1
School of Medicine, I-Shou University, Kaohsiung City 82445, Taiwan
2
Department of Plastic Surgery, E-Da Hospital, I-Shou University, Kaohsiung City 82445, Taiwan
3
Department of Physical Therapy, I-Shou University, Kaohsiung City 82445, Taiwan
4
School of Chinese Medicine for Post Baccalaureate, I-Shou University, Kaohsiung City 82445, Taiwan
5
Department of Pathology, E-Da Hospital, I-Shou University, Kaohsiung City 82445, Taiwan
6
Department of Biological Science and Technology, I-Shou University, Kaohsiung City 82445, Taiwan
7
Institute of Medicine, Chung Shan Medical University, Taichung City 40201, Taiwan
*
Authors to whom correspondence should be addressed.
Biomedicines 2023, 11(4), 1192; https://doi.org/10.3390/biomedicines11041192
Submission received: 5 April 2023
/
Accepted: 14 April 2023
/
Published: 17 April 2023
(This article belongs to the Special Issue Roles of p53 Family in Cancers and Their Therapeutic Approaches)
{kind=link}
{kind=link}
The protein p53 is a well-known tumor suppressor that plays a crucial role in preventing cancer development. In Chinese philosophy, the concept of Yin and Yang is used to describe inseparable opposites, where Yin represents the negative or inhibitory force, and Yang represents the positive or promoting force. In this context, the wild type of p53 can be considered Yin, as it acts as a negative regulator of cancer progression, while most p53 mutations can be considered Yang, as they promote cancer progression.
Some p53 mutants, such as p53 S46F and S121F, exhibit higher transactivation activity than wild type, which can be considered as a super Yin of p53 [1,2,3]. Double mutations of p53 in mice, such as p53 F53Q and F54S, have been shown to enhance transactivation function and tumor suppression capacity, which can be considered as a super Yin of p53 [4]. The p53 protein forms a tetramer with four subunits, which are responsible for its fully transactivation function (Figure 1A) [5,6]. If we consider only the effect of mutated p53 on blocking wild type p53 activity, it takes three mutated p53 molecules within the tetramer to block the whole p53 tetramer transactivation activity (Figure 1B) [7]. In contrast, only one molecule of ΔNp53 isoform within the tetramer can break down the p53 tetramer transactivation activity (Figure 1C) [7]. Recent studies have revealed that some p53 mutants not only lose their transactivation function, but also gain oncogenic functions. One such function is their ability to become prion-like aggregation proteins that can pull down other tumor suppressor genes [8,9,10,11].
It has been known that some mutated p53 can appear in the cytosol [12], and cytosolic mutated p53 can block autophagy [13]. However, autophagy can also promote the degradation of mutated p53 [14,15]. The major aggregative proteins detected by the A11 antibody, which is specific to staining aggregative proteins, were found to co-localize with the signal detected by the p53 antibody within the nucleus of cells with p53 R280K mutation [9]. Previously, our research has shown that most p53 R175H molecules co-localize with thioflavin T (a stain reagent used to detect aggregative proteins) in the nucleus. However, some of these co-localized signals also appear in the cytosol [16]. A recent study reported that the presence of cytoplasmic mutant p53 aggregates is associated with poor prognosis in patients with high-grade serous ovarian carcinoma [17]. Therefore, cytosolic aggregated mutated p53 may be considered a super Yang molecule that promotes cancer progression. Some p53 mutants gain oncogenic functions by activating the PI3K-AKT pathway [18,19], which in turn inhibits autophagy [20,21]. Cytosolic aggregated mutated p53 may have an even greater effect in blocking autophagy through AKT or other oncogenic pathways. Moreover, autophagy may find it more difficult to clear aggregated mutated p53 in the cytosol. Additionally, AKT can phosphorylate MDM2, leading to the degradation of wild type p53 [22]. Functional members of the p53 family, such as p53, p63, and p73, can activate autophagy-related genes (ATGs) such as α2 and γ subunits of AMPK [23,24]. AMPK is a positive regulator of autophagy [25] and can also phosphorylate p53 family members to enhance their anticancer function [26,27]. p53, p63, and p73 can activate PUMA, which induces cell apoptosis [28,29,30]. However, the PI3K-AKT pathway can suppress PUMA expression [31,32], and as shown in Figure 2, mutant p53 can prevent cell apoptosis through the PI3K-AKT-PUMA pathway. We previously summarized the activators of p63 or p73 in p53 mutant cancers [33], and it is possible that the super Yang effect of cytosolic aggregated mutated p53 may confer higher drug resistance, making it harder to induce cancer cell apoptosis. Identifying drugs that can remove the super Yang effect of cytosolic aggregated mutated p53 or introducing/reactivating super Yin p53 family members to activate autophagy for cleaning mutated p53 are key issues in developing anticancer therapies.
