

Screening for Neuroprotective and Rapid Antidepressant-like Effects of 20 Essential Oils
by
 , , ,
, , ,
Khoa Nguyen Tran
† ,
,
Nhi Phuc Khanh Nguyen
† ,
,
Ly Thi Huong Nguyen
 ,
,
Heung-Mook Shin
* and
In-Jun Yang
* Department of Physiology, College of Korean Medicine, Dongguk University, Gyeongju 38066, Republic of Korea
*
Authors to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Biomedicines 2023, 11(5), 1248; https://doi.org/10.3390/biomedicines11051248
Submission received: 13 March 2023
/
Revised: 13 April 2023
/
Accepted: 21 April 2023
/
Published: 23 April 2023
(This article belongs to the Special Issue Neurodegeneration in Cognitive Impairment and Mood Disorders for Experimental, Clinical and Translational Neuropsychiatry)
Abstract
:Depression is a serious psychiatric disorder with high prevalence, and the delayed onset of antidepressant effects remains a limitation in the treatment of depression. This study aimed to screen essential oils that have the potential for rapid-acting antidepressant development. PC12 and BV2 cells were used to identify essential oils with neuroprotective effects at doses of 0.1 and 1 µg/mL. The resulting candidates were treated intranasally (25 mg/kg) to ICR mice, followed by a tail suspension test (TST) and an elevated plus maze (EPM) after 30 min. In each effective essential oil, five main compounds were computationally analyzed, targeting glutamate receptor subunits. As a result, 19 essential oils significantly abolished corticosterone (CORT)-induced cell death and lactate dehydrogenase (LDH) leakage, and 13 reduced lipopolysaccharide (LPS)-induced tumor necrosis factor alpha (TNF-α) and interleukin 6 (IL-6). From in vivo experiments, six essential oils decreased the immobility time of mice in the TST, in which Chrysanthemum morifolium Ramat. and Myristica fragrans Houtt. also increased time and entries into the open arms of the EPM. Four compounds including atractylon, α-curcumene, α-farnesene, and selina-4(14),7(11)-dien-8-one had an affinity toward GluN1, GluN2B, and Glu2A receptor subunits surpassed that of the reference compound ketamine. Overall, Atractylodes lancea (Thunb.) DC and Chrysanthemum morifolium Ramat essential oils are worthy of further research for fast-acting antidepressants through interactions with glutamate receptors, and their main compounds (atractylon, α-curcumene, α-farnesene, and selina-4(14),7(11)-dien-8-one) are predicted to underlie the fast-acting effect.
Keywords:
depression; anxiety; rapid-acting effect; essential oil; glutamate; neurotoxicity; neuroinflammation
1. Introduction
Depression is a complex psychiatric disorder, characterized by fatigue, constant feelings of sadness, and loss of pleasure [1]. Statistics show that depression remains a major burden in society, affecting approximately 280 million people worldwide [2]. Several pathological mechanisms of depression have been documented, including alterations in neurotransmitter systems, hypothalamus–pituitary–adrenal (HPA) axis activity, neuroinflammation, and changes in brain structures [3]. Norepinephrine and dopamine are neurotransmitters that play a role in reward processing and motivation, and their abnormal serum levels have been reported in depressed rodents [4]. Although serotonin has been the most extensively studied neurotransmitter in depression, researchers recently offer compelling data rejecting the relationship between serotonin activity to depression [5]. Stress-induced biological alterations, therefore, are receiving more attention as significant contributors to depression [6]. The HPA axis is a complex system that regulates the body’s stress response. Dysregulation of the HPA axis and the resulting elevations in corticotrophin-releasing hormone, adrenocorticotropic hormone, and cortisol or corticosterone (CORT) levels have been proposed as key mechanisms underlying the development of depression [7,8]. A dysregulated HPA axis activity is proposed to be accompanied by the overproduction of pro-inflammatory cytokines such as tumor necrosis factor alpha (TNF-α), interleukin (IL)-6, or IL-1β, which leads to the loss of hippocampal neurogenesis and promotion of depression-like behaviors [9,10]. In addition, chronic exposure to glucocorticoids such as cortisol or CORT can lead to dysfunction prefrontal cortex or atrophy of the hippocampus and structural deficits in the dentate gyrus area, which are all brain regions responsible for mood regulation [11,12]. In clinical practice, classical antidepressants exert their effects by inhibiting neurotransmitter-degrading enzymes (monoamine oxidase inhibitors) or inhibiting neurotransmitter reabsorption (selective serotonin reuptake inhibitors (SSRIs), selective serotonin and noradrenaline reuptake inhibitors, and tricyclic antidepressants) [13]. However, safety concerns have been raised, with side effects including weight gain, constipation, drowsiness, or even lethal hypertension [14,15]. Furthermore, the delayed onset of antidepressant action is another major limitation. For instance, acute treatment with SSRIs was found to initially elevate serotonin levels only in the cell body and dendrites, not in axons, which then immediately inhibit serotonin neuronal firing via an action at 5HT1A somatodendritic autoreceptors. In long-term treatment, SSRIs can cause a desensitization of 5HT1A autoreceptors, increasing the firing rate of neurons and serotonin release at axon terminals to postsynaptic receptors [16]. This partly explains the slow onset response of existing antidepressants, which usually take two to three weeks to manifest their effects [17]. Therefore, these limitations lead to an urgent need for safer agents with rapid antidepressant action.
In recent years, there has been a growing interest in research using essential oils due to their purported ability to alleviate a wide range of health issues, including inflammation, cancer, insomnia, anxiety, and depression with fewer side effects [18]. Essential oil is a mixture of secondary metabolites derived from plants [19]. Over 60 components have been identified in essential oils, with major compounds being benzenoids, phenylpropanoids, and terpenoids [20]. Their small, lipophilic components can rapidly and easily penetrate the blood–brain barrier (BBB) to access brain tissues, interacting with the thalamus, cerebral cortex, and limbic system, suggesting their potential use in rapidly reducing the symptoms of anxiety and depression [21,22]. For instance, lavender and citrus essential oil exerted anxiolytic-like and antidepressant-like effects in rodent models by restoring the decrease of monoamine neurotransmitter levels with downregulation of BDNF in serum or in the hippocampus [23,24,25].
Intranasal administration has been emphasized as a noninvasive method for the rapid management of neuropsychiatric disorders [26]. Intranasal agents directly stimulate the olfactory and trigeminal chemoreceptors, further enhancing the production of neurotransmitters and regulating the neuroendocrine system. In addition, mucosal epithelial pathways with blood vessel-dense nasal mucosa also contribute to rapid substance absorption and subsequent systemic effects, avoiding the hepatic first-pass effect and thus improving drug bioavailability [26,27]. Intranasal delivery of berberine, curcumin, and genipin using a thermosensitive hydrogel system has been reported to improve depressant-like activities in rodent models by enhancing monoamine neurotransmitter concentrations in the hippocampus and striatum. Additionally, at a lower dosage, these treatments exerted superior effects compared with intragastric or intraperitoneal routes [28,29,30].
Ketamine is a widely known medication that can rapidly alleviate the symptoms of depression through interacting with N-methyl-D-aspartate receptors (NMDARs), including GluN1 and GluN2B receptor subunits [31]. Notably, the GluN2B subunit is involved in many neurological disorders, and therefore significantly responsible for the biophysical and pharmacological properties of the NMDARs [32]. Ifenprodil, a GluN2B-selective NMDAR antagonist, was also found to exert a rapid-acting antidepression-like effect, compared with traditional medications that regulate the monoaminergic system [33]. In addition to NMDARs, the role of alpha-amino-3-hydroxy-5-methyl-4-isoxazole propionic acid receptors (AMPARs) including GluA1 and GluA2 subunits was also revealed in the rapid and long-lasting effects of antidepressants, for instance, ketamine and TAK-653 that potentiate AMPAR activity in maintaining synaptic plasticity [34,35,36]. Therefore, studies on the rapid action of new antidepressants should consider mechanisms via NMDARs and AMPARs.
Against this background, the present study was performed to identify and recommend essential oils that might have a rapid antidepression-like effect. We examined the in vitro neuroprotective and anti-neuroinflammatory effects of 20 essential oils to investigate their potential antidepression-like effects. Potentially antidepressant essential oils were then evaluated via intranasal administration in ICR mice using the tail suspension test (TST) and elevated plus maze test (EPM). Molecular docking was subsequently performed to predict the interactions between the major compounds in the effective essential oils and NMDAR and AMPAR subunits.
2. Materials and Methods
2.1. Preparation of Essential Oils
Twenty herbs were purchased from Omni Herb (Daegu, Republic of Korea), and each herbal material was immersed in distilled water (1:10 w/w) and hydrodistilled for 4 h using a steam distillation solvent extraction (SDE) apparatus, and the distilled oil was captured in n-hexane (Chemicals Duksan Corp., Ansan, Gyeonggi, Republic of Korea). The oil was dehydrated using a separating funnel, and n-hexane was evaporated. The essential oil was weighed (Thuja orientalis L. (0.28% w/w), Acorus gramineus Sol. (1.79%), Foeniculum vulgare Mill. (1.09%), Magnolia biondii Pamp. (0.55%), Ligusticum striatum DC. (0.46%), Prunella vulgaris L. (0.03%), Chrysanthemum morifolium Ramat. (0.28%), Cinnamomum cassia (Nees & T.Nees) J.Presl (0.75%), Zingiber officinale Roscoe (0.11%), Santalum album L. (0.39%), Nardostachys jatamansi (D. Don) DC. (1.38%), Angelica acutiloba (Siebold & Zucc.) Kitag. (0.20%), Aucklandia lappa DC. (0.21%), Mentha arvensis L. (yield 0.73%), Perilla frutescens (L.) Britton (0.39%), Syzygium aromaticum (L.) Merr. & L.M.Perry (6.75%), Citrus reticulata Blanco (0.41%), Atractylodes lancea (Thunb.) DC. (0.11%), Agastache rugosa (Fisch. & C.A.Mey.) Kuntze (0.20%), Myristica fragrans Houtt. (0.87%)) and stored at −20 °C until further use.
2.2. Animal Experiments and Treatments
Five-week-old male ICR outbred mice weighing 25–29 g were obtained from Koatech Lab Animal Inc. (Pyeongtaek, Gyeonggi, Republic of Korea) and acclimated for one week before the behavioral test. All mice had free access to water and a commercial pellet diet (5L79; PMI Nutrition, St. Louis, MO, USA). All experimental animal procedures were approved by the Institutional Animal Care and Use Committee of Dongguk University (IACUC-2021-15). The mice were randomly divided into the following groups: control group (CON), essential oil treatment groups (EO), and memantine group (MEM), which were offered vehicle (saline and Tween 80, 3% v/v), essential oil (25 mg/kg), and memantine (3 mg/kg), respectively. On the day of the experiment, all animals were habituated to the test room for two hours prior to receiving a 10 µL intranasal administration (CON and EO groups) or 200 µL intraperitoneal injection (MEM group). Thirty minutes after the treatment, a behavioral test was performed.
2.3. Tail Suspension Test (TST)
The effect of essential oils on depression-like behaviors in mice using the TST test was based on the fact that rodents subjected to the short-term, inescapable stress of being suspended by their tail will develop an immobile posture [37]. The tail of the mouse was attached to a bar using 15 cm adhesive tape, and the distance from the bar to the ground was fixed at 50 cm. All 6 min trials were recorded, and the total mobility time of each mouse was measured by manual scoring. The strong shaking of the body and movement of the four limbs akin to running were counted as mobility, whereas small movements that were confined to the front legs but without the involvement of the hind legs were not. The total mobility time was then subtracted from the 360 s of test time and was then stated as the immobility time. After each trial, the suspension box was wiped with 70% ethanol to remove unwanted odors.
2.4. Elevated Plus Maze Test (EPM)
The EPM test, which is based on the innate tendency of rodents to avoid elevated and open spaces and explore novel environments, was performed to assess the anxiety-like behaviors of mice after essential oil treatment [38]. The equipment consisted of four 29 cm × 5 cm arms elevated 40 cm above the ground, with two “closed arms” enclosed by 14 cm black walls. Each mouse was placed in the central area of the maze for a 5-min free exploration period. The arms were cleaned with 70% ethanol after every trial to remove unwanted odors, urine, and feces. The distance traveled, number of entries into, and time spent in the open arms were recorded and analyzed using Smart V3.0 software (Panlab Harvard Apparatus, Holliston, MA, USA).
2.5. Cell Culture and Treatments
PC12 cells (a rat adrenal medullary pheochromocytoma cell line) were cultured in Roswell Park Memorial Institute (RPMI) medium (Welgene Inc., Gyeongsan, Gyeongsangbuk-do, Republic of Korea) supplemented with 10% fetal bovine serum (FBS) (Merck KGaA, Darmstadt, Germany) and 1% penicillin-streptomycin (Thermo Fisher Scientific, Waltham, MA, USA) at 37 °C in a 5% CO2 humidified environment. PC12 cells were incubated with the SDE extract of each essential oil (0.1 or 1 μg/mL) for 1 h and then stimulated with corticosterone (CORT) (200 μM) for 24 h.
BV2 cells (an immortalized mouse microglial cell line) were cultured in Dulbecco’s Modified Eagle Medium (DMEM) supplemented with 10% FBS (Merck KGaA, Darmstadt, Germany) and 1% penicillin-streptomycin (Thermo Fisher Scientific, Waltham, MA, USA) at 37 °C in a 5% CO2 humidified environment. For the cell viability assay, BV2 cells were treated with the SDE extract of each essential oil (0.1 or 1 μg/mL). To test the anti-neuroinflammatory effects of the essential oils, BV2 cells were pretreated with non-toxic doses of each herb extract for 6 h and then stimulated with lipopolysaccharide (LPS) (1 μg/mL) for 18 h.
2.6. Water-Soluble Tetrazolium Salt Assay (WST)
The effects of the essential oils on the viability of PC12 and BV2 cells were examined using the EZ-Cytox assay kit (DoGenBio, Seoul, Republic of Korea). Cells were seeded at a density of 5 × 104 cells/100 µL/well in 96-well plates and incubated at 37 °C and 5% CO2 for 24 h. After treating the cells and incubating for 24 h, 10 μL of the WST solution was added to each well, and the plates were incubated for 3–4 h. Absorbances were measured at a detection wavelength of 450 nm and a reference wavelength of 650 nm using a Tecan microplate reader (Männedorf, Switzerland).
2.7. Lactate Dehydrogenase Assay (LDH)
LDH released into the culture supernatant was measured using an EZ-LDH assay kit (DoGenBio, Seoul, Republic of Korea). PC12 cells were seeded and incubated under the same conditions as those used for the WST assay. After 24 h of treatment, the cell culture plates were centrifuged at 600× g for 5 min. Subsequently, 10 µL of supernatant from each well was transferred to a new 96-well plate and mixed with 100 μL of LDH reaction mixture for 30 min in darkness. LDH levels in the culture medium were determined by measuring the absorbance at 450 nm (reference wavelength, 650 nm) using a Tecan microplate reader (Männedorf, Switzerland).
2.8. Enzyme-Linked Immunosorbent Assay (ELISA)
The levels of inflammatory cytokines TNF-α and IL-6 secreted by BV2 cells in the culture media were measured to evaluate the anti-inflammatory activity of the essential oils. The cells were seeded at a density of 2 × 105 cells/1 mL/well in 12-well plates and incubated at 37 °C and 5% CO2 for one day. 24 h after treatments, cell culture media were collected and centrifuged at 1500× g rpm for 10 min at 4 °C to remove particulates. The next steps were performed using ELISA kits (LABISKOMA, Seoul, Republic of Korea) following the manufacturer’s protocols. The absorbance at 450 nm was measured to detect cytokine levels, using a Tecan microplate reader (Männedorf, Switzerland).
2.9. Molecular Docking
The crystal structures of the GluN1 and GluN2B-NMDAR subunits and GluA2-AMPAR subunit (PDB ID:5H8Q, 5EWM, and 5ZG3, respectively) were downloaded from the Protein Data Bank (RCSB PDB) as previously described [39,40,41]. Then, all the heteroatoms and water molecules of the proteins were removed. Finally, Kollman charges were added to the proteins, and the macromolecules were exported into a PDBQT format for molecular docking. Information about the main compounds of essential oils was collected from the PubMed database and Traditional Chinese Medicine Systems Pharmacology Database and Analysis Platform (TCMSP). Only compounds with a molecular weight ≤ 500 Da and blood–brain barrier index ≥ 0.3 were selected. The 3D structures of the compounds were retrieved from the PubChem database. All ligands were then converted into PDBQT format using AutoDockTools version 1.5.6 (The Scripps Research Institute, San Diego, CA, USA).
Molecular docking of GluN1, GluN2B, and GluA2 was performed using AutoDock Vina, version 1.2.0 (The Scripps Research Institute, San Diego, CA, USA). According to the native ligand, the grid boxes that covered the active sites of GluN1, GluN2B, and GluA2 were defined using the following parameters: center_x = 14.66, center_y = −14.25, center_z = −25.74, size_x = 30, size_y = 30, and size_z = 30; center_x = 81.00, center_y = 5.90, center_z = −32.42, size_x = 30, size_y = 30, and size_z = 30; and center_x = 33.47, center_y = −55.89, center_z = 19.58, size_x = 30, size_y = 30, and size_z = 30, respectively. Before the docking study was performed, the docking parameters and algorithm were validated by re-docking the native ligand to the target receptor. The re-docked conformation was then superimposed onto the co-crystallized one using Discovery Studio Visualizer 2021 (Dassault Systèmes BIOVIA, San Diego, CA, USA) and the root mean square deviation (RMSD) was calculated. An RMSD lower than 2 Å suggested that the method could consistently predict the natural conformation of the ligand receptor [42,43].
In this study, ketamine, ifenprodil, and TAK-653, allosteric NMDAR antagonists and an allosteric AMPAR potentiator, respectively, were selected as reference compounds to check whether they strongly interacted with proteins inside selected binding pockets. Within the same grid boxes, herbal compounds with good protein affinity were suggested to exert similar effects as the reference compounds. Favorable conformations were selected based on the lowest binding energy. Finally, Discovery Studio Visualizer 2021 was used to visualize the molecular interactions between proteins and ligands.
2.10. Statistical Analysis
All experiments were performed with at least three independent experiments. GraphPad Prism 8.0 (GraphPad Software, San Diego, CA, USA) was used for statistical analysis. The results are presented as means ± standard deviations (SDs) followed by statistical significance (two-tailed unpaired Student’s t-test) defined as a p-value < 0.05.
3. Results
3.1. Effects of Essential Oils on CORT-Induced Neurotoxicity in PC12 Cells
To determine the appropriate concentration of CORT for inducing cell damage, PC12 cells were incubated for 24 h with 100, 200, 300, and 400 µM CORT. Exposure to CORT reduced survival in a dose-dependent manner by 25, 38, 70, and 85%, respectively (Figure 1A). The cell viability of the 200 µM CORT-treated group decreased to about half that of the control group; this concentration was therefore used in subsequent experiments to induce neurotoxicity.
The effects of 0.1 and 1.0 µg/mL of each essential oil on the viability of PC12 cells were also assessed. As shown in Figure 1B, all essential oils did not cause toxicity to PC12 cells at doses of 0.1 and 1.0 µg/mL; these doses were then used to treat the cells for 1 h prior to incubation with CORT for 24 h. As shown in Figure 2A, compared with the control group, the cell survival rate significantly decreased in the model group treated with 200 µM CORT (p < 0.05). Foeniculum vulgare Mill., Chrysanthemum morifolium Ramat., Angelica acutiloba (Siebold & Zucc.) Kitag., Aucklandia lappa DC., Syzygium aromaticum (L.) Merr. & L.M. Perry, Citrus reticulata Blanco, Atractylodes lancea (Thunb.) DC., and Myristica fragrans Houtt. essential oils showed a dose-response relationship, while Acorus gramineus Sol. did not exhibit a protective effect at either of the two doses, and the remaining 11 essential oils exerted an effect only at the higher dose.
As LDH leakage from cells is widely used as a cellular damage marker, LDH release was measured in the culture medium to assess PC12 cell injury. LDH secreted from cells was significantly increased in the model group compared to that in the control group; the percentage of LDH leakage increased from 100% (control) to 288 ± 6.9%. However, pretreatment with essential oils reduced LDH leakage (Figure 2B), which was consistent with the data from the WST assay. These results indicated that essential oil pretreatment could prevent CORT-induced injury in PC12 cells.
3.2. Effects of Essential Oils on LPS-Induced Neuroinflammation in BV2 Cells
To test whether essential oil treatments were toxic to the viability of BV2 cells, a WST assay was performed using concentrations of 0.1 and 1.0 µg/mL of each essential oil. From the results, all essential oils did not cause toxicity to BV2 cells at doses of 0.1 and 1.0 µg/mL. Although Thuja orientalis L. at 1.0 µg/mL decreased cell viability to approximately 75%, this reduction was not statistically significant (Figure 3). Hence, doses of 0.1 and 1.0 µg/mL of all essential oils were used for the subsequent experiments.
As shown in Figure 4, LPS (1 µg/mL) remarkably increased the production of the inflammatory cytokines TNF-α and IL-6 compared to that in the control BV2 cells, but this effect was blunted by pretreatment with essential oils at different levels. In particular, 10 essential oils (Ligusticum striatum DC., Cinnamomum cassia (Nees & T.Nees) J.Presl, Santalum album L., Aucklandia lappa DC., Mentha arvensis L., Perilla frutescens (L.) Britton, Syzygium aromaticum (L.) Merr. & L.M. Perry, Citrus reticulata Blanco, Atractylodes lancea (Thunb.) DC., and Myristica fragrans Houtt.) reduced IL-6 release, yet only six essential oils (Foeniculum vulgare Mill., Ligusticum striatum DC., Prunella vulgaris L., Chrysanthemum morifolium Ramat., Cinnamomum cassia (Nees & T.Nees) J.Presl, and Santalum album L.) abrogated the increase in TNF-α levels (Figure 4). Of the 20 examined essential oils, eight including Chrysanthemum morifolium Ramat., Cinnamomum cassia (Nees & T.Nees) J.Presl, Santalum album L., Perilla frutescens (L.) Britton, Syzygium aromaticum (L.) Merr. & L.M. Perry, Citrus reticulata Blanco, Atractylodes lancea (Thunb.) DC., and Myristica fragrans Houtt. suppressed TNF-α or IL-6 production at a low dose of 0.1 µg/mL. Moreover, three essential oils (Ligusticum striatum DC., Cinnamomum cassia (Nees & T.Nees) J.Presl, and Santalum album L.) significantly decreased the levels of both the assessed inflammatory cytokines (Figure 4). At the investigated doses, no beneficial effects were observed for Thuja orientalis L., Acorus gramineus Sol., Magnolia biondii Pamp., Zingiber officinale Roscoe, Nardostachys jatamansi (D. Don) DC., Angelica acutiloba (Siebold & Zucc.) Kitag., and Agastache rugosa (Fisch. & C.A.Mey.) Kuntze.
3.3. Rapid-Acting Effect of Essential Oils on Behavior Changes of Mice in the TST and EPM
Based on the in vitro results, 19 potential essential oils were used for the in vivo experiments. Thirty minutes after intranasal administration, the TST and EPM were conducted to investigate the effects of essential oils on the behavior of mice. During the TST, Chrysanthemum morifolium Ramat., Zingiber officinale Roscoe, Santalum album L., Citrus reticulata Blanco, Atractylodes lancea (Thunb.) DC., and Myristica fragrans Houtt. essential oil treatments exhibited antidepressant-like effects by decreasing the immobility time compared with the control mice (Figure 5). There was a reduction in the immobility time of mice in the TST after memantine treatment, while no statistically significant change was observed in the EPM (Figure 5 and Figure 6). As shown in Figure 6, both Chrysanthemum morifolium Ramat. and Myristica fragrans Houtt. essential oils enhanced the percentage of distance covered in the open arms in the EPM test. In addition, Myristica fragrans Houtt. significantly increased the number of open arm entries, while Chrysanthemum morifolium Ramat. treatment increased the time of mice exploring the open arms, indicating antianxiety-like effects. In contrast, Magnolia biondii Pamp. exerted the opposite effect, significantly reducing exploration time in the open arms.
3.4. Molecular Docking Analysis of Essential Oil Main Compounds
Molecular docking was conducted to investigate the five major compounds of the seven essential oils that modified depressant- or anxiety-related behaviors in animal tests (Table 1). After removing duplicates, 27 compounds remained (Table 2). The docking protocol was validated through a re-docking experiment using the native ligand. Root mean square deviation (RMSD) values of less than 2 Å were observed, suggesting that the ligand-receptor conformations had a high docking accuracy (Figure 7). The molecular docking of compounds against the GluN1, GluN2B, and GluA2 proteins obtained using Autodock Vina are presented in Table 3, with higher negative binding energies indicating better affinity between components. Among the 27 natural compounds, 24, 22, and 13 had docking scores of less than −6.0 kcal/mol toward GluN1, GluN2B, and GluA2, respectively, suggesting a highly stable complex. When compared with the reference compounds, four compounds, including atractylon, α-curcumene, α-farnesene, and selina-4(14),7(11)-dien-8-one, had lower energy binding than ketamine when interacting with all three proteins, while none had lower docking scores than ifenprodil and TAK-653. The 2D conformations are visualized in Figure 8, Figure 9 and Figure 10.
4. Discussion
Side effects and delayed onset of action are the limitations of current antidepressants. Ketamine was reported to alleviate depressive symptoms within hours post-treatment, but rapid-acting antidepressants have not yet made any other striking progress [65,66]. Essential oils possessing antianxiety and antidepressant properties with fewer side effects have been reported [67]. They mainly consist of small and lipophilic molecules, and therefore can quickly and easily bypass the BBB to target brain tissues, acting on the cerebral cortex, thalamus, and limbic system, indicating their potential application in alleviating symptoms of anxiety and depression in a rapid manner [21,22]. Additionally, recent studies have revealed the advantages of intranasal administration over other routes of administration [26]. Hence, the purpose of this study was to identify essential oils with rapid-acting antidepressant effects when administered intranasally. As expected, our results suggested that different essential oils exerted neuroprotective effects in vitro and rapid antidepressant-like effects in a normal mouse model (Table 4). In addition, the effects of essential oils were comparable to those of MEM, a common medication used to treat moderate-to-severe dementia, whose acute antidepressant-like effects have also been reported in previous studies [68,69].
Neuroinflammation is a common feature in patients with depression. Several studies have reported that inflammation in the brain and the associated over-release of pro-inflammatory cytokines TNF-α, IL-6, or IL-1β can lead to a loss of hippocampal neurogenesis, promoting depression-like behaviors [10]. The underlying mechanism may be related to microglial cells, which influence neuronal excitability and neurotransmission [70]. Under threats such as the LPS challenge, microglia are activated to undergo morphological changes and secrete pro-inflammatory cytokines [71]. These cytokines can decrease the levels of serotonin and norepinephrine, which are neurotransmitters involved in mood regulation [72]. In addition, pro-inflammatory cytokines can affect the function of the hippocampus, a brain region that is important for learning, memory, and mood regulation [12]. In the present study, we treated the murine microglial cell line BV2 with LPS and essential oils, with the hypothesis that essential oils that could reduce LPS-induced pro-inflammatory cytokines would have potential antidepressant effects. Essential oils such as Chrysanthemum morifolium Ramat., Santalum album L., Citrus reticulata Blanco, and Atractylodes lancea (Thunb.) DC., which inhibited TNF-α or IL-6 released from LPS-treated BV2 cells, significantly decreased the immobility time of mice in TST, suggesting their antidepressant effects occurred through inactivating glial cells and reducing neuroinflammation.
Depression is also accompanied by the overproduction of CORT, which affects numerous physiological processes in neurons and astrocytes, such as apoptosis and synaptic plasticity [73]. High concentrations of CORT can increase the production of reactive oxygen species (ROS) and pro-inflammatory cytokines, which can damage neuronal cells [74,75]. Additionally, through activating glucocorticoid receptors (GRs) in neurons, CORT can cause a cascade of apoptosis-related enzymes known as caspases that are responsible for initiating programmed cell death [76]. GRs are mainly distributed in the hippocampus, and CORT-induced GR overexpression can lead to structural changes such as hippocampal atrophy, loss of dendritic synapses, and synaptic plasticity alternation, resulting in the development of anxiety and depression [77,78,79,80,81]. In this study, PC12 cells, a classical neuronal cell line, were treated with CORT to mimic the features of depression, and essential oils that protect cells from CORT-induced toxicity may have the potential to treat depression. PC12 cell viability was evaluated using a WST assay based on mitochondrial succinate dehydrogenase activity, indicative of cellular energy capacity, compared to LDH leakage, indicative of apoptosis [82]. Our results showed that 19 of the 20 essential oils included in this study inhibited CORT-induced cell death and LDH leakage, indicating their potential in the treatment of depression, probably by decreasing neuronal apoptosis. In addition, a very small dose of essential oil (SDE extract), including 0.1 or 1 g/mL, was sufficient to exhibit anti-inflammatory and neuroprotective effects. Previous studies using ethanol or methanol extracts of herbal materials normally required an effective dose of 10–100 g/mL. This demonstrates one benefit of essential oils in drug development applications [83,84,85,86].
The TST is an important behavioral model that is widely used to screen for new antidepressants. In animals subjected to inescapable stress, immobility has been suggested as a key indicator of behavioral despair, reminiscent of depressive disorders in humans [37]. In the present study, a significant decrease in immobility time in treated mice was observed with six essential oils only 30 min after intranasal administration, indicating their fast antidepressant-like activity. This is the first time the rapid antidepressant effects of Chrysanthemum morifolium Ramat., Zingiber officinale Roscoe, Santalum album L., Citrus reticulata Blanco, Atractylodes lancea (Thunb.) DC., and Myristica fragrans Houtt. essential oils have been reported, particularly through intranasal administration. Although the antidepressant activity of nutmeg (Myristica fragrans) seeds has been reported in previous studies, its effect was observed after oral administration for three days [87,88].
The EPM is an extensively used model for the assessment of novel anxiolytic agents, based on the natural tendency of rodents to avoid open and elevated areas [38]. Because high levels of anxiety result in depression, and that approximately 85% of patients with depression have significant anxiety, anxiety is considered one of the symptoms of depression [89,90]. Among the two effective essential oils, Chrysanthemum morifolium Ramat. increased the time mice spent in the open arms, whereas Myristica fragrans Houtt. increased open-arm entries. Although no essential oils significantly elevated either parameter, increasing trends were observed. Moreover, mice treated with the two essential oils also showed notably higher total distances in the open arms than that demonstrated in the control group. Therefore, these essential oils might have exerted anti-anxiety effects to a certain extent, as evidenced by an increased exploration in the open elevated space, which contributed to the reduction of depression-like behaviors.
Ketamine and ifenprodil, an NMDAR antagonist and a GluN2B-selective NMDAR antagonist, respectively, have been proven to be rapid-acting antidepressant therapies in clinical and in vivo studies. Their mechanisms may be related to the de-inhibition of mTOR activity, which induces protein biosynthesis, leading to synaptic potentiation [33,91]. In addition, by blocking spontaneous NMDAR-mediated neurotransmission, ketamine can suppress eukaryotic elongation factor 2 (eEF2) kinase function, preventing eEF2 substrate phosphorylation and enhancing BDNF translation. Ketamine can also upregulate AMPAR, especially the GluA1 and GluA2 subunits, thereby enhancing synaptic strengthening and transmission [91,92]. TAK-653, an AMPAR potentiator, exerts antidepressant effects in vivo. It can also directly activate AMPAR, stimulating the mammalian target of rapamycin (mTOR) and BDNF signaling in vitro [36]. In 2015, a study revealed that ketamine conferred fast antidepressant effects in mice 30 min after intraperitoneal injection, favorably suppressing NMDAR subunits GluN1 and GluN2B functioning in the hippocampus [31]. Additionally, Li et al. suggested that ketamine exerts antidepressant effects by enhancing AMPAR expression [34]. Indeed, current concepts regarding glutamate receptors suggest that drugs that increase AMPAR signaling or decrease NMDAR function may be effective antidepressants. Therefore, we performed a molecular docking analysis of the main compounds from the essential oils toward GluN1, GluN2B, and GluA2 proteins using ketamine, ifenprodil, and TAK-653 as reference compounds.
Molecular docking is a computational technique that predicts the binding orientation and affinity of small molecules (compounds) to a target protein or macromolecule (receptor subunits). It provides insights into the mechanisms of drug action and allows researchers to identify potential drug candidates and optimize their binding properties before conducting costly and time-consuming experiments [93,94]. Besides, through simulating the binding interactions between molecules and targets, the technique may also suggest whether a candidate drug has similar actions to other reference compounds. Here, we showed that in comparison with ketamine (with docking scores of −8.2, −5.9, and −4.9 kcal/mol toward GluN1, GluN2B, and GluA2, respectively), four compounds including atractylon, α-curcumene, α-farnesene, and selina-4(14),7(11)-dien-8-one had much lower docking scores, indicating stronger binding to the targeted receptors. Hence, these compounds that are main components of Atractylodes lancea (Thunb.) DC. and Chrysanthemum morifolium Ramat. essential oils are of greater concern.
Atractylodes lancea (Thunb.) DC. is a type of Atractylodis Rhizoma, whose extract has been commonly used for the treatment of digestive disorders owing to its anti-inflammatory and gastroprotective effects [59]. Atractylodes lancea (Thunb.) DC. may also have the potential to treat depression. A previous study suggested that drugs inhibiting gastric acid secretion can be used to treat depressive symptoms and antidepressants can be an effective treatment for stress ulcers, indicating a similar mechanism of pathogenesis shared by the two ailments [95]. This is consistent with our results showing that Atractylodes lancea (Thunb.) DC. essential oil significantly reduced depression-like behaviors in mice in the TST. Furthermore, atractylon, one of the major constituents of this essential oil, was recently found to improve cognitive dysfunction in mice by inhibiting microglial activation and exerting anti-inflammatory activity in vitro through the inhibition of cyclooxygenase-2 (COX-2) and inducible nitric oxide synthase (iNOS) expression [96,97]. Such an action has been previously revealed to curb depressive symptoms by reducing neuroinflammation [12,72]. Compared with ketamine, atractylon has a lower binding energy for the three glutamate receptor subunits, indicating a stronger binding capacity than ketamine. Notably, atractylon can interact with GluN1 and GluA2 subunits through the same amino acids as that of ketamine, suggesting a similar mechanism of action for its rapid-acting antidepressant effect (Figure 8 and Figure 10). Regarding selina-4(14),7(11)-dien-8-one, also a major component of Atractylodes lancea (Thunb.) DC., the pharmacological activity of this compound has not been reported to date. However, based on the molecular docking results, this compound was able to interact with the GluA2 subunit at sites similar to those of ketamine and ifenprodil (Figure 10). Hence, selina-4(14),7(11)-dien-8-one is also a compound with potential for the research and development of antidepressants.