Author Contributions
B.-H.C. and I.-L.H. wrote the manuscript. B.-H.C., Y.-T.S., C.-C.C., J.-F.S. and I.-L.H. conducted the review and editing. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the grants from the National Science and Technology Council, Taiwan (NSTC 112-2823-8-214-001 to professor J.-F.S.), and I-Shou University and E-Da Medical Con-sortium Legal Person Research Cooperation Project [(ISU-110-IUC-08 to B.-H.C. and C.-C.C.).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Nakamura, Y.; Futamura, M.; Kamino, H.; Yoshida, K.; Arakawa, H. Identification of p53–46F as a super p53 with an enhanced ability to induce p53-dependent apoptosis. Cancer Sci. 2006, 97, 633–641. [Google Scholar] [CrossRef] [PubMed]
- Anazawa, Y.; Arakawa, H.; Nakagawa, H.; Nakamura, Y. Identification of STAG1 as a key mediator of a p53-dependent apoptotic pathway. Oncogene 2004, 23, 7621–7627. [Google Scholar] [CrossRef] [PubMed]
- Cai, B.H.; Chen, J.Y.; Lu, M.H.; Chang, L.T.; Lin, H.C.; Chang, Y.M.; Chao, C.F. Functional four-base A/T gap core sequence CATTAG of P53 response elements specifically bound tetrameric P53 differently than two-base A/T gap core sequence CATG bound both dimeric and tetrameric P53. Nucleic Acids Res. 2009, 37, 1984–1990. [Google Scholar] [CrossRef] [PubMed]
- Mello, S.S.; Valente, L.J.; Raj, N.; Seoane, J.A.; Flowers, B.M.; McClendon, J.; Bieging-Rolett, K.T.; Lee, J.; Ivanochko, D.; Kozak, M.M.; et al. A p53 Super-tumor Suppressor Reveals a Tumor Suppressive p53-Ptpn14-Yap Axis in Pancreatic Cancer. Cancer Cell 2017, 32, 460–473. [Google Scholar] [CrossRef]
- McLure, K.G.; Lee, P.W. How p53 binds DNA as a tetramer. EMBO J. 1998, 17, 3342–3350. [Google Scholar] [CrossRef]
- Cai, B.H.; Chao, C.F.; Huang, H.C.; Lee, H.Y.; Kannagi, R.; Chen, J.Y. Roles of p53 Family Structure and Function in Non-Canonical Response Element Binding and Activation. Int. J. Mol. Sci. 2019, 20, 3681. [Google Scholar] [CrossRef]
- Chan, W.M.; Siu, W.Y.; Lau, A.; Poon, R.Y. How many mutant p53 molecules are needed to inactivate a tetramer? Mol. Cell. Biol. 2004, 24, 3536–3551. [Google Scholar] [CrossRef]
- Xu, J.; Reumers, J.; Couceiro, J.R.; De Smet, F.; Gallardo, R.; Rudyak, S.; Cornelis, A.; Rozenski, J.; Zwolinska, A.; Marine, J.C.; et al. Gain of function of mutant p53 by coaggregation with multiple tumor suppressors. Nat. Chem. Biol. 2011, 7, 285–295. [Google Scholar] [CrossRef]
- Ano Bom, A.P.; Rangel, L.P.; Costa, D.C.; de Oliveira, G.A.; Sanches, D.; Braga, C.A.; Gava, L.M.; Ramos, C.H.; Cepeda, A.O.; Stumbo, A.C.; et al. Mutant p53 aggregates into prion-like amyloid oligomers and fibrils: Implications for cancer. J. Biol. Chem. 2012, 287, 28152–28162. [Google Scholar] [CrossRef]