Chrysanthemum morifolium Ramat. is a traditional herb widely used for the treatment of fever, headache, sore throat, and hypertension, due to the variety of flavonoids, anthocyanins, alkaloids, and phenolic acid components found in its extract [98]. Notably, neuroprotective and antioxidant effects, as well as its ability to reduce ROS levels and lipid peroxidation, have also been reported [99,100]. ROS and oxidative stress are known to cause dysfunction in neurotransmission and the HPA axis, reduced neuroplasticity, and neuroinflammation, all of which are involved in the pathogenesis of depression [101]. Hence, the antidepressant potential of Chrysanthemum morifolium Ramat. should not be overlooked. However, until now, only one previous study revealed the chronic antidepressant effect of Chrysanthemum morifolium on CORT-injected C57BL/6 mice, evidenced by significantly elevated sucrose consumption and serum serotonin levels [102]. Our results showed that Chrysanthemum morifolium Ramat. essential oil improved both anxiety-like and depression-like symptoms after only a single dose, suggesting an acute effect on depression. We predict that such an effect may originate from the two main compounds of the essential oil, α-curcumene, and α-farnesene, although only the anti-inflammatory effect of α-farnesene has been reported to date [103]. We found that the interaction of these two compounds with the glutamate receptor subunits were comparable to that of reference compounds. α-Curcumene and α-farnesene have binding energies for the three subunits surpassing that of ketamine, interact with GluN1 and GluN2B at many amino acids, similar to ketamine and ifenprodil, and form more bonds with the proteins than ketamine. This suggests that α-curcumene and α-farnesene not only bind well to NMDAR subunits but can also mimic the mechanisms by which ketamine and ifenprodil improve depression.
Myristica fragrans Houtt. or nutmeg is a well-characterized herb with various pharmacological properties such as antidiarrheal, antimicrobial, antifungal, antioxidant, cardioprotective, especially sedative and antidepressant effects [104]. However, to our best knowledge, this study is the first to report the neuroprotective effect in vitro as well as the rapid-acting antianxiety-like and antidepression-like effects in vivo of Myristica fragrans Houtt. essential oil via intranasal delivery. Unlike Atractylodes lancea (Thunb.) DC. and Chrysanthemum morifolium Ramat., fast actions of Myristica fragrans Houtt. appeared not to mainly exert via glutamate receptors since its main compounds such as limonene, β-myrcene, α-pinene, and β-pinene do not have a strong affinity towards GluN1 and GluA2 subunits. It should be noticed that not only glutamate receptors drive the rapid anxiolytic and antidepressant effects, but others might also do that function, for instance, gamma-aminobutyric acid B, or 5-hydroxytryptamine receptor 4 receptors [105,106,107]. A previous study showed that 3-day treatments of Myristica fragrans n-hexane extract reduced depression-like behaviors in mice by targeting the serotonergic and noradrenergic nervous systems [87,88]. Short-term inhalation of limonene has been found to restore chronic unpredictable mild stress-induced depression-like behavior by modulating the activity of the HPA axis, BDNF receptors, and monoamine neurotransmitter levels [23]. The acute anxiolytic effect of inhaled α-pinene after one day of treatment has also been reported, yet its underlying mechanism remains to be studied [108]. In short, though Myristica fragrans Houtt. essential oil is a potential material for rapid-acting antidepressant development, other mechanisms of action rather than glutamate pathway should be considered.
Notably, the rapid antidepressant-like effect of essential oils via intranasal administration is a novelty of this study. Nasal delivery has recently been shown to have advantages over other routes, such as oral administration or intraperitoneal injection. Since intranasal intervention directly transports exogenous materials from the nasal cavity to the brain, avoiding first-pass metabolism in the liver, it results in a fast onset of action and higher bioavailability of drugs [109,110]. In our study, intranasal administration of essential oils only 30 min prior to the behavioral test produced positive changes in the behavior of mice. We postulate that the olfactory bulb receives sensory information from the olfactory receptors in the lining of the nasal cavity and projects signals to certain brain regions, including the hippocampus and amygdala, which are involved in emotional processing and linked to the regulation of anxiety and depression [21,111]. In addition, compared to previous work, with the same herbal material but different forms of extract, essential oils delivered intranasally required a very small dosage but exerted their effects within a shorter time course [102]. Given these advantages, the use of essential oils for antidepressant development and its intranasal delivery are promising strategies and should be the focus of future research.
This study has some limitations that could be addressed in future research. First, the safety assessment of essential oils in mice administered intranasally was overlooked. Nasal delivery has been reported to be associated with irritation and dryness of the nasal mucosa, resulting in cracking, inflammation, and nosebleeds [112,113]. Hence, toxicity evaluation for the nasal mucosa and olfactory tissues is necessary before intranasal administration. The second limitation of this study was the tested dose (25 mg/kg), which was administered to mice for all 20 essential oils. Although we chose the dose based on previous studies [114,115,116,117], it should be noted that each essential oil has its own properties and characteristics; therefore, the effective doses and pathways in which they are absorbed or distributed are different. However, finding the right dose for each essential oil, including those that had never previously been studied or published, was relatively unfeasible. Thirdly, the compounds described in the molecular docking experiment were derived from a literature search, not a GC-MS analysis from our own samples. GC-MS analysis would be more appropriate for future in-depth research on each essential oil suggested in this paper. Finally, the resolution of the protein structure determines the accuracy of docking, yet there are currently no high-resolution data on other AMPAR subunits; hence, molecular docking could only be performed for the GluA2 subunit.
5. Conclusions
Chrysanthemum morifolium Ramat., Zingiber officinale Roscoe, Santalum album L., Citrus reticulata Blanco, Atractylodes lancea (Thunb.) DC., and Myristica fragrans Houtt. essential oils have been shown to possess neuroprotective or anti-neuroinflammatory effects in vitro and to exert rapid-acting antidepressant effects through intranasal delivery in mice. Notably, Chrysanthemum morifolium Ramat. and Atractylodes lancea (Thunb.) DC. essential oils and their volatile compounds, including atractylon, α-curcumene, α-farnesene, and selina-4(14),7(11)-dien-8-one, may be useful candidates for development as rapid-acting intranasal antidepressants through interactions with NMDARs and AMPARs. It is important for future work to validate the safety of these essential oils and compounds for intranasal delivery and the exact mechanism underlying their rapid-acting effects against depression. Additionally, they should be studied using a specific animal disease model.
Author Contributions
Conceptualization, I.-J.Y., K.N.T. and N.P.K.N.; methodology, K.N.T. and N.P.K.N.; software, K.N.T. and N.P.K.N.; formal analysis, K.N.T. and N.P.K.N.; writing—original draft preparation, K.N.T. and N.P.K.N.; writing—review and editing, I.-J.Y., H.-M.S. and L.T.H.N.; supervision, I.-J.Y., H.-M.S. and L.T.H.N.; project administration, I.-J.Y.; funding acquisition, I.-J.Y. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Dongguk University Research Fund (2023) and the National Research Foundation (NRF-2021R1I1A2048979).
Institutional Review Board Statement
The study was conducted according to the guidelines of the Declaration of Helsinki and approved by the Institutional Review Board (or Ethics Committee) of the Institutional Animal Care and Use Committee of Dongguk University (protocol code IACUC-2021-15).
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available in this article.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design, execution, interpretation, or writing of the study.
References
- Chand, S.P.; Arif, H. Depression. In StatPearls; StatPearls Publishing LLC.: Treasure Island, FL, USA, 2023. Available online: https://www.ncbi.nlm.nih.gov/books/NBK430847/ (accessed on 7 April 2023).
- World Health Organization. Depression; World Health Organization: Geneva, Switzerland, 2021. Available online: https://www.who.int/news-room/fact-sheets/detail/depression (accessed on 7 April 2023).
- Peng, S.; Zhou, Y.; Lu, M.; Wang, Q. Review of Herbal Medicines for the Treatment of Depression. Nat. Prod. Commun. 2022, 17, 1934578X221139082. [Google Scholar] [CrossRef]
- Zhang, H.; Ren, Y.; Lv, M.; Xie, M.; Wang, K.; Yang, M.; Lv, C.; Li, X. Anti-depression effect and mechanism of Suanzaoren Decoction on mice with depression. IOP Conf. Ser. Earth Environ. Sci. 2021, 714, 022065. [Google Scholar] [CrossRef]
- Borgogna, N.C.; Aita, S.L. Is the serotonin hypothesis dead? If so, how will clinical psychology respond? Front. Psychol. 2022, 13, 1027375. [Google Scholar] [CrossRef] [PubMed]
- Moncrieff, J.; Cooper, R.E.; Stockmann, T.; Amendola, S.; Hengartner, M.P.; Horowitz, M.A. The serotonin theory of depression: A systematic umbrella review of the evidence. Mol. Psychiatry 2022. [Google Scholar] [CrossRef] [PubMed]
- Fiksdal, A.; Hanlin, L.; Kuras, Y.; Gianferante, D.; Chen, X.; Thoma, M.V.; Rohleder, N. Associations between symptoms of depression and anxiety and cortisol responses to and recovery from acute stress. Psychoneuroendocrinology 2019, 102, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Pariante, C.M.; Lightman, S.L. The HPA axis in major depression: Classical theories and new developments. Trends Neurosci. 2008, 31, 464–468. [Google Scholar] [CrossRef]
- Bendezú, J.J.; Calhoun, C.D.; Vinograd, M.; Patterson, M.W.; Rudolph, K.D.; Giletta, M.; Hastings, P.; Nock, M.K.; Slavich, G.M.; Prinstein, M.J. Exploring joint HPA-inflammatory stress response profiles in adolescent girls: Implications for developmental models of neuroendocrine dysregulation. Dev. Psychobiol. 2022, 64, e22247. [Google Scholar] [CrossRef]
- Troubat, R.; Barone, P.; Leman, S.; Desmidt, T.; Cressant, A.; Atanasova, B.; Brizard, B.; El Hage, W.; Surget, A.; Belzung, C.; et al. Neuroinflammation and depression: A review. Eur. J. Neurosci. 2021, 53, 151–171. [Google Scholar] [CrossRef]
- Mizoguchi, K.; Ishige, A.; Aburada, M.; Tabira, T. Chronic stress attenuates glucocorticoid negative feedback: Involvement of the prefrontal cortex and hippocampus. Neuroscience 2003, 119, 887–897. [Google Scholar] [CrossRef]
- Seguin, J.A.; Brennan, J.; Mangano, E.; Hayley, S. Proinflammatory cytokines differentially influence adult hippocampal cell proliferation depending upon the route and chronicity of administration. Neuropsychiatr. Dis. Treat. 2009, 5, 5–14. [Google Scholar] [PubMed]
- Feighner, J.P. Mechanism of action of antidepressant medications. J. Clin. Psychiatry 1999, 60 (Suppl. S4), 4–13. [Google Scholar] [PubMed]
- Wang, S.M.; Han, C.; Bahk, W.M.; Lee, S.J.; Patkar, A.A.; Masand, P.S.; Pae, C.U. Addressing the Side Effects of Contemporary Antidepressant Drugs: A Comprehensive Review. Chonnam Med. J. 2018, 54, 101–112. [Google Scholar] [CrossRef] [PubMed]
- Chamberlain, S.R.; Baldwin, D.S. Monoamine Oxidase Inhibitors (MAOIs) in Psychiatric Practice: How to Use them Safely and Effectively. CNS Drugs 2021, 35, 703–716. [Google Scholar] [CrossRef] [PubMed]
- Commons, K.G.; Linnros, S.E. Delayed Antidepressant Efficacy and the Desensitization Hypothesis. ACS Chem. Neurosci. 2019, 10, 3048–3052. [Google Scholar] [CrossRef] [PubMed]
- Machado-Vieira, R.; Salvadore, G.; Luckenbaugh, D.A.; Manji, H.K.; Zarate, C.A., Jr. Rapid onset of antidepressant action: A new paradigm in the research and treatment of major depressive disorder. J. Clin. Psychiatry 2008, 69, 946–958. [Google Scholar] [CrossRef]
- Soares, G.A.B.E.; Bhattacharya, T.; Chakrabarti, T.; Tagde, P.; Cavalu, S. Exploring Pharmacological Mechanisms of Essential Oils on the Central Nervous System. Plants 2021, 11, 21. [Google Scholar] [CrossRef]
- Liang, J.; Zhang, Y.; Chi, P.; Liu, H.; Jing, Z.; Cao, H.; Du, Y.; Zhao, Y.; Qin, X.; Zhang, W.; et al. Essential oils: Chemical constituents, potential neuropharmacological effects and aromatherapy—A review. Pharmacol. Res.—Mod. Chin. Med. 2023, 6, 100210. [Google Scholar] [CrossRef]
- Hanif, M.A.; Nisar, S.; Khan, G.S.; Mushtaq, Z.; Zubair, M. Essential Oils. In Essential Oil Research: Trends in Biosynthesis, Analytics, Industrial Applications and Biotechnological Production; Malik, S., Ed.; Springer International Publishing: Cham, Switzerland, 2019; pp. 3–17. [Google Scholar] [CrossRef]