- Rangel, L.P.; Costa, D.C.; Vieira, T.C.; Silva, J.L. The aggregation of mutant p53 produces prion-like properties in cancer. Prion 2014, 8, 75–84. [Google Scholar] [CrossRef]
- de Oliveira, G.A.P.; Petronilho, E.C.; Pedrote, M.M.; Marques, M.A.; Vieira, T.C.R.G.; Cino, E.A.; Silva, J.L. The Status of p53 Oligomeric and Aggregation States in Cancer. Biomolecules 2020, 10, 548. [Google Scholar] [CrossRef]
- Iwao, C.; Shidoji, Y. Induction of nuclear translocation of mutant cytoplasmic p53 by geranylgeranoic acid in a human hepatoma cell line. Sci. Rep. 2014, 4, 4419. [Google Scholar] [CrossRef]
- Morselli, E.; Tasdemir, E.; Maiuri, M.C.; Galluzzi, L.; Kepp, O.; Criollo, A.; Vicencio, J.M.; Soussi, T.; Kroemer, G. Mutant p53 protein localized in the cytoplasm inhibits autophagy. Cell Cycle 2008, 7, 3056–3061. [Google Scholar] [CrossRef]
- Garufi, A.; Pucci, D.; D’Orazi, V.; Cirone, M.; Bossi, G.; Avantaggiati, M.L.; D’Orazi, G. Degradation of mutant p53H175 protein by Zn(II) through autophagy. Cell Death Dis. 2014, 5, e1271. [Google Scholar] [CrossRef]
- Binayke, A.; Mishra, S.; Suman, P.; Das, S.; Chander, H. Awakening the “guardian of genome”: Reactivation of mutant p53. Cancer Chemother. Pharm. 2019, 83, 1–15. [Google Scholar] [CrossRef]
- Cai, B.H.; Bai, Z.Y.; Lien, C.F.; Yu, S.J.; Lu, R.Y.; Wu, M.H.; Wu, W.C.; Chen, C.C.; Hsu, Y.C. NAMPT Inhibitor and P73 Activator Represses P53 R175H Mutated HNSCC Cell Proliferation in a Synergistic Manner. Biomolecules 2022, 12, 438. [Google Scholar] [CrossRef]
- Iwahashi, N.; Ikezaki, M.; Komohara, Y.; Fujiwara, Y.; Noguchi, T.; Nishioka, K.; Sakai, K.; Nishio, K.; Ueda, M.; Ihara, Y.; et al. Cytoplasmic p53 aggregates accumulated in p53-mutated cancer correlate with poor prognosis. PNAS Nexus 2022, 1, pgac128. [Google Scholar] [CrossRef]
- Qin, Z.Q.; Li, Q.G.; Yi, H.; Lu, S.S.; Huang, W.; Rong, Z.X.; Tang, Y.Y.; Xiao, Z.Q. Heterozygous p53-R280T Mutation Enhances the Oncogenicity of NPC Cells Through Activating PI3K-Akt Signaling Pathway. Front. Oncol. 2020, 10, 104. [Google Scholar] [CrossRef]
- Yue, X.; Wu, F.; Li, Y.; Liu, J.; Boateng, M.; Mandava, K.; Zhang, C.; Feng, Z.; Gao, J.; Hu, W. Gain of function mutant p53 protein activates AKT through the Rac1 signaling to promote tumorigenesis. Cell Cycle 2020, 19, 1338–1351. [Google Scholar] [CrossRef]
- Wang, R.C.; Wei, Y.; An, Z.; Zou, Z.; Xiao, G.; Bhagat, G.; White, M.; Reichelt, J.; Levine, B. Akt-mediated regulation of autophagy and tumorigenesis through Beclin 1 phosphorylation. Science 2012, 338, 956–959. [Google Scholar] [CrossRef]
- Choi, A.M.; Ryter, S.W.; Levine, B. Autophagy in human health and disease. N. Engl. J. Med. 2013, 368, 651–662. [Google Scholar] [CrossRef] [PubMed]
- Ogawara, Y.; Kishishita, S.; Obata, T.; Isazawa, Y.; Suzuki, T.; Tanaka, K.; Masuyama, N.; Gotoh, Y. Akt enhances Mdm2-mediated ubiquitination and degradation of p53. J. Biol. Chem. 2002, 277, 21843–21850. [Google Scholar] [CrossRef] [PubMed]
- Napoli, M.; Flores, E.R. The family that eats together stays together: New p53 family transcriptional targets in autophagy. Genes Dev. 2013, 27, 971–974. [Google Scholar] [CrossRef] [PubMed]
- Kenzelmann Broz, D.; Spano Mello, S.; Bieging, K.T.; Jiang, D.; Dusek, R.L.; Brady, C.A.; Sidow, A.; Attardi, L.D. Global genomic profiling reveals an extensive p53-regulated autophagy program contributing to key p53 responses. Genes Dev. 2013, 27, 1016–1031. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Kundu, M.; Viollet, B.; Guan, K.L. AMPK and mTOR regulate autophagy through direct phosphorylation of Ulk1. Nat. Cell Biol. 2011, 13, 132–141. [Google Scholar] [CrossRef]
- Adamovich, Y.; Adler, J.; Meltser, V.; Reuven, N.; Shaul, Y. AMPK couples p73 with p53 in cell fate decision. Cell Death Differ. 2014, 21, 1451–1459. [Google Scholar] [CrossRef]
- Yi, Y.; Zhang, W.; Yi, J.; Xiao, Z.X. Role of p53 Family Proteins in Metformin Anti-Cancer Activities. J. Cancer 2019, 10, 2434–2442. [Google Scholar] [CrossRef]
- Nakano, K.; Vousden, K.H. PUMA, a novel proapoptotic gene, is induced by p53. Mol. Cell. 2001, 7, 683–694. [Google Scholar] [CrossRef]
- Ming, L.; Sakaida, T.; Yue, W.; Jha, A.; Zhang, L.; Yu, J. Sp1 and p73 activate PUMA following serum starvation. Carcinogenesis 2008, 29, 1878–1884. [Google Scholar] [CrossRef]
- Cai, B.H.; Chao, C.F.; Lin, H.C.; Huang, H.Y.; Kannagi, R.; Chen, J.Y. A/T gap tolerance in the core sequence and flanking sequence requirements of non-canonical p53 response elements. J. Biochem. 2016, 159, 563–572. [Google Scholar] [CrossRef]
- Amente, S.; Zhang, J.; Lavadera, M.L.; Lania, L.; Avvedimento, E.V.; Majello, B. Myc and PI3K/AKT signaling cooperatively repress FOXO3a-dependent PUMA and GADD45a gene expression. Nucleic Acids Res. 2011, 39, 9498–9507. [Google Scholar] [CrossRef]
- Coloff, J.L.; Mason, E.F.; Altman, B.J.; Gerriets, V.A.; Liu, T.; Nichols, A.N.; Zhao, Y.; Wofford, J.A.; Jacobs, S.R.; Ilkayeva, O.; et al. Akt requires glucose metabolism to suppress puma expression and prevent apoptosis of leukemic T cells. J. Biol. Chem. 2011, 286, 5921–5933. [Google Scholar] [CrossRef]
- Cai, B.H.; Hsu, Y.C.; Yeh, F.Y.; Lin, Y.R.; Lu, R.Y.; Yu, S.J.; Shaw, J.F.; Wu, M.H.; Tsai, Y.Z.; Lin, Y.C.; et al. P63 and P73 Activation in Cancers with p53 Mutation. Biomedicines 2022, 10, 1490. [Google Scholar] [CrossRef]
Figure 1.