- Fung, T.K.H.; Lau, B.W.M.; Ngai, S.P.C.; Tsang, H.W.H. Therapeutic Effect and Mechanisms of Essential Oils in Mood Disorders: Interaction between the Nervous and Respiratory Systems. Int. J. Mol. Sci. 2021, 22, 4844. [Google Scholar] [CrossRef]
- Cui, J.; Li, M.; Wei, Y.; Li, H.; He, X.; Yang, Q.; Li, Z.; Duan, J.; Wu, Z.; Chen, Q.; et al. Inhalation Aromatherapy via Brain-Targeted Nasal Delivery: Natural Volatiles or Essential Oils on Mood Disorders. Front. Pharmacol. 2022, 13, 860043. [Google Scholar] [CrossRef]
- Zhang, L.-L.; Yang, Z.-Y.; Fan, G.; Ren, J.-N.; Yin, K.-J.; Pan, S.-Y. Antidepressant-like Effect of Citrus sinensis (L.) Osbeck Essential Oil and Its Main Component Limonene on Mice. J. Agric. Food Chem. 2019, 67, 13817–13828. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Vidaña, D.I.; Po, K.K.; Fung, T.K.; Chow, J.K.; Lau, W.K.; So, P.K.; Lau, B.W.; Tsang, H.W. Lavender essential oil ameliorates depression-like behavior and increases neurogenesis and dendritic complexity in rats. Neurosci. Lett. 2019, 701, 180–192. [Google Scholar] [CrossRef] [PubMed]
- Chioca, L.R.; Ferro, M.M.; Baretta, I.P.; Oliveira, S.M.; Silva, C.R.; Ferreira, J.; Losso, E.M.; Andreatini, R. Anxiolytic-like effect of lavender essential oil inhalation in mice: Participation of serotonergic but not GABAA/benzodiazepine neurotransmission. J. Ethnopharmacol. 2013, 147, 412–418. [Google Scholar] [CrossRef] [PubMed]
- Jin, Z.; Han, Y.; Zhang, D.; Li, Z.; Jing, Y.; Hu, B.; Sun, S. Application of Intranasal Administration in the Delivery of Antidepressant Active Ingredients. Pharmaceutics 2022, 14, 2070. [Google Scholar] [CrossRef]
- Turner, P.V.; Brabb, T.; Pekow, C.; Vasbinder, M.A. Administration of substances to laboratory animals: Routes of administration and factors to consider. J. Am. Assoc. Lab. Anim. Sci. 2011, 50, 600–613. [Google Scholar] [PubMed]
- Qi, X.J.; Liu, X.Y.; Tang, L.M.; Li, P.F.; Qiu, F.; Yang, A.H. Anti-depressant effect of curcumin-loaded guanidine-chitosan thermo-sensitive hydrogel by nasal delivery. Pharm. Dev. Technol. 2020, 25, 316–325. [Google Scholar] [CrossRef]
- Wang, Q.-S.; Li, K.; Gao, L.-N.; Zhang, Y.; Lin, K.-M.; Cui, Y.-L. Intranasal delivery of berberine via in situ thermoresponsive hydrogel with non-invasive therapy exhibits better antidepressant-like effects. Biomater. Sci. 2020, 8, 2853–2865. [Google Scholar] [CrossRef]
- Qi, X.-J.; Xu, D.; Tian, M.-L.; Zhou, J.-F.; Wang, Q.-S.; Cui, Y.-L. Thermosensitive hydrogel designed for improving the antidepressant activities of genipin via intranasal delivery. Mater. Des. 2021, 206, 109816. [Google Scholar] [CrossRef]
- Xia, B.; Zhang, H.; Xue, W.; Tao, W.; Chen, C.; Wu, R.; Ren, L.; Tang, J.; Wu, H.; Cai, B.; et al. Instant and Lasting Down-Regulation of NR1 Expression in the Hippocampus is Associated Temporally with Antidepressant Activity after Acute Yueju. Cell. Mol. Neurobiol. 2016, 36, 1189–1196. [Google Scholar] [CrossRef]
- Hall, B.J.; Ripley, B.; Ghosh, A. NR2B signaling regulates the development of synaptic AMPA receptor current. J. Neurosci. 2007, 27, 13446–13456. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Ju, P.; Liu, H.; Wu, X.; Niu, Z.; Zhu, Y.; Zhang, C.; Fang, Y. Ifenprodil rapidly ameliorates depressive-like behaviors, activates mTOR signaling and modulates proinflammatory cytokines in the hippocampus of CUMS rats. Psychopharmacology 2020, 237, 1421–1433. [Google Scholar] [CrossRef]
- Li, J.-M.; Liu, L.-L.; Su, W.-J.; Wang, B.; Zhang, T.; Zhang, Y.; Jiang, C.-L. Ketamine may exert antidepressant effects via suppressing NLRP3 inflammasome to upregulate AMPA receptors. Neuropharmacology 2019, 146, 149–153. [Google Scholar] [CrossRef] [PubMed]
- Koike, H.; Iijima, M.; Chaki, S. Involvement of AMPA receptor in both the rapid and sustained antidepressant-like effects of ketamine in animal models of depression. Behav. Brain Res. 2011, 224, 107–111. [Google Scholar] [CrossRef] [PubMed]
- Hara, H.; Suzuki, A.; Kunugi, A.; Tajima, Y.; Yamada, R.; Kimura, H. TAK-653, an AMPA receptor potentiator with minimal agonistic activity, produces an antidepressant-like effect with a favorable safety profile in rats. Pharmacol. Biochem. Behav. 2021, 211, 173289. [Google Scholar] [CrossRef]
- Can, A.; Dao, D.T.; Terrillion, C.E.; Piantadosi, S.C.; Bhat, S.; Gould, T.D. The tail suspension test. J. Vis. Exp. 2012, 59, e3769. [Google Scholar] [CrossRef]
- Hogg, S. A review of the validity and variability of the Elevated Plus-Maze as an animal model of anxiety. Pharmacol. Biochem. Behav. 1996, 54, 21–30. [Google Scholar] [CrossRef]
- Joyce Gem, M.C.; Joanna, J.O.; Junie, B.B. In Sılıco Screenıng for Neuroreceptor Targets and Derıvatızatıon of Alkaloıds from Phaeanthus Ophthalmıcus. Pharmacophore 2022, 13, 27–43. [Google Scholar] [CrossRef]
- Kaniakova, M.; Korabecny, J.; Holubova, K.; Kleteckova, L.; Chvojkova, M.; Hakenova, K.; Prchal, L.; Novak, M.; Dolezal, R.; Hepnarova, V.; et al. 7-phenoxytacrine is a dually acting drug with neuroprotective efficacy in vivo. Biochem. Pharmacol. 2021, 186, 114460. [Google Scholar] [CrossRef]
- Wei, L.; Qi, X.; Yu, X.; Zheng, Y.; Luo, X.; Wei, Y.; Ni, P.; Zhao, L.; Wang, Q.; Ma, X.; et al. 3,4-Dihydrobenzo[e][1,2,3] oxathiazine 2,2-dioxide analogs act as potential AMPA receptor potentiators with antidepressant activity. Eur. J. Med. Chem. 2023, 251, 115252. [Google Scholar] [CrossRef] [PubMed]
- Sweilam, S.H.; Alqarni, M.H.; Youssef, F.S. Antimicrobial Alkaloids from Marine-Derived Fungi as Drug Leads versus COVID-19 Infection: A Computational Approach to Explore their Anti-COVID-19 Activity and ADMET Properties. Evid. Based Complement. Altern. Med. 2022, 2022, 5403757. [Google Scholar] [CrossRef]
- Ramadhan, D.S.F.; Siharis, F.; Abdurrahman, S.; Isrul, M.; Fakih, T.M. In silico analysis of marine natural product from sponge (Clathria Sp.) for their activity as inhibitor of SARS-CoV-2 Main Protease. J. Biomol. Struct. Dyn. 2022, 40, 11526–11532. [Google Scholar] [CrossRef]
- Wu, W. GC-MS analysis of chemical components in essential oil from Flos magnoliae. Zhong Yao Cai 2000, 23, 538–541. [Google Scholar] [PubMed]
- Hu, M.; Bai, M.; Ye, W.; Wang, Y.; Wu, H. Variations in Volatile Oil Yield and Composition of “Xin-yi” (Magnolia biondii Pamp. Flower Buds) at Different Growth Stages. J. Oleo Sci. 2018, 67, 779–787. [Google Scholar] [CrossRef]
- Zeng, Z.; Xie, R.; Zhang, T.; Zhang, H.; Chen, J.Y. Analysis of volatile compositions of Magnolia biondii pamp by steam distillation and headspace solid phase micro-extraction. J. Oleo Sci. 2011, 60, 591–596. [Google Scholar] [CrossRef] [PubMed]
- Kuang, C.L.; Lv, D.; Shen, G.H.; Li, S.S.; Luo, Q.Y.; Zhang, Z.Q. Chemical composition and antimicrobial activities of volatile oil extracted from Chrysanthemum morifolium Ramat. J. Food Sci. Technol. 2018, 55, 2786–2794. [Google Scholar] [CrossRef]
- Peng, A.; Lin, L.; Zhao, M. Screening of key flavonoids and monoterpenoids for xanthine oxidase inhibitory activity-oriented quality control of Chrysanthemum morifolium Ramat. ‘Boju’ based on spectrum-effect relationship coupled with UPLC-TOF-MS and HS-SPME-GC/MS. Food Res. Int. 2020, 137, 109448. [Google Scholar] [CrossRef]
- Li, X.; Ao, M.; Zhang, C.; Fan, S.; Chen, Z.; Yu, L. Zingiberis Rhizoma Recens: A Review of Its Traditional Uses, Phytochemistry, Pharmacology, and Toxicology. Evid. Based Complement. Altern. 2021, 2021, 6668990. [Google Scholar] [CrossRef]
- Yu, D.-X.; Guo, S.; Wang, J.-M.; Yan, H.; Zhang, Z.-Y.; Yang, J.; Duan, J.-A. Comparison of Different Drying Methods on the Volatile Components of Ginger (Zingiber officinale Roscoe) by HS-GC-MS Coupled with Fast GC E-Nose. Foods 2022, 11, 1611. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Pandotra, P.; Ram, G.; Anand, R.; Gupta, A.P.; Husain, K.; Bedi, Y.S.; Mallavarapu, G.R. Composition of a monoterpenoid-rich essential oil from the rhizome of Zingiber officinale from north western Himalayas. Nat. Prod. Commun. 2011, 6, 93–96. [Google Scholar] [CrossRef]
- de Groot, A.C.; Schmidt, E. Essential Oils, Part VI: Sandalwood Oil, Ylang-Ylang Oil, and Jasmine Absolute. Dermatitis 2017, 28, 14–21. [Google Scholar] [CrossRef]
- Braun, N.A.; Meier, M.; Pickenhagen, W. Isolation and Chiral GC Analysis of β-Bisabolols—Trace Constituents from the Essential Oil of Santalum album L. (Santalaceae). J. Essent. Oil Res. 2003, 15, 63–65. [Google Scholar] [CrossRef]
- Mohankumar, A.; Kalaiselvi, D.; Levenson, C.; Shanmugam, G.; Thiruppathi, G.; Nivitha, S.; Sundararaj, P. Antioxidant and stress modulatory efficacy of essential oil extracted from plantation-grown Santalum album L. Ind. Crop. Prod. 2019, 140, 111623. [Google Scholar] [CrossRef]
- Fahmy, N.M.; Elhady, S.S.; Bannan, D.F.; Malatani, R.T.; Gad, H.A. Citrus reticulata Leaves Essential Oil as an Antiaging Agent: A Comparative Study between Different Cultivars and Correlation with Their Chemical Compositions. Plants 2022, 11, 3335. [Google Scholar] [CrossRef] [PubMed]
- Duan, L.; Guo, L.; Dou, L.-L.; Zhou, C.-L.; Xu, F.-G.; Zheng, G.-D.; Li, P.; Liu, E.H. Discrimination of Citrus reticulata Blanco and Citrus reticulata ‘Chachi’ by gas chromatograph-mass spectrometry based metabolomics approach. Food Chem. 2016, 212, 123–127. [Google Scholar] [CrossRef] [PubMed]
- Chutia, M.; Deka Bhuyan, P.; Pathak, M.G.; Sarma, T.C.; Boruah, P. Antifungal activity and chemical composition of Citrus reticulata Blanco essential oil against phytopathogens from North East India. LWT—Food Sci. Technol. 2009, 42, 777–780. [Google Scholar] [CrossRef]
- Lota, M.-L.; de Rocca Serra, D.; Tomi, F.; Joseph, C. Chemical variability of peel and leaf essential oils of mandarins from Citrus reticulata Blanco. Biochem. Syst. Ecol. 2000, 28, 61–78. [Google Scholar] [CrossRef]
- Zhang, W.J.; Zhao, Z.Y.; Chang, L.K.; Cao, Y.; Wang, S.; Kang, C.Z.; Wang, H.Y.; Zhou, L.; Huang, L.Q.; Guo, L.P. Atractylodis Rhizoma: A review of its traditional uses, phytochemistry, pharmacology, toxicology and quality control. J. Ethnopharmacol. 2021, 266, 113415. [Google Scholar] [CrossRef]
- Xu, R.; Lu, J.; Wu, J.; Yu, D.; Chu, S.; Guan, F.; Liu, W.; Hu, J.; Peng, H.; Zha, L. Comparative analysis in different organs and tissue-specific metabolite profiling of Atractylodes lancea from four regions by GC-MS and laser microdissection. J. Sep. Sci. 2022, 45, 1067–1079. [Google Scholar] [CrossRef]
- Lei, H.; Yue, J.; Yin, X.Y.; Fan, W.; Tan, S.H.; Qin, L.; Zhao, Y.N.; Bai, J.H. HS-SPME coupled with GC-MS for elucidating differences between the volatile components in wild and cultivated Atractylodes chinensis. Phytochem. Anal. 2023, 34, 317–328. [Google Scholar] [CrossRef]
- Ashokkumar, K.; Simal-Gandara, J.; Murugan, M.; Dhanya, M.K.; Pandian, A. Nutmeg (Myristica fragrans Houtt.) essential oil: A review on its composition, biological, and pharmacological activities. Phytother. Res. 2022, 36, 2839–2851. [Google Scholar] [CrossRef]
- Gupta, A.D.; Bansal, V.K.; Babu, V.; Maithil, N. Chemistry, antioxidant and antimicrobial potential of nutmeg (Myristica fragrans Houtt). J. Genet. Eng. Biotechnol. 2013, 11, 25–31. [Google Scholar] [CrossRef]
- Ogunwande, I.A.; Olawore, N.O.; Adeleke, K.A.; Ekundayo, O. Chemical Composition of Essential Oil of Myristica Fragrans Houtt (Nutmeg) From Nigeria. J. Essent. Oil Bear. Plants 2003, 6, 21–26. [Google Scholar] [CrossRef]
- Berman, R.M.; Cappiello, A.; Anand, A.; Oren, D.A.; Heninger, G.R.; Charney, D.S.; Krystal, J.H. Antidepressant effects of ketamine in depressed patients. Biol. Psychiatry 2000, 47, 351–354. [Google Scholar] [CrossRef]
- Zarate, C.A., Jr.; Singh, J.B.; Carlson, P.J.; Brutsche, N.E.; Ameli, R.; Luckenbaugh, D.A.; Charney, D.S.; Manji, H.K. A randomized trial of an N-methyl-D-aspartate antagonist in treatment-resistant major depression. Arch. Gen. Psychiatry 2006, 63, 856–864. [Google Scholar] [CrossRef] [PubMed]
- de Sousa, D.; Hocayen, P.D.A.S.; Andrade, L.N.; Andreatini, R. A Systematic Review of the Anxiolytic-Like Effects of Essential Oils in Animal Models. Molecules 2015, 20, 18620–18660. [Google Scholar] [CrossRef]
- Almeida, R.C.; Souza, D.G.; Soletti, R.C.; López, M.G.; Rodrigues, A.L.; Gabilan, N.H. Involvement of PKA, MAPK/ERK and CaMKII, but not PKC in the acute antidepressant-like effect of memantine in mice. Neurosci. Lett. 2006, 395, 93–97. [Google Scholar] [CrossRef] [PubMed]
- McShane, R.; Westby, M.J.; Roberts, E.; Minakaran, N.; Schneider, L.; Farrimond, L.E.; Maayan, N.; Ware, J.; Debarros, J. Memantine for dementia. Cochrane Database Syst. Rev. 2019, 3, Cd003154. [Google Scholar] [CrossRef]
- Pascual, O.; Ben Achour, S.; Rostaing, P.; Triller, A.; Bessis, A. Microglia activation triggers astrocyte-mediated modulation of excitatory neurotransmission. Proc. Natl. Acad. Sci. USA 2012, 109, E197–E205. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, Y.; Si, Q.S.; Kataoka, K. Lipopolysaccharide-induced microglial activation in culture: Temporal profiles of morphological change and release of cytokines and nitric oxide. Neurosci. Res. 1999, 35, 95–100. [Google Scholar] [CrossRef]
- Dunn, A.J.; Wang, J.; Ando, T. Effects of cytokines on cerebral neurotransmission. Comparison with the effects of stress. Adv. Exp. Med. Biol. 1999, 461, 117–127. [Google Scholar] [CrossRef]
- Zhao, Z.; Zhang, L.; Guo, X.D.; Cao, L.L.; Xue, T.F.; Zhao, X.J.; Yang, D.D.; Yang, J.; Ji, J.; Huang, J.Y.; et al. Rosiglitazone Exerts an Anti-depressive Effect in Unpredictable Chronic Mild-Stress-Induced Depressive Mice by Maintaining Essential Neuron Autophagy and Inhibiting Excessive Astrocytic Apoptosis. Front. Mol. Neurosci. 2017, 10, 293. [Google Scholar] [CrossRef] [PubMed]
- Mo, F.; Tang, Y.; Du, P.; Shen, Z.; Yang, J.; Cai, M.; Zhang, Y.; Li, H.; Shen, H. GPR39 protects against corticosterone-induced neuronal injury in hippocampal cells through the CREB-BDNF signaling pathway. J. Affect. Disord. 2020, 272, 474–484. [Google Scholar] [CrossRef] [PubMed]
- Jin, W.; Xu, X.; Chen, X.; Qi, W.; Lu, J.; Yan, X.; Zhao, D.; Cong, D.; Li, X.; Sun, L. Protective effect of pig brain polypeptides against corticosterone-induced oxidative stress, inflammatory response, and apoptosis in PC12 cells. Biomed. Pharmacother. 2019, 115, 108890. [Google Scholar] [CrossRef] [PubMed]
- Gruver-Yates, A.L.; Cidlowski, J.A. Tissue-specific actions of glucocorticoids on apoptosis: A double-edged sword. Cells 2013, 2, 202–223. [Google Scholar] [CrossRef]
- Zhang, S.Q.; Cao, L.L.; Liang, Y.Y.; Wang, P. The Molecular Mechanism of Chronic High-Dose Corticosterone-Induced Aggravation of Cognitive Impairment in APP/PS1 Transgenic Mice. Front. Mol. Neurosci. 2020, 13, 613421. [Google Scholar] [CrossRef]
- Wang, Q.; Van Heerikhuize, J.; Aronica, E.; Kawata, M.; Seress, L.; Joels, M.; Swaab, D.F.; Lucassen, P.J. Glucocorticoid receptor protein expression in human hippocampus; stability with age. Neurobiol. Aging 2013, 34, 1662–1673. [Google Scholar] [CrossRef] [PubMed]
- Gradin, V.B.; Pomi, A. The role of hippocampal atrophy in depression: A neurocomputational approach. J. Biol. Phys. 2008, 34, 107–120. [Google Scholar] [CrossRef]
- Hao, Y.; Shabanpoor, A.; Metz, G.A. Stress and corticosterone alter synaptic plasticity in a rat model of Parkinson’s disease. Neurosci. Lett. 2017, 651, 79–87. [Google Scholar] [CrossRef]
- Xu, L.; Holscher, C.; Anwyl, R.; Rowan, M.J. Glucocorticoid receptor and protein/RNA synthesis-dependent mechanisms underlie the control of synaptic plasticity by stress. Proc. Natl. Acad. Sci. USA 1998, 95, 3204–3208. [Google Scholar] [CrossRef]
- Kumar, P.; Nagarajan, A.; Uchil, P.D. Analysis of Cell Viability by the Lactate Dehydrogenase Assay. Cold Spring Harb. Protoc. 2018, 2018. [Google Scholar] [CrossRef]
- Park, Y.-k.; Chung, Y.S.; Kim, Y.S.; Kwon, O.Y.; Joh, T.H. Inhibition of gene expression and production of iNOS and TNF-α in LPS-stimulated microglia by methanol extract of Phellodendri cortex. Int. Immunopharmacol. 2007, 7, 955–962. [Google Scholar] [CrossRef]
- Lee, S.H.; Lee, J.H.; Oh, E.Y.; Kim, G.Y.; Choi, B.T.; Kim, C.; Choi, Y.H. Ethanol extract of Cnidium officinale exhibits anti-inflammatory effects in BV2 microglial cells by suppressing NF-κB nuclear translocation and the activation of the PI3K/Akt signaling pathway. Int. J. Mol. Med. 2013, 32, 876–882. [Google Scholar] [CrossRef]
- Wu, F.; Li, H.; Zhao, L.; Li, X.; You, J.; Jiang, Q.; Li, S.; Jin, L.; Xu, Y. Protective effects of aqueous extract from Acanthopanax senticosus against corticosterone-induced neurotoxicity in PC12 cells. J. Ethnopharmacol. 2013, 148, 861–868. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Jin, W.; Cui, Y.; Ao, M.; Liu, H.; Xu, H.; Yu, L. Protective effects of macamides from Lepidium meyenii Walp. against corticosterone-induced neurotoxicity in PC12 cells. RSC Adv. 2019, 9, 23096–23108. [Google Scholar] [CrossRef]
- Dhingra, D.; Sharma, A. Antidepressant-like activity of n-hexane extract of nutmeg (Myristica fragrans) seeds in mice. J. Med. Food 2006, 9, 84–89. [Google Scholar] [CrossRef] [PubMed]
- Iwata, N.; Kobayashi, D.; Kawashiri, T.; Kubota, T.; Kawano, K.; Yamamuro, Y.; Miyagi, A.; Deguchi, Y.; Chijimatsu, T.; Shimazoe, T. Mechanisms and Safety of Antidepressant-Like Effect of Nutmeg in Mice. Biol. Pharm. Bull. 2022, 45, 738–742. [Google Scholar] [CrossRef]
- Muris, P.; Merckelbach, H.; Schmidt, H.; Gadet, B.; Bogie, N. Anxiety and depression as correlates of self-reported behavioural inhibition in normal adolescents. Behav. Res. Ther. 2001, 39, 1051–1061. [Google Scholar] [CrossRef]
- Tiller, J.W. Depression and anxiety. Med. J. Aust. 2013, 199, S28–S31. [Google Scholar] [CrossRef] [PubMed]
- Zanos, P.; Gould, T.D. Mechanisms of ketamine action as an antidepressant. Mol. Psychiatry 2018, 23, 801–811. [Google Scholar] [CrossRef]
- Sanacora, G.; Schatzberg, A.F. Ketamine: Promising Path or False Prophecy in the Development of Novel Therapeutics for Mood Disorders? Neuropsychopharmacology 2015, 40, 259–267. [Google Scholar] [CrossRef] [PubMed]
- Shamsi, A.; Shahwan, M.; Khan, M.S.; Husain, F.M.; Alhumaydhi, F.A.; Aljohani, A.S.M.; Rehman, M.T.; Hassan, M.I.; Islam, A. Elucidating the Interaction of Human Ferritin with Quercetin and Naringenin: Implication of Natural Products in Neurodegenerative Diseases: Molecular Docking and Dynamics Simulation Insight. ACS Omega 2021, 6, 7922–7930. [Google Scholar] [CrossRef]
- Khan, A.; Mohammad, T.; Shamsi, A.; Hussain, A.; Alajmi, M.F.; Husain, S.A.; Iqbal, M.A.; Hassan, M.I. Identification of plant-based hexokinase 2 inhibitors: Combined molecular docking and dynamics simulation studies. J. Biomol. Struct. Dyn. 2022, 40, 10319–10331. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Xu, Z.; Gao, Y.; Wu, Y.; Li, Z.; Liu, H.; Zhang, C. Bidirectional crosstalk between stress-induced gastric ulcer and depression under chronic stress. PLoS ONE 2012, 7, e51148. [Google Scholar] [CrossRef]
- Lin, Y.; Liu, X.; Tan, D.; Jiang, Z. Atractylon treatment prevents sleep-disordered breathing-induced cognitive dysfunction by suppression of chronic intermittent hypoxia-induced M1 microglial activation. Biosci. Rep. 2020, 40, BSR20192800. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.G.; Jan, Y.S.; Tsai, P.W.; Norimoto, H.; Michihara, S.; Murayama, C.; Wang, C.C. Anti-inflammatory and Antinociceptive Constituents of Atractylodes japonica Koidzumi. J. Agric. Food Chem. 2016, 64, 2254–2262. [Google Scholar] [CrossRef]
- Lin, L.-Z.; Harnly, J.M. Identification of the phenolic components of chrysanthemum flower (Chrysanthemum morifolium Ramat). Food Chem. 2010, 120, 319–326. [Google Scholar] [CrossRef]
- Duh, P.-D.; Tu, Y.-Y.; Yen, G.-C. Antioxidant Activity of Water Extract of Harng Jyur (Chrysanthemum morifolium Ramat). LWT—Food Sci. Technol. 1999, 32, 269–277. [Google Scholar] [CrossRef]
- Yuan, H.; Jiang, S.; Liu, Y.; Daniyal, M.; Jian, Y.; Peng, C.; Shen, J.; Liu, S.; Wang, W. The flower head of Chrysanthemum morifolium Ramat. (Juhua): A paradigm of flowers serving as Chinese dietary herbal medicine. J. Ethnopharmacol. 2020, 261, 113043. [Google Scholar] [CrossRef]
- Ibi, M.; Yabe-Nishimura, C. Chapter 1—The role of reactive oxygen species in the pathogenic pathways of depression. In Oxidative Stress and Dietary Antioxidants in Neurological Diseases; Martin, C.R., Preedy, V.R., Eds.; Academic Press: Cambridge, MA, USA, 2020; pp. 3–16. [Google Scholar] [CrossRef]
- Liu, T.; Zhou, N.; Xu, R.; Cao, Y.; Zhang, Y.; Liu, Z.; Zheng, X.; Feng, W. A metabolomic study on the anti-depressive effects of two active components from Chrysanthemum morifolium. Artif. Cells Nanomed. Biotechnol. 2020, 48, 718–727. [Google Scholar] [CrossRef] [PubMed]
- Schepetkin, I.A.; Özek, G.; Özek, T.; Kirpotina, L.N.; Khlebnikov, A.I.; Klein, R.A.; Quinn, M.T. Neutrophil Immunomodulatory Activity of Farnesene, a Component of Artemisia dracunculus Essential Oils. Pharmaceuticals 2022, 15, 642. [Google Scholar] [CrossRef]
- Ha, M.T.; Vu, N.K.; Tran, T.H.; Kim, J.A.; Woo, M.H.; Min, B.S. Phytochemical and pharmacological properties of Myristica fragrans Houtt.: An updated review. Arch. Pharmacal Res. 2020, 43, 1067–1092. [Google Scholar] [CrossRef]
- Workman, E.R.; Niere, F.; Raab-Graham, K.F. mTORC1-dependent protein synthesis underlying rapid antidepressant effect requires GABABR signaling. Neuropharmacology 2013, 73, 192–203. [Google Scholar] [CrossRef] [PubMed]
- Mendez-David, I.; David, D.J.; Darcet, F.; Wu, M.V.; Kerdine-Römer, S.; Gardier, A.M.; Hen, R. Rapid Anxiolytic Effects of a 5-HT4 Receptor Agonist Are Mediated by a Neurogenesis-Independent Mechanism. Neuropsychopharmacology 2014, 39, 1366–1378. [Google Scholar] [CrossRef]
- Faye, C.; Hen, R.; Guiard, B.P.; Denny, C.A.; Gardier, A.M.; Mendez-David, I.; David, D.J. Rapid Anxiolytic Effects of RS67333, a Serotonin Type 4 Receptor Agonist, and Diazepam, a Benzodiazepine, Are Mediated by Projections from the Prefrontal Cortex to the Dorsal Raphe Nucleus. Biol. Psychiatry 2020, 87, 514–525. [Google Scholar] [CrossRef] [PubMed]
- Satou, T.; Kasuya, H.; Maeda, K.; Koike, K. Daily inhalation of α-pinene in mice: Effects on behavior and organ accumulation. Phytother. Res. 2014, 28, 1284–1287. [Google Scholar] [CrossRef] [PubMed]
- Shringarpure, M.; Gharat, S.; Momin, M.; Omri, A. Management of epileptic disorders using nanotechnology-based strategies for nose-to-brain drug delivery. Expert Opin. Drug Deliv. 2021, 18, 169–185. [Google Scholar] [CrossRef]
- Long, Y.; Yang, Q.; Xiang, Y.; Zhang, Y.; Wan, J.; Liu, S.; Li, N.; Peng, W. Nose to brain drug delivery—A promising strategy for active components from herbal medicine for treating cerebral ischemia reperfusion. Pharmacol. Res. 2020, 159, 104795. [Google Scholar] [CrossRef]
- Walf, A.A.; Frye, C.A. A Review and Update of Mechanisms of Estrogen in the Hippocampus and Amygdala for Anxiety and Depression Behavior. Neuropsychopharmacology 2006, 31, 1097–1111. [Google Scholar] [CrossRef]
- Meltzer, E.O. Formulation considerations of intranasal corticosteroids for the treatment of allergic rhinitis. Ann. Allergy Asthma Immunol. 2007, 98, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Caimmi, D.; Neukirch, C.; Louis, R.; Malard, O.; Thabut, G.; Demoly, P. Effect of the Use of Intranasal Spray of Essential Oils in Patients with Perennial Allergic Rhinitis: A Prospective Study. Int. Arch. Allergy Immunol. 2021, 182, 182–189. [Google Scholar] [CrossRef]
- Vale, T.G.; Matos, F.J.A.; de Lima, T.C.M.; Viana, G.S.B. Behavioral effects of essential oils from Lippia alba (Mill.) N.E. Brown chemotypes. J. Ethnopharmacol. 1999, 67, 127–133. [Google Scholar] [CrossRef]
- Guo, J.; Duan, J.A.; Tang, Y.; Jia, N.; Li, X.; Zhang, J. Fast onset of action and the analgesic and sedative efficacy of essential oil from Rhizoma Chuanxiong after nasal administration. Pharmazie 2010, 65, 296–299. [Google Scholar] [PubMed]
- Guo, J.; Duan, J.A.; Tang, Y.; Li, Y. Sedative and anticonvulsant activities of styrax after oral and intranasal administration in mice. Pharm. Biol. 2011, 49, 1034–1038. [Google Scholar] [CrossRef] [PubMed]
- Can, Ö.D.; Demir Özkay, Ü.; Kıyan, H.T.; Demirci, B. Psychopharmacological profile of Chamomile (Matricaria recutita L.) essential oil in mice. Phytomedicine 2012, 19, 306–310. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Effects of (A) CORT and (B) essential oils on the viability of PC12 cells. Results are presented as means ± SDs (n = 3 per experiment). * p < 0.05 vs. CON.
Figure 1.
Effects of (A) CORT and (B) essential oils on the viability of PC12 cells. Results are presented as means ± SDs (n = 3 per experiment). * p < 0.05 vs. CON.


Figure 2.
Effects of essential oils on CORT-induced neurotoxicity in PC12 cells. (A) Effects of essential oils on the cell viability in CORT-stimulated PC12 cells. (B) Effects of essential oils on the LDH release in CORT-stimulated PC12 cells. Results are presented as means ± SDs (n = 3 per experiment). * p < 0.05 vs. CON, # p < 0.05 vs. CORT-treated cells.
Figure 2.
Effects of essential oils on CORT-induced neurotoxicity in PC12 cells. (A) Effects of essential oils on the cell viability in CORT-stimulated PC12 cells. (B) Effects of essential oils on the LDH release in CORT-stimulated PC12 cells. Results are presented as means ± SDs (n = 3 per experiment). * p < 0.05 vs. CON, # p < 0.05 vs. CORT-treated cells.
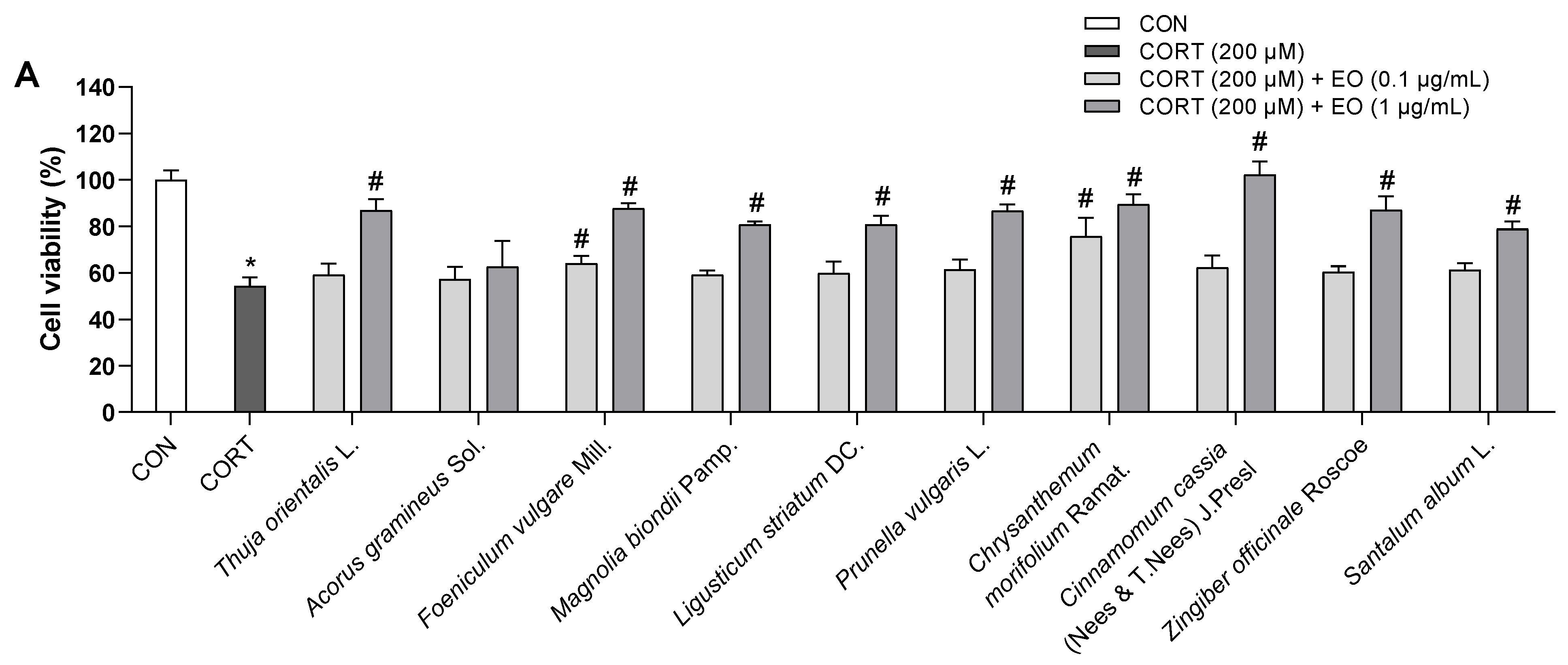

Figure 3.
Effects of essential oils on the viability of BV2 cells. Results are presented as means ± SDs (n = 3 per experiment). * p < 0.05 vs. CON.
Figure 3.
Effects of essential oils on the viability of BV2 cells. Results are presented as means ± SDs (n = 3 per experiment). * p < 0.05 vs. CON.

Figure 4.
Effects of essential oils on LPS-induced neuroinflammation in BV2 cells. (A) TNF-α levels (pg/mL). (B) IL-6 levels (pg/mL). Results are presented as means ± SDs (n = 3 per experiment). * p < 0.05 vs. CON, # p < 0.05 vs. LPS-treated cells.
Figure 4.
Effects of essential oils on LPS-induced neuroinflammation in BV2 cells. (A) TNF-α levels (pg/mL). (B) IL-6 levels (pg/mL). Results are presented as means ± SDs (n = 3 per experiment). * p < 0.05 vs. CON, # p < 0.05 vs. LPS-treated cells.
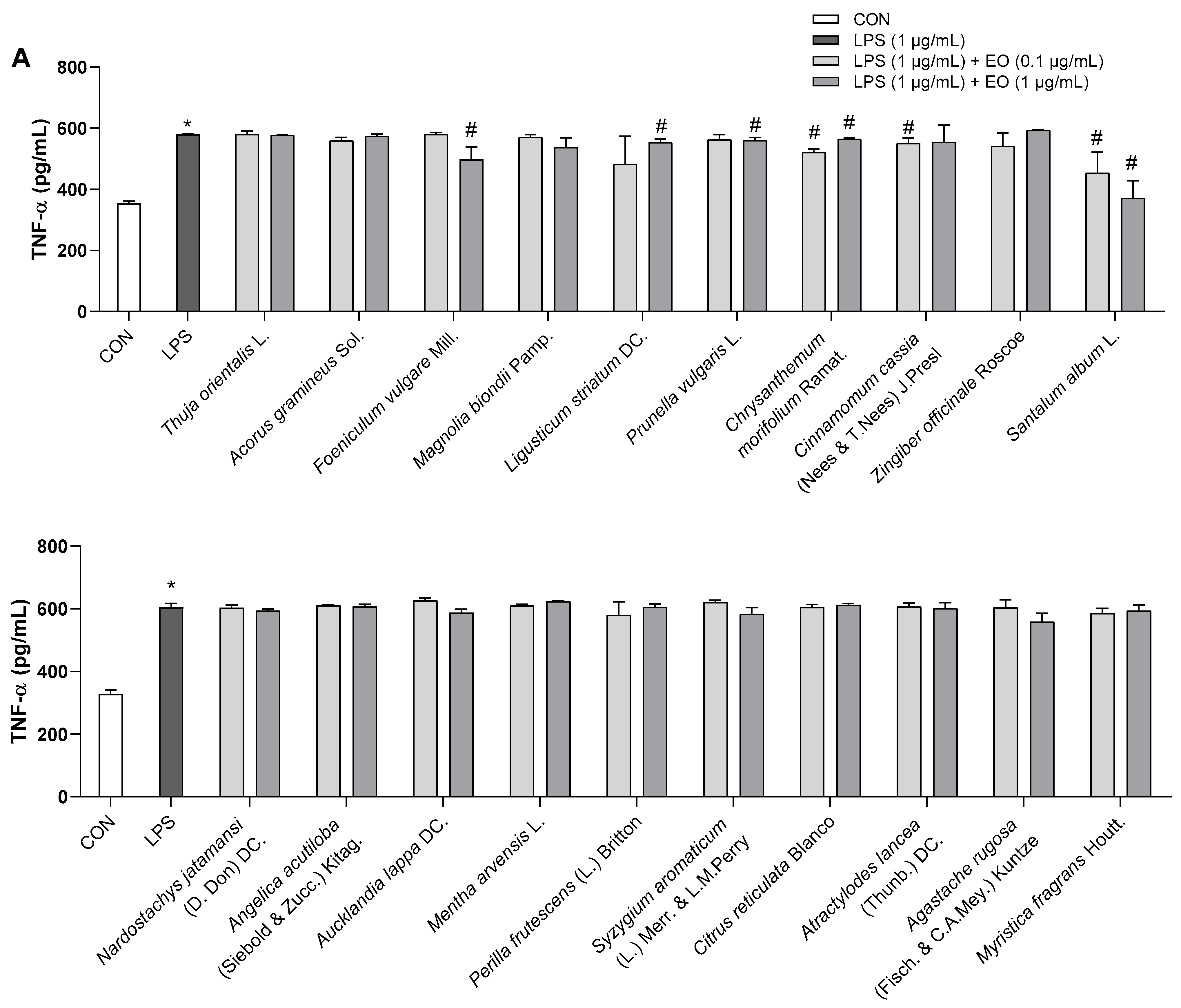

Figure 5.