Effects of mutant p53 and ΔNp53 on the transactivity of wild type p53. (A) Wild type p53 binds to DNA response elements as a tetramer and activates the transcription of its target genes. (B) Mutant p53 can fully impair the transcriptional activity of wild-type p53 by forming a heterotetramer consisting of three mutant p53 subunits and one wild-type p53 subunit. (C) ΔNp53 is an isoform of p53 that lacks the transactivation domain. When one ΔNp53 subunit forms a heterotetramer with three wild type p53 subunits, the transcriptional activity of wild type p53 is completely abolished.
Figure 1.
Effects of mutant p53 and ΔNp53 on the transactivity of wild type p53. (A) Wild type p53 binds to DNA response elements as a tetramer and activates the transcription of its target genes. (B) Mutant p53 can fully impair the transcriptional activity of wild-type p53 by forming a heterotetramer consisting of three mutant p53 subunits and one wild-type p53 subunit. (C) ΔNp53 is an isoform of p53 that lacks the transactivation domain. When one ΔNp53 subunit forms a heterotetramer with three wild type p53 subunits, the transcriptional activity of wild type p53 is completely abolished.

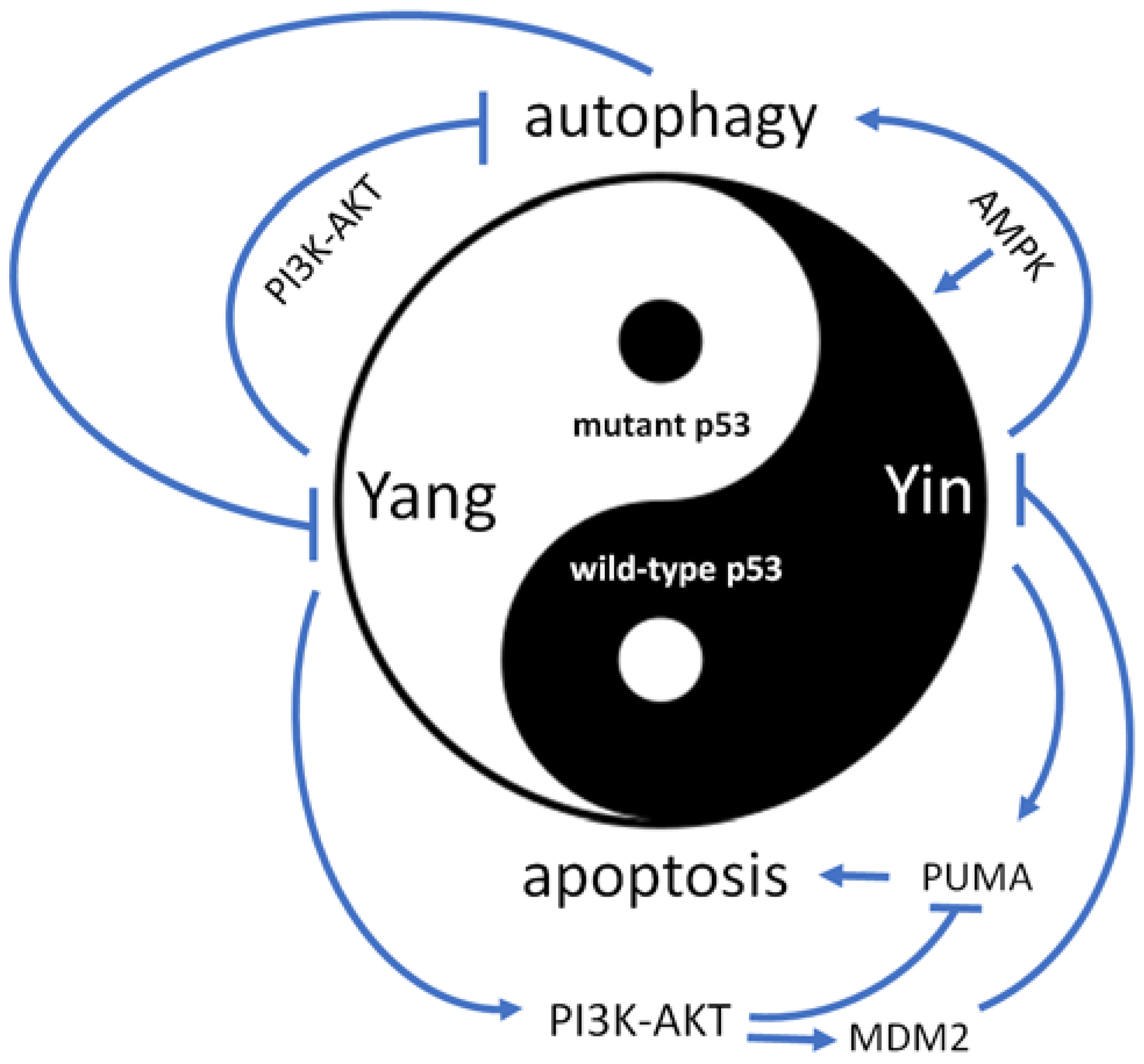
Figure 2.
The competition between wild type and mutant p53 plays a crucial role in tumor progression. While wild type p53 acts as a negative factor (Yin), mutant p53 serves as a positive factor (Yang) in cancer development. Mutant p53 can activate the PI3K-AKT signaling pathway, which induces the MDM2-mediated degradation of wild type p53 and inhibits autophagy. On the other hand, wild type p53 can directly activate AMPK, which in turn activates autophagy and up-regulates p53′s anticancer function. Autophagy also plays a role in decreasing the expression of mutant p53. Wild type p53 can activate the pro-apoptotic protein PUMA, while the PI3K-AKT pathway can inhibit its expression. Mutant p53 can thus prevent cell apoptosis by activating the PI3K-AKT pathway.