Effects of essential oils on the behavioral changes of mice in TST test. The immobility time in TST was recorded. Each black triangle indicates the value of each mouse subject in groups. Results are presented as means percentage relative to control ± SDs (n = 4 mice per group). * p < 0.05 vs. CON.
Figure 5.
Effects of essential oils on the behavioral changes of mice in TST test. The immobility time in TST was recorded. Each black triangle indicates the value of each mouse subject in groups. Results are presented as means percentage relative to control ± SDs (n = 4 mice per group). * p < 0.05 vs. CON.
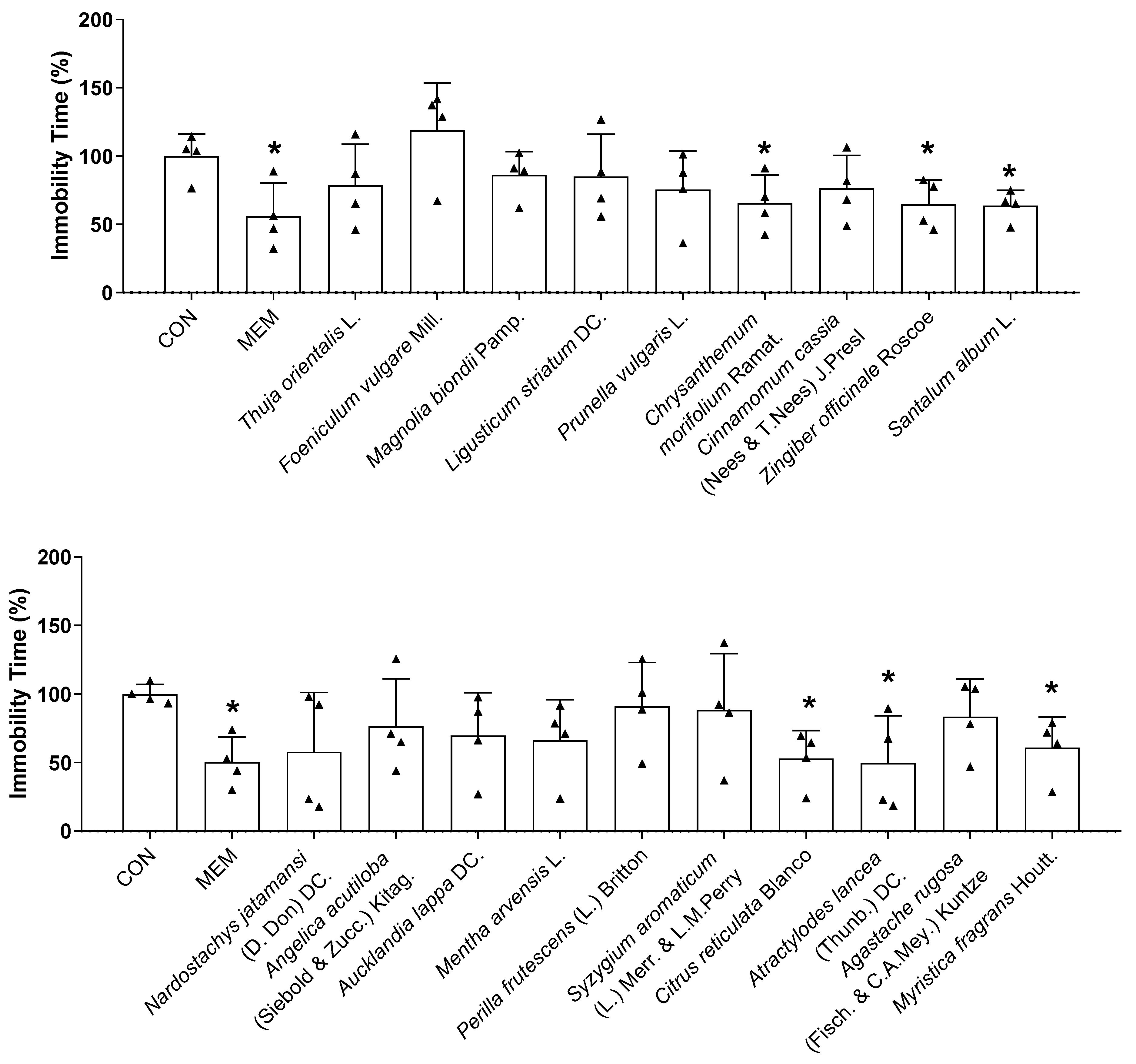
Figure 6.
Effects of essential oils on the behavioral changes of mice in EPM test. The (A) number of entries, (B) time, and (C) distance in open arms were recorded. Each black triangle indicates the value of each mouse subject in groups. Results are presented as means percentage relative to control ± SDs (n = 4 mice per group). * p < 0.05 vs. CON.
Figure 6.
Effects of essential oils on the behavioral changes of mice in EPM test. The (A) number of entries, (B) time, and (C) distance in open arms were recorded. Each black triangle indicates the value of each mouse subject in groups. Results are presented as means percentage relative to control ± SDs (n = 4 mice per group). * p < 0.05 vs. CON.

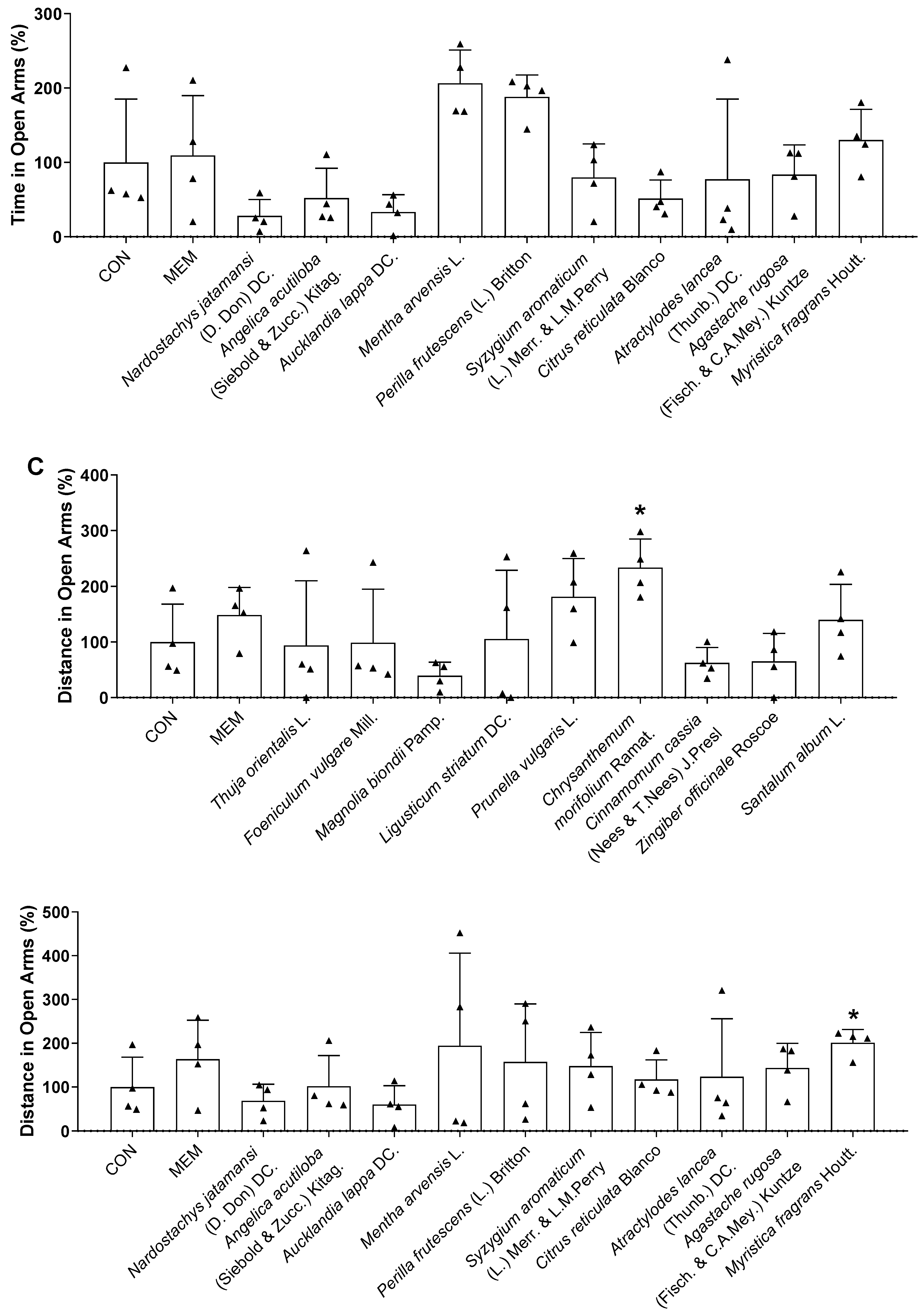
Figure 7.
Superimposed zoomed-in image of native ligand and redocked native ligand in the active site of (A) GluN1, (B) GluN2B, and (C) GluA2 (native ligand: gray color, redocked ligand: yellow color).
Figure 7.
Superimposed zoomed-in image of native ligand and redocked native ligand in the active site of (A) GluN1, (B) GluN2B, and (C) GluA2 (native ligand: gray color, redocked ligand: yellow color).
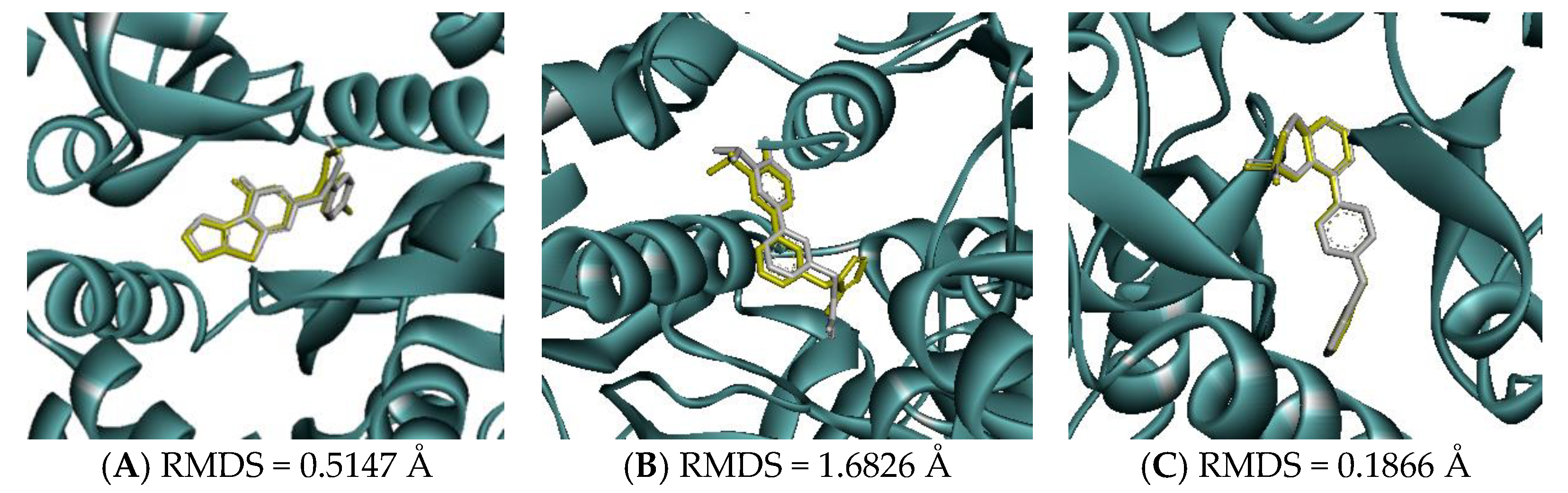
Figure 8.
2D interactions between (A) ketamine, (B) ifenprodil, (C) TAK-653, (D) atractylon, (E) α-curcumene, (F) α-farnesene, and (G) selina-4(14),7(11)-dien-8-one with GluN1 protein.
Figure 8.
2D interactions between (A) ketamine, (B) ifenprodil, (C) TAK-653, (D) atractylon, (E) α-curcumene, (F) α-farnesene, and (G) selina-4(14),7(11)-dien-8-one with GluN1 protein.

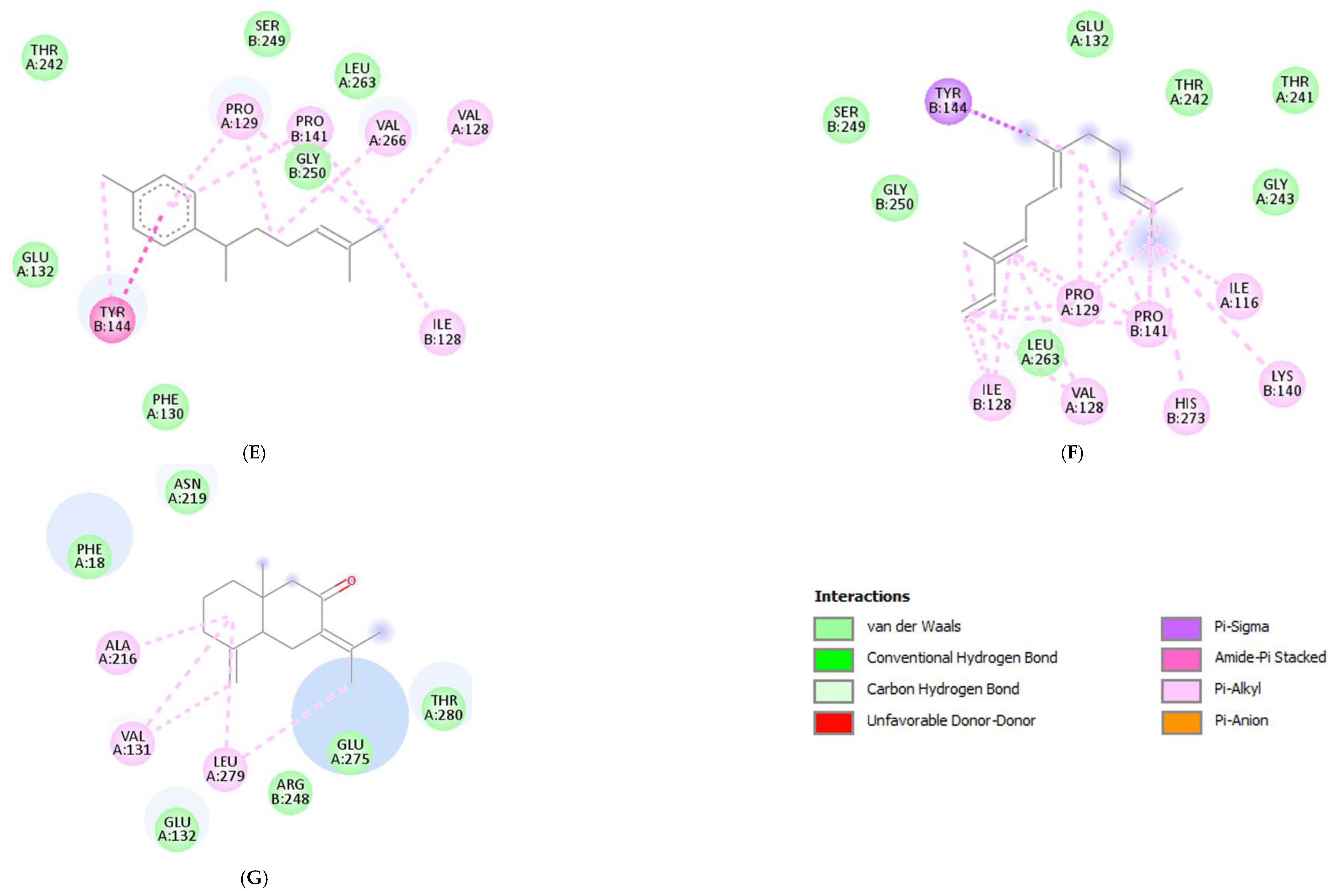
Figure 9.
2D interactions between (A) ketamine, (B) ifenprodil, (C) TAK-653, (D) atractylon, (E) α-curcumene, (F) α-farnesene, and (G) selina-4(14),7(11)-dien-8-one with GluN2B protein.
Figure 9.
2D interactions between (A) ketamine, (B) ifenprodil, (C) TAK-653, (D) atractylon, (E) α-curcumene, (F) α-farnesene, and (G) selina-4(14),7(11)-dien-8-one with GluN2B protein.
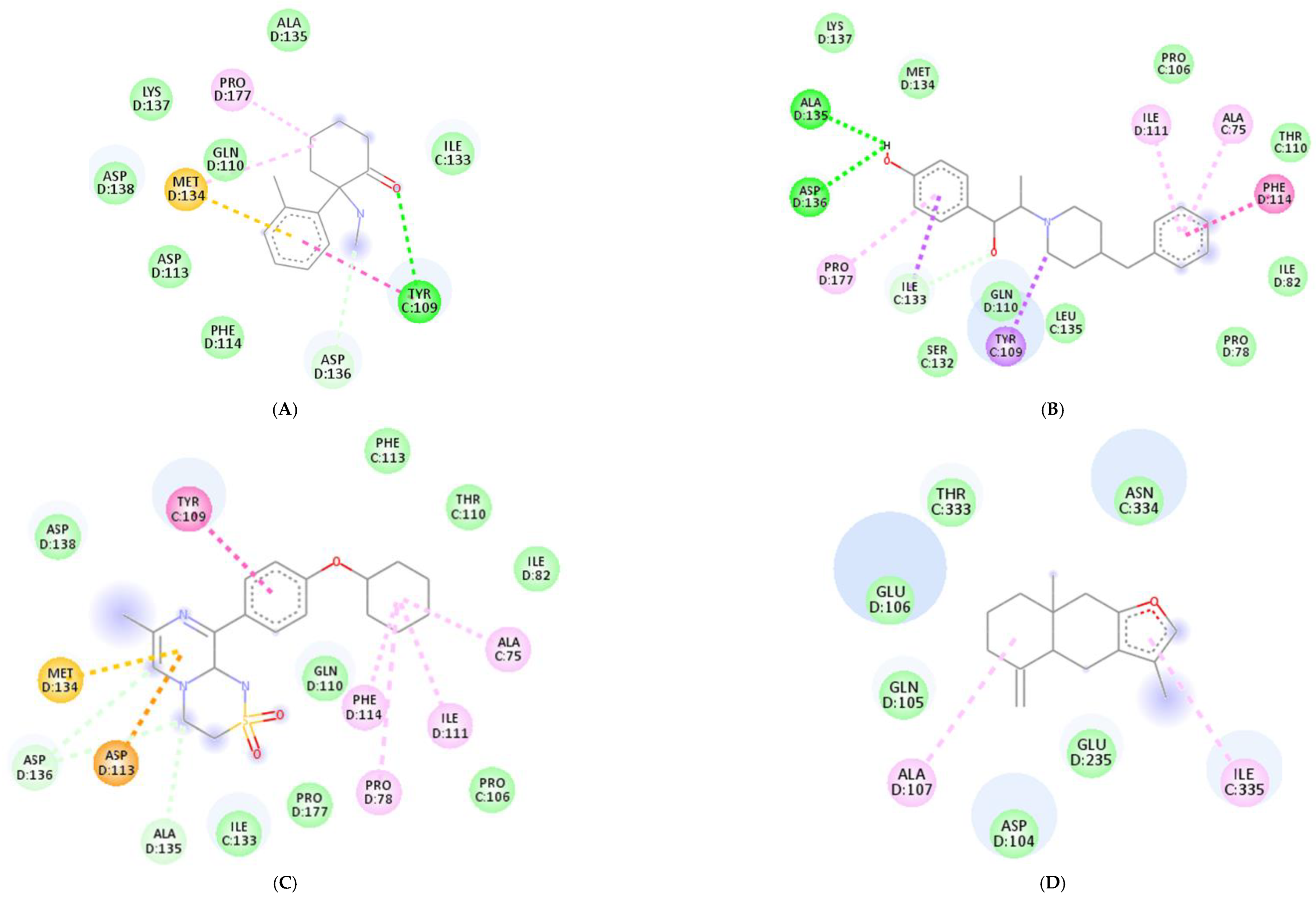
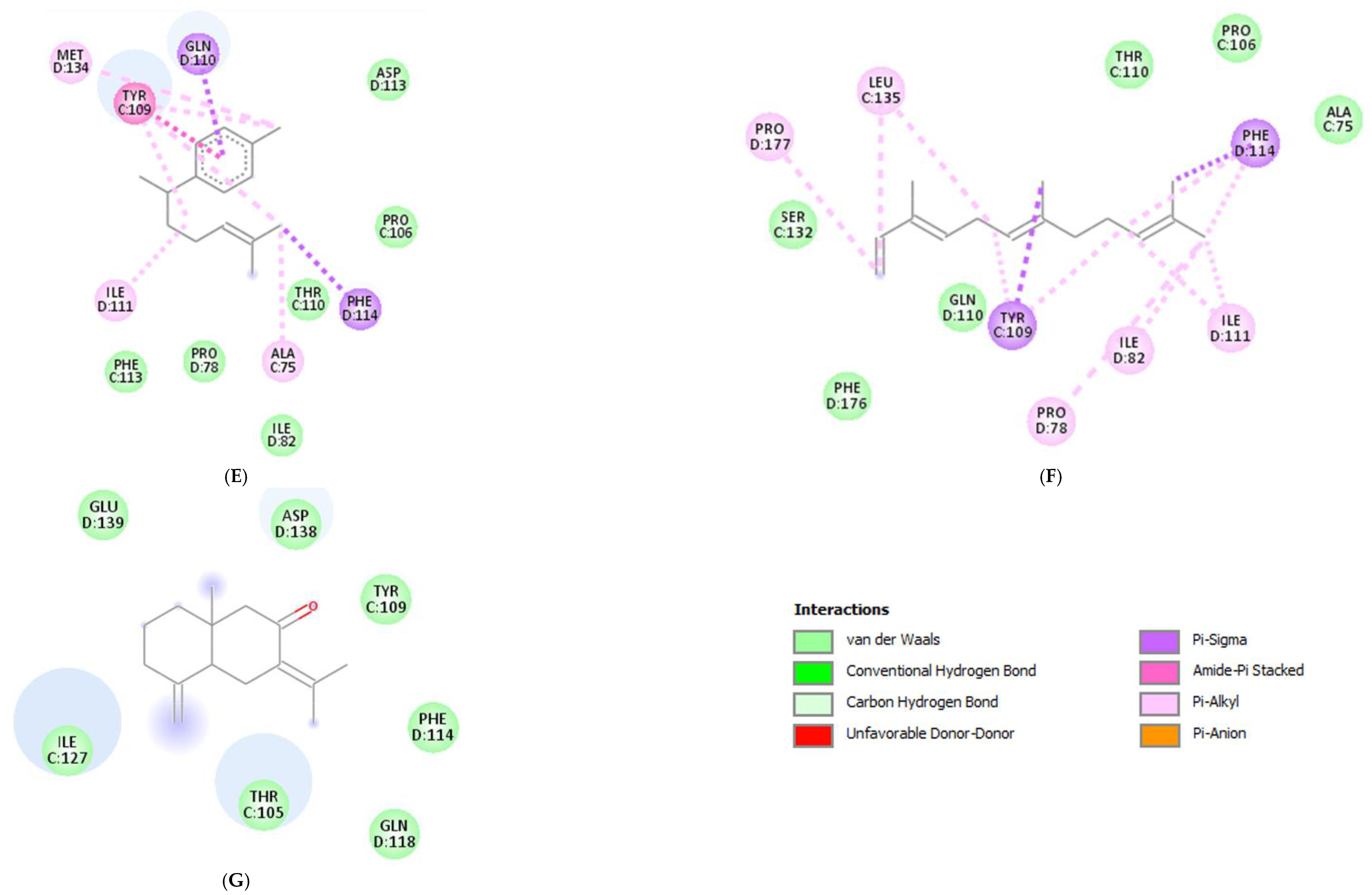
Figure 10.
2D interactions between (A) ketamine, (B) ifenprodil, (C) TAK-653, (D) atractylon, (E) α-curcumene, (F) α-farnesene, and (G) selina-4(14),7(11)-dien-8-one with GluA2 protein.
Figure 10.
2D interactions between (A) ketamine, (B) ifenprodil, (C) TAK-653, (D) atractylon, (E) α-curcumene, (F) α-farnesene, and (G) selina-4(14),7(11)-dien-8-one with GluA2 protein.
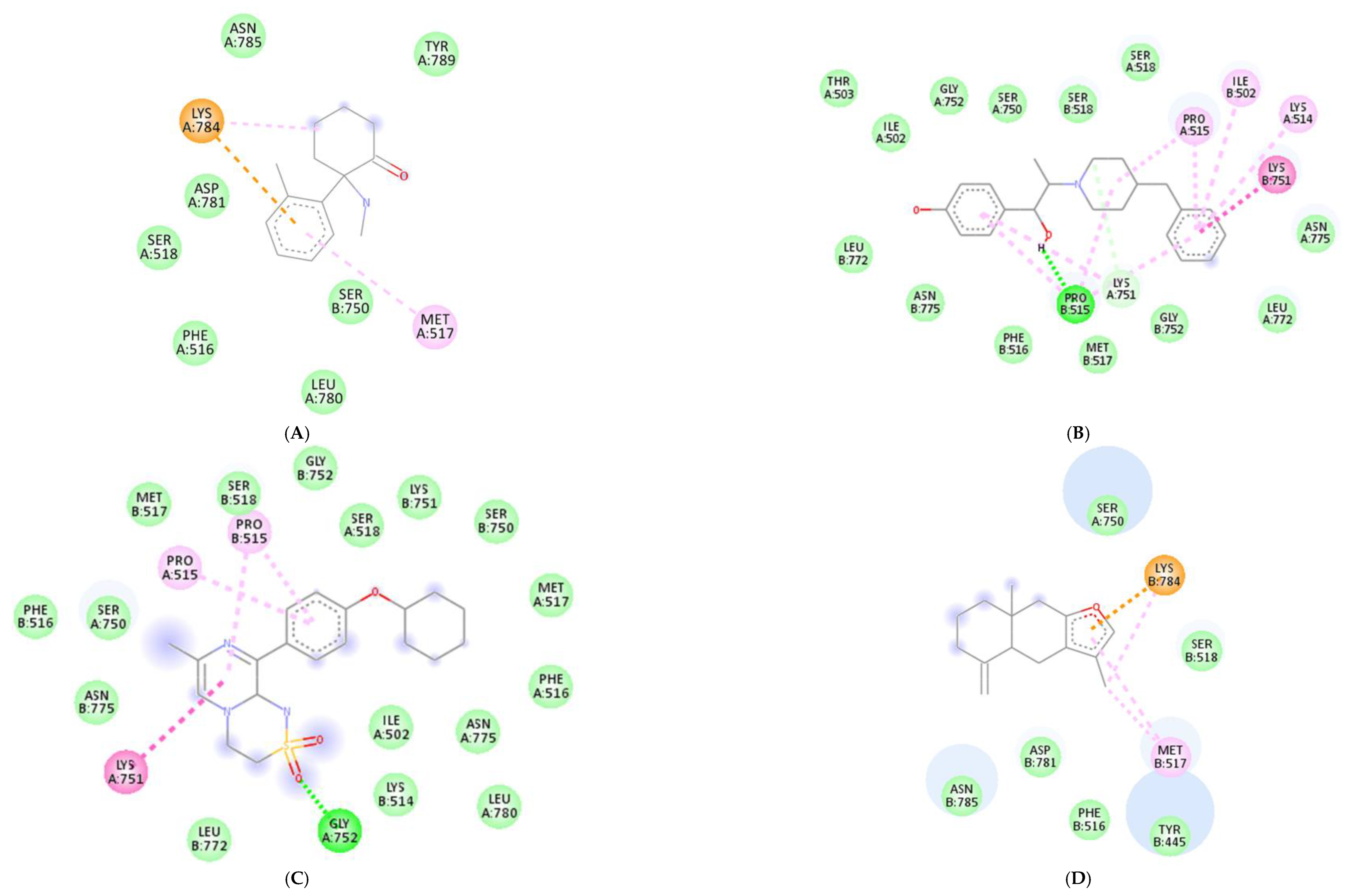


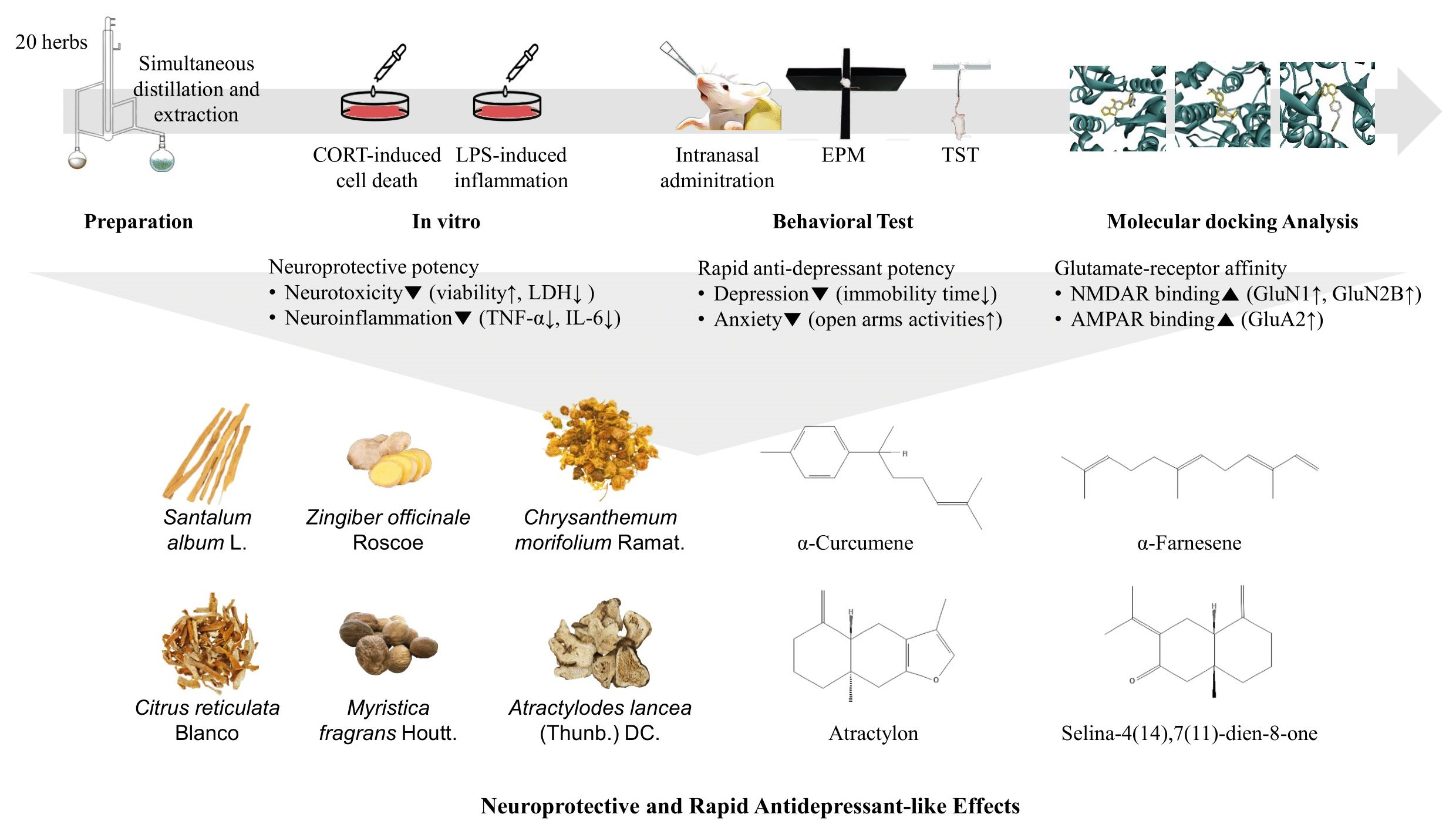{kind=link}
{kind=link}
{kind=link}
{kind=link}
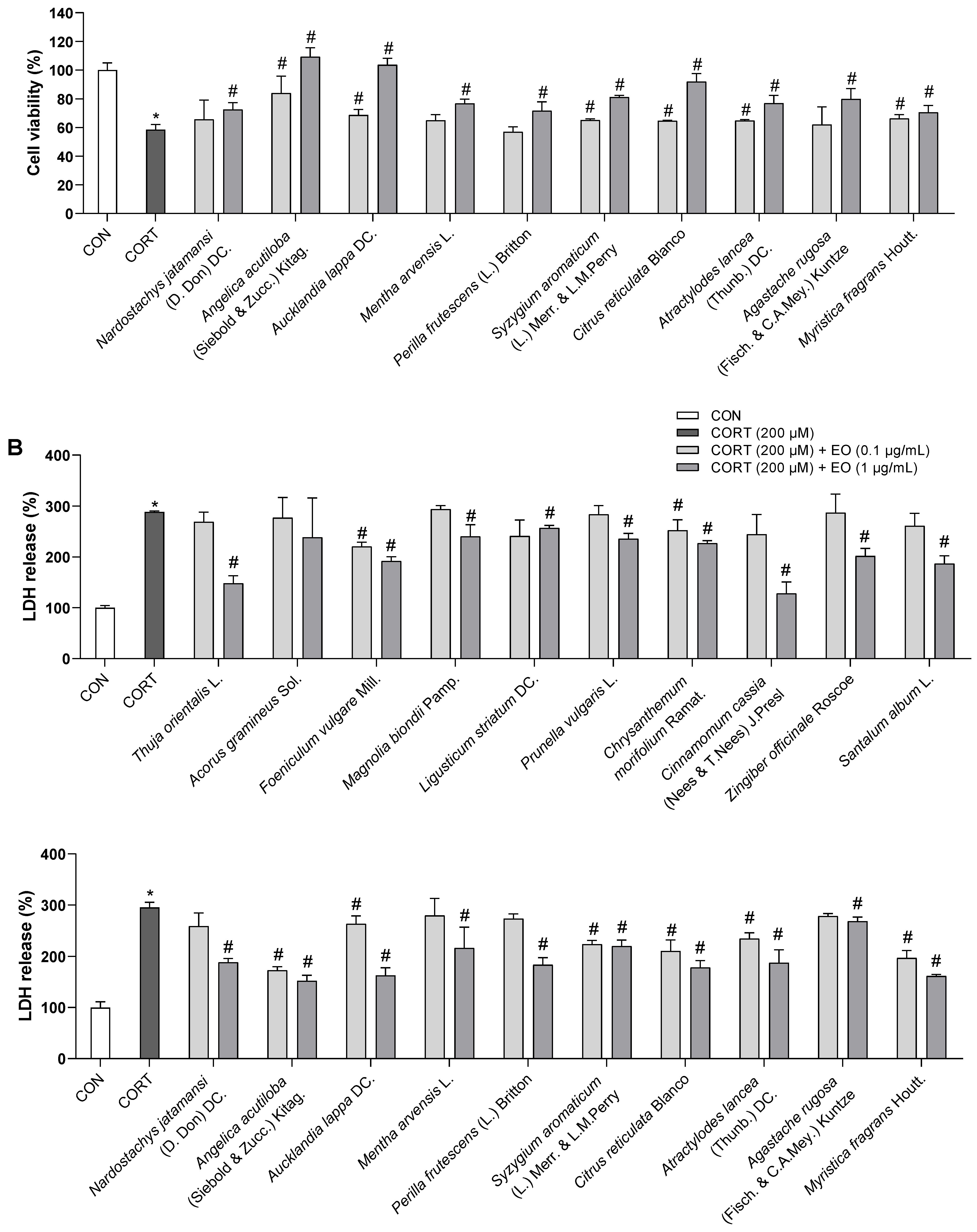{kind=link}
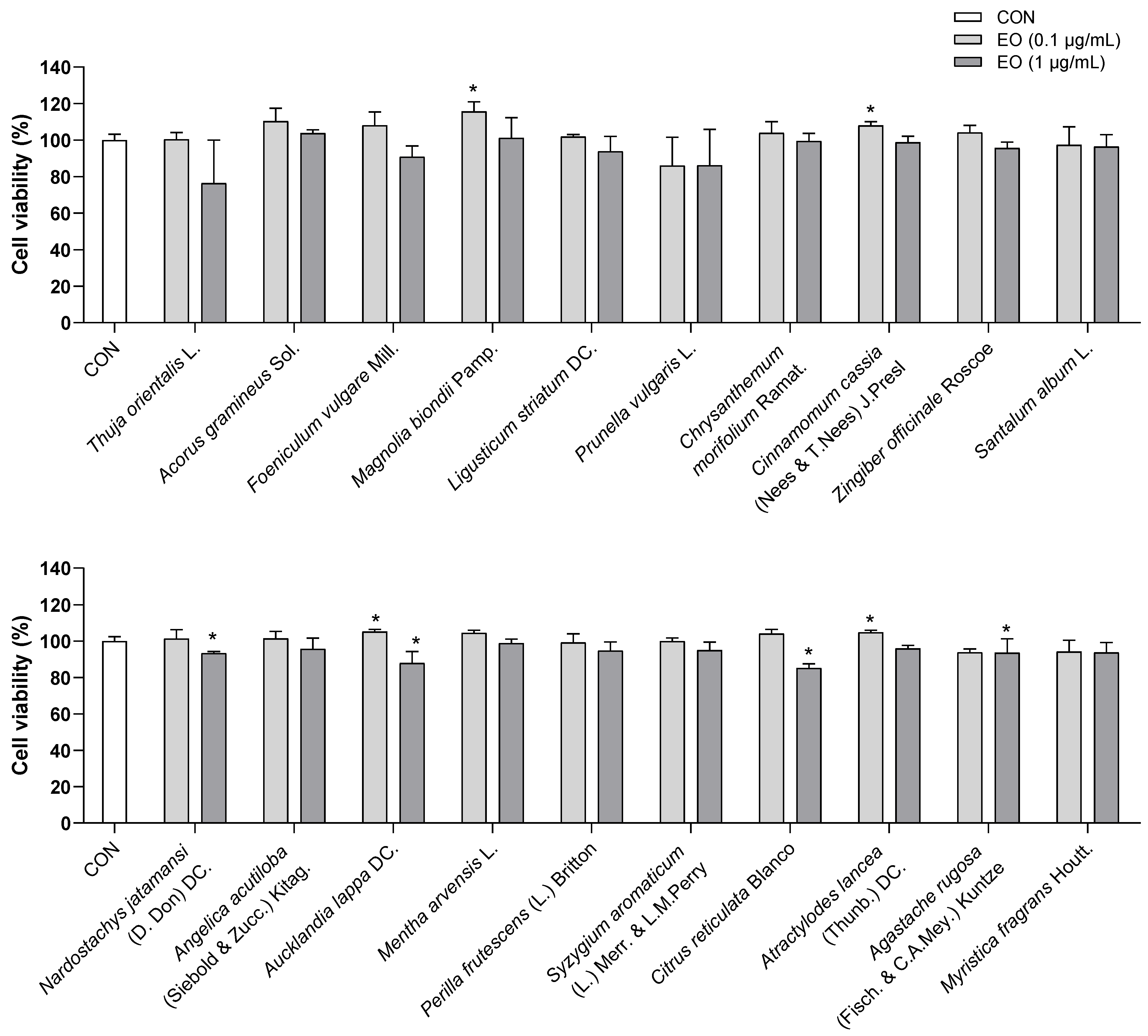{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
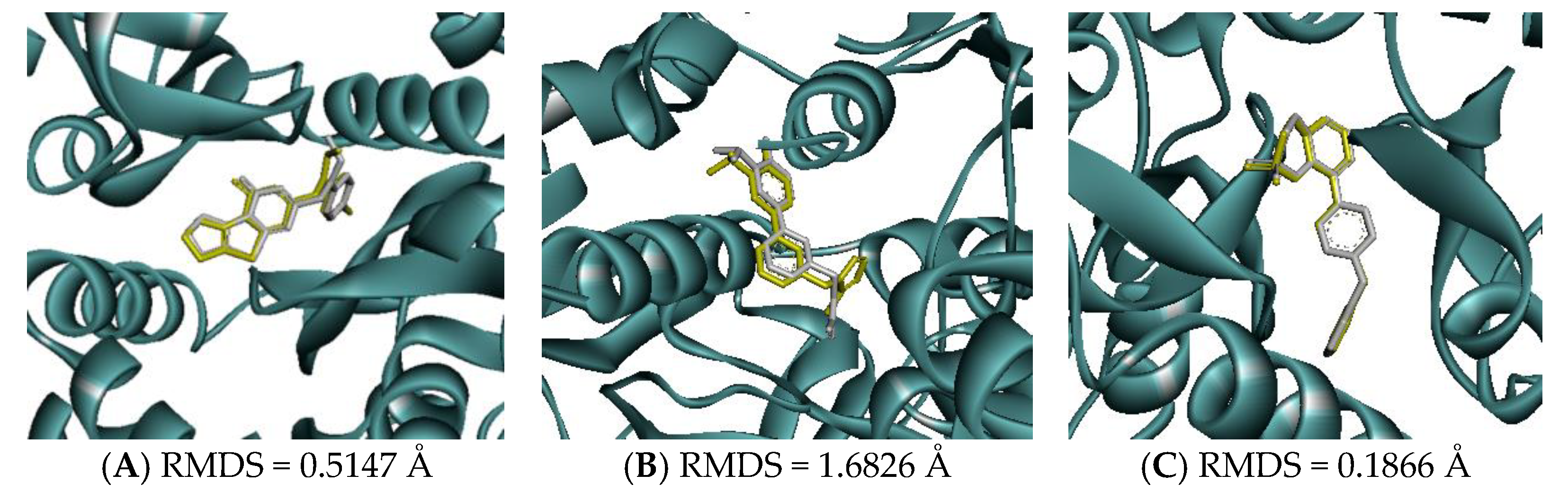{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Main compounds of essential oils.
| No. | Essential Oil | Main Compounds | Ref. | ||||
|---|---|---|---|---|---|---|---|
| 1 | Magnolia biondii Pamp. | Camphor | trans-Caryophyllene | 1,8-Cineole | α-Pinene | β-Pinene | [44,45,46] |
| 2 | Chrysanthemum morifolium Ramat. | α-Curcumene | α-Farnesene | n-Heptadecane | Linoleic Acid | Nonadecane | [47,48] |
| 3 | Zingiber officinale Roscoe | Camphene | 1,8-Cineole | β-Myrcene | β-Phellandrene | α-Pinene | [49,50,51] |
| 4 | Santalum album L. | (Z)-trans-α-Bergamotol | (Z)-Lanceol | (E)-Nuciferol | (Z)-α-Santalol | (E)-β-Santalol | [52,53,54] |
| 5 | Citrus reticulata Blanco | Limonene | Linalool | β-Myrcene | α-Pinene | γ-Terpinene | [55,56,57,58] |
| 6 | Atractylodes lancea (Thunb.) DC. | Atractylodin | Atractylon | β-Eudesmol | Elemol | Selina-4(14),7(11)-dien-8-one | [59,60,61] |
| 7 | Myristica fragrans Houtt. | Limonene | β-Myrcene | α-Phellandrene | α-Pinene | β-Pinene | [62,63,64] |
Table 2.
List of ligands.
| No. | Compound | Pubchem CID |
|---|---|---|
| Reference compounds | ||
| 1 | Ketamine | 3821 |
| 2 | Ifenprodil | 3689 |
| 3 | TAK-653 | 56655833 |
| Herbal compounds | ||
| 1 | Atractylodin | 5321047 |
| 2 | Atractylon | 3080635 |
| 3 | (Z)-trans-α-Bergamotol | 5368743 |
| 4 | Camphene | 6616 |
| 5 | Camphor | 2537 |
| 6 | trans-Caryophyllene | 5281515 |
| 7 | 1,8-Cineole | 2758 |
| 8 | α-Curcumene | 92139 |
| 9 | Elemol | 92138 |
| 10 | β-Eudesmol | 91457 |
| 11 | α-Farnesene | 5281516 |
| 12 | n-Heptadecane | 12398 |
| 13 | (Z)-Lanceol | 15560069 |
| 14 | Limonene | 22311 |
| 15 | Linalool | 6549 |
| 16 | Linoleic Acid | 5280450 |
| 17 | β-Myrcene | 31253 |
| 18 | Nonadecane | 12401 |
| 19 | (E)-Nuciferol | 6429177 |
| 20 | α-Phellandrene | 7460 |
| 21 | β-Phellandrene | 11142 |
| 22 | α-Pinene | 6654 |
| 23 | β-Pinene | 14896 |
| 24 | (Z)-α-Santalol | 11085337 |
| 25 | (E)-β-Santalol | 11031396 |
| 26 | Selina-4(14),7(11)-dien-8-one | 13986099 |
| 27 | γ-Terpinene | 7461 |
Table 3.
Docking results of ligands towards GluN1, GluN2B, and GluA2 proteins.
| No. | Ligand | Binding Energy (kcal/mol) | ||
|---|---|---|---|---|
| GluN1 | GluN2B | GluA2 | ||
| Native ligand | −10.4 | −10.7 | −9.9 | |
| 1 | Ketamine | −8.2 | −5.9 | −4.9 |
| 2 | Ifenprodil | −9.6 | −9.7 | −9.2 |
| 3 | TAK-653 | −8.6 | −8.6 | −9.2 |
| 4 | Atractylodin | −7.2 | −7.2 | −6.0 |
| 5 | Atractylon | −8.7 | −8.8 | −6.1 |
| 6 | (Z)-trans-α-Bergamotol | −7.9 | −7.0 | −5.8 |
| 7 | Camphene | −5.8 | −5.3 | −4.7 |
| 8 | Camphor | −5.9 | −4.6 | −4.3 |
| 9 | trans-Caryophyllene | −7.0 | −6.4 | −5.6 |
| 10 | 1,8-Cineole | −6.0 | −5.6 | −4.5 |
| 11 | α-Curcumene | −8.8 | −9.3 | −7.0 |
| 12 | Elemol | −7.2 | −7.8 | −6.4 |
| 13 | β-Eudesmol | −7.0 | −6.0 | −6.2 |
| 14 | α-Farnesene | −8.2 | −8.4 | −6.7 |
| 15 | n-Heptadecane | −6.3 | −6.6 | −5.8 |