Figure 2.
The competition between wild type and mutant p53 plays a crucial role in tumor progression. While wild type p53 acts as a negative factor (Yin), mutant p53 serves as a positive factor (Yang) in cancer development. Mutant p53 can activate the PI3K-AKT signaling pathway, which induces the MDM2-mediated degradation of wild type p53 and inhibits autophagy. On the other hand, wild type p53 can directly activate AMPK, which in turn activates autophagy and up-regulates p53′s anticancer function. Autophagy also plays a role in decreasing the expression of mutant p53. Wild type p53 can activate the pro-apoptotic protein PUMA, while the PI3K-AKT pathway can inhibit its expression. Mutant p53 can thus prevent cell apoptosis by activating the PI3K-AKT pathway.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Cai, B.-H.; Sung, Y.-T.; Chen, C.-C.; Shaw, J.-F.; Hsin, I.-L. The Competition of Yin and Yang: Exploring the Role of Wild-Type and Mutant p53 in Tumor Progression. Biomedicines 2023, 11, 1192. https://doi.org/10.3390/biomedicines11041192
AMA Style
Cai B-H, Sung Y-T, Chen C-C, Shaw J-F, Hsin I-L. The Competition of Yin and Yang: Exploring the Role of Wild-Type and Mutant p53 in Tumor Progression. Biomedicines. 2023; 11(4):1192. https://doi.org/10.3390/biomedicines11041192
Chicago/Turabian StyleCai, Bi-He, Yu-Te Sung, Chia-Chi Chen, Jei-Fu Shaw, and I-Lun Hsin. 2023. "The Competition of Yin and Yang: Exploring the Role of Wild-Type and Mutant p53 in Tumor Progression" Biomedicines 11, no. 4: 1192. https://doi.org/10.3390/biomedicines11041192
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.