| 16 | (Z)-Lanceol | −7.8 | −8.1 | −6.9 |
| 17 | Limonene | −6.6 | −7.5 | −6.0 |
| 18 | Linalool | −6.4 | −6.7 | −5.7 |
| 19 | Linoleic Acid | −7.6 | −7.2 | −6.6 |
| 20 | β-Myrcene | −6.4 | −6.9 | −5.0 |
| 21 | Nonadecane | −6.8 | −8.1 | −6.9 |
| 22 | (E)-Nuciferol | −8.1 | −8.4 | −6.9 |
| 23 | α-Phellandrene | −6.7 | −7.4 | −6.1 |
| 24 | β-Phellandrene | −6.4 | −7.5 | −6.1 |
| 25 | α-Pinene | −6.1 | −5.7 | −4.6 |
| 26 | β-Pinene | −6.1 | −6.2 | −4.4 |
| 27 | (Z)-α-Santalol | −7.4 | −6.4 | −5.7 |
| 28 | (E)-β-Santalol | −7.2 | −5.5 | −6.0 |
| 29 | Selina-4(14),7(11)-dien-8-one | −8.8 | −6.8 | −7.6 |
| 30 | γ-Terpinene | −7.0 | −7.4 | −5.9 |
The colors indicate binding energy values corresponding to binding affinity of ligands to the receptor subunits. Redder color: lower energy value, higher binding affinity; Whiter color: higher energy value, lower binding affinity.
Table 4.
Summary of key research findings: neuroprotective, anti-neuroinflammatory effects in vitro and rapid-acting anti-anxiety and anti-depressant effects in vivo of 20 essential oils.
Table 4.
Summary of key research findings: neuroprotective, anti-neuroinflammatory effects in vitro and rapid-acting anti-anxiety and anti-depressant effects in vivo of 20 essential oils.
| No. | Herb | In Vitro | In Vivo | |||||
|---|---|---|---|---|---|---|---|---|
| Neuroprotective Dose (µg/mL) | Anti-Neuroinflammatory Dose (µg/mL) | Rapid Anti-Depressant Dose (mg/kg) | Rapid Anti-Anxiety Dose (mg/kg) | |||||
| TNF-α | IL-6 | |||||||
| 1 | Thuja orientalis L. | 1 | - | - | - | - | ||
| 2 | Acorus gramineus Sol. | - | - | - | - | - | ||
| 3 | Foeniculum vulgare Mill. | 0.1, 1 | 1 | - | - | - | ||
| 4 | Magnolia biondii Pamp. | 1 | - | - | - | - | ||
| 5 | Ligusticum striatum DC. | 1 | 1 | 1 | - | - | ||
| 6 | Prunella vulgaris L. | 1 | 1 | - | - | - | ||
| 7 | Chrysanthemum morifolium Ramat. | 0.1, 1 | 0.1, 1 | - | 25 | 25 | ||
| 8 | Cinnamomum cassia (Nees & T.Nees) J.Presl | 1 | 0.1 | 0.1, 1 | - | - | ||
| 9 | Zingiber officinale Roscoe | 1 | - | - | 25 | - | ||
| 10 | Santalum album L. | 1 | 0.1, 1 | 1 | 25 | - | ||
| 11 | Nardostachys jatamansi (D. Don) DC. | 1 | - | - | - | - | ||
| 12 | Angelica acutiloba (Siebold & Zucc.) Kitag. | 0.1, 1 | - | - | - | - | ||
| 13 | Aucklandia lappa DC. | 0.1, 1 | - | 1 | - | - | ||
| 14 | Mentha arvensis L. | 1 | - | 1 | - | - | ||
| 15 | Perilla frutescens (L.) Britton | 1 | - | 0.1, 1 | - | - | ||
| 16 | Syzygium aromaticum (L.) Merr. & L.M.Perry | 0.1, 1 | - | 0.1, 1 | - | - | ||
| 17 | Citrus reticulata Blanco | 0.1, 1 | - | 0.1, 1 | 25 | - | ||
| 18 | Atractylodes lancea (Thunb.) DC. | 0.1, 1 | - | 0.1, 1 | 25 | - | ||
| 19 | Agastache rugosa (Fisch. & C.A.Mey.) Kuntze. | 1 | - | - | - | - | ||
| 20 | Myristica fragrans Houtt. | 0.1, 1 | - | 0.1, 1 | 25 | 25 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Tran, K.N.; Nguyen, N.P.K.; Nguyen, L.T.H.; Shin, H.-M.; Yang, I.-J. Screening for Neuroprotective and Rapid Antidepressant-like Effects of 20 Essential Oils. Biomedicines 2023, 11, 1248. https://doi.org/10.3390/biomedicines11051248
AMA Style
Tran KN, Nguyen NPK, Nguyen LTH, Shin H-M, Yang I-J. Screening for Neuroprotective and Rapid Antidepressant-like Effects of 20 Essential Oils. Biomedicines. 2023; 11(5):1248. https://doi.org/10.3390/biomedicines11051248
Chicago/Turabian StyleTran, Khoa Nguyen, Nhi Phuc Khanh Nguyen, Ly Thi Huong Nguyen, Heung-Mook Shin, and In-Jun Yang. 2023. "Screening for Neuroprotective and Rapid Antidepressant-like Effects of 20 Essential Oils" Biomedicines 11, no. 5: 1248. https://doi.org/10.3390/biomedicines11051248
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